
Evaluating the Enzyme Activities and Soil Physicochemical Properties of Four Typical Halophytic Communities in Saline-Sodic Soil
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Overview of the Study Area
2.2. Sample Collection
2.3. Analysis Methods
2.4. Statistical Analysis
3. Results
3.1. Physicochemical Characteristics of Representative Salt-Tolerant Plant Soils in Saline-Sodic Soil
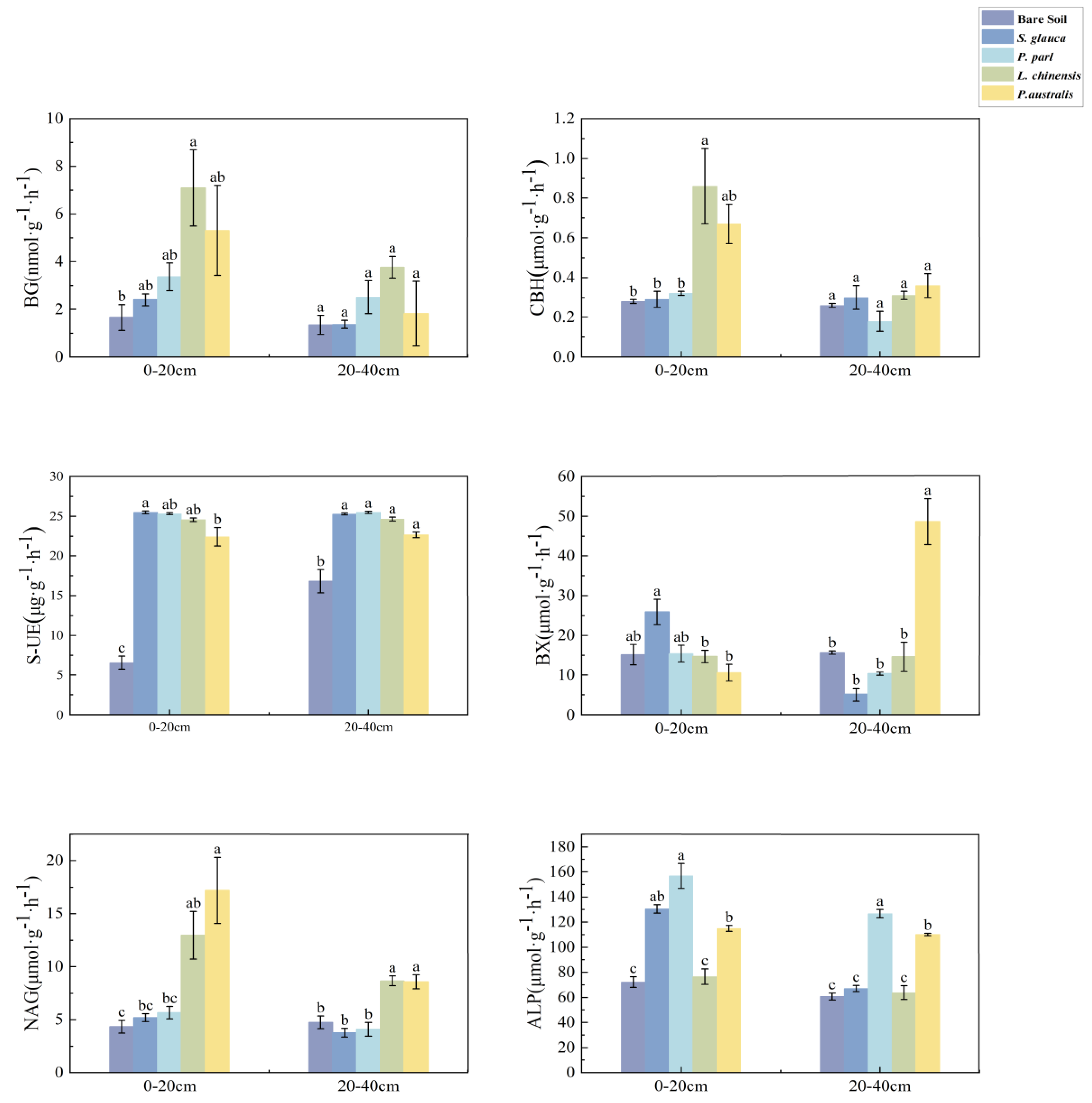
3.2. Investigation of the Varying Enzymatic Activity in Soil across Distinct Halophytic Plant Species
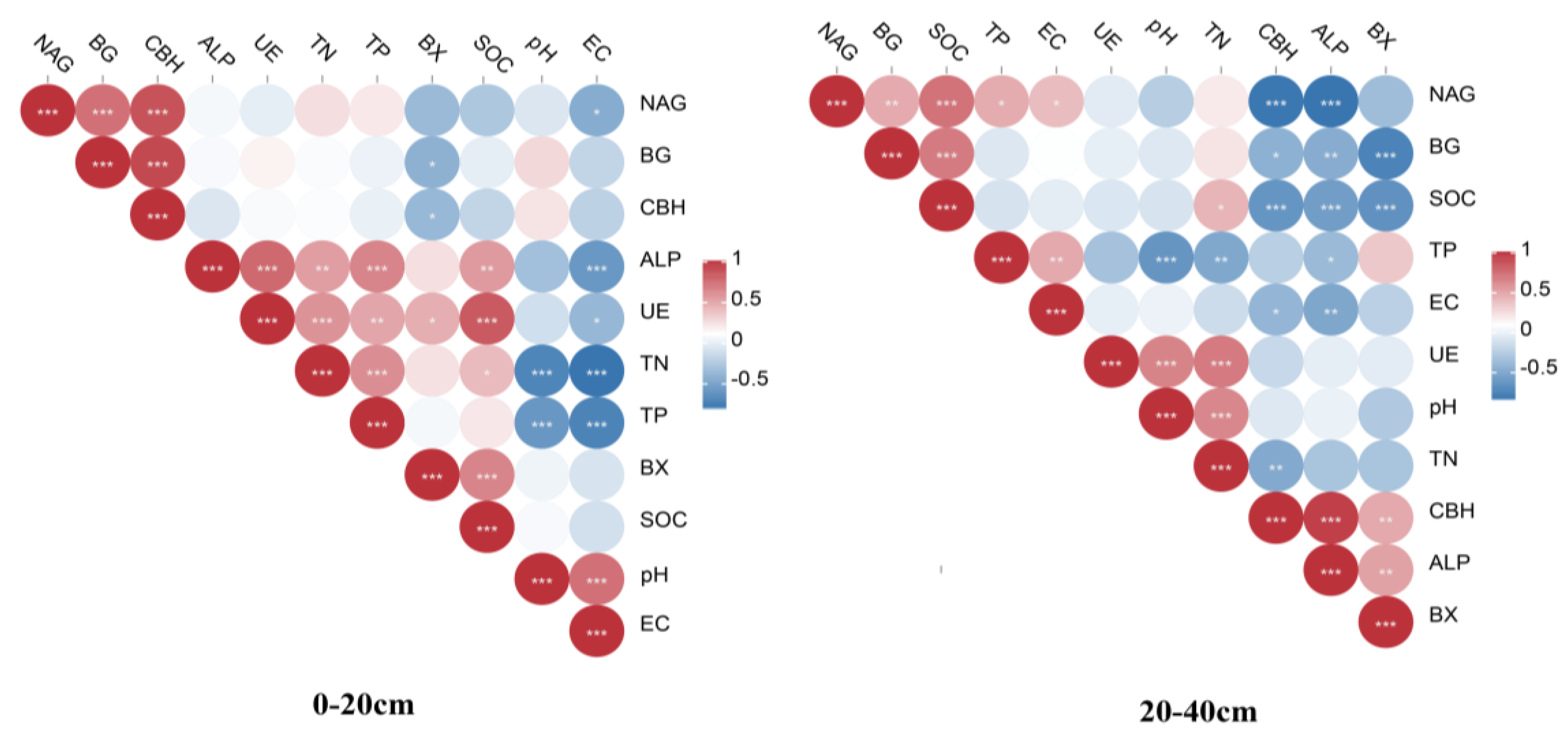
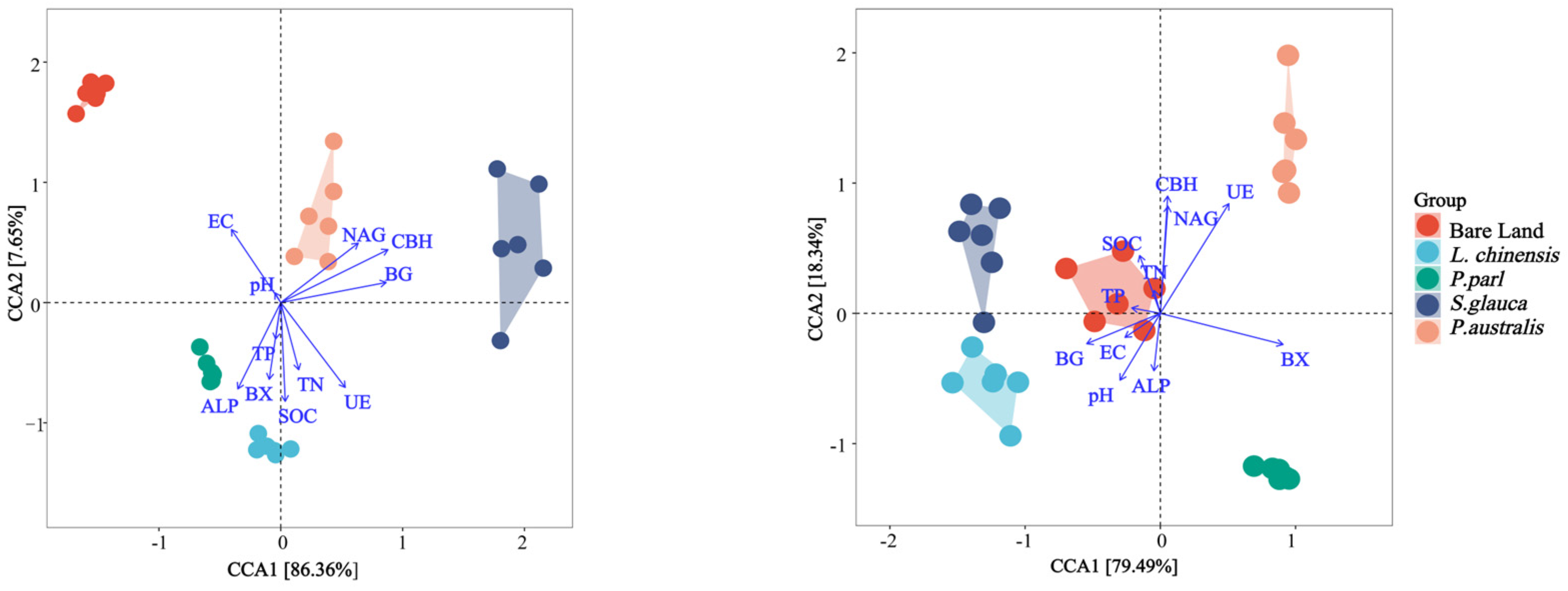
3.3. Investigating the Relationship between Soil Properties and Soil Enzyme Activity throughout Various Soil Layers
4. Discussion
4.1. Effect of Vegetation on Soil Enzyme Activity
4.2. Effects of Environmental Factors on Enzyme Activities
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wang, J.; Feng, W.; Zhang, H.; Sun, J.; Zhao, Q. Experimental study on the effect of deep pine technology on water and salt transport in soda saline land. Arab. J. Geosci. 2021, 14, 571. [Google Scholar] [CrossRef]
- Li, Y.; Li, Q.; Guo, D.; Liang, S.; Wang, Y. Ecological stoichiometry homeostasis of Leymus chinensis in degraded grassland in western Jilin Province, NE China. Ecol. Eng. 2016, 90, 387–391. [Google Scholar] [CrossRef]
- Li, Y.; Liang, S.; Zhao, Y.; Li, W.; Wang, Y. Machine learning for the prediction of L. chinensis carbon, nitrogen and phosphorus contents and understanding of mechanisms underlying grassland degradation. J. Environ. Manag. 2017, 192, 116–123. [Google Scholar] [CrossRef]
- Amini, S.; Ghadiri, H.; Chen, C.; Marschner, P. Salt-affected soils, reclamation, carbon dynamics, and biochar: A review. J. Soils Sediments 2016, 16, 939–953. [Google Scholar] [CrossRef]
- Luo, J.-Q.; Wang, L.-L.; Li, Q.-S.; Zhang, Q.-K.; He, B.-Y.; Wang, Y.; Qin, L.-P.; Li, S.-S. Improvement of hard saline–sodic soils using polymeric aluminum ferric sulfate (PAFS). Soil Tillage Res. 2015, 149, 12–20. [Google Scholar] [CrossRef]
- Li, F.; Li, H.-R.; Zhou, J.; Liu, Q.; Guo, R.; Xia, X. Eco-physiological responses of linseed (Linum usitatissimum) to salt and alkali stresses. Chin. J. Plant Ecol. 2016, 40, 69–79. [Google Scholar]
- Qadir, M.; Qureshi, R.H.; Ahmad, N. Reclamation of a saline-sodic soil by gypsum and Leptochloa fusca. Geoderma 1996, 74, 207–217. [Google Scholar] [CrossRef]
- Robbins, C.W. Sodic Calcareous Soil Reclamation as Affected by Different Amendments and Crops. Agron. J. 1986, 78, 916–920. [Google Scholar] [CrossRef]
- Li, N.; Shao, T.; Zhou, Y.; Cao, Y.; Hu, H.; Sun, Q.; Long, X.; Yue, Y.; Gao, X.; Rengel, Z. Effects of planting Melia azedarach L. on soil properties and microbial community in saline-alkali soil. Land Degrad. Dev. 2021, 32, 2951–2961. [Google Scholar] [CrossRef]
- Cao, D.; Shi, F.; Koike, T.; Lu, Z.; Sun, J. Halophyte Plant Communities Affecting Enzyme Activity and Microbes in Saline Soils of the Yellow River Delta in China. CLEAN—Soil Air Water 2014, 42, 1433–1440. [Google Scholar] [CrossRef]
- Zhao, Q.; Tang, J.; Li, Z.; Yang, W.; Duan, Y. The Influence of Soil Physico-Chemical Properties and Enzyme Activities on Soil Quality of Saline-Alkali Agroecosystems in Western Jilin Province, China. Sustainability 2018, 10, 1529. [Google Scholar] [CrossRef]
- Ma, X.; Zarebanadkouki, M.; Kuzyakov, Y.; Blagodatskaya, E.; Pausch, J.; Razavi, B.S. Spatial patterns of enzyme activities in the rhizosphere: Effects of root hairs and root radius. Soil Biol. Biochem. 2018, 118, 69–78. [Google Scholar] [CrossRef]
- Burns, R.G.; Dick, R.P. Enzymes in the Environment: Activity, Ecology, and Applications; CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar]
- Das, S.K.; Varma, A. Role of Enzymes in Maintaining Soil Health. In Soil Enzymology; Shukla, G., Varma, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 25–42. [Google Scholar]
- Prasad, S.; Malav, L.C.; Choudhary, J.; Kannojiya, S.; Kundu, M.; Kumar, S.; Yadav, A.N. Soil Microbiomes for Healthy Nutrient Recycling. Current Trends in Microbial Biotechnology for Sustainable Agriculture; Springer: Berlin/Heidelberg, Germany, 2021; pp. 1–21. [Google Scholar]
- Kotroczó, Z.; Veres, Z.; Fekete, I.; Krakomperger, Z.; Tóth, J.A.; Lajtha, K.; Tóthmérész, B. Soil enzyme activity in response to long-term organic matter manipulation. Soil Biol. Biochem. 2014, 70, 237–243. [Google Scholar] [CrossRef]
- Ge, G.; Li, Z.; Fan, F.; Chu, G.; Hou, Z.; Liang, Y. Soil biological activity and their seasonal variations in response to long-term application of organic and inorganic fertilizers. Plant Soil 2010, 326, 31–44. [Google Scholar] [CrossRef]
- Dick, W.A.; Cheng, L.; Wang, P. Soil acid and alkaline phosphatase activity as pH adjustment indicators. Soil Biol. Biochem. 2000, 32, 1915–1919. [Google Scholar] [CrossRef]
- Xu, H.; Qu, Q.; Chen, Y.; Liu, G.; Xue, S. Responses of soil enzyme activity and soil organic carbon stability over time after cropland abandonment in different vegetation zones of the Loess Plateau of China. CATENA 2021, 196, 104812. [Google Scholar] [CrossRef]
- Gong, X.; Liu, C.; Li, J.; Luo, Y.; Yang, Q.; Zhang, W.; Yang, P.; Feng, B. Responses of rhizosphere soil properties, enzyme activities and microbial diversity to intercropping patterns on the Loess Plateau of China. Soil Tillage Res. 2019, 195, 104355. [Google Scholar] [CrossRef]
- Veres, Z.; Kotroczó, Z.; Fekete, I.; Tóth, J.A.; Lajtha, K.; Townsend, K.; Tóthmérész, B. Soil extracellular enzyme activities are sensitive indicators of detrital inputs and carbon availability. Appl. Soil Ecol. 2015, 92, 18–23. [Google Scholar] [CrossRef]
- Li, J. Flora of China. Harv. Pap. Bot. 2007, 13, 301–302. [Google Scholar] [CrossRef]
- Ding, Y.; Liu, Y.; Liu, S.; Li, Z.; Tan, X.; Huang, X.; Zeng, G.; Zhou, L.; Zheng, B. Biochar to improve soil fertility. A review. Agron. Sustain. Dev. 2016, 36, 36. [Google Scholar] [CrossRef]
- Rutigliano, F.A.; Castaldi, S.; D’Ascoli, R.; Papa, S.; Carfora, A.; Marzaioli, R.; Fioretto, A. Soil activities related to nitrogen cycle under three plant cover types in Mediterranean environment. Appl. Soil Ecol. 2009, 43, 40–46. [Google Scholar] [CrossRef]
- Zhou, L.; Zhao, B.; Zhuang, W.-Q. Double-edged sword effects of dissimilatory nitrate reduction to ammonium (DNRA) bacteria on anammox bacteria performance in an MBR reactor. Water Res. 2023, 233, 119754. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Su, C.; Fan, C.; Cao, L.; Liang, Z.; Xu, Y.; Chen, Z.; Wang, Q.; Chen, M. Metagenomic and extracellular polymeric substances analysis reveals the mechanism of exogenous N-hexanoyl-L-homoserine lactone in alleviating the inhibition of perfluorooctanoic acid on anammox process. Bioresour. Technol. 2023, 369, 128482. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ma, J.; Xiao, C.; Li, Y. Effects of climate factors and soil properties on soil nutrients and elemental stoichiometry across the Huang–Huai–Hai River Basin, China. J. Soils Sediments 2020, 20, 1970–1982. [Google Scholar] [CrossRef]
- Dai, W.; Li, Y.; Fu, W.; Jiang, P.; Zhao, K.; Li, Y.; Penttinen, P. Spatial variability of soil nutrients in forest areas: A case study from subtropical China. J. Plant Nutr. Soil Sci. 2018, 181, 827–835. [Google Scholar] [CrossRef]
- Bogunovic, I.; Pereira, P.; Brevik, E.C. Spatial distribution of soil chemical properties in an organic farm in Croatia. Sci. Total Environ. 2017, 584–585, 535–545. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Liu, G.B.; Xue, S.; Zhu, B. Changes in soil physico-chemical and microbiological properties during natural succession on abandoned farmland in the Loess Plateau. Environ. Earth Sci. 2011, 62, 915–925. [Google Scholar] [CrossRef]
- Qu, Y.; Tang, J.; Li, Z.; Zhou, Z.; Wang, J.; Wang, S.; Cao, Y. Soil Enzyme Activity and Microbial Metabolic Function Diversity in Soda Saline–Alkali Rice Paddy Fields of Northeast China. Sustainability 2020, 12, 10095. [Google Scholar] [CrossRef]
- Yin, R.; Deng, H.; Wang, H.-l.; Zhang, B. Vegetation type affects soil enzyme activities and microbial functional diversity following re-vegetation of a severely eroded red soil in sub-tropical China. CATENA 2014, 115, 96–103. [Google Scholar] [CrossRef]
- Hartman, W.H.; Richardson, C.J.; Vilgalys, R.; Bruland, G.L. Environmental and anthropogenic controls over bacterial communities in wetland soils. Proc. Natl. Acad. Sci. USA 2008, 105, 17842–17847. [Google Scholar] [CrossRef]
- Cui, J.; Song, D.; Dai, X.; Xu, X.; He, P.; Wang, X.; Liang, G.; Zhou, W.; Zhu, P. Effects of long-term cropping regimes on SOC stability, soil microbial community and enzyme activities in the Mollisol region of Northeast China. Appl. Soil Ecol. 2021, 164, 103941. [Google Scholar] [CrossRef]
- Wu, G.; Chen, Z.; Jiang, N.; Jiang, H.; Chen, L. Effects of long-term no-tillage with different residue application rates on soil nitrogen cycling. Soil Tillage Res. 2021, 212, 105044. [Google Scholar] [CrossRef]
- Broeckling, C.D.; Broz, A.K.; Bergelson, J.; Manter, D.K.; Vivanco, J.M. Root Exudates Regulate Soil Fungal Community Composition and Diversity. Appl. Environ. Microbiol. 2008, 74, 738–744. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Richardson, A.E.; O’Callaghan, M.; DeAngelis, K.M.; Jones, E.E.; Stewart, A.; Firestone, M.K.; Condron, L.M. Effects of selected root exudate components on soil bacterial communities. FEMS Microbiol. Ecol. 2011, 77, 600–610. [Google Scholar] [CrossRef] [PubMed]
- Rath, K.M.; Fierer, N.; Murphy, D.V.; Rousk, J. Linking bacterial community composition to soil salinity along environmental gradients. ISME J. 2019, 13, 836–846. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Mei, L.; Wei, Q.; Li, B.; Zhang, P.; Sun, S.; Cui, G. Leymus chinensis resists degraded soil stress by modulating root exudate components to attract beneficial microorganisms. Front. Microbiol. 2022, 13, 951838. [Google Scholar] [CrossRef]
- Liu, T.; Wang, S.; Chen, Y.; Luo, J.; Hao, B.; Zhang, Z.; Yang, B.; Guo, W. Bio-organic fertilizer promoted phytoremediation using native plant leymus chinensis in heavy Metal(loid)s contaminated saline soil. Environ. Pollut. 2023, 327, 121599. [Google Scholar] [CrossRef] [PubMed]
- Gianfreda, L.; Ruggiero, P. Enzyme Activities in Soil. In Nucleic Acids and Proteins in Soil; Springer: Berlin/Heidelberg, Germany, 2006; pp. 257–311. [Google Scholar]
- Nannipieri, P.; Giagnoni, L.; Renella, G.; Puglisi, E.; Ceccanti, B.; Masciandaro, G.; Fornasier, F.; Moscatelli, M.C.; Marinari, S. Soil enzymology: Classical and molecular approaches. Biol. Fertil. Soils 2012, 48, 743–762. [Google Scholar] [CrossRef]
- Jiao, X.-G.; Gao, C.-S.; Lü, G.-H.; Sui, Y.-Y. Effect of Long-Term Fertilization on Soil Enzyme Activities Under Different Hydrothermal Conditions in Northeast China. Agric. Sci. China 2011, 10, 412–422. [Google Scholar] [CrossRef]
- Jourgholami, M.; Ramineh, A.; Zahedi Amiri, G.; Labelle, E.R. The Influence of Slope Positions on the Recovery Response of Compacted Soil Properties and Enzyme Activity in an Oriental Beech Stand in the Hyrcanian Forests, Iran. Sustainability 2019, 11, 1940. [Google Scholar] [CrossRef]
- Flores Fernández, J.L.; Rubin, L.; Hartmann, P.; Puhlmann, H.; von Wilpert, K. Initial recovery of soil structure of a compacted forest soil can be enhanced by technical treatments and planting. For. Ecol. Manag. 2019, 431, 54–62. [Google Scholar] [CrossRef]
- Naether, A.; Foesel, B.U.; Naegele, V.; Wüst, P.K.; Weinert, J.; Bonkowski, M.; Alt, F.; Oelmann, Y.; Polle, A.; Lohaus, G.; et al. Environmental Factors Affect Acidobacterial Communities below the Subgroup Level in Grassland and Forest Soils. Appl. Environ. Microbiol. 2012, 78, 7398–7406. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.; Zhang, B.; Yin, R.; Wang, H.-l.; Mitchell, S.M.; Griffiths, B.S.; Daniell, T.J. Long-term effect of re-vegetation on the microbial community of a severely eroded soil in sub-tropical China. Plant Soil 2010, 328, 447–458. [Google Scholar] [CrossRef]
- Garcia, C.; Roldan, A.; Hernandez, T. Ability of different plant species to promote microbiological processes in semiarid soil. Geoderma 2005, 124, 193–202. [Google Scholar] [CrossRef]
- Stone, M.M.; DeForest, J.L.; Plante, A.F. Changes in extracellular enzyme activity and microbial community structure with soil depth at the Luquillo Critical Zone Observatory. Soil Biol. Biochem. 2014, 75, 237–247. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, X.; Kuzyakov, Y.; Wang, H.; Fu, X.; Yang, Y.; Chen, F.; Dungait, J.A.J.; Green, S.M.; Fang, X. Responses of C-, N- and P-acquiring hydrolases to P and N fertilizers in a subtropical Chinese fir plantation depend on soil depth. Appl. Soil Ecol. 2020, 150, 103465. [Google Scholar] [CrossRef]
- Steinauer, K.; Chatzinotas, A.; Eisenhauer, N. Root exudate cocktails: The link between plant diversity and soil microorganisms? Ecol. Evol. 2016, 6, 7387–7396. [Google Scholar] [CrossRef] [PubMed]
- Neemisha; Sharma, S. Soil Enzymes and Their Role in Nutrient Cycling. In Structure and Functions of Pedosphere; Springer: Berlin/Heidelberg, Germany, 2022; pp. 173–188. [Google Scholar]
- Zhao, H.; Jiang, Y.; Ning, P.; Liu, J.; Zheng, W.; Tian, X.; Shi, J.; Xu, M.; Liang, Z.; Shar, A.G. Effect of Different Straw Return Modes on Soil Bacterial Community, Enzyme Activities and Organic Carbon Fractions. Soil Sci. Soc. Am. J. 2019, 83, 638–648. [Google Scholar] [CrossRef]
- Yan, B.; Wang, X.; Sun, Y.; Fan, B.; Shi, L.; Liu, G. Vegetation rehabilitation increases soil enzyme activities in degraded land via carbon supply and nitrogen retention. Eur. J. Soil Biol. 2020, 98, 103186. [Google Scholar] [CrossRef]
- Dong, W.Y.; Zhang, X.Y.; Liu, X.Y.; Fu, X.L.; Chen, F.S.; Wang, H.M.; Sun, X.M.; Wen, X.F. Responses of soil microbial communities and enzyme activities to nitrogen and phosphorus additions in Chinese fir plantations of subtropical China. Biogeosciences 2015, 12, 5537–5546. [Google Scholar] [CrossRef]
- Wei, K.; Sun, T.; Tian, J.; Chen, Z.; Chen, L. Soil microbial biomass, phosphatase and their relationships with phosphorus turnover under mixed inorganic and organic nitrogen addition in a Larix gmelinii plantation. For. Ecol. Manag. 2018, 422, 313–322. [Google Scholar] [CrossRef]
- Miransari, M. Soil microbes and plant fertilization. Appl. Microbiol. Biotechnol. 2011, 92, 875–885. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, D.; Bhardwaj, S.; Landi, M.; Sharma, A.; Ramakrishnan, M.; Sharma, A. The Impact of Drought in Plant Metabolism: How to Exploit Tolerance Mechanisms to Increase Crop Production. Appl. Sci. 2020, 10, 5692. [Google Scholar] [CrossRef]
- Canarini, A.; Kaiser, C.; Merchant, A.; Richter, A.; Wanek, W. Root Exudation of Primary Metabolites: Mechanisms and Their Roles in Plant Responses to Environmental Stimuli. Front. Plant Sci. 2019, 10, 157. [Google Scholar] [CrossRef]
- Park, J.-H.; Kalbitz, K.; Matzner, E. Resource control on the production of dissolved organic carbon and nitrogen in a deciduous forest floor. Soil Biol. Biochem. 2002, 34, 813–822. [Google Scholar] [CrossRef]
- Pugnaire, F.I.; Morillo, J.A.; Peñuelas, J.; Reich, P.B.; Bardgett, R.D.; Gaxiola, A.; Wardle, D.A.; van der Putten, W.H. Climate change effects on plant-soil feedbacks and consequences for biodiversity and functioning of terrestrial ecosystems. Sci. Adv. 2019, 5, eaaz1834. [Google Scholar] [CrossRef]
- Ding, X.; Chen, S.; Zhang, B.; Liang, C.; He, H.; Horwath, W.R. Warming increases microbial residue contribution to soil organic carbon in an alpine meadow. Soil Biol. Biochem. 2019, 135, 13–19. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Vegetation | Soil Depth (cm) | pH | EC (mS/cm) | SOC (%) | TN (g/kg) | TP (g/kg) | AP (mg/kg) |
|---|---|---|---|---|---|---|---|
| Bare soil | 0–20 | 10.39 ± 0.01 a | 5.95 ± 0.26 a | 0.54 ± 0.06 bc | 0.33 ± 0.04 d | 0.38 ± 0.03 a | 36.97 ± 2.38 a |
| 20–40 | 10.54 ± 0.01 a | 3.20 ± 0.19 a | 0.52 ± 0.08 bc | 0.32 ± 0.02 c | 0.63 ± 0.10 ab | 22.79 ± 0.81 a | |
| Puccinellia parl | 0–20 | 10.38 ± 0.01 a | 1.48 ± 0.03 bc | 0.78 ± 0.02 ab | 0.59 ± 0.03 c | 0.74 ± 0.11 a | 19.15 ± 0.59 b |
| 20–40 | 10.43 ± 0.02 a | 1.42 ± 0.06 b | 0.37 ± 0.06 c | 0.50 ± 0.04 c | 0.45 ± 0.01 b | 19.65 ± 0.58 a | |
| Suaeda glauca | 0–20 | 10.48 ± 0.03 a | 2.14 ± 0.28 b | 0.74 ± 0.01 abc | 0.53 ± 0.03 c | 0.42 ± 0.01 a | 18.16 ± 2.24 b |
| 20–40 | 10.44 ± 0.01 a | 1.90 ± 0.14 b | 0.48 ± 0.09 c | 0.43 ± 0.03 c | 0.41 ± 0.01 b | 22.89 ± 1.35 a | |
| Leymus chinensis | 0–20 | 9.41 ± 0.17 c | 0.28 ± 0.06 d | 0.94 ± 0.08 a | 1.17 ± 0.08 a | 0.97 ± 0.32 a | 3.73 ± 0.55 d |
| 20–40 | 9.50 ± 0.18 c | 0.35 ± 0.09 c | 0.85 ± 0.04 a | 0.98 ± 0.04 a | 1.28 ± 0.34 a | 4.64 ± 1.15 b | |
| Phragmites australis | 0–20 | 9.85 ± 0.08 b | 0.69 ± 0.22 cd | 0.50 ± 0.05 c | 0.94 ± 0.04 b | 0.89 ± 0.15 a | 10.43 ± 1.04 c |
| 20–40 | 10.02 ± 0.08 b | 0.62 ± 0.13 c | 0.77 ± 0.04 ab | 0.78 ± 0.08 b | 0.79 ± 0.07 ab | 3.50 ± 0.23 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, J.; Miao, Y.; Guo, L.; Zhang, T.; Nie, Z.; Luo, X.; Yang, F.; Wang, Z. Evaluating the Enzyme Activities and Soil Physicochemical Properties of Four Typical Halophytic Communities in Saline-Sodic Soil. Agronomy 2024, 14, 141. https://doi.org/10.3390/agronomy14010141
Zhou J, Miao Y, Guo L, Zhang T, Nie Z, Luo X, Yang F, Wang Z. Evaluating the Enzyme Activities and Soil Physicochemical Properties of Four Typical Halophytic Communities in Saline-Sodic Soil. Agronomy. 2024; 14(1):141. https://doi.org/10.3390/agronomy14010141
Chicago/Turabian StyleZhou, Jie, Yue Miao, Liangliang Guo, Tieyi Zhang, Zhaoyang Nie, Xuejiao Luo, Fan Yang, and Zhichun Wang. 2024. "Evaluating the Enzyme Activities and Soil Physicochemical Properties of Four Typical Halophytic Communities in Saline-Sodic Soil" Agronomy 14, no. 1: 141. https://doi.org/10.3390/agronomy14010141