
The Mechanisms of Different Light Supply Patterns in the Nutrient Uptake and Chlorophyll Fluorescence of Hydroponic Lettuce
 ,
,
Abstract
:1. Introduction
1.1. The Application of Light Supply Modes in Plant Factories
1.2. The Characteristics of Light Supply Modes
1.3. The Effects of Light Supply Mode on Plants
1.4. The Research Purpose of This Article
2. Materials and Methods
2.1. Experimental Site
2.2. Plant Materials, Hydroponic Setup and Growing Conditions
2.3. Experimental Design and Treatment
2.4. Nutrient Solution Preparation and Management
2.5. Growth and Yield of Lettuce
2.6. Nutrient Contents of Lettuce
2.6.1. C, N, P, K Content
2.6.2. Ca, Mg, Fe, Mn, Zn, Cu Content
2.7. Photosynthetic Pigment Measurement
2.8. Photosynthetic Parameters
2.9. Chlorophyll Fluorescence Parameters
2.10. Data Processing and Analysis
3. Results
3.1. Growth Measurement
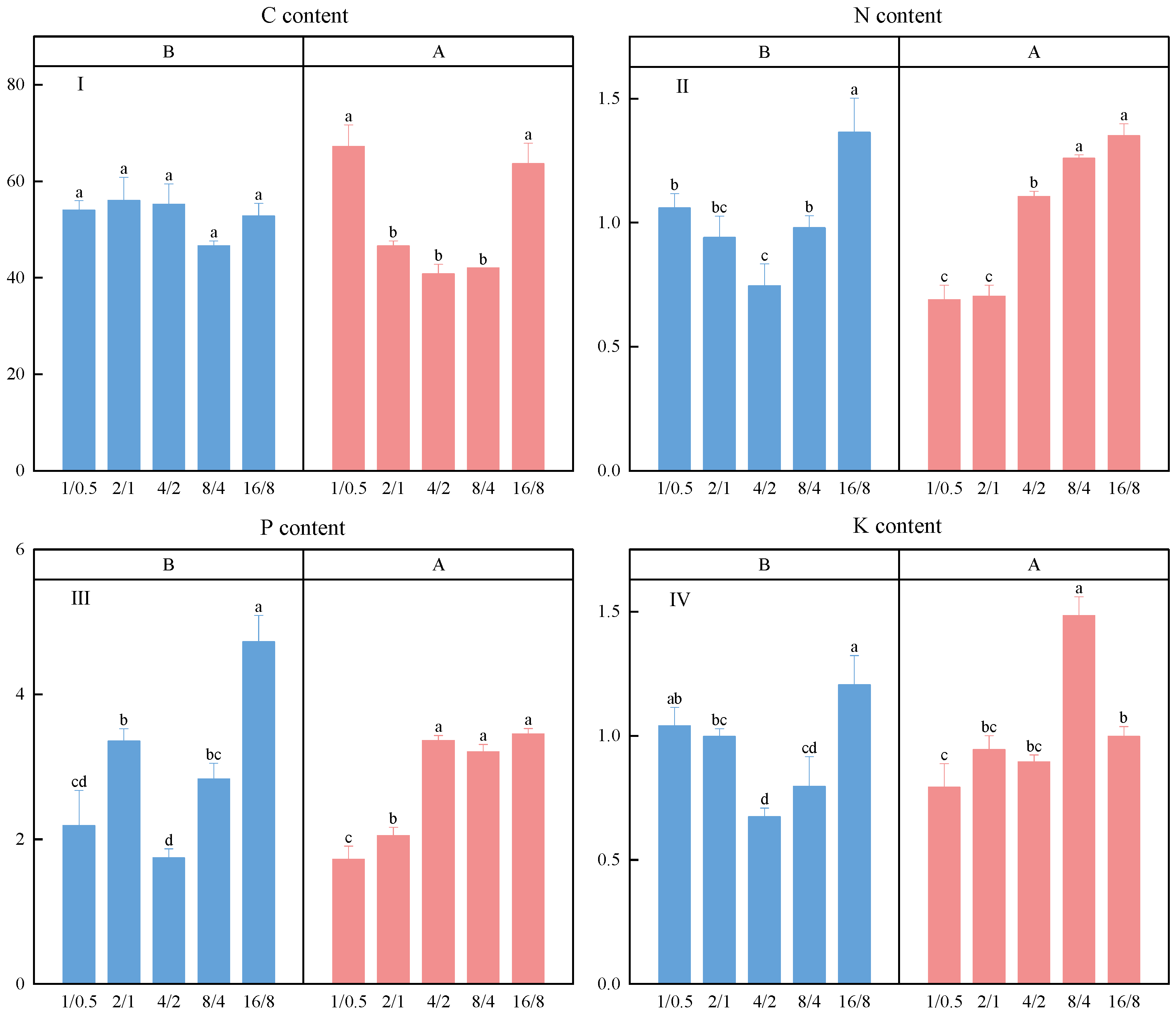
3.2. C, N, P, K Content
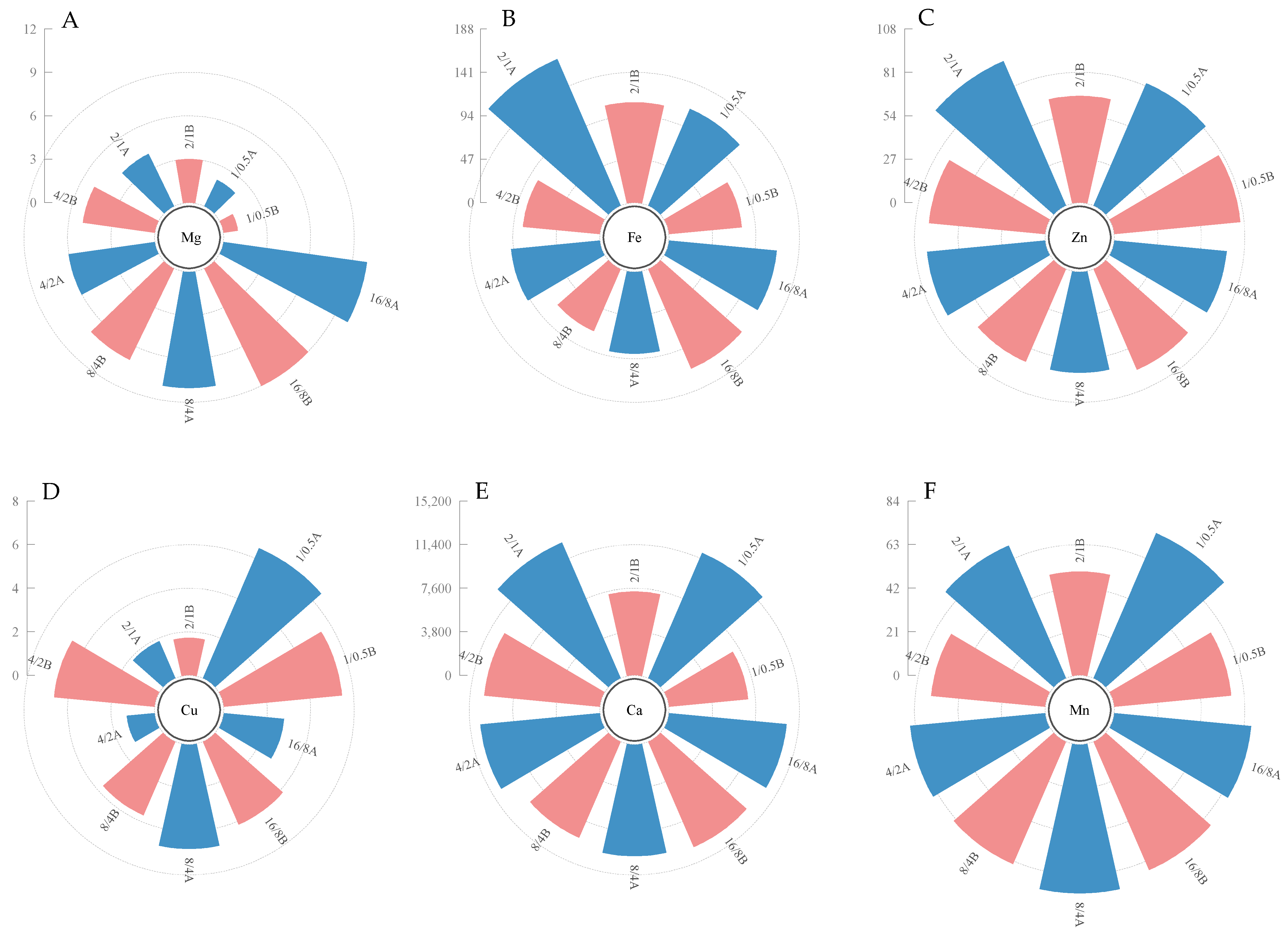
3.3. Mg, Ca, Fe, Mn, Zn, Cu Content
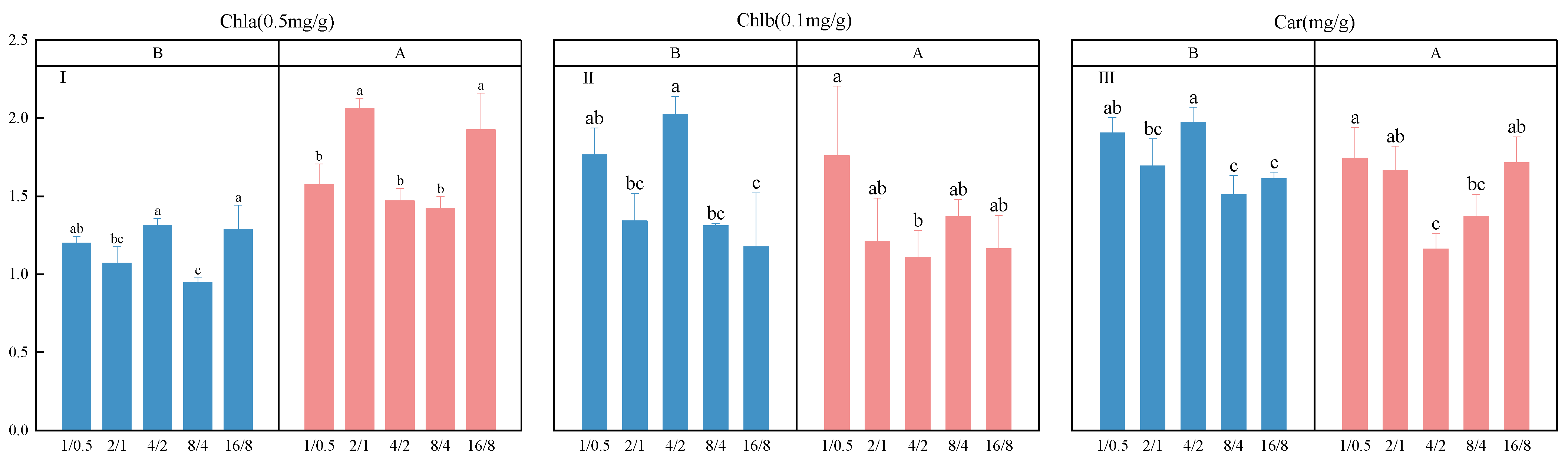
3.4. Photosynthetic Pigment Measurement
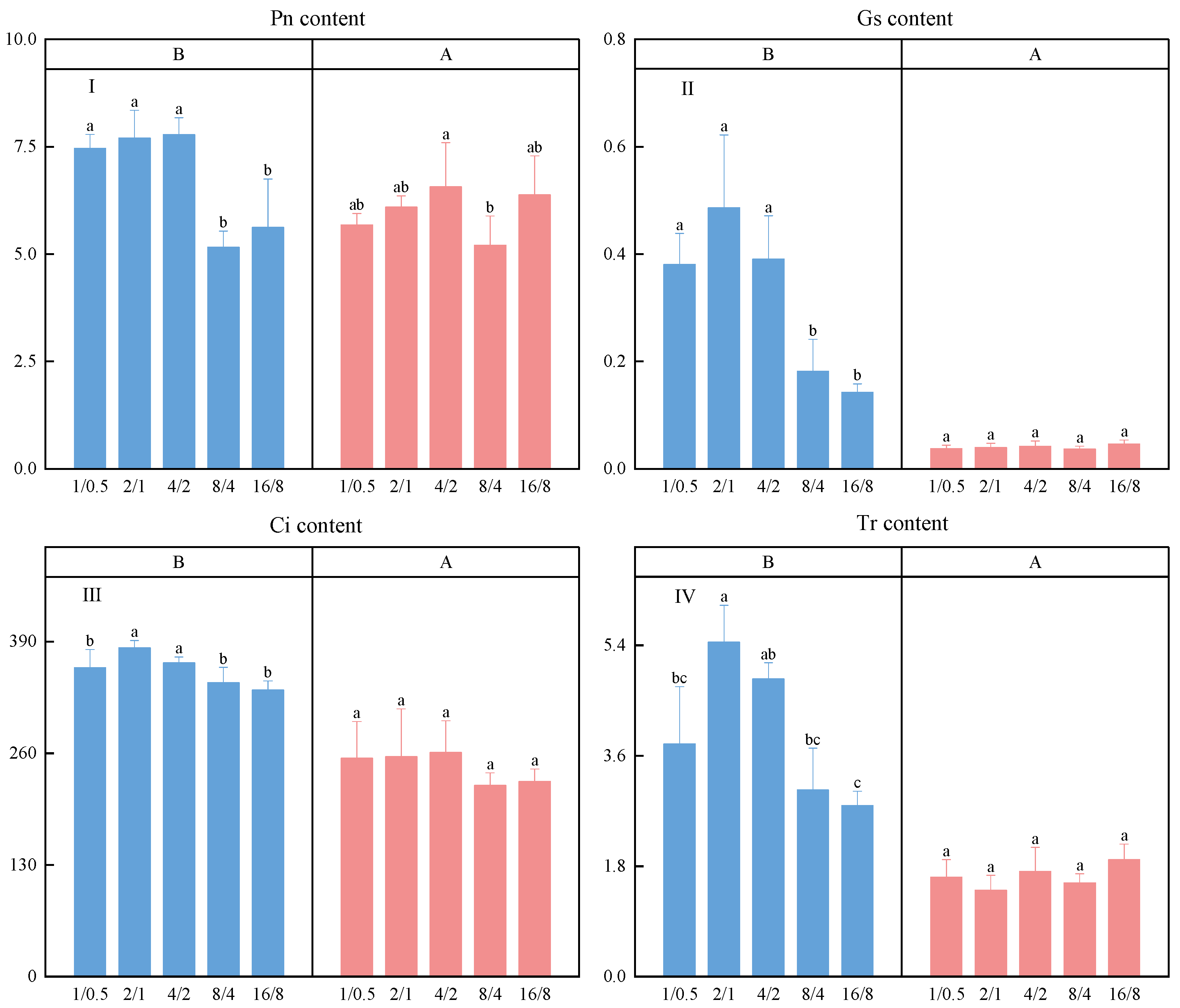
3.5. Photosynthetic Parameters
3.6. Chlorophyll Fluorescence Parameters
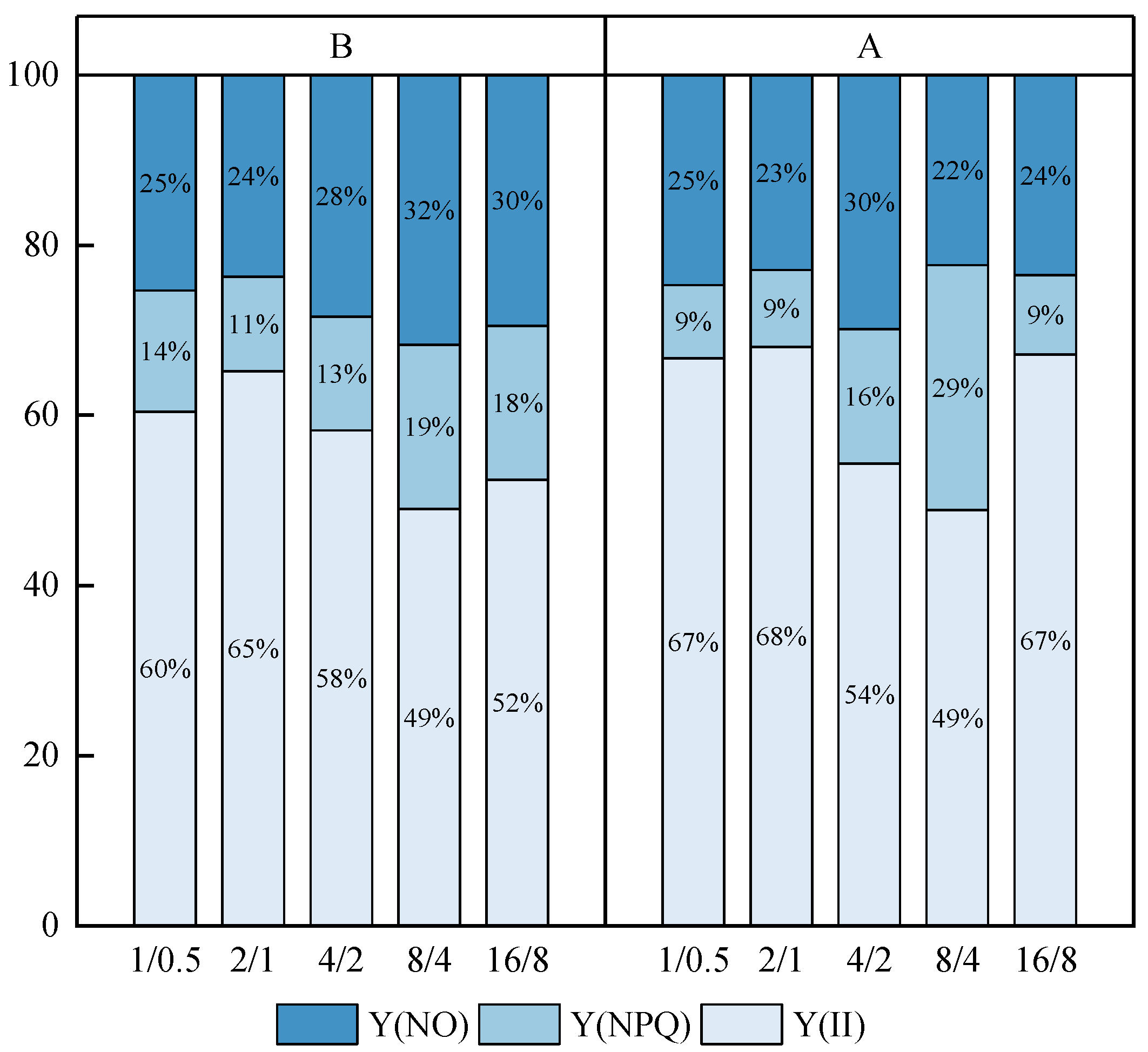
3.6.1. Energy Allocation of Antenna Pigment Absorption [Y(II), Y(NPQ), Y(NO)]
3.6.2. Effects of Light Stress on Photosynthetic Apparatus (Fv/Fm, φP0, PIABS, PICS)
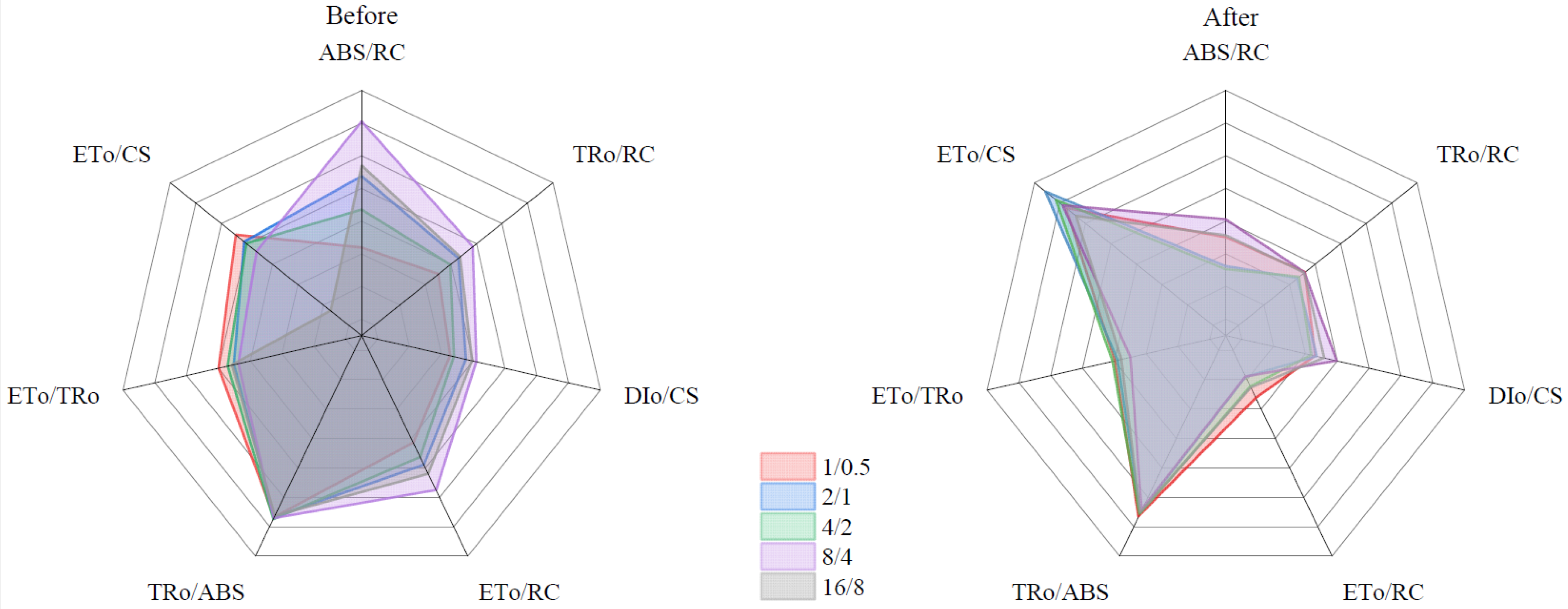
3.6.3. PSII Reaction Center Changes
4. Discussion
4.1. The Effect of Light Supply Modes on the Yield and Quality of Lettuce
4.2. The Effect of Light Supply Mode on Nutrient Absorption
4.3. Response Mechanism of Light Stress to Light Supply Mode in Lettuce
5. Conclusions
- i.
- Intermittent light supply has significant effects on growth indicators, C, N, P, K, Mg, Fe, Ca, Mn, Zn, and Cu contents, chlorophyll a, chlorophyll b, carotene, photosynthetic indicators, and chlorophyll fluorescence indicators. The 2/1 treatment group had the best response in terms of lettuce growth, nutrient absorption and photosynthesis.
- ii.
- After continuous light supply, the growth indicators of lettuce all significantly increased; the N, P, K, and Cu contents significantly increased in the 4/2 and 8/4 treatments. Continuous light supply also stabilized the Mg, Ca, and Mn contents between the treatment groups. Stomatal closure leads to a decrease in the net photosynthesis rate, transpiration rate, and intercellular CO2 concentration. In summary, short-term continuous light supply can improve the yield and nutrient absorption of lettuce, but photosynthesis is hindered by stomatal closure.
- iii.
- Two light supply modes cause light stress in lettuce, which forces it to develop the ability to resist adversity, as evidenced by an increase in qI and ET0/CS.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, D.; Basu, C.; Meinhardt, W.M.; Roth, B. LEDs for energy efficient greenhouse lighting. Renew. Sustain. Energy Rev. 2015, 49, 139–147. [Google Scholar] [CrossRef] [Green Version]
- Darko, E.; Heydarizadeh, P.; Schoefs, B.; Sabzalian, M.R. Photosynthesis under artificial light: The shift in primary and secondary metabolism. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murase, H. The Latest Development of Laser Application Research in Plant Factory. Agric. Agric. Sci. Procedia 2015, 3, 4–8. [Google Scholar] [CrossRef] [Green Version]
- Panjai, L.; Noga, G.; Hunsche, M.; Fiebig, A. Optimal red light irradiation time to increase health-promoting compounds in tomato fruit postharvest. Sci. Hortic. 2019, 251, 189–196. [Google Scholar] [CrossRef]
- Dong, C.; Shao, L.; Liu, G.; Wang, M.; Liu, H.; Xie, B.; Li, B.; Fu, Y.; Liu, H. Photosynthetic characteristics, antioxidant capacity and biomass yield of wheat exposed to intermittent light irradiation with millisecond-scale periods. J. Plant Physiol. 2015, 184, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Folta1, K.M.; Childers, K.S. Light as a Growth Regulator: Controlling Plant Biology with Narrow-bandwidth Solid-state Lighting Systems. HortScience 2008, 43, 1957–1964. [Google Scholar] [CrossRef] [Green Version]
- Jung, W.S.; Chung, I.M.; Hwang, M.H.; Kim, S.H.; Yu, C.Y.; Ghimire, B.K. Application of Light-Emitting Diodes for Improving the Nutritional Quality and Bioactive Compound Levels of Some Crops and Medicinal Plants. Molecules 2021, 26, 1477. [Google Scholar] [CrossRef]
- Ahmed, H.A.; Tong, Y.-X.; Yang, Q.-C. Optimal control of environmental conditions affecting lettuce plant growth in a controlled environment with artificial lighting: A review. S. Afr. J. Bot. 2020, 130, 75–89. [Google Scholar] [CrossRef]
- Colonna, E.; Rouphael, Y.; Barbieri, G.; De Pascale, S. Nutritional quality of ten leafy vegetables harvested at two light intensities. Food Chem. 2016, 199, 702–710. [Google Scholar] [CrossRef]
- Braidot, E.; Petrussa, E.; Peresson, C.; Patui, S.; Bertolini, A.; Tubaro, F.; Wahlby, U.; Coan, M.; Vianello, A.; Zancani, M. Low-intensity light cycles improve the quality of lamb’s lettuce (Valerianella olitoria [L.] Pollich) during storage at low temperature. Postharvest Biol. Technol. 2014, 90, 15–23. [Google Scholar] [CrossRef]
- Paradiso, R.; Proietti, S. Light-Quality Manipulation to Control Plant Growth and Photomorphogenesis in Greenhouse Horticulture: The State of the Art and the Opportunities of Modern LED Systems. J. Plant Growth Regul. 2021, 41, 742–780. [Google Scholar] [CrossRef]
- He, R.; Zhang, Y.; Song, S.; Su, W.; Hao, Y.; Liu, H. UV-A and FR irradiation improves growth and nutritional properties of lettuce grown in an artificial light plant factory. Food Chem. 2021, 345, 128727. [Google Scholar] [CrossRef]
- Demotes, M.S.; Péron, T.; Corot, A.; Bertheloot, J.; Le Gourrierec, J.; Pelleschi, T.S.; Crespel, L.; Morel, P.; Huché Thélier, L.; Boumaza, R.; et al. Plant responses to red and far-red lights, applications in horticulture. Environ. Exp. Bot. 2016, 121, 4–21. [Google Scholar] [CrossRef]
- Chen, X.L.; Xue, X.Z.; Guo, W.Z.; Wang, L.C.; Qiao, X.J. Growth and nutritional properties of lettuce affected by mixed irradiation of white and supplemental light provided by light-emitting diode. Sci. Hortic. 2016, 200, 111–118. [Google Scholar] [CrossRef]
- Ginzburg, D.N.; Klein, J.D. LED pre-exposure shines a new light on drought tolerance complexity in lettuce (Lactuca sativa) and rocket (Eruca sativa). Environ. Exp. Bot. 2020, 180, 104240. [Google Scholar] [CrossRef]
- Heo, J.; Lee, C.; Chakrabarty, D.; Paek, K. Growth responses of marigold and salvia bedding plants as affected by monochromic or mixture radiation provided by a Light-Emitting Diode (LED). Plant Growth Regul. 2002, 38, 225–230. [Google Scholar] [CrossRef]
- Ohtake, N.; Ishikura, M.; Suzuki, H.; Yamori, W.; Goto, E. Continuous Irradiation with Alternating Red and Blue Light Enhances Plant Growth While Keeping Nutritional Quality in Lettuce. HortScience 2018, 53, 1804–1809. [Google Scholar] [CrossRef] [Green Version]
- Kuno, Y.; Shimizu, H.; Nakashima, H.; Miyasaka, J.; Ohdoi, K. Effects of Irradiation Patterns and Light Quality of Red and Blue Light-Emitting Diodes on Growth of Leaf Lettuce (Lactuca sativa L. “Greenwave”). Environ. Control Biol. 2017, 55, 129–135. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.L.; Yang, Q.C.; Song, W.P.; Wang, L.C.; Guo, W.Z.; Xue, X.Z. Growth and nutritional properties of lettuce affected by different alternating intervals of red and blue LED irradiation. Sci. Hortic. 2017, 223, 44–52. [Google Scholar] [CrossRef]
- Velez-Ramirez, A.I.; Heuvelink, E.; van Ieperen, W.; Vreugdenhil, D.; Millenaar, F.F. Continuous Light as a Way to Increase Greenhouse Tomato Production: Expected Challenges. In Proceedings of the VII International Symposium on Light in Horticulture, Wageningen, The Netherlands, 14–18 October 2012; pp. 51–57. [Google Scholar]
- Matsuda, R.; Yamano, T.; Murakami, K.; Fujiwara, K. Effects of spectral distribution and photosynthetic photon flux density for overnight LED light irradiation on tomato seedling growth and leaf injury. Sci. Hortic. 2016, 198, 363–369. [Google Scholar] [CrossRef]
- Zha, L.Y.; Zhang, Y.B.; Li, Z.G.; Liu, W.K. Effect of Continuous Red/Blue LED Light and Its Light Intensity on Growth and Mineral Elements Absorption of Lettuce. Spectrosc. Spectr. Anal. 2019, 39, 2474–2480. [Google Scholar] [CrossRef]
- Zha, L.; Zhang, Y.; Liu, W. Dynamic Responses of Ascorbate Pool and Metabolism in Lettuce to Long-term Continuous Light Provided by Red and Blue LEDs. Environ. Exp. Bot. 2019, 163, 15–23. [Google Scholar] [CrossRef]
- Avgoustaki, D.D.; Li, J.Y.; Xydis, G. Basil plants grown under intermittent light stress in a small-scale indoor environment: Introducing energy demand reduction intelligent technologies. Food Control 2020, 118, 107389. [Google Scholar] [CrossRef]
- Chen, X.L.; Yang, Q.C. Effects of intermittent light exposure with red and blue light emitting diodes on growth and carbohydrate accumulation of lettuce. Sci. Hortic. 2018, 234, 220–226. [Google Scholar] [CrossRef]
- Chen, Z.; Su, B. Influence of Medium Frequency Light/Dark Cycles on the Cultivation of Auxenochlorella pyrenoidosa. Appl. Sci. 2020, 10, 5093. [Google Scholar] [CrossRef]
- Liu, W.K.; Zhang, Y.B.; Zha, L.Y.; Liu, Y.F. Effects of Continuous Light Before Harvest on Nutrient Element Contents of Hydroponic Lettuce Cultivated Supplied With Three Nitrogen Levels and Two LED Red and Blue Light Qualities. Spectrosc. Spectr. Anal. 2020, 40, 3884–3889. [Google Scholar] [CrossRef]
- Shao, M.J.; Liu, W.K.; Zhou, C.B.; Wang, Q.; Li, B.S. Effects of High Light Duration and Frequencies on Growth and Nutrient Element Contents of Hydroponic Lettuce Cultivated Under LED Red and Blue Light LED. Spectrosc. Spectr. Anal. 2021, 41, 2853–2858. [Google Scholar] [CrossRef]
- Zha, L.Y.; Liu, W.K. Effect of continuous light with red and blue LED lamps on growth and characteristics of photosynthesis and chlorophyll fluorescence of five lettuce cultivars. J. Plant Physiol. 2017, 53, 1735–1741. [Google Scholar]
- Berger, S.; Benediktyová, Z.; Matouš, K.; Bonfig, K.; Mueller, M.J.; Nedbal, L.; Roitsch, T. Visualization of dynamics of plant–pathogen interaction by novel combination of chlorophyll fluorescence imaging and statistical analysis: Differential effects of virulent and avirulent strains of P. syringae and of oxylipins on A. thaliana. J. Exp. Bot. 2007, 58, 797–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant 2016, 38, 102. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, A.M.; Noga, G.; Hunsche, M. Alternating high and low intensity of blue light affects PSII photochemistry and raises the contents of carotenoids and anthocyanins in pepper leaves. Plant Growth Regul. 2015, 79, 275–285. [Google Scholar] [CrossRef]
- Lavaud, J.; Rousseau, B.; van Gorkom, H.J.; Etienne, A.L. Influence of the diadinoxanthin pool size on photoprotection in the marine planktonic diatom Phaeodactylum tricornutum. Plant Physiol. 2002, 129, 1398–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoagland, D.R.; Arnon, D.I. The water culture method for growing plants without soil. Circ. Calif. Agric. Exp. Stn. 1950, 347, 32. [Google Scholar]
- Li, H. (Ed.) Principles and Techniques of Plant Physiology and Biochemistry Experiments; Higher Education Press: Beijing, China, 2000; Volume 2, pp. 71–91. [Google Scholar]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Chen, X.L.; Guo, W.Z.; Xue, X.Z.; Beauty, M.M. Effects of LED spectrum combinations on the absorption of mineral elements of hydroponic lettuce. Spectrosc. Spectr. Anal. 2014, 34, 1394–1397. [Google Scholar] [CrossRef]
- Liu, W.K.; Zhang, Y.B.; Zha, L.Y. Effect of LED Red and Blue Continuous Lighting before Harvest on Growth and Nutrient Absorption of Hydroponic Lettuce Cultivated under Different Nitrogen Forms and Light Qualities LED. Spectrosc. Spectr. Anal. 2020, 40, 2215–2221. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Schansker, G.; Ladle, R.J.; Goltsev, V.; Bosa, K.; Allakhverdiev, S.I.; Brestic, M.; Bussotti, F.; Calatayud, A.; Dabrowski, P.; et al. Frequently asked questions about in vivo chlorophyll fluorescence: Practical issues. Photosynth. Res. 2014, 122, 121–158. [Google Scholar] [CrossRef] [PubMed]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [Green Version]
- Muller, P.; Li, X.P.; Niyogi, K.K. Non-photochemical quenching. A response to excess light energy. Plant Physiol. 2001, 125, 1558–1566. [Google Scholar] [CrossRef] [Green Version]
- Tholen, D.; Boom, C.; Noguchi, K.; Ueda, S.; Katase, T.; Terashima, I. The chloroplast avoidance response decreases internal conductance to CO2 diffusion in Arabidopsis thaliana leaves. Plant Cell Physiol. 2008, 31, 1688–1700. [Google Scholar] [CrossRef]
- Ko, S.S.; Jhong, C.M.; Lin, Y.J.; Wei, C.Y.; Lee, J.Y.; Shih, M.C. Blue Light Mediates Chloroplast Avoidance and Enhances Photoprotection of Vanilla Orchid. Int. J. Mol. Sci. 2020, 21, 8022. [Google Scholar] [CrossRef] [PubMed]
- Campbell, D.A.; Tyystjarvi, E. Parameterization of photosystem II photoinactivation and repair. Biochim. Biophys. Acta 2012, 1817, 258–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peers, G.; Truong, T.B.; Ostendorf, E.; Busch, A.; Elrad, D.; Grossman, A.R.; Hippler, M.; Niyogi, K.K. An ancient light-harvesting protein is critical for the regulation of algal photosynthesis. Nature 2009, 462, 518–521. [Google Scholar] [CrossRef] [PubMed]
- Banks, J.M. Continuous excitation chlorophyll fluorescence parameters: A review for practitioners. Tree Physiol. 2017, 37, 1128–1136. [Google Scholar] [CrossRef] [PubMed]
- Roháček, K.; Bertrand, M.; Moreau, B.; Jacquette, B.; Caplat, C.; Morant-Manceau, A.; Schoefs, B. Relaxation of the non-photochemical chlorophyll fluorescence quenching in diatoms: Kinetics, components and mechanisms. Philos. Trans. R. Soc. 2014, 369, 20130241. [Google Scholar] [CrossRef] [Green Version]
- Roeber, V.M.; Bajaj, I.; Rohde, M.; Schmulling, T.; Cortleven, A. Light acts as a stressor and influences abiotic and biotic stress responses in plants. Plant Cell Environ. 2021, 44, 645–664. [Google Scholar] [CrossRef]
- Velez-Ramirez, A.I.; van Ieperen, W.; Vreugdenhil, D.; Millenaar, F.F. Plants under continuous light. Trends Plant Sci. 2011, 16, 310–318. [Google Scholar] [CrossRef] [Green Version]
- Velez-Ramirez, A.I.; Vreugdenhil, D.; Millenaar, F.F.; van Ieperen, W. Phytochrome A Protects Tomato Plants From Injuries Induced by Continuous Light. Front. Plant Sci. 2019, 10, 19. [Google Scholar] [CrossRef]
- Hu, M.C.; Chen, Y.H.; Huang, L.C. A sustainable vegetable supply chain using plant factories in Taiwanese markets: A Nash–Cournot model. Int. J. Prod. Econ. 2014, 152, 49–56. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1/0.5 | 2/1 | 4/2 | 8/4 | 16/8 | |
|---|---|---|---|---|---|
| Before continuous light supply (B) | 1/0.5 B | 2/1 B | 4/2 B | 8/4 B | 16/8 B |
| After continuous light supply (A) | 1/0.5 A | 2/1 A | 4/2 A | 8/4 A | 16/8 A |
| Formulae and Terms | Illustrations |
|---|---|
| F0 | Minimal recorded fluorescence intensity |
| Fm | Maximal recorded fluorescence intensity |
| Fv/Fm = (Fm − F0)/Fm | Maximum quantum efficiency of PSII photochemistry |
| φP0 = TR0/ABS | Maximum quantum yield for primary photochemistry |
| Ψ0 = ET0/TR0 | Probability that a trapped exciton moves an electron into the electro transport chain beyond QA− (at t = 0) |
| Y(II) = Fv′/Fm′ | Maximum efficiency of PSII |
| Y(NPQ) = F/Fm′ − F/Fm | Heat energy dissipated by a photoprotective mechanism |
| Y(NO) = F/Fm | Passive dissipation of heat and fluorescing energy |
| PIABS ≡ (RC/ABS)[φP0/(1 − φP0)][ψ0/(1 − ψ0)] | Performance index on an absorption basis |
| PICS ≡ (RC/CS0)[φP0/(1 − φP0)][ψ0/(1 − ψ0)] | Performance index on a cross-section basis (at t = 0) |
| ABS/RC | Absorption flux per RC (reaction center) |
| TR0/RC | Trapped energy flux per RC (at t = 0) |
| ET0/RC | Electron transport flux per RC (at t = 0) |
| DI0/CS | Dissipated energy flux per CS (cross-section) (at t = 0) |
| ET0/CS | Electron transport flux per CS (at t = 0) |
| RC/CS | Density of reaction centers |
| Treatment | Plant Height (cm) | Root Length (cm) | Overground Part (g) | Underground Part (g) | Root Shoot Ratio | ||
|---|---|---|---|---|---|---|---|
| Fresh Weight | Dry Weight | Fresh Weight | Dry Weight | ||||
| 1/0.5 B | 13.92 ± 0.15 b | 25.60 ± 0.56 b | 43.59 ±2.36 b | 2.43 ± 0.23 b | 4.61 ± 0.20 c | 0.41 ± 0.02 b | 5.92 ± 0.19 ab |
| 2/1 B | 15.87 ± 0.49 a | 31.61 ± 0.33 a | 58.61 ±1.16 a | 4.13 ± 0.11 a | 9.59 ± 0.43 a | 0.95 ± 0.06 a | 4.36 ± 0.17 b |
| 4/2 B | 14.17 ± 0.43 b | 32.23 ± 1.17 a | 45.46 ±3.88 b | 2.50 ± 0.11 b | 6.15 ± 0.05 b | 0.47 ± 0.03 b | 5.33 ± 0.23 b |
| 8/4 B | 15.83 ± 0.38 a | 17.36 ± 0.63 c | 21.31 ±2.98 c | 1.07 ± 0.15 c | 1.62 ± 0.24 d | 0.14 ± 0.06 c | 8.36 ± 2.31 a |
| 16/8 B | 15.53 ± 0.55 a | 27.58 ± 2.49 b | 46.56 ±2.85 b | 2.78 ± 0.32 b | 6.06 ± 0.63 b | 0.44 ± 0.04 b | 6.41 ± 1.16 ab |
| 1/0.5 A | 16.08 ± 0.42 d | 32.59 ± 1.89 a | 44.38 ±2.26 c | 2.68 ± 0.03 c | 4.87 ± 0.13 b | 0.56 ± 0.04 c | 4.81 ± 0.25 a |
| 2/1 A | 18.15 ± 0.23 a | 34.14 ± 0.79 a | 77.81 ±1.67 a | 5.41 ± 0.16 a | 9.62 ± 1.07 a | 1.01 ± 0.12 a | 5.43 ± 0.48 a |
| 4/2 A | 16.39 ± 0.32 cd | 33.43 ± 0.60 a | 63.13 ±2.59 b | 3.46 ± 0.22 b | 8.85 ± 0.44 a | 0.72 ± 0.09 bc | 4.88 ± 0.87 a |
| 8/4 A | 17.44 ± 0.52 ab | 31.54 ± 1.75 a | 29.24 ±1.05 d | 1.70 ± 0.14 d | 3.83 ± 0.30 b | 0.26 ± 0.04 d | 6.69 ± 1.26 a |
| 16/8 A | 17.07 ± 0.16 bc | 34.35 ± 0.65 a | 67.76 ±2.51 b | 5.61 ± 0.38 a | 9.18 ± 0.94 a | 0.92 ± 0.14 ab | 6.17 ± 0.72 a |
| Treatment | Fv/Fm | φP0 | PIABS | PICS |
|---|---|---|---|---|
| 1/0.5 B | 0.768 | 0.833 | 2.823 | 1502.909 |
| 1/0.5 A | 0.762 | 0.825 | 2.299 | 1533.408 |
| 2/1 B | 0.776 | 0.836 | 2.341 | 1055.216 |
| 2/1 A | 0.781 | 0.824 | 1.943 | 1497.348 |
| 4/2 B | 0.779 | 0.830 | 3.237 | 1333.866 |
| 4/2 A | 0.795 | 0.833 | 2.112 | 1472.879 |
| 8/4 B | 0.764 | 0.815 | 1.908 | 828.164 |
| 8/4 A | 0.791 | 0.829 | 1.270 | 1032.986 |
| 16/8 B | 0.771 | 0.821 | 2.186 | 438.015 |
| 16/8 A | 0.770 | 0.818 | 1.673 | 1091.067 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Cao, N.; Shi, X.; Meng, F.; Zhou, Y.; Wang, H.; Yang, Q. The Mechanisms of Different Light Supply Patterns in the Nutrient Uptake and Chlorophyll Fluorescence of Hydroponic Lettuce. Agronomy 2023, 13, 1910. https://doi.org/10.3390/agronomy13071910
Liu Y, Cao N, Shi X, Meng F, Zhou Y, Wang H, Yang Q. The Mechanisms of Different Light Supply Patterns in the Nutrient Uptake and Chlorophyll Fluorescence of Hydroponic Lettuce. Agronomy. 2023; 13(7):1910. https://doi.org/10.3390/agronomy13071910
Chicago/Turabian StyleLiu, Yanwei, Ni Cao, Xiaolan Shi, Fei Meng, Yingjie Zhou, Haidong Wang, and Qiliang Yang. 2023. "The Mechanisms of Different Light Supply Patterns in the Nutrient Uptake and Chlorophyll Fluorescence of Hydroponic Lettuce" Agronomy 13, no. 7: 1910. https://doi.org/10.3390/agronomy13071910