
Individual and Combined Effects of Predatory Bug Engytatus nicotianae and Trichoderma atroviride in Suppressing the Tomato Potato Psyllid Bactericera cockerelli in Greenhouse Grown Tomatoes

 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Inoculation with T. atroviridae and Plant Growth Conditions
2.2. Experimental Design
2.3. Infestation of Plants with TPP and Introduction of E. nicotianae
2.4. Weekly Data Collection
2.5. VOC Sampling
2.6. Statistical Analyses
3. Results
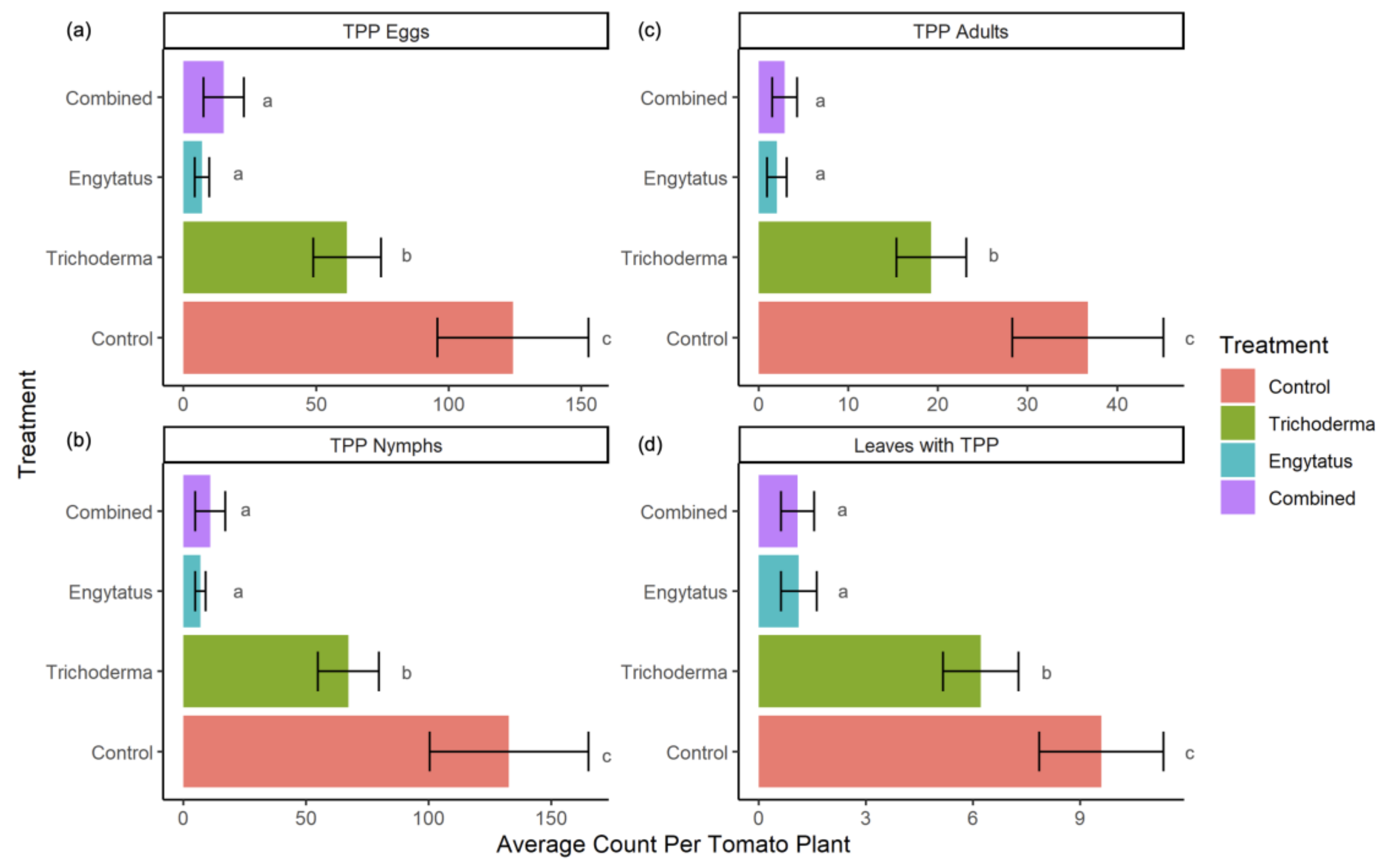
3.1. Average Rates of TPP Suppression
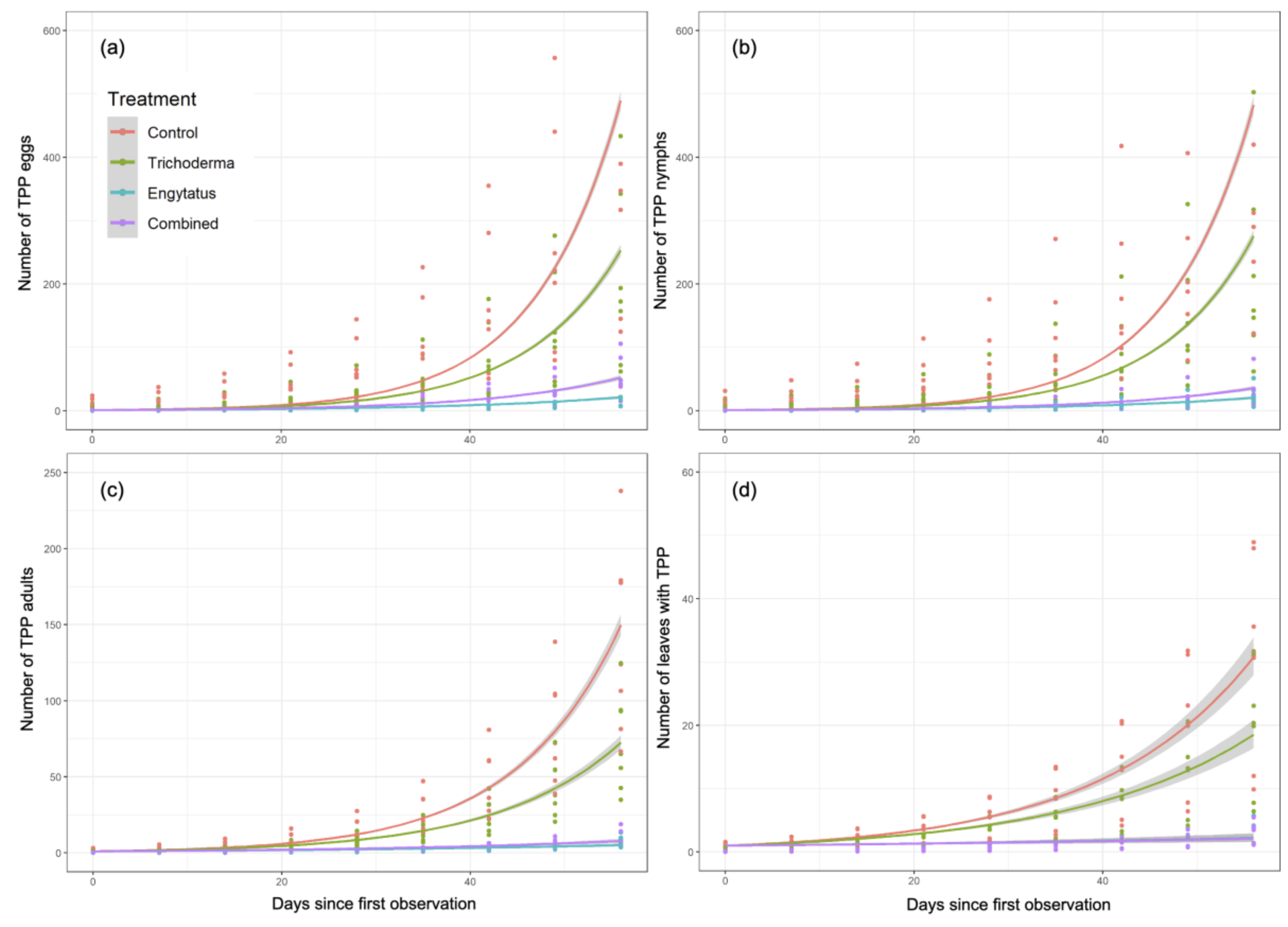
3.2. Comparison of TPP Growth Rates
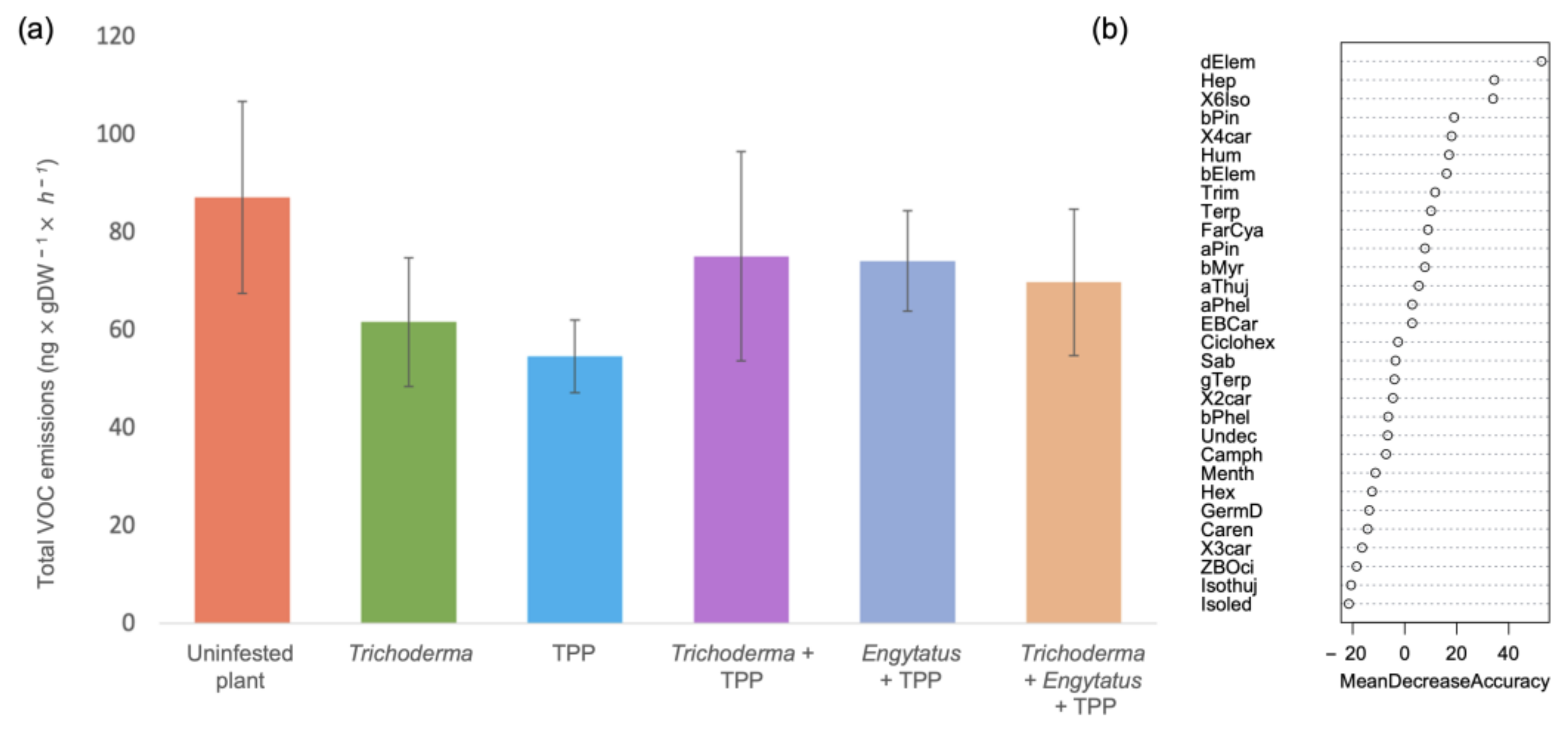
3.3. Treatment Effect of on Plant VOC Emissions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, D.; Trumble, J.T. Comparative fitness of invasive and native populations of the potato psyllid (Bactericera cockerelli). Entomol. Exp. Appl. 2007, 123, 35–42. [Google Scholar] [CrossRef]
- Liu, D.; Trumble, J.T. Interactions of plant resistance and insecticides on the development and survival of Bactericerca cockerelli [Sulc](Homoptera: Psyllidae). Crop Prot. 2005, 24, 111–117. [Google Scholar] [CrossRef]
- Butler, C.D.; Trumble, J.T. The potato psyllid, Bactericera cockerelli (Sulc)(Hemiptera: Triozidae): Life history, relationship to plant diseases, and management strategies. Terr. Arthropod Rev. 2012, 5, 87–111. [Google Scholar] [CrossRef]
- Teulon, D.; Workman, P.; Thomas, K.; Nielsen, M. Bactericera cockerelli incursion dispersal and current distribution on vegetable crops in New Zealand. N. Z. Plant Prot. 2009, 62, 136–144. [Google Scholar] [CrossRef]
- Munyaneza, J.E. Zebra chip disease of potato: Biology, epidemiology, and management. Am. J. Potato Res. 2012, 89, 329–350. [Google Scholar] [CrossRef]
- Hansen, A.; Trumble, J.; Stouthamer, R.; Paine, T. A new huanglongbing species, “Candidatus Liberibacter psyllaurous,” found to infect tomato and potato, is vectored by the psyllid Bactericera cockerelli (Sulc). Appl. Environ. Microbiol. 2008, 74, 5862–5865. [Google Scholar] [CrossRef] [PubMed]
- Liefting, L.W.; Sutherland, P.W.; Ward, L.I.; Paice, K.L.; Weir, B.S.; Clover, G.R. A new ‘Candidatus Liberibacter’species associated with diseases of solanaceous crops. Plant Dis. 2009, 93, 208–214. [Google Scholar] [CrossRef]
- Munyaneza, J.E. Zebra chip disease, Candidatus Liberibacter, and potato psyllid: A global threat to the potato industry. Am. J. Potato Res. 2015, 92, 230–235. [Google Scholar] [CrossRef]
- Soliman, T.; Mourits, M.; Oude Lansink, A.; Van Der Werf, W. Economic justification for quarantine status–the case study of ‘Candidatus Liberibacter solanacearum’ in the European Union. Plant Pathol. 2013, 62, 1106–1113. [Google Scholar] [CrossRef]
- Cameron, P.; Walker, G.; Hodson, A.; Kale, A.; Herman, T. Trends in IPM and insecticide use in processing tomatoes in New Zealand. Crop Prot. 2009, 28, 421–427. [Google Scholar] [CrossRef]
- Goldson, S.; Bourdôt, G.; Brockerhoff, E.; Byrom, A.; Clout, M.; McGlone, M.; Nelson, W.; Popay, A.; Suckling, D.; Templeton, M. New Zealand pest management: Current and future challenges. J. R. Soc. N. Z. 2015, 45, 31–58. [Google Scholar] [CrossRef]
- Walker, J.; Park, N.; Clothier, B.; Manktelow, D.; Van_Den_Dijssel, C.; Hodson, A.; Barley, M.; Hodson-Kersey, L. Progress in pesticide risk reduction in New Zealand horticulture. N. Z. Plant Prot. 2009, 62, 321–327. [Google Scholar] [CrossRef]
- Rubio Covarrubias, O.Á.; Almeyda León, I.H.; Ireta Moreno, J.; Sánchez Salas, J.A.; Fernández Sosa, R.; Borbón Soto, J.T.; Díaz Hernández, C.; Garzón Tiznado, J.A.; Rocha Rodríguez, R.; Cadena Hinojosa, M.A. Distribución de la punta morada y Bactericera cockerelli Sulc. en las principales zonas productoras de papa en México. Agric. Técnica México 2006, 32, 201–211. [Google Scholar]
- Vega-Gutierrez, M.T.; Rodríguez-Maciel, J.C.; Diaz-Gomez, O.; Bujanos-Muniz, R.; Mota-Sanchez, D.; Martínez-Carrillo, J.L.; Lagunes-Tejeda, A.; Garzon-Tiznado, J.A. Susceptibility to insecticides in two Mexican populations of tomato-potato psyllid, Bactericera cockerelli (Sulc.) (Hemiptera: Triozidae). Agrociencia 2008, 42, 463–471. [Google Scholar]
- Guenthner, J.; Goolsby, J.; Greenway, G. Use and cost of insecticides to control potato psyllids and zebra chip on potatoes. Southwest. Entomol. 2012, 37, 263–270. [Google Scholar] [CrossRef]
- Anderson, J.A.; Walker, G.P.; Alspach, P.A.; Jeram, M.; Wright, P.J. Assessment of susceptibility to zebra chip and Bactericera cockerelli of selected potato cultivars under different insecticide regimes in New Zealand. Am. J. Potato Res. 2013, 90, 58–65. [Google Scholar] [CrossRef]
- Cerna, E.; Ochoa, Y.; Aguirre, L.; Flores, M.; Landeros, J. Determination of insecticide resistance in four populations of potato psillid Bactericera cockerelli (Sulc.) (Hemiptera: Triozidae). Phyton 2013, 82, 63. [Google Scholar]
- Chávez, E.C.; Bautista, O.H.; Flores, J.L.; Uribe, L.A.; Fuentes, Y.M.O. Insecticide-resistance ratios of three populations of Bactericera cockerelli (Hemiptera: Psylloidea: Triozidae) in regions of northern Mexico. Fla. Entomol. 2015, 98, 950–953. [Google Scholar] [CrossRef]
- Szczepaniec, A.; Varela, K.A.; Kiani, M.; Paetzold, L.; Rush, C.M. Incidence of resistance to neonicotinoid insecticides in Bactericera cockerelli across Southwest US. Crop Prot. 2019, 116, 188–195. [Google Scholar] [CrossRef]
- Malhotra, N.; Chen, K.H.-C.; Huang, J.-C.; Lai, H.-T.; Uapipatanakul, B.; Roldan, M.J.M.; Macabeo, A.P.G.; Ger, T.-R.; Hsiao, C.-D. Physiological effects of neonicotinoid insecticides on non-target aquatic animals—An updated review. Int. J. Mol. Sci. 2021, 22, 9591. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F. Insecticides mode of action in relation to their toxicity to non-target organisms. J. Environ. Anal. Toxicol. 2012, 4, S4-002. [Google Scholar]
- Mulé, R.; Sabella, G.; Robba, L.; Manachini, B. Systematic review of the effects of chemical insecticides on four common butterfly families. Front. Environ. Sci. 2017, 5, 32. [Google Scholar] [CrossRef]
- Özkara, A.; Akyıl, D.; Konuk, M. Pesticides, environmental pollution, and health. In Environmental Health Risk-Hazardous Factors to Living Species; IntechOpen: London, UK, 2016. [Google Scholar]
- Sánchez-Bayo, F. Ecological impacts of insecticides. In Insecticides: Advances in Integrated Pest Management; IntechOpen: London, UK, 2012; pp. 61–90. [Google Scholar]
- Yadav, I.C.; Devi, N.L. Pesticides classification and its impact on human and environment. Environ. Sci. Eng. 2017, 6, 140–158. [Google Scholar]
- Veronesi, E.R.; Thompson, C.J.; Goldson, S.L. Insect biological control of the tomato-potato psyllid Bactericera cockerelli, a review. N. Z. J. Crop Hortic. Sci. 2023, 1–17. [Google Scholar] [CrossRef]
- Veronesi, E.R.; Olaniyan, O.; London, H.; Saville, D.J.; Wratten, S.D. Potential inter-guild interactions to enhance biological control of Bactericera cockerelli on tomatoes: A laboratory and cage study. BioControl 2021, 66, 343–353. [Google Scholar] [CrossRef]
- Veronesi, E.R.; Wratten, S.D.; van Koten, C.; Goldson, S.L. Potential of the mirid bug Engytatus nicotianae, and the parasitic wasp Tamarixia triozae for the biological control of the tomato-potato psyllid; a cage greenhouse assay. N. Z. J. Crop Hortic. Sci. 2022, 1–8. [Google Scholar] [CrossRef]
- Veronesi, E.R.; Saville, D.J.; van Koten, C.; Wratten, S.D.; Goldson, S.L. Potential of the mirid bug, Engytatus nicotianae, for the biological control of the tomato-potato psyllid in greenhouses. Crop Prot. 2022, 156, 105941. [Google Scholar] [CrossRef]
- Harman, G.E. Multifunctional fungal plant symbionts: New tools to enhance plant growth and productivity. New Phytol. 2011, 189, 647–649. [Google Scholar] [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef]
- Kandula, D.R.W.; Stewart, A.; McDermid, J.; Hunt, J.S. Improving pasture establishment and yield with a Trichoderma bio-inoculant. In Proceedings of the 16th Biennial Australasian Plant Pathology Society Conference, Adelaide, Australia, 24–27 September 2007. [Google Scholar]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.; Marra, R.; Barbetti, M.; Li, H.; Woo, S.L.; Lorito, M. A novel role for Trichoderma secondary metabolites in the interactions with plants. Physiol. Mol. Plant Pathol. 2008, 72, 80–86. [Google Scholar] [CrossRef]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Ruocco, M.; Woo, S.; Lorito, M. Trichoderma secondary metabolites that affect plant metabolism. Nat. Prod. Commun. 2012, 7, 1934578X1200701133. [Google Scholar] [CrossRef]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Woo, S.L.; Lorito, M. Trichoderma–plant–pathogen interactions. Soil Biol. Biochem. 2008, 40, 1–10. [Google Scholar] [CrossRef]
- Ruano Rosa, D.; Lopez Herrera, C.J. Evaluation of Trichoderma spp. as biocontrol agents against avocado white root rot. Biol. Control 2009, 51, 66–71. [Google Scholar] [CrossRef]
- Di Lelio, I.; Coppola, M.; Comite, E.; Molisso, D.; Lorito, M.; Woo, S.L.; Pennacchio, F.; Rao, R.; Digilio, M.C. Temperature differentially influences the capacity of Trichoderma species to induce plant defense responses in tomato against insect pests. Front. Plant Sci. 2021, 12, 678830. [Google Scholar] [CrossRef] [PubMed]
- Coppola, M.; Cascone, P.; Di Lelio, I.; Woo, S.L.; Lorito, M.; Rao, R.; Pennacchio, F.; Guerrieri, E.; Digilio, M.C. Trichoderma atroviride P1 colonization of tomato plants enhances both direct and indirect defense barriers against insects. Front. Physiol. 2019, 10, 813. [Google Scholar] [CrossRef] [PubMed]
- Coppola, M.; Cascone, P.; Chiusano, M.L.; Colantuono, C.; Lorito, M.; Pennacchio, F.; Rao, R.; Woo, S.L.; Guerrieri, E.; Digilio, M.C. Trichoderma harzianum enhances tomato indirect defense against aphids. Insect Sci. 2017, 24, 1025–1033. [Google Scholar] [CrossRef]
- Hoitink, H.; Madden, L.; Dorrance, A. Systemic resistance induced by Trichoderma spp.: Interactions between the host, the pathogen, the biocontrol agent, and soil organic matter quality. Phytopathology 2006, 96, 186–189. [Google Scholar] [CrossRef]
- McCormick, A.C.; Unsicker, S.B.; Gershenzon, J. The specificity of herbivore-induced plant volatiles in attracting herbivore enemies. Trends Plant Sci. 2012, 17, 303–310. [Google Scholar] [CrossRef]
- Clavijo McCormick, A.; Boeckler, G.A.; Köllner, T.G.; Gershenzon, J.; Unsicker, S.B. The timing of herbivore-induced volatile emission in black poplar (Populus nigra) and the influence of herbivore age and identity affect the value of individual volatiles as cues for herbivore enemies. BMC Plant Biol. 2014, 14, 304. [Google Scholar] [CrossRef]
- van Zijll de Jong, E.; Kandula, J.; Rostás, M.; Kandula, D.; Hampton, J.; Mendoza-Mendoza, A. Fungistatic activity mediated by volatile organic compounds is isolate-dependent in Trichoderma sp. “atroviride B”. J. Fungi 2023, 9, 238. [Google Scholar] [CrossRef]
- Effah, E.; Barrett, D.P.; Peterson, P.G.; Wargent, J.J.; Potter, M.A.; Holopainen, J.K.; Clavijo McCormick, A. Herbivory and attenuated UV radiation affect volatile emissions of the invasive weed Calluna vulgaris. Molecules 2020, 25, 3200. [Google Scholar] [CrossRef] [PubMed]
- Effah, E.; Barrett, D.P.; Peterson, P.G.; Godfrey, A.J.R.; Potter, M.A.; Holopainen, J.K.; Clavijo McCormick, A. Natural variation in volatile emissions of the invasive weed Calluna vulgaris in New Zealand. Plants 2020, 9, 283. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Pérez-Hedo, M.; Bouagga, S.; Jaques, J.A.; Flors, V.; Urbaneja, A. Tomato plant responses to feeding behavior of three zoophytophagous predators (Hemiptera: Miridae). Biol. Control 2015, 86, 46–51. [Google Scholar] [CrossRef]
- Snyder, W.E.; Wise, D.H. Predator interference and the establishment of generalist predator populations for biocontrol. Biol. Control 1999, 15, 283–292. [Google Scholar] [CrossRef]
- Xu, X.; Robinson, J.; Jeger, M.; Jeffries, P. Using combinations of biocontrol agents to control Botrytis cinerea on strawberry leaves under fluctuating temperatures. Biocontrol Sci. Technol. 2010, 20, 359–373. [Google Scholar] [CrossRef]
- Chong, J.-H.; Oetting, R.D. Intraguild predation and interference by the mealybug predator Cryptolaemus montrouzieri on the parasitoid Leptomastix dactylopii. Biocontrol Sci. Technol. 2007, 17, 933–944. [Google Scholar] [CrossRef]
- Holopainen, J.K.; Gershenzon, J. Multiple stress factors and the emission of plant VOCs. Trends Plant Sci. 2010, 15, 176–184. [Google Scholar] [CrossRef]
- Turlings, T.C.; Bernasconi, M.; Bertossa, R.; Bigler, F.; Caloz, G.; Dorn, S. The induction of volatile emissions in maize by three herbivore species with different feeding habits: Possible consequences for their natural enemies. Biol. Control 1998, 11, 122–129. [Google Scholar] [CrossRef]
- Tun, K.M.; Minor, M.; Jones, T.; McCormick, A.C. Volatile profiling of fifteen willow species and hybrids and their responses to giant willow aphid infestation. Agronomy 2020, 10, 1404. [Google Scholar] [CrossRef]
- Dicke, M. Induced plant volatiles: Plant body odours structuring ecological networks. New Phytol. 2016, 210, 10–12. [Google Scholar] [CrossRef] [PubMed]
- Schwartzberg, E.G.; Böröczky, K.; Tumlinson, J.H. Pea aphids, Acyrthosiphon pisum, suppress induced plant volatiles in broad bean, Vicia faba. J. Chem. Ecol. 2011, 37, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Najar-Rodriguez, A.J.; Friedli, M.; Klaiber, J.; Dorn, S. Aphid-deprivation from Brassica plants results in increased isothiocyanate release and parasitoid attraction. Chemoecology 2015, 25, 303–311. [Google Scholar] [CrossRef]
- Frago, E.; Mala, M.; Weldegergis, B.T.; Yang, C.; McLean, A.; Godfray, H.C.J.; Gols, R.; Dicke, M. Symbionts protect aphids from parasitic wasps by attenuating herbivore-induced plant volatiles. Nat. Commun. 2017, 8, 1860. [Google Scholar] [CrossRef]
- Casteel, C.L.; Hansen, A.K.; Walling, L.L.; Paine, T.D. Manipulation of plant defense responses by the tomato psyllid (Bactericera cockerelli) and its associated endosymbiont Candidatus Liberibacter psyllaurous. PLoS ONE 2012, 7, e35191. [Google Scholar] [CrossRef]
- Lins, J.C.; van Loon, J.J.; Bueno, V.H.; Lucas-Barbosa, D.; Dicke, M.; van Lenteren, J.C. Response of the zoophytophagous predators Macrolophus pygmaeus and Nesidiocoris tenuis to volatiles of uninfested plants and to plants infested by prey or conspecifics. BioControl 2014, 59, 707–718. [Google Scholar] [CrossRef]
- Van Oudenhove, L.; Mailleret, L.; Fauvergue, X. Infochemical use and dietary specialization in parasitoids: A meta-analysis. Ecol. Evol. 2017, 7, 4804–4811. [Google Scholar] [CrossRef]
- Dickens, J.C. Predator–prey interactions: Olfactory adaptations of generalist and specialist predators. Agric. For. Entomol. 1999, 1, 47–54. [Google Scholar] [CrossRef]
- Clavijo Mccormick, A.; Gershenzon, J.; Unsicker, S.B. Little peaks with big effects: Establishing the role of minor plant volatiles in plant–insect interactions. Plant Cell Environ. 2014, 37, 1836–1844. [Google Scholar] [CrossRef]
- Cardé, R.T.; Bell, W.J.; Vet, L.E.; Lewis, W.J.; Cardé, R.T. Parasitoid foraging and learning. In Chemical Ecology of Insects 2; Springer: Berlin/Heidelberg, Germany, 1995; pp. 65–101. [Google Scholar]
- Vet, L.E.; Groenewold, A.W. Semiochemicals and learning in parasitoids. J. Chem. Ecol. 1990, 16, 3119–3135. [Google Scholar] [CrossRef]
- Ardanuy, A.; Albajes, R.; Turlings, T.C. Innate and learned prey-searching behavior in a generalist predator. J. Chem. Ecol. 2016, 42, 497–507. [Google Scholar] [CrossRef] [PubMed]
- de Boer, J.; Dicke, M. Olfactory learning by predatory arthropods. Anim. Biol. 2006, 56, 143–155. [Google Scholar] [CrossRef]
- Koricheva, J.; Nykänen, H.; Gianoli, E. Meta-analysis of trade-offs among plant antiherbivore defenses: Are plants jacks-of-all-trades, masters of all? Am. Nat. 2004, 163, E64–E75. [Google Scholar] [CrossRef] [PubMed]
- Ballhorn, D.J.; Kautz, S.; Lion, U.; Heil, M. Trade-offs between direct and indirect defences of lima bean (Phaseolus lunatus). J. Ecol. 2008, 96, 971–980. [Google Scholar] [CrossRef]
- Rudgers, J.A.; Strauss, S.Y.; Wendel, J.F. Trade-offs among anti-herbivore resistance traits: Insights from Gossypieae (Malvaceae). Am. J. Bot. 2004, 91, 871–880. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatment | Eggs | Nymphs | Adults | Leaves with TPP |
|---|---|---|---|---|
| Control | 6.82% | 6.82% | 8.00% | 6.72% |
| Trichoderma | 6.88% | 6.18% *** | 8.44% | 6.61% |
| Engytatus | 3.25% *** | 1.11% *** | 5.44% *** | 4.50% ** |
| Combined | 5.87% *** | 7.14% | 6.72% * | 3.98% *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Veronesi, E.R.; Cairns, S.M.; Alizadeh, H.; Hampton, J.; Maris, R.; Godsoe, W.; Goldson, S.L.; Clavijo McCormick, A. Individual and Combined Effects of Predatory Bug Engytatus nicotianae and Trichoderma atroviride in Suppressing the Tomato Potato Psyllid Bactericera cockerelli in Greenhouse Grown Tomatoes. Agronomy 2023, 13, 3019. https://doi.org/10.3390/agronomy13123019
Veronesi ER, Cairns SM, Alizadeh H, Hampton J, Maris R, Godsoe W, Goldson SL, Clavijo McCormick A. Individual and Combined Effects of Predatory Bug Engytatus nicotianae and Trichoderma atroviride in Suppressing the Tomato Potato Psyllid Bactericera cockerelli in Greenhouse Grown Tomatoes. Agronomy. 2023; 13(12):3019. https://doi.org/10.3390/agronomy13123019
Chicago/Turabian StyleVeronesi, Emiliano R., Sarah M. Cairns, Hossein Alizadeh, John Hampton, Robbie Maris, William Godsoe, Stephen L. Goldson, and Andrea Clavijo McCormick. 2023. "Individual and Combined Effects of Predatory Bug Engytatus nicotianae and Trichoderma atroviride in Suppressing the Tomato Potato Psyllid Bactericera cockerelli in Greenhouse Grown Tomatoes" Agronomy 13, no. 12: 3019. https://doi.org/10.3390/agronomy13123019