
Evaluation of the ECOSSE Model for Estimating Soil Respiration from Eight European Permanent Grassland Sites
 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Study Sites
2.2. CO2 Flux Measurements
2.3. The ECOSSE Model
2.4. Model Sensitivity and Attribution
2.5. ECOSSE Evaluation
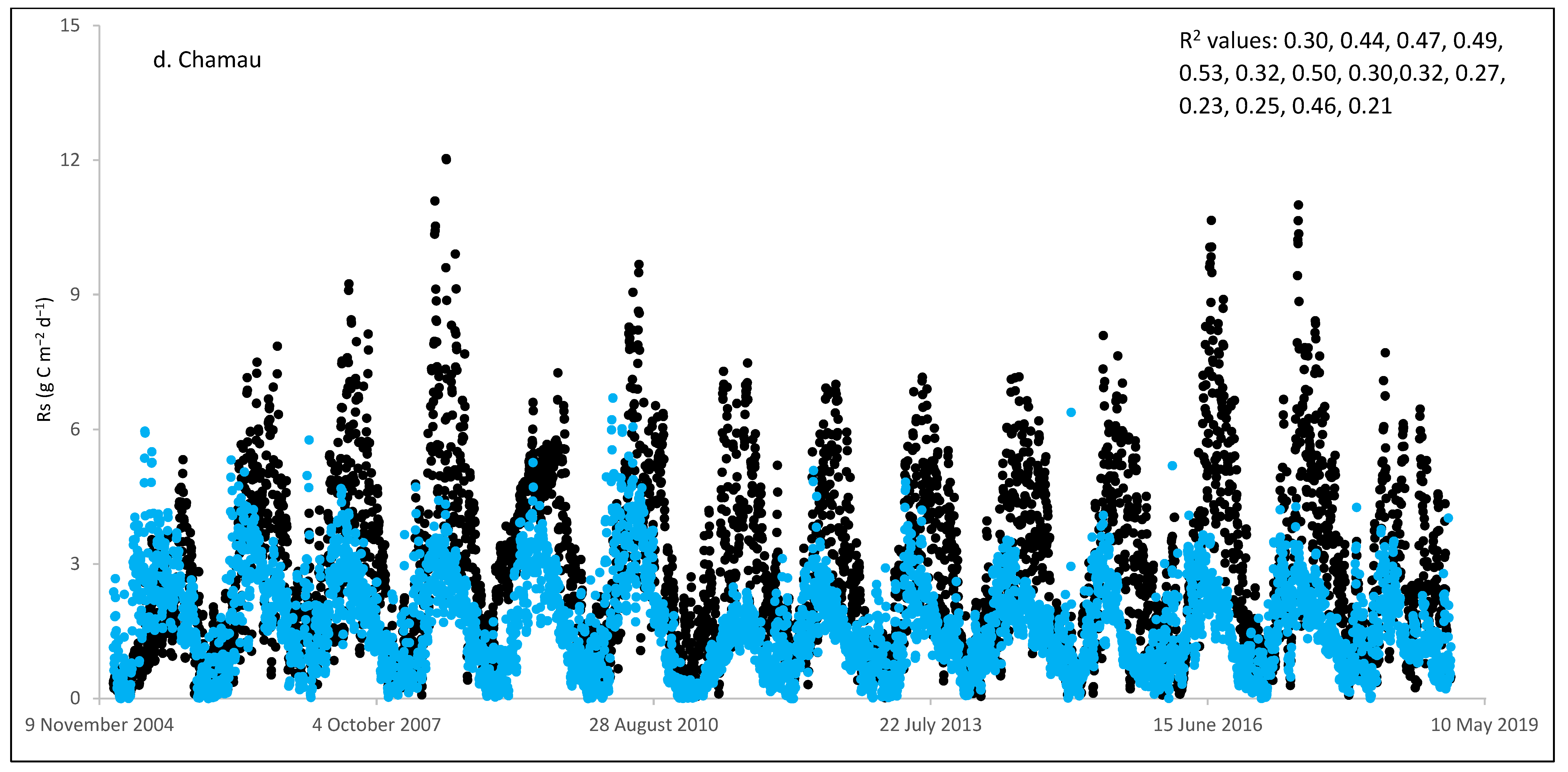
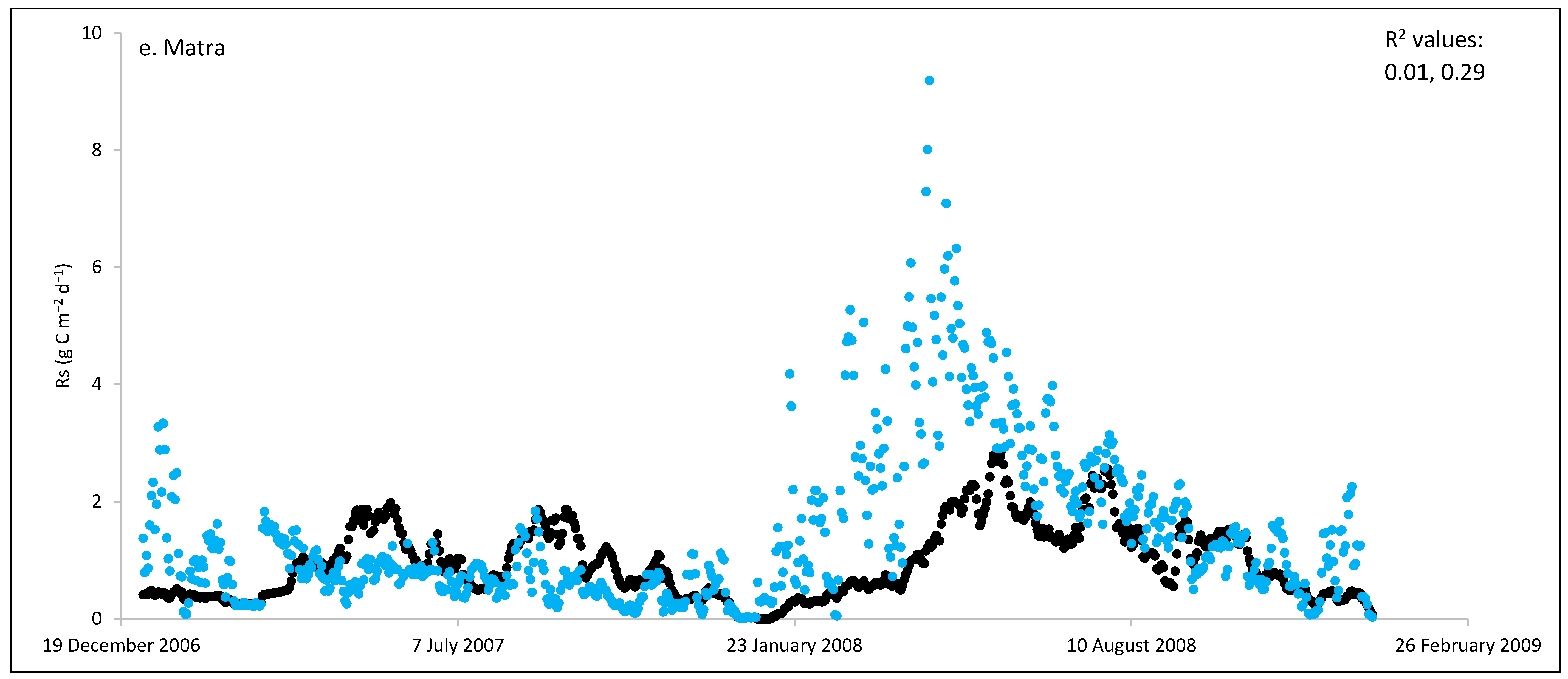
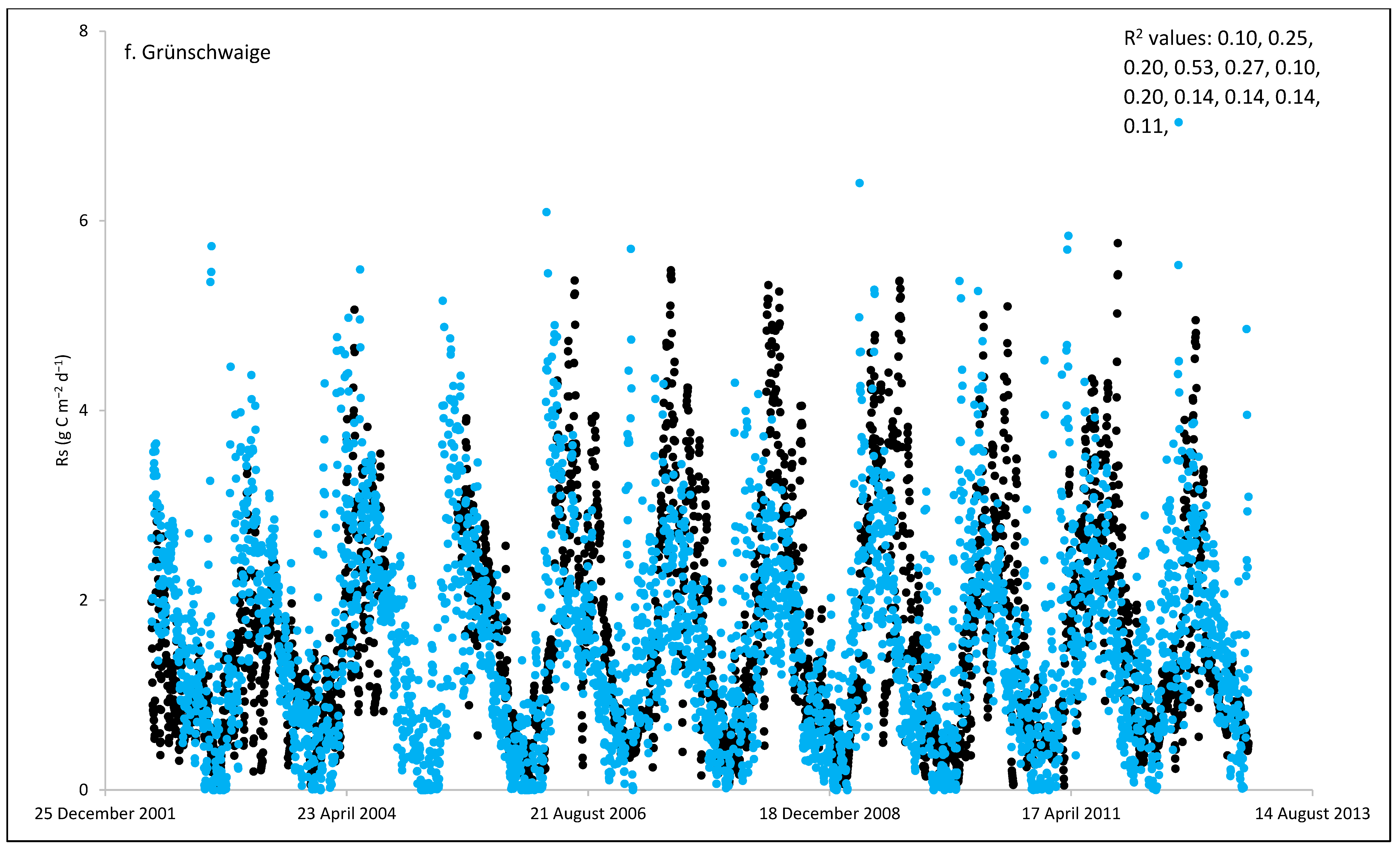
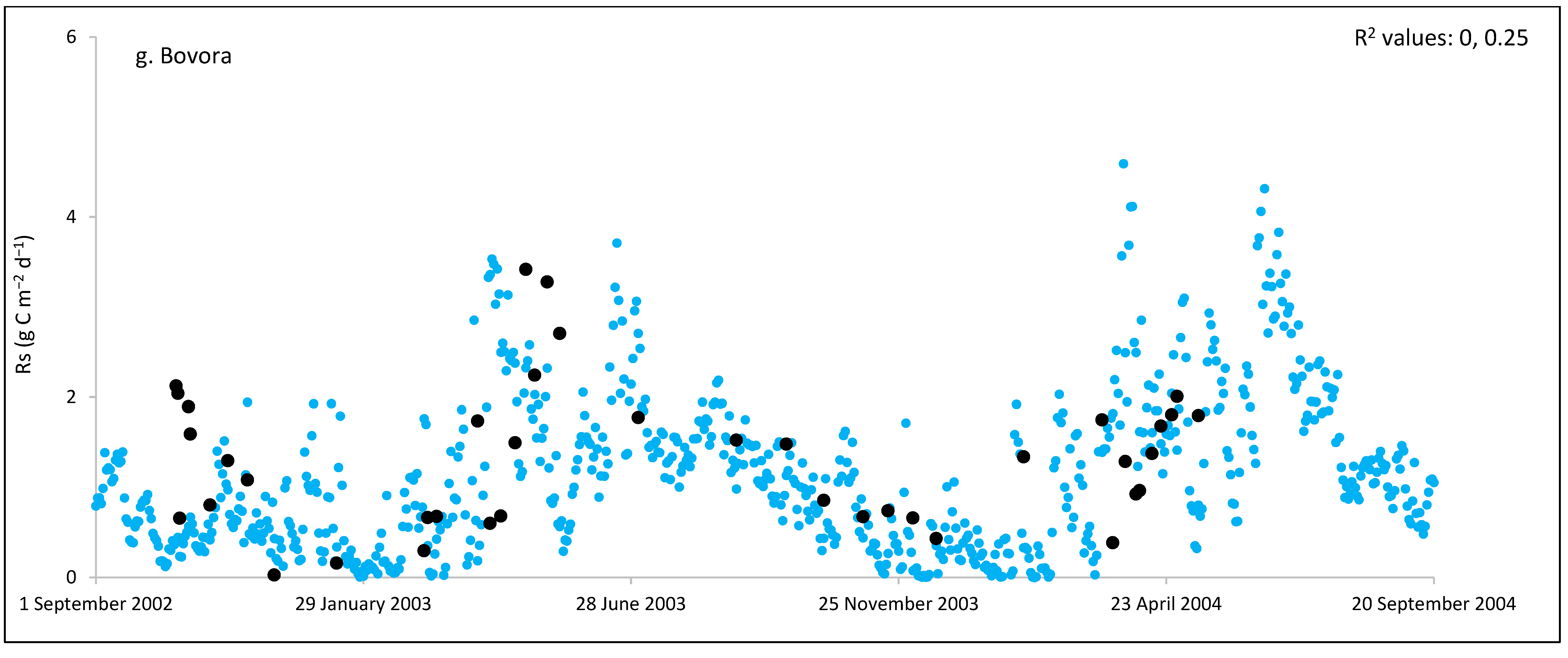
3. Results and Discussion
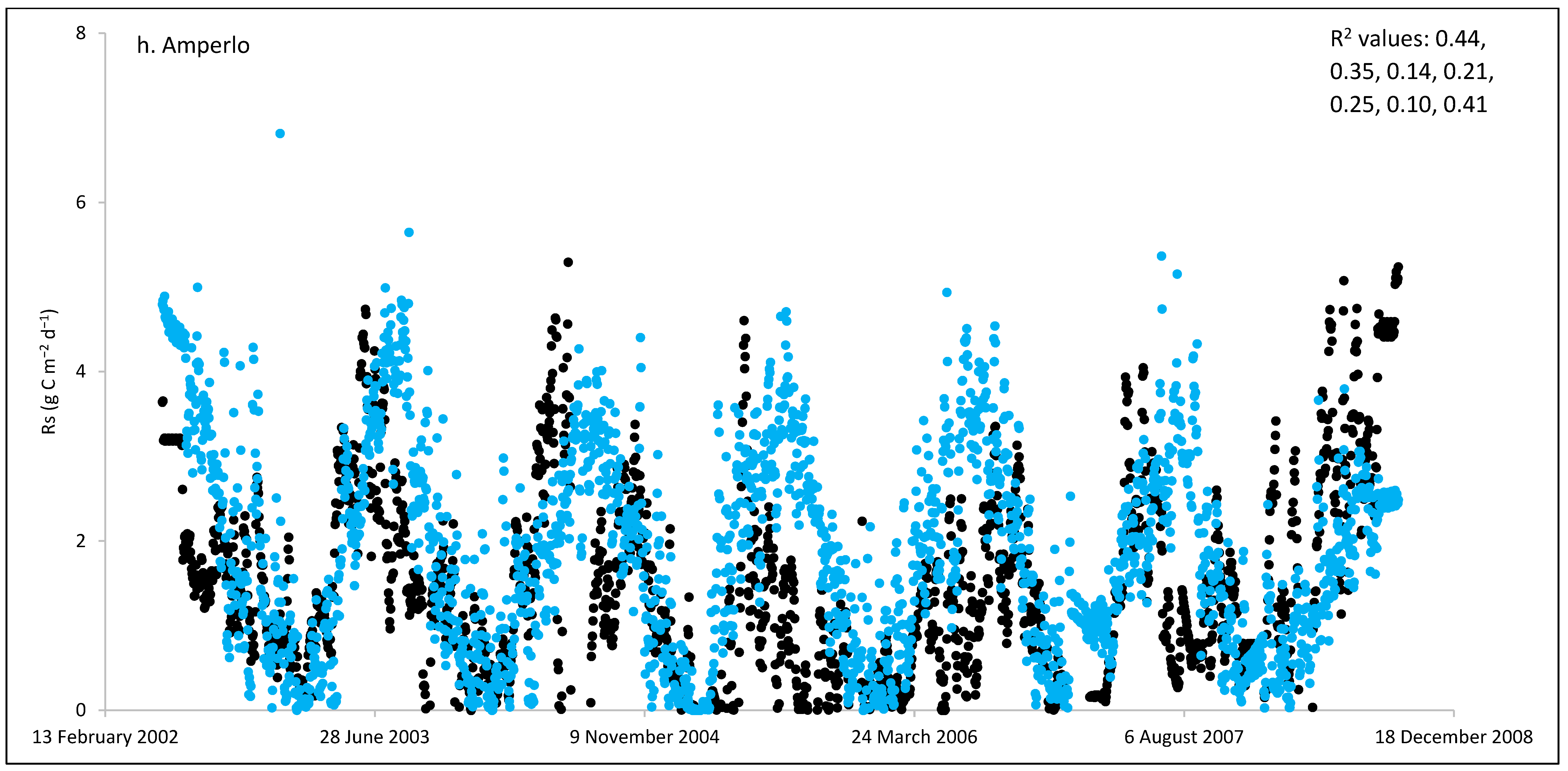
3.1. Field Estimated Soil Respiration (Rs) from Reco
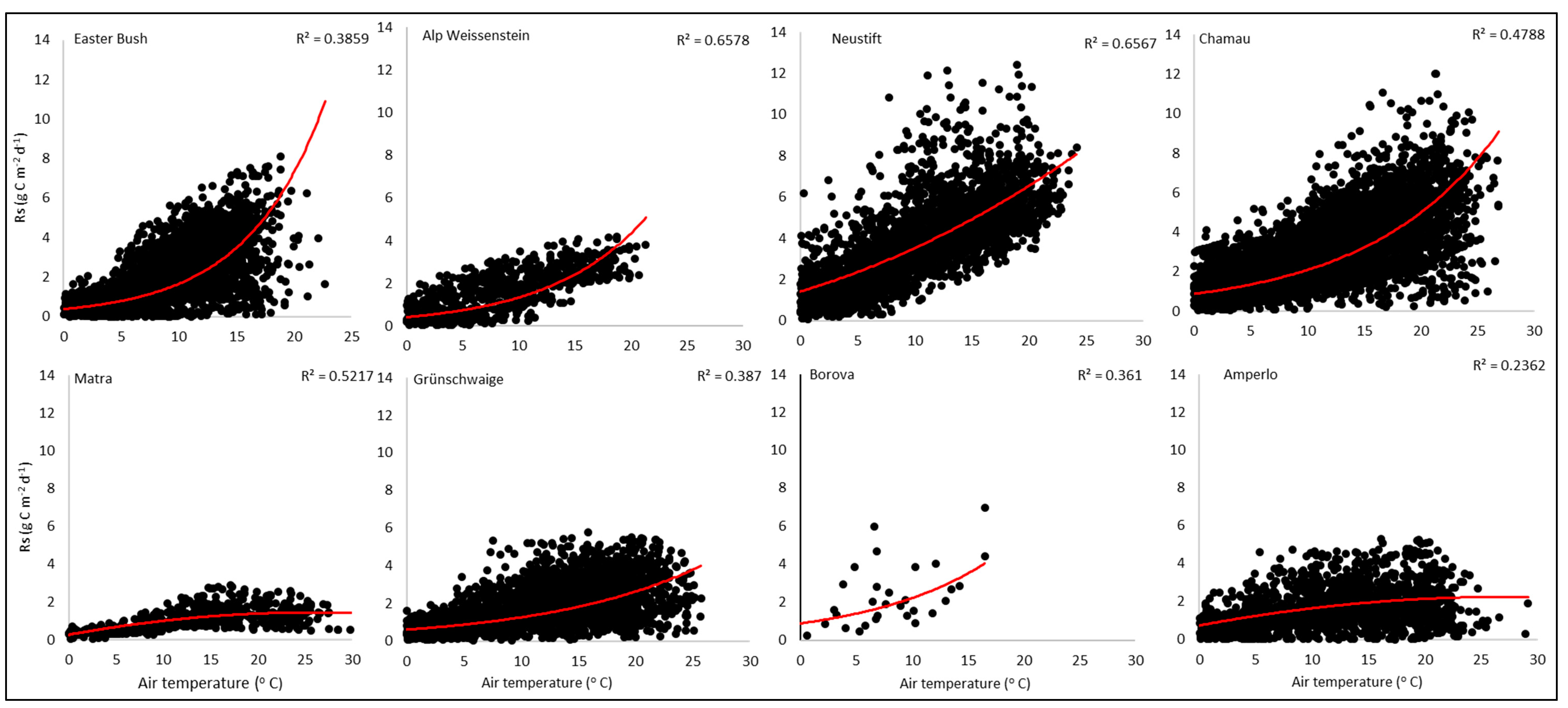
3.2. Model Sensitivity and Attribution
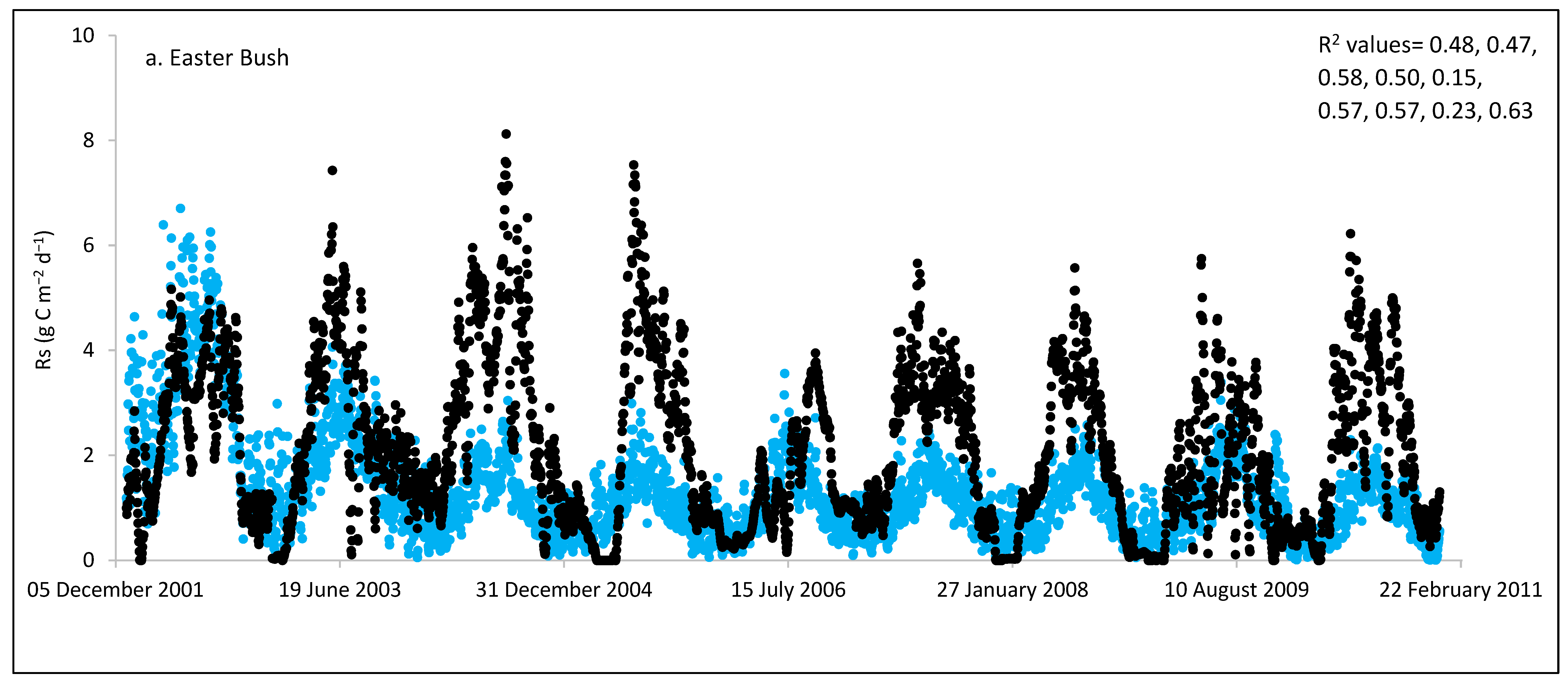
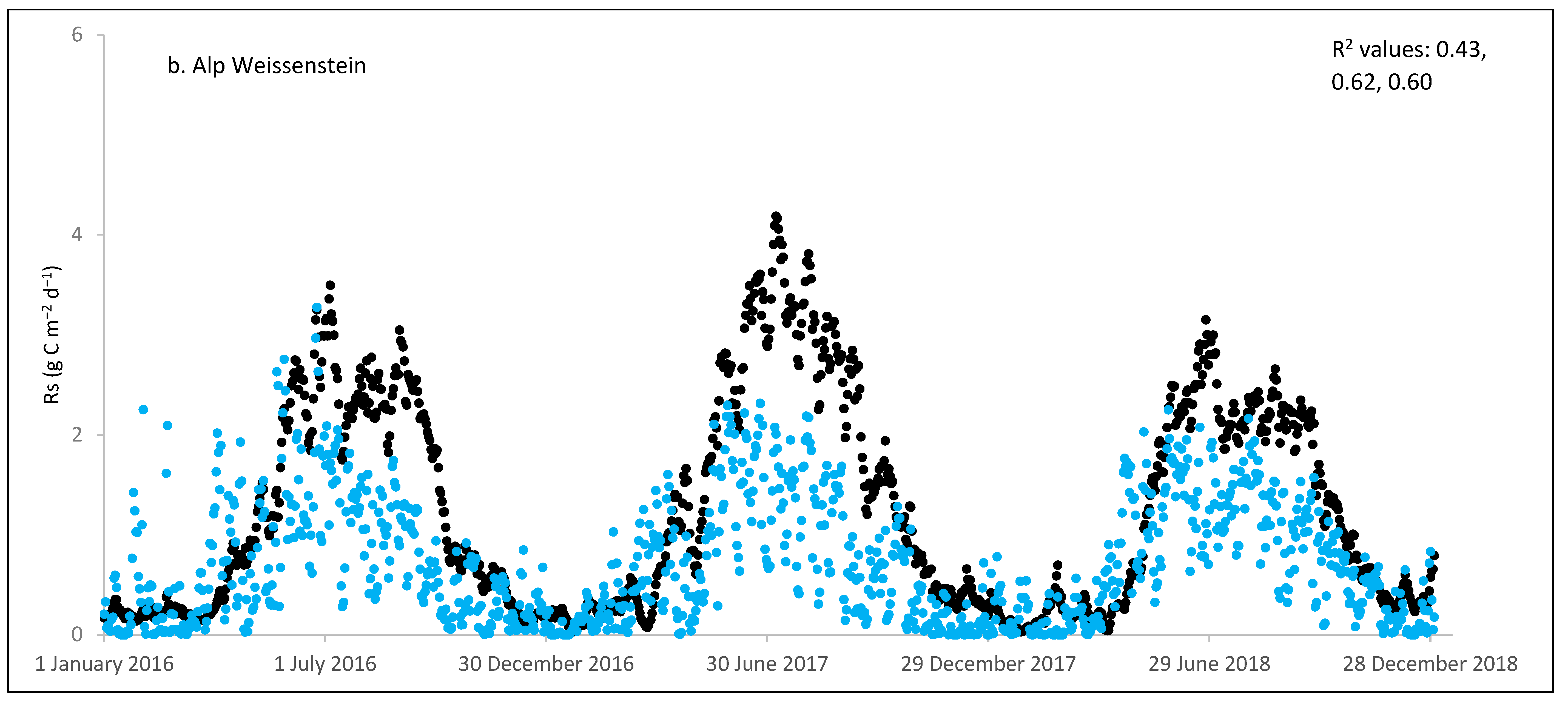
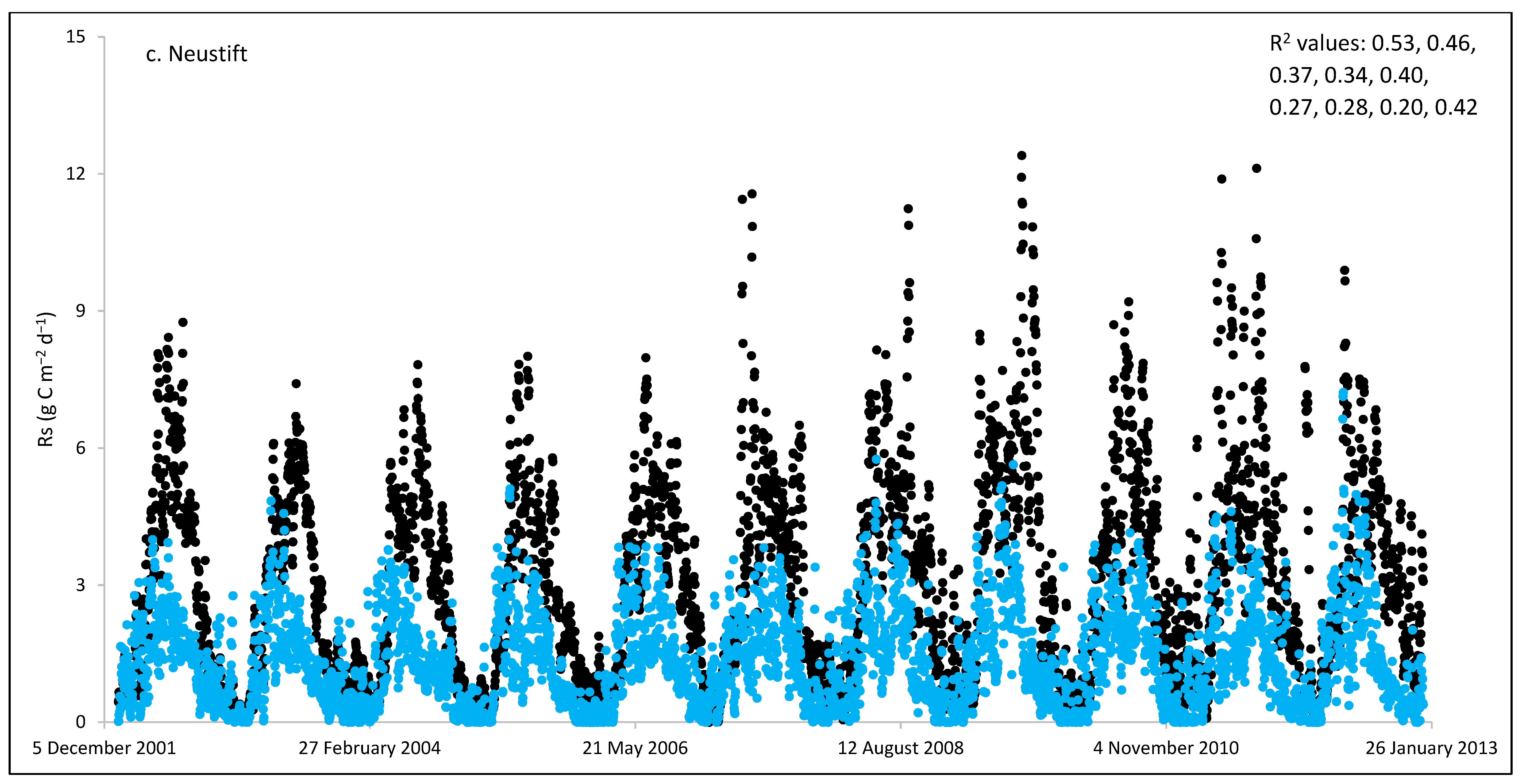
3.3. ECOSSE Simulations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2021; Available online: https://www.ipcc.ch/report/ar6/wg1/ (accessed on 9 November 2022).
- NOAA Climate. National Oceanic and Atmospheric Administration. Climate Change: Atmospheric Carbon Dioxide. 2022. Available online: https://www.climate.gov/news-features/understanding-climate/climate-change-atmospheric-carbon-dioxide#:~:text=Based%20on%20the%20annual%20report,setting%20a%20new%20record%20high (accessed on 9 November 2022).
- Caldeira, K.; Wickett, M.E. Anthropogenic carbon and ocean pH. Nature 2003, 425, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byrne, R.H.; Mecking, S.; Feely, R.A.; Liu, X. Direct observations of basin-wide acidification of the North Pacific Ocean. Geophys. Res. Lett. 2010, 37, L02601. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, G.E.; Smith, J.E.; Johnson, K.S.; Send, U.; Levin, L.A.; Micheli, F.; Paytan, A.; Price, N.N.; Peterson, B.; Takeshita, Y.; et al. High-Frequency Dynamics of Ocean pH: A Multi-Ecosystem Comparison. PLoS ONE 2011, 6, e28983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schils, R.L.; Bufe, C.; Rhymer, C.M.; Francksen, R.M.; Klaus, V.H.; Abdalla, M.; Milazzo, F.; Lellei-Kovács, E.; ten Berge, H.; Bertora, C.; et al. Permanent grasslands in Europe: Land use change and intensification decrease their multifunctionality. Agric. Ecosyst. Environ. 2022, 330, 107891. [Google Scholar] [CrossRef]
- European Commission. Directorate-General for Agriculture and Rural Development. Commission Publishes External Study on Future of EU Livestock (europa.eu). 2020. Available online: https://agriculture.ec.europa.eu/news/commission-publishes-external-study-future-eu-livestock-2020-10-14_en#:~:text=Once%20emissions%20related%20to%20the,of%20total%20agricultural%20GHG%20emissions (accessed on 9 November 2022).
- Soussana, J.-F.; Tallec, T.; Blanfort, V. Mitigating the greenhouse gas balance of ruminant production systems through carbon sequestration in grasslands. Int. J. Anim. Biosci. 2010, 4, 334–350. [Google Scholar] [CrossRef] [Green Version]
- Van Den Pol, A.; Becker, T.; Botana Fernandez, A.; Hennessy, T.; Peratoner, G. Social and economic impacts of grass based ruminant production. In Proceedings of the 27th European Grassland Federation General Meeting Cork (EGF 2018), Cork, Ireland, 17–21 June 2018; Horan, B., Hennessy, D., O’Donovan, M., Kennedy, E., McCarthy, B., Finn, J.A., O’Brien, B., Eds.; European Grassland Federation EGF: Zürich, Switzerland, 2018; pp. 697–708. [Google Scholar]
- Hussain, R.I.; Walcher, R.; Eder, R.; Allex, B.; Wallner, P.; Hutter, H.-P.; Bauer, N.; Arnberger, A.; Zaller, J.G.; Frank, T. Management of mountainous meadows associated with biodiversity attributes, perceived health benefits and cultural ecosystem services. Sci. Rep. 2019, 9, 14977. [Google Scholar] [CrossRef] [Green Version]
- Eurostat. Share of Main Land Types in Utilised Agricultural Area (UAA) by NUTS 2 Regions. 2020. Available online: https://ec.europa.eu/eurostat/statistics-explained/SEPDF/cache/73319.pdf (accessed on 9 November 2022).
- Eurostat. Eurostat Regional Yearbook 2013. 2013. Available online: https://ec.europa.eu/eurostat/web/products-statistical-books/-/ks-ha-13-001 (accessed on 9 November 2022).
- Follett, R.F.; Reed, D.A. Soil Carbon Sequestration in Grazing Lands: Societal Benefits and Policy Implications. Rangel. Ecol. Manag. 2010, 63, 4–15. [Google Scholar] [CrossRef] [Green Version]
- Conant, R.T.; Cerri, C.E.P.; Osborne, B.B.; Paustian, K. Grassland management impacts on soil carbon stocks: A new synthesis. Ecol. Appl. 2017, 27, 662–668. [Google Scholar] [CrossRef] [Green Version]
- Abdalla, M.; Espenberg, M.; Zavattaro, L.; Lellei-Kovacs, E.; Mander, U.; Smith, K.; Thorman, R.; Damatirca, C.; Schils, R.; Ten-Berge, H.; et al. Does liming grasslands increase biomass productivity without causing detrimental impacts on net greenhouse gas emissions? Environ. Pollut. 2022, 300, 118999. [Google Scholar] [CrossRef]
- Folger, P. The Carbon Cycle: Implications for Climate Change and Congress. CRS Report for Congress. 2009. Available online: www.fas.org/sgp/crs/misc/RL34059.pdf (accessed on 9 November 2022).
- Bernhardt, E.S.; Barber, J.J.; Pippen, J.S.; Taneva, L.; Andrews, J.A.; Schlesinger, W.H. Long-term effects of Free Air CO2 Enrichment (FACE) on soil respiration. Biogeochemistry 2006, 77, 91–116. [Google Scholar] [CrossRef]
- Wang, W.; Fang, J. Soil respiration and human effects on global grasslands. Glob. Planet. Change 2009, 67, 20–28. [Google Scholar] [CrossRef]
- Abdalla, M.A.; Hastings, M.; Bell, J.; Smith, J.U.; Richards, M.; Nilsson, M.B.; Peichl, M.; Löfvenius, M.O.; Lund, M.; Helfter, C.; et al. Simulation of CO2 and attribution analysis at six European peatland sites using the ECOSSE model. Wat Air Soil Pollut. 2014, 225, 2182. [Google Scholar] [CrossRef]
- Meyer, N.; Welp, G.; Amelung, W. The temperature sensitivity (Q10) of soil respiration: Controlling factors and spatial prediction at regional scale based on environmental soil classes. Glob. Biogeochem. Cycles 2018, 32, 306–323. [Google Scholar] [CrossRef]
- Dacal, M.; Delgado-Baquerizo, M.; Barquero, J.; Berhe, A.A.; Gallardo, A.; Maestre, F.T.; Garcı’a-Palacios, P. Temperature Increases Soil Respiration Across Ecosystem Types and Soil Development, But Soil Properties Determine the Magnitude of This Effect. Ecosystems 2022, 25, 184–198. [Google Scholar] [CrossRef]
- Smith, S.D.; Huxman, T.E.; Zitzer, S.E.; Charlet, T.N.; Housman, D.C.; Coleman, S.; Fenstermaker, L.K.; Seeman, J.R.; Nowak, R.S. Elevated CO2 increases productivity and invasive species success in an arid ecosystem. Nature 2000, 408, 79–82. [Google Scholar] [CrossRef]
- Abdalla, M.; Kumar, S.; Jones, M.; Burke, J.; Williams, M. Testing DNDC model for simulating soil respiration and assessing the effects of climate change on the CO2 gas flux from Irish agriculture. Glob. Planet. Change 2011, 78, 106–115. [Google Scholar] [CrossRef]
- Zak, D.R.; Pregitzer, K.S.; King, J.S.; Holmes, W.E. Elevated atmospheric CO2, fine roots and the response of soil microorganisms: A review and hypothesis. New Phytol. 2000, 147, 201–222. [Google Scholar] [CrossRef] [Green Version]
- Jenkinson, D.S.; Rayner, J.H. The turnover of soil organic matter in some of the Rothamsted classical experiments. Soil Sci. 1977, 123, 298–305. [Google Scholar] [CrossRef]
- Jenkinson, D.S.; Hart, P.B.S.; Rayner, J.H.; Parry, L.C. Modelling the turnover of organic matter in long-term experiments at Rothamsted. Intecol Bull. 1987, 15, 1–8. [Google Scholar]
- Coleman, K.; Jenkinson, D.S. RothC-26.3: A model for the turnover of carbon in soil. In Evaluation of Soil Organic Matter Models Using Existing, Long-Term Datasets, NATO ASI Series I; Powlson, D.S., Smith, P., Smith, J.U., Eds.; Springer: Berlin/Heidelberg, Germany, 1996; Volume 38, pp. 237–246. [Google Scholar]
- Bradbury, N.J.; Whitmore, A.P.; Hart, P.B.S.; Jenkinson, D.S. Modelling the fate of nitrogen in crop and soil in the years following the application of 15N-labelled fertilizer to winter wheat. J. Agric. Sci. 1993, 121, 363–379. [Google Scholar] [CrossRef]
- Smith, J.U.; Glendining, M.J. A decision support system for optimising the use of nitrogen in crop rotations. Rotations and cropping systems. Asp. Appl. Biol. 1996, 47, 103–110. [Google Scholar]
- Smith, J.; Gottschalk, P.; Bellarby, J.; Richards, M.; Nayak, D.; Coleman, K.; Hillier, J.; Wattenbach, M.; Aitkenhead, M.; Yeluripurti, J.; et al. Model to Estimate Carbon in Organic Soils-Sequestration and Emissions (ECOSSE) User-Manual; University of Aberdeen: Aberdeen, UK, 2010; pp. 1–76. [Google Scholar]
- Reth, S.; Riechstein, M.; Falge, E. The effect of soil water content, soil temperature, soil pH-value and the root mass on soil CO2 efflux–A modified model. Plant Soil 2005, 268, 21–33. [Google Scholar] [CrossRef]
- McMillen, R.T. An eddy correlation technique with extended applicability to non-simple terrain. Bound Layer Meteorol. 1988, 43, 231–245. [Google Scholar] [CrossRef]
- Aubinet, M.; Vesala, T.; Papale, D. Eddy Covariance: A Practical Guide to Measurement and Data Analysis; Spring Atmospheric Science: London, UK, 2012; ISSN 2194-5217. [Google Scholar]
- Jones, S.K.; Helfter, C.; Anderson, M.; Coyle, M.; Campbell, C.; Famulari, D.; Di Marco, C.; van Dijk, N.; Topp, C.F.E.; Kiese, R.; et al. The nitrogen, carbon and greenhouse gas budget of a grazed, cut and fertilised temperate grassland. Biogeosciences 2017, 14, 2069–2088. [Google Scholar] [CrossRef] [Green Version]
- Hiller, R.M.; Zeeman, J.; Eugster, W. Eddy-covariance flux measurements in the complex terrain of an alpine valley in Switzerland. Bound. Layer Meteorol. 2008, 127, 449–467. [Google Scholar] [CrossRef] [Green Version]
- Wohlfahrt, G.; Anderson-Dunn, M.; Bahn, M.; Balzarolo, M.; Berninger, F.; Campbell, C.; Carrara, A.; Cescatti, A.; Christensen, T.; Dore, S.; et al. Biotic, abiotic and management controls on the net ecosystem CO2 exchange of European mountain grasslands. Ecosystems 2008, 11, 1338–1351. [Google Scholar] [CrossRef]
- Fuchs, K.; Hörtnagl, L.; Buchmann, N.; Eugster, W.; Snow, V.; Merbold, L. Management matters: Testing a mitigation strategy for nitrous oxide emissions using legumes on intensively managed grassland. Biogeosciences 2018, 15, 5519–5543. [Google Scholar] [CrossRef] [Green Version]
- Pintér, K.; Barcza, Z.; Balogh, J.; Czóbel Sz Csintalan Zs Tuba, Z.; Zoltán, N. Interannual variability of grasslands’ carbon balance depends on soil type. Community Ecol. 2008, 9 (Suppl. S1), 43–48. [Google Scholar] [CrossRef]
- Hirl, R.T.; Schnyder, H.; Ostler, U.; Schäufele, R.; Schleip, I.; Vetter, S.; Auerswald, K.; Baca Cabrera, J.C.; Wingate, L.; Barbour, M.M.; et al. The 18O ecohydrology of a grassland ecosystem—Predictions and observations. Hydrol. Earth Syst. Sci. 2019, 23, 2581–2600. [Google Scholar] [CrossRef] [Green Version]
- Cuhel, J.; šimek, M.; Laughlin, R.J.; Bru, D.; Chèneby, D.; Watson, C.J.; Philippot, L. Insights into the effect of soil pH on N2O and N2 emissions and denitrifier community size and activity. Appl. Environ. Microbiol. 2010, 76, 870–1878. [Google Scholar] [CrossRef] [Green Version]
- Balzarolo, M.; Balzarolo, M.; Boussetta, S.; Balsamo, G.; Beljaars, A.; Maignan, F.; Calvet, J.-C.; Lafont, S.; Barbu, A.; Poulter, B.; et al. Evaluating the potential of large-scale simulations to predict carbon fluxes of terrestrial ecosystems over a European Eddy Covariance network. Biogeosciences 2014, 11, 266–2678. [Google Scholar] [CrossRef] [Green Version]
- Hardie, S.M.L.; Garnett, M.H.; Fallick, A.E.; Ostle, N.J.; Rowland, A.P. Bomb 14C analysis of ecosystem respiration reveals that peatland vegetation facilitates release of old carbon. Geoderma 2009, 153, 393–401. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; McGechan, M.B. A review of carbon and nitrogen processes in four soil nitrogen dynamics models. J. Agric. Eng. Res. 1998, 69, 279–305. [Google Scholar] [CrossRef]
- Thornthwaite, C.W. An approach toward a rational classification of climate. Geogr. Rev. 1948, 38, 55–94. [Google Scholar] [CrossRef]
- Smith, J.; Smith, P. Environmental Modelling: An Introduction; Oxford University Press: Oxford, UK, 2007; pp. 1–178. [Google Scholar]
- Smith, P.; Smith, J.U.; Powlson, D.S.; McGill, W.B.; Arah, J.R.M.; Chertov, O.G.; Coleman, K.; Franko, U.; Frolking, S.; Jenkinson, D.S.; et al. A comparison of the performance of nine soil organic matter models using seven long-term experimental datasets. Geoderma 2007, 81, 153–225. [Google Scholar] [CrossRef]
- Cai, Z.; Swamoto, T.; Li, C.; Kang, G.; Boonjawat, J.; Mosier, A.; Wassmann, R.; Tsuruta, H. Field validation of the DNDC-model for greenhouse gas emissions in East Asian cropping systems. Glob. Biogeochem. Cycles 2003, 17, 1107. [Google Scholar] [CrossRef] [Green Version]
- Bahn, M.; Rodeghiero, M.; Anderson-Dunn, M.; Dore, S.; Gimeno, C.; Drösler, M.; Williams, M.; Ammann, C.; Berninger, F.; Flechard, C.; et al. Soil Respiration in European Grasslands in Relation to Climate and Assimilate Supply. Ecosystems 2008, 11, 1352–1367. [Google Scholar] [CrossRef] [Green Version]
- Schär, C.; Vidale, P.; Lüthi, D.; Frei, C.; Häberli, C.; Liniger, M.; Appenzeller, C. The role of increasing temperature variability in European summer heatwaves. Nature 2004, 427, 332–336. [Google Scholar] [CrossRef]
- Li, J.; Pei, J.; Pendall, E.; Fang, C. Ming Ni Spatial heterogeneity of temperature sensitivity of soil respiration: A global analysis of field observations. Soil Biol. Biochem. 2020, 141, 107675. [Google Scholar] [CrossRef]
- Carbone, M.S.; Winston, G.C.; Trumbore, S.E. Soil respiration in perennial grass and shrub ecosystems: Linking environmental controls with plant and microbial sources on seasonal and diel timescales. J. Geophys. Res. 2008, 113, G02022. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.Z.; Wan, S.Q.; Su, B.; Hui, D.F.; Luo, Y.Q. Response of soil CO2 efflux to water manipulation in a tallgrass prairie ecosystem. Plant Soil 2002, 240, 213–223. [Google Scholar] [CrossRef]
- Xu, L.K.; Baldocchi, D.D.; Tang, J.W. How soil moisture, rain pulses, and growth alter the response of ecosystem respiration to temperature. Glob. Biogeochem. Cycles 2004, 18, GB4002. [Google Scholar] [CrossRef]
- Frank, A.B.; Liebig, M.A.; Hanson, J.D. Soil carbon dioxide fluxes in northern semiarid grasslands. Soil Biol. Biochem. 2002, 34, 1235–1241. [Google Scholar] [CrossRef]
- Hamza, M.A.; Anderson, W.K. Soil compaction in cropping systems: A review of the nature, causes and possible solutions. Soil Tillage Res. 2005, 82, 121–145. [Google Scholar] [CrossRef]
- Solomon, S.; Qin, D.; Manning, M.; Chen, Z.; Marquis, M.; Averyt, K.B.; Tignor, M.; Miller, H.L. IPCC Fourth Assessment Report: Climate Change 2007, Cambridge Univ Press, Cambridge, UK, Climate Change 2007: Working Group I: The Physical Science Basis. 2008. Available online: https://www.ipcc.ch/site/assets/uploads/2018/02/ar4-wg1-frontmatter-1.pdf (accessed on 9 November 2022).
- Cantarel, A.A.M.; Bloor, J.M.G.; Pommier, T.; Guillaumaud, N.; Moirot, C.; Soussana, J.-F.; Poly, F. Four years of experimental climate change modifies the microbial drivers of N2O fluxes in an upland grassland ecosystem. Glob. Change Biol. 2012, 18, 2520–2531. [Google Scholar] [CrossRef]
- Wieder, W.R.; Bonan, G.B.; Allison, S.D. Global soil carbon projections are improved by modelling microbial processes. Nat. Clim. Change 2013, 3, 909–912. [Google Scholar] [CrossRef] [Green Version]
- De Goede, R.G.M.; Brussaard, L.; Akkermans, A.D.L. On-farm impact of cattle slurry manure management on biological soil quality. NJAS Wagening. J. Life Sci. 2003, 51, 103–133. [Google Scholar] [CrossRef] [Green Version]
- Asmuß, T.; Bechtold, M.; Tiemeyer, B. On the Potential of Sentinel-1 for High Resolution Monitoring of Water Table Dynamics in Grasslands on Organic Soils. Remote Sens. 2019, 11, 1659. [Google Scholar] [CrossRef] [Green Version]
- Clair, S.B.S.T.; Sudderth, E.A.; Fischer, M.L.; Torn, M.S.; Stuart, S.A.; Salve, R.; Eggett, D.L.; Ackerly, D.D. Soil drying and nitrogen availability modulate carbon and water exchange over a range of annual precipitation totals and grassland vegetation types. Glob. Change Biol. 2009, 15, 3018–3030. [Google Scholar] [CrossRef]
- Davidson, E.A.; Belk, E.; Boone, R.D. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Glob. Change Biol. 1998, 4, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Shaver, G.R.; Canadell, J.; Chapin, F.S.I.I.I.; Gurevitch, J.; Harte, J.; Henry, G.; Ineson, P.; Jonasson, S.; Melillo, J.; Pitelka, L.; et al. Global warming and terrestrial ecosystems: A conceptual framework for analysis. Bioscience 2000, 50, 871–882. [Google Scholar] [CrossRef]
- Shaw, M.R.; Harte, J. Control of litter decomposition in a subalpine meadow-sagebrush steppe ecotone under climate change. Ecol. Appl. 2001, 11, 1206–1223. [Google Scholar]
- Chen, D.; Li, J.; Lan, Z.; Hu, S.; Bai, Y. Soil acidification exerts a greater control on soil respiration than soil nitrogen availability in grasslands subjected to long-term nitrogen enrichment. Funct. Ecol. 2016, 30, 658–669. [Google Scholar] [CrossRef]
- Apostolakis, A.; Schöning, I.; Michalzik, B.; Klaus, V.H.; Boeddinghaus, R.S.; Kandeler, E.; Marhan, S.; Bolliger, R.; Fischer, M.; Prati, D.; et al. Drivers of soil respiration across a management intensity gradient in temperate grasslands under drought. Nutr. Cycl. Agroecosyst. 2022, 124, 101–116. [Google Scholar] [CrossRef]
- Sankaran, M.; Augustine, D.J. Large herbivores suppress decomposer abundance in a semiarid grazing ecosystem. Ecology 2004, 85, 1052–1061. [Google Scholar] [CrossRef] [Green Version]
- Barneze, A.S.; Abdalla, M.; Whitaker, J.; McNamara, N.P.; Ostle, N.J. Predicted Soil Greenhouse Gas Emissions from Climate X Management Interactions in Temperate Grassland. Agronomy 2022, 12, 3055. [Google Scholar] [CrossRef]
- Scharlemann, J.P.W.; Tanner, E.V.J.; Hiederer, R.; Kapos, V. Global soil carbon: Understanding and managing the largest terrestrial carbon pool. Carbon Manag. 2014, 5, 81–91. [Google Scholar] [CrossRef]
- Moinet, G.Y.K.; Midwood, A.J.; Hunt, J.E.; Rumpel, C.; Millard, P.; Chabbi, A. Grassland Management Influences the Response of Soil Respiration to Drought. Agronomy 2019, 9, 124. [Google Scholar] [CrossRef] [Green Version]
- Sándor, R.; Ehrhardt, F.; Brilli, L.; Carozzi, M.; Recous, S.; Smith, P.; Snow, V.; Soussana, J.-F.; Dorich, C.D.; Fuchs, K.; et al. The use of biogeochemical models to evaluate mitigation of greenhouse gas emissions from managed grasslands. Sci. Total Environ. 2018, 642, 292–306. [Google Scholar] [CrossRef] [Green Version]
- Subke, J.A.; Bahn, M. On the ‘temperature sensitivity’ of soil respiration: Can we use the immeasurable to predict the unknown? Soil Biol. Biochem. 2010, 42, 1653–1656. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site/Location * | Coordinates | MAT (°C) | MAP (mm) | Type of Grassland | Site Management | Soil Type | Investigated Years |
|---|---|---|---|---|---|---|---|
| Easter Bush/UK | 55°52′ N, 03°02′ W | 9.0 | 947 | Lolium perenne/clover | Fertilised, grazed, cut | Sandy clay loam | 2002–2010 |
| Alp Weissenstein/CH | 46°34′ N, 09°47′ E | 4.0 | 918 | Mixed grasslands | Extensively grazed | Sandy loam | 2016–2018 |
| Neustift/AT | 47°12′ N, 11°32′ E | 6.5 | 852 | Mixed grasslands | Grazed, cut, manure | Sandy loam | 2002–2012 |
| Chamau/CH | 47°12′ N, 08°24′ E | 9.1 | 1151 | Ryegrass/clover | Cut, grazed, fertilised | Sandy loam to silty loam | 2005–2018 |
| Matra/HU | 47°52′ N, 19°56′ E | 10.2 | 622 | Mixed grasslands | Cut | Clay soil | 2007–2008 |
| Grünschwaige/DE | 48°23′ N, 11°50′ E | 9.0 | 775 | Ryegrass/clover | Grazed, manure | Silt loam | 2002–2012 |
| Borova/CZ | 48°52′ N, 14°13′ E | 7.0 | 650 | Ryegrass/clover | Extensively grazed, fertilised | Sandy loam | 2002–2004 |
| Amperlo/IT | 41°90′ N, 13°22′ E | 10.0 | 1365 | Mixed grasslands | Grazed | Clay soil | 2002–2008 |
| Site/Location | Bulk Density (g cm−3) | Soil pH | Clay (%) | Water Table Depth (cm) | Measured/Estimated SOC * (t ha−1) |
|---|---|---|---|---|---|
| Easter Bush/UK | 1.3 | 5.1 | 23 | 85 | 120 |
| Alp Weissenstein/CH | 1.1 | 6.4 | 20 | 125 | 32.5 |
| Neustift/AT | 0.7 | 5.5 | 5.5 | 125 | 43.9 |
| Chamau/CH | 1.1 | 5.0 | 20 | 125 | 36.7 |
| Matra/HU | 1.3 | 5.4 | 35 | 400 | 19.5 |
| Grünschwaige/DE | 1.1 | 6.4 | 26 | 125 | 31.3 |
| Borova/CZ | 1.1 | 7.8 | 6.0 | 25 | 27.9 |
| Amperlo/IT | 1.1 | 6.6 | 56 | 125 | 54.2 |
| Site/Location | Years of Measurements | Cumulative Annual Estimated Rs (g C m−2) | Cumulative Annual Simulated Rs (g C m−2) | Mean Daily Estimated Rs (g C m−2) | Mean Daily Simulated Rs (g C m−2) | RD * (%) | RMSE ▲ (g C m−2 d−1) | d | R2-Range ¶ |
|---|---|---|---|---|---|---|---|---|---|
| Easter Bush/UK | 9 | 770 | 490 | 2.11 | 1.35 | −36 | 1.56 | 0.86 | 0.15–0.63 |
| Alp Weissenstein/CH | 3 | 460 | 260 | 1.25 | 0.72 | −42 | 0.91 | 0.87 | 0.43–0.62 |
| Neustift/AT | 11 | 1120 | 490 | 3.07 | 1.34 | −56 | 2.51 | 0.28 | 0.20–0.53 |
| Chamau/CH | 14 | 810 | 430 | 2.92 | 1.55 | −47 | 3.30 | 0.55 | 0.21–0.53 |
| Matra/HU | 2 | 350 | 530 | 0.95 | 1.45 | 51 | 1.28 | 0.67 | 0.01–0.29 |
| Grünschwaige/DE | 11 | 530 | 550 | 1.60 | 1.58 | 4 | 1.28 | 0.54 | 0.10–0.53 |
| Borova/CZ | 3 | 483 | 394 | 1.32 | 1.08 | −18 | 1.05 | 0.87 | 0.0–0.25 ▼ |
| Amperlo/IT | 7 | 450 | 560 | 1.55 | 1.95 | 24 | 1.35 | 0.55 | 0.14–0.44 |
| Site Name | Management | Date (Month/Year) | Estimated Rs (% Change) | Simulated Rs (% Change) |
|---|---|---|---|---|
| Easter Bush | Grass cut | June 2002 | −21 | −10 |
| August 2002 | −12 | −13 | ||
| May 2003 | 20 | 25 | ||
| Slurry application | September 2004 | 15 | 15 | |
| April 2005 | 33 | 0 | ||
| Chamau | Grass cut | May 2005 | −36 | −32 |
| June 2005 | −44 | −31 | ||
| July 2005 | 17 | 8 | ||
| August 2005 | −10 | −14 | ||
| July 2006 | −56 | −24 | ||
| July 2006 | −2 | 2 | ||
| September 2006 | 90 | 55 | ||
| Slurry application | May 2005 | 84 | 67 | |
| June 2005 | 46 | 31 | ||
| August 2005 | 13 | 10 | ||
| December 2005 | −29 | 27 | ||
| March 2006 | 58 | >100 | ||
| May 2006 | −43 | −1 | ||
| June 2006 | >100 | 3 | ||
| July 2006 | 38 | 16 | ||
| August 2006 | −16 | −19 |
| Site/Location | Temp+ | Temp− | Rain+ | Rain− | SOC+ | SOC− | pH+ | pH− | WT+ |
|---|---|---|---|---|---|---|---|---|---|
| Easter Bush/UK | 20.1 | −23.0 | −0.6 | 0.0 | 4.3 | −0.8 | 0.4 | −2.2 | 0.0 |
| Alp Weissenstein/CH | 25.5 | −22.4 | −17.7 | 10.9 | 2.2 | −2.6 | 0.0 | −6.3 | 11.3 |
| Neustift/AT | 21.6 | −19.1 | −10.1 | 1.6 | 7.4 | −2.5 | 0.0 | −5.1 | 14.1 |
| Chamau/CH | 21.4 | −19.2 | −5.5 | 0.0 | 3.1 | −2.5 | 0.0 | −11.1 | 4.7 |
| Matra/HU | 16.0 | −11.3 | −6.0 | −5.8 | 4.5 | 1.9 | 11.2 | −2.5 | 0.0 |
| Grünschwaige/DE | 21.8 | −20.1 | −1.4 | 0.0 | 2.1 | −2.2 | 0.0 | −11.5 | 4.8 |
| Borova/CZ | 22.4 | −21.5 | −1.2 | 1.6 | 0.6 | −0.6 | 0.0 | −6.4 | 16.2 |
| Amperlo/IT | 24.4 | −22.1 | −7.2 | 5.7 | 15.6 | −2.2 | 5.7 | −15.7 | 8.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdalla, M.; Feigenwinter, I.; Richards, M.; Vetter, S.H.; Wohlfahrt, G.; Skiba, U.; Pintér, K.; Nagy, Z.; Hejduk, S.; Buchmann, N.; et al. Evaluation of the ECOSSE Model for Estimating Soil Respiration from Eight European Permanent Grassland Sites. Agronomy 2023, 13, 1734. https://doi.org/10.3390/agronomy13071734
Abdalla M, Feigenwinter I, Richards M, Vetter SH, Wohlfahrt G, Skiba U, Pintér K, Nagy Z, Hejduk S, Buchmann N, et al. Evaluation of the ECOSSE Model for Estimating Soil Respiration from Eight European Permanent Grassland Sites. Agronomy. 2023; 13(7):1734. https://doi.org/10.3390/agronomy13071734
Chicago/Turabian StyleAbdalla, Mohamed, Iris Feigenwinter, Mark Richards, Sylvia Helga Vetter, Georg Wohlfahrt, Ute Skiba, Krisztina Pintér, Zoltán Nagy, Stanislav Hejduk, Nina Buchmann, and et al. 2023. "Evaluation of the ECOSSE Model for Estimating Soil Respiration from Eight European Permanent Grassland Sites" Agronomy 13, no. 7: 1734. https://doi.org/10.3390/agronomy13071734