
Comparative Study of Photosynthesis Performance of Herbicide-Treated Young Triticale Plants during Drought and Waterlogging Stress




Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatments
2.2. Biometric Parameters
2.3. Leaf Pigment Content
2.4. Chlorophyll a Fluorescence
2.5. Gas Exchange Parameters
2.6. Recovery and Resilience Indices of Selected Parameters
2.7. Statistical Analysis
3. Results
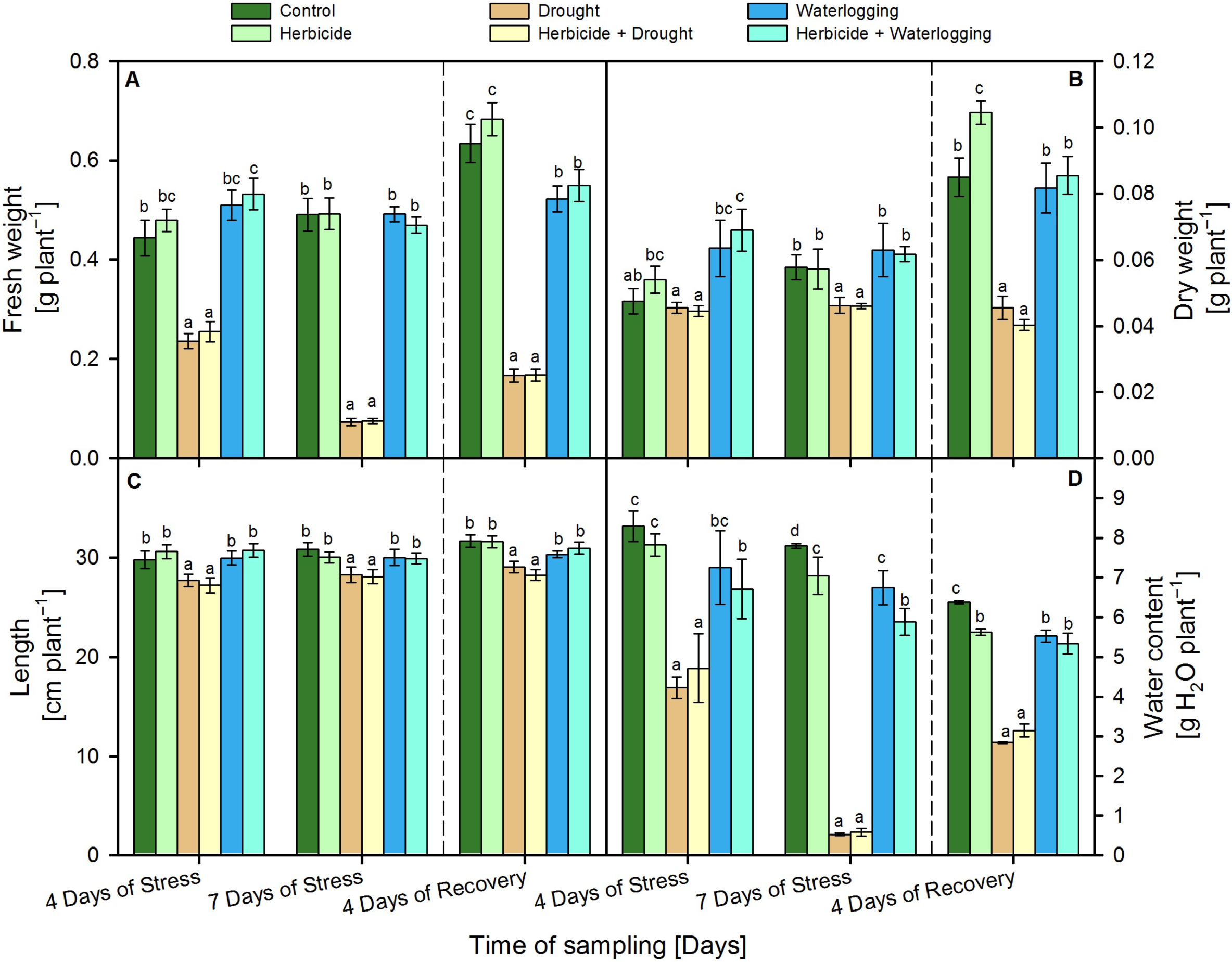
3.1. Biometric Parameters
3.2. Recovery and Resilience Indices of Biometric Parameters
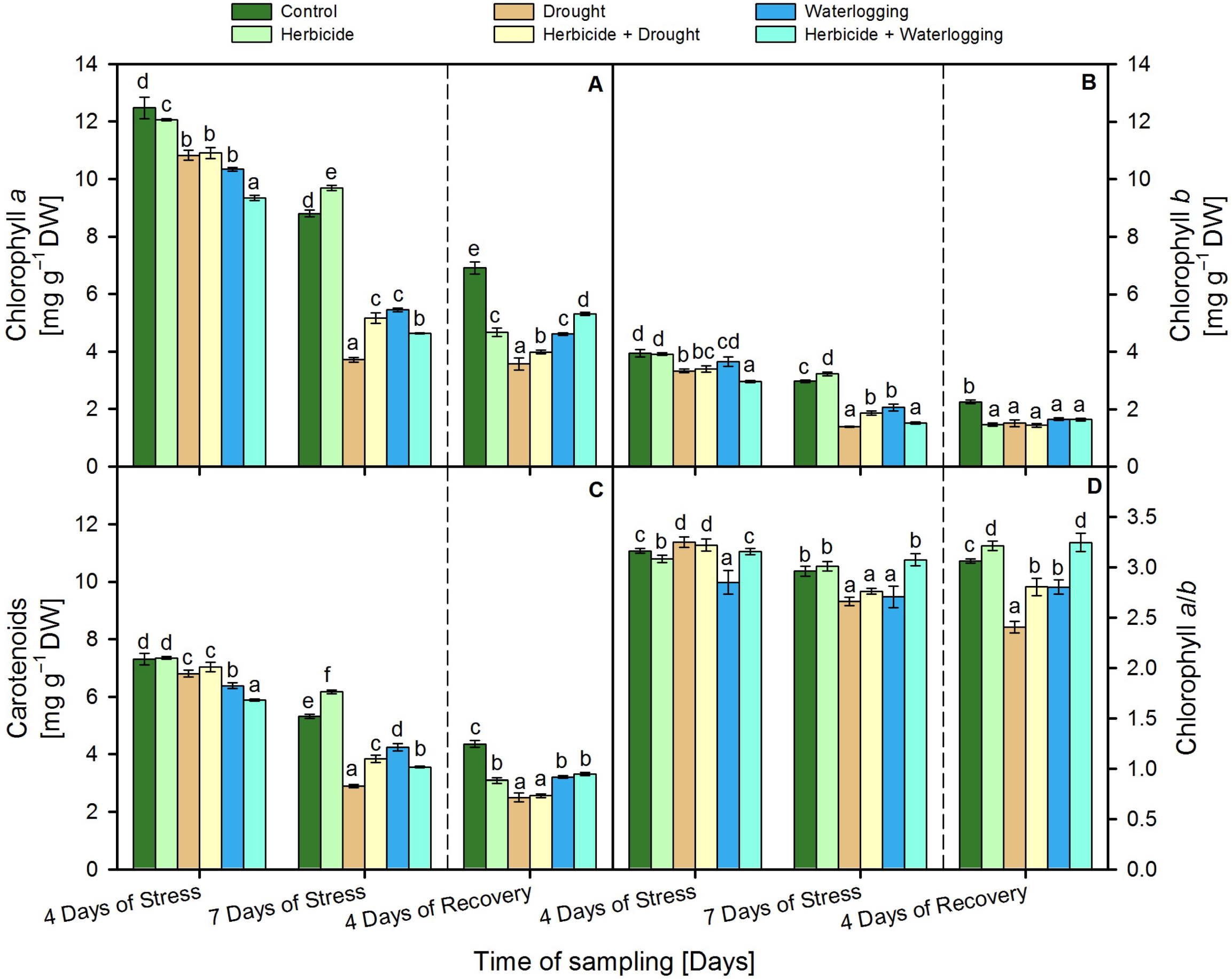
3.3. Leaf Pigment Content
3.4. Leaf Gas Exchange Parameters
3.5. Indices of Recovery and Resilience of Photosynthesis
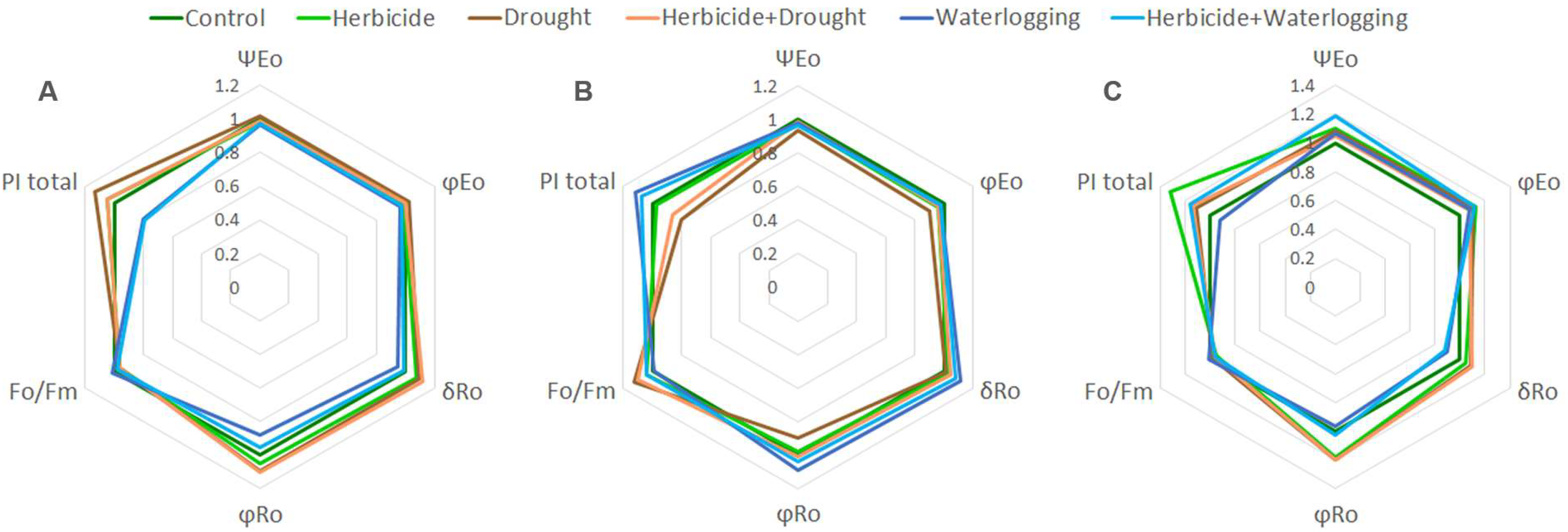
3.6. Parameters of Chlorophyll a Fluorescence
3.7. Spider Plot Presentation of Parameters of Chlorophyll a Fluorescence
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Nakka, S.; Jugulam, M.; Peterson, D.; Asif, M. Herbicide resistance: Development of wheat production systems and current status of resistant weeds in wheat cropping systems. Crop J. 2019, 7, 750–760. [Google Scholar] [CrossRef]
- Pesticide Properties DataBase. Available online: https://sitem.herts.ac.uk/aeru/ppdb/en/atoz.htm (accessed on 24 July 2023).
- Mutava, R.N.; Prince, S.J.K.; Syed, N.H.; Song, L.; Valliyodan, B.; Chen, W.; Nguyen, H.T. Understanding abiotic stress tolerance mechanisms in soybean: A comparative evaluation of soybean response to drought and flooding stress. Plant Physiol. Biochem. 2015, 86, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Qi, M.; Liu, X.; Li, Y.; Song, H.; Yin, Z.; Zhang, F.; He, Q.; Xu, Z.; Zhou, G. Photosynthetic resistance and resilience under drought, flooding and rewatering in maize plants. Photosynth. Res. 2021, 148, 1–15. [Google Scholar] [CrossRef]
- Xu, Z.; Zhou, G.; Shimizu, H. Plant responses to drought and rewatering. Plant Signal. Behav. 2010, 5, 649–654. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Sharif, R.; Xu, X.; Chen, X. Mechanisms of waterlogging tolerance in plants: Research progress and prospects. Front. Plant Sci. 2021, 11, 627331. [Google Scholar] [CrossRef]
- Todorova, D.; Aleksandrov, V.; Anev, S.; Sergiev, I. Photosynthesis Alterations in Wheat Plants Induced by Herbicide, Soil Drought or Flooding. Agronomy 2022, 12, 390. [Google Scholar] [CrossRef]
- Blum, A. The Abiotic Stress Response and Adaptation of Triticale—A Review. Cereal Res. Commun. 2014, 42, 359–375. [Google Scholar] [CrossRef] [Green Version]
- Arsenuik, E. Triticale Abiotic Stresses—An Overview. In Triticale; Eudes, A., Ed.; Springer International Publishing: Cham, Switzerland, 2015; pp. 69–81. [Google Scholar] [CrossRef]
- Mergoum, M.; Singh, P.K.; Peña, R.J.; Lozano-del Río, A.J.; Cooper, K.V.; Salmon, D.F.; Gómez Macpherson, H. Triticale: A ‘‘New’’ Crop with Old Challenges. In Cereals. Handbook of Plant Breeding; Carena, M.J., Ed.; Springer: New York, NY, USA, 2009; Volume 3, pp. 267–287. [Google Scholar] [CrossRef]
- Tounekti, T.; Hernández, I.; Müller, M.; Khemira, H.; Munné-Bosch, S. Kinetin applications alleviate salt stress and improve the antioxidant composition of leaf extracts in Salvia officinalis. Plant Physiol. Biochem. 2011, 49, 1165–1176. [Google Scholar] [CrossRef] [PubMed]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Strasser, R.J.; Tsimilli-Michael, M.; Qiang, S.; Goltsev, V. Simultaneous in vivo recording of prompt and delayed fluorescence and 820-nm reflection changes during drying and after rehydration of the resurrection plant Haberlea rhodopensis. Biochim. Biophys. Acta BBA—Bioenerg. 2010, 1797, 1313–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li-Cor. Using the LI-6800 Portable Photosynthesis System, 2nd ed. 2022. Available online: https://www.licor.com/env/support/LI-6800/docs.html (accessed on 24 July 2023).
- Malik, A.I.; Colmer, D.T.; Lambers, H.; Schortemeyer, M. Changes in physiological and morphological traits of roots and shoots of wheat in response to different depths of waterlogging. Aust. J. Plant Physiol. 2001, 28, 1121–1131. [Google Scholar] [CrossRef]
- Abid, M.; Ali, S.; Qi, L.K.; Zahoor, R.; Tian, Z.; Jiang, D.; Snider, J.L.; Dai, T. Physiological and biochemical changes during drought and recovery periods at tillering and jointing stages in wheat (Triticum aestivum L.). Sci. Rep. 2018, 8, 4615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katam, R.; Shokri, S.; Murthy, N.; Singh, S.K.; Suravajhala, P.; Khan, M.N.; Bahmani, M.; Sakata, K.; Reddy, K.R. Proteomics, physiological, and biochemical analysis of cross tolerance mechanisms in response to heat and water stresses in soybean. PLoS ONE 2020, 15, e0233905. [Google Scholar] [CrossRef] [PubMed]
- Hura, T.; Hura, K.; Ostrowska, A.; Grzesiak, M.; Dziurka, K. The cell wall-bound phenolics as a biochemical indicator of soil drought resistance in winter triticale. Plant Soil Environ. 2013, 59, 189–195. [Google Scholar] [CrossRef]
- Meena, V.; Kaushik, M.K.; Verma, A.; Upadhayay, B.; Meena, S.K.; Bhimwal, J.P. Effect of herbicide and their combinations on growth and productivity of wheat (Triticum aestivum L.) under late sown condition. Int. J. Chem. Stud. 2017, 5, 1512–1516. [Google Scholar]
- Zarzecka, K.; Gugała, M.; Mystkowska, I.; Sikorska, A. Changes in dry weight and starch content in potatounder the effect of herbicides and biostimulants. Plant Soil Environ. 2021, 67, 202–207. [Google Scholar] [CrossRef]
- Zulfiqar, S.; Sharif, S.; Saeed, M.; Tahir, A. Role of Carotenoids in Photosynthesis. In Carotenoids: Structure and Function in the Human Body; Zia-Ul-Haq, M., Dewanjee, S., Riaz, M., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 147–187. [Google Scholar] [CrossRef]
- Wingler, A.; Lea, P.J.; Quick, W.P.; Leegood, R.C. Photorespiration: Metabolic pathways and their role in stress protection. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2000, 355, 1517–1529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 102. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Moustakas, M.; Sperdouli, I.; Moustaka, J.; Șaș, B.; İșgören, S.; Morales, F. Mechanistic Insights on Salicylic Acid Mediated Enhancement of Photosystem II Function in Oregano Seedlings Subjected to Moderate Drought Stress. Plants 2023, 12, 518. [Google Scholar] [CrossRef]
- Niyogi, K.K. Safety valves for photosynthesis. Curr. Opin. Plant Biol. 2000, 3, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Mattila, M.; Mishra, S.; Tyystjärvi, T.; Tyystjärvi, E. Singlet oxygen production by photosystem II is caused by misses of the oxygen evolving complex. New Phytol. 2023, 237, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Tyystjärvi, E. Photoinhibition of Photosystem II and photodamage of the oxygen evolving manganese cluster. Coord. Chem. Rev. 2008, 252, 361–376. [Google Scholar] [CrossRef]
- Pandey, J.; Devadasu, E.; Saini, D.; Marriboina, K.; Raghavendra, A.S.; Subramanyam, R. Reversible changes in structure and function of photosynthetic apparatus of pea (Pisum sativum) leaves under drought stress. Plant J. 2023, 113, 60–74. [Google Scholar] [CrossRef]
- Larouk, C.; Gabon, F.; Kehel, Z.; Djekoun, A.; Nachit, M.; Amri, A. Chlorophyll Fluorescence and Drought Tolerance in a Mapping Population of Durum Wheat. Contemp. Agricult. 2021, 70, 123–134. [Google Scholar] [CrossRef]
- Sherstneva, O.; Khlopkov, A.; Gromova, E.; Yudina, L.; Vetrova, Y.; Pecherina, A.; Kuznetsova, D.; Krutova, E.; Sukhov, V.; Vodeneev, V. Analysis of chlorophyll fluorescence parameters as predictors of biomass accumulation and tolerance to heat and drought stress of wheat (Triticum aestivum) plants. Funct. Plant Biol. 2022, 49, 155–169. [Google Scholar] [CrossRef]
- Barboričová, M.; Filaček, A.; Mlynáriková Vysoká, D.; Gašparovič, K.; Živčák, M.; Brestič, M. Sensitivity of fast chlorophyll fluorescence parameters to combined heat and drought stress in wheat genotypes. CAAS Agricult. J. 2022, 68, 309–316. [Google Scholar] [CrossRef]
- Wu, X.; Tang, Y.; Li, C.; Wu, C.; Huang, G. Chlorophyll Fluorescence and Yield Responses of Winter Wheat to Waterlogging at Different Growth Stages. Plant Prod. Sci. 2015, 18, 284–294. [Google Scholar] [CrossRef] [Green Version]
- Stefanov, M.; Rashkov, G.; Borisova, P.; Apostolova, E. Sensitivity of the Photosynthetic Apparatus in Maize and Sorghum under Different Drought Levels. Plants 2023, 12, 1863. [Google Scholar] [CrossRef] [PubMed]
- Vasquez-Robinet, C.; Mane, S.P.; Ulanov, A.V.; Watkinson, J.I.; Stromberg, V.K.; De Koeyer, D.; Schafleitner, R.; Willmot, D.B.; Bonierbale, M.; Bohnert, H.J.; et al. Physiological and molecular adaptations to drought in Andean potato genotypes. J. Exp. Bot. 2008, 59, 2109–2123. [Google Scholar] [CrossRef] [Green Version]
- Souza, R.P.; Machado, E.C.; Silva, J.A.B.; Lagôa, A.M.M.A.; Silveira, J.A.G. Photosynthetic gas exchange, chlorophyll fluorescence and some associated metabolic changes in cowpea (Vigna unguiculata) during water stress and recovery. Environ. Exp. Bot. 2004, 51, 45–56. [Google Scholar] [CrossRef]
- Li, Y.; Fang, A.; Zhang, T.; Zhang, S.; Zhu, W.; Zhou, Y. Exogenous Calcium Improves Photosynthetic Capacity of Pinus sylvestris var. mongolica under Drought. Forests 2022, 13, 2155. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Treatment | Recovery | Resilience |
|---|---|---|---|
| FW | Herbicide | 39 ⮙ | 8 ⮘⮚ |
| Drought | 129 ⮙⮙⮙⮙ | −74 ⮛⮛⮛ | |
| Herbicide + Drought | 123 ⮙⮙⮙⮙ | −74 ⮛⮛⮛ | |
| Waterlogging | 6 ⮘⮚ | −18 ⮛ | |
| Herbicide + Waterlogging | 17 ⮙ | −13 ⮛ | |
| DW | Herbicide | 82 ⮙⮙⮙ | 23 ⮙ |
| Drought | −2 ⮘⮚ | −46 ⮛⮛ | |
| Herbicide + Drought | −12 ⮛ | −53 ⮛⮛ | |
| Waterlogging | 30 ⮙ | −4 ⮘⮚ | |
| Herbicide + Waterlogging | 38 ⮙ | 1 ⮘⮚ | |
| WC | Herbicide | −20 ⮛ | −12 ⮛ |
| Drought | 443 ⮙⮙⮙⮙ | −55 ⮛⮛ | |
| Herbicide + Drought | 446 ⮙⮙⮙⮙ | −51 ⮛⮛ | |
| Waterlogging | −18 ⮛ | −13 ⮛ | |
| Herbicide + Waterlogging | −9 ⮘⮚ | −16 ⮛ |
| Trait | Treatment | Recovery | Resilience |
|---|---|---|---|
| An | Herbicide | 12 ⮙ | 16 ⮙ |
| Drought | −277 ⮛⮛⮛⮛ | −34 ⮛ | |
| Herbicide + Drought | −175 ⮛⮛⮛⮛ | −71 ⮛⮛⮛ | |
| Waterlogging | 322 ⮙⮙⮙⮙ | −27 ⮛ | |
| Herbicide + Waterlogging | 649 ⮙⮙⮙⮙ | −21 ⮛ | |
| E | Herbicide | 21 ⮙ | 57 ⮙⮙ |
| Drought | 1133 ⮙⮙⮙⮙ | −38 ⮛ | |
| Herbicide + Drought | 467 ⮙⮙⮙⮙ | −56 ⮛⮛ | |
| Waterlogging | 81 ⮙⮙⮙ | −39 ⮛ | |
| Herbicide + Waterlogging | 180 ⮙⮙⮙⮙ | −35 ⮛ | |
| Gs | Herbicide | 42 ⮙⮙ | 84 ⮙⮙⮙ |
| Drought | 1705 ⮙⮙⮙⮙ | −6 ⮘⮚ | |
| Herbicide + Drought | 510 ⮙⮙⮙⮙ | −51 ⮛⮛ | |
| Waterlogging | 159 ⮙⮙⮙⮙ | −8 ⮘⮚ | |
| Herbicide + Waterlogging | 144 ⮙⮙⮙⮙ | −23 ⮛ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Todorova, D.; Aleksandrov, V.; Anev, S.; Sergiev, I. Comparative Study of Photosynthesis Performance of Herbicide-Treated Young Triticale Plants during Drought and Waterlogging Stress. Agronomy 2023, 13, 1992. https://doi.org/10.3390/agronomy13081992
Todorova D, Aleksandrov V, Anev S, Sergiev I. Comparative Study of Photosynthesis Performance of Herbicide-Treated Young Triticale Plants during Drought and Waterlogging Stress. Agronomy. 2023; 13(8):1992. https://doi.org/10.3390/agronomy13081992
Chicago/Turabian StyleTodorova, Dessislava, Vladimir Aleksandrov, Svetoslav Anev, and Iskren Sergiev. 2023. "Comparative Study of Photosynthesis Performance of Herbicide-Treated Young Triticale Plants during Drought and Waterlogging Stress" Agronomy 13, no. 8: 1992. https://doi.org/10.3390/agronomy13081992