

Absorption, Translocation, and Metabolism of Glyphosate and Imazethapyr in Smooth Pigweed with Multiple Resistance
 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
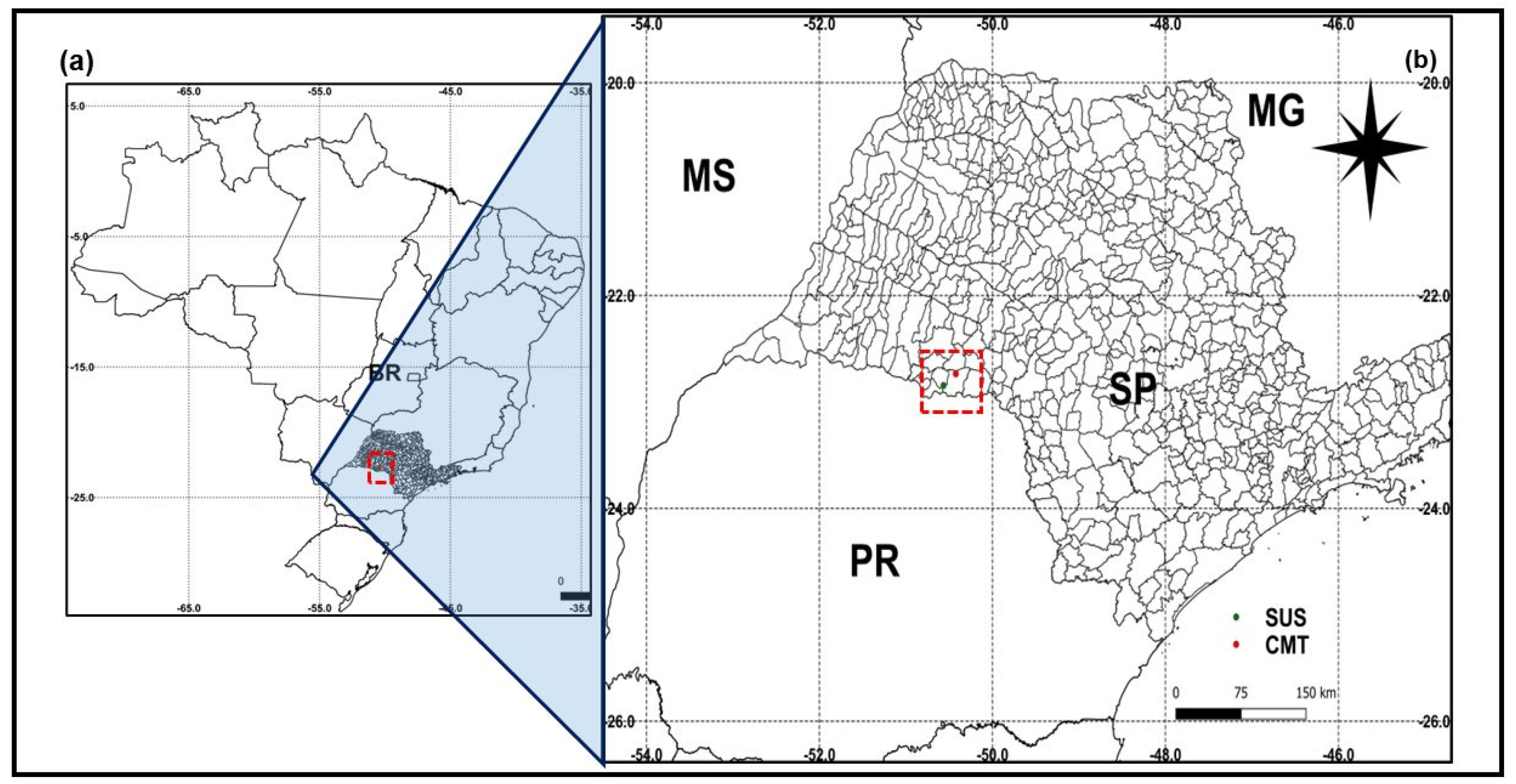
2.1. Plant Material for the Dose–Response Test
2.2. Resistant Population Confirmation
2.2.1. DNA Extraction
2.2.2. Amaranthus Species Determination
2.3. Dose–Response Assay
2.4. Absorption, Translocation, and Metabolism Study
2.4.1. Preparation of 14C-Glyphosate and 14C-Imazethapyr Solution
2.4.2. Absorption, Translocation, and Metabolism of 14C-Glyphosate and 14C-Imazethapyr
2.5. Statistical Analysis
3. Results
3.1. Amaranthus Species Determination
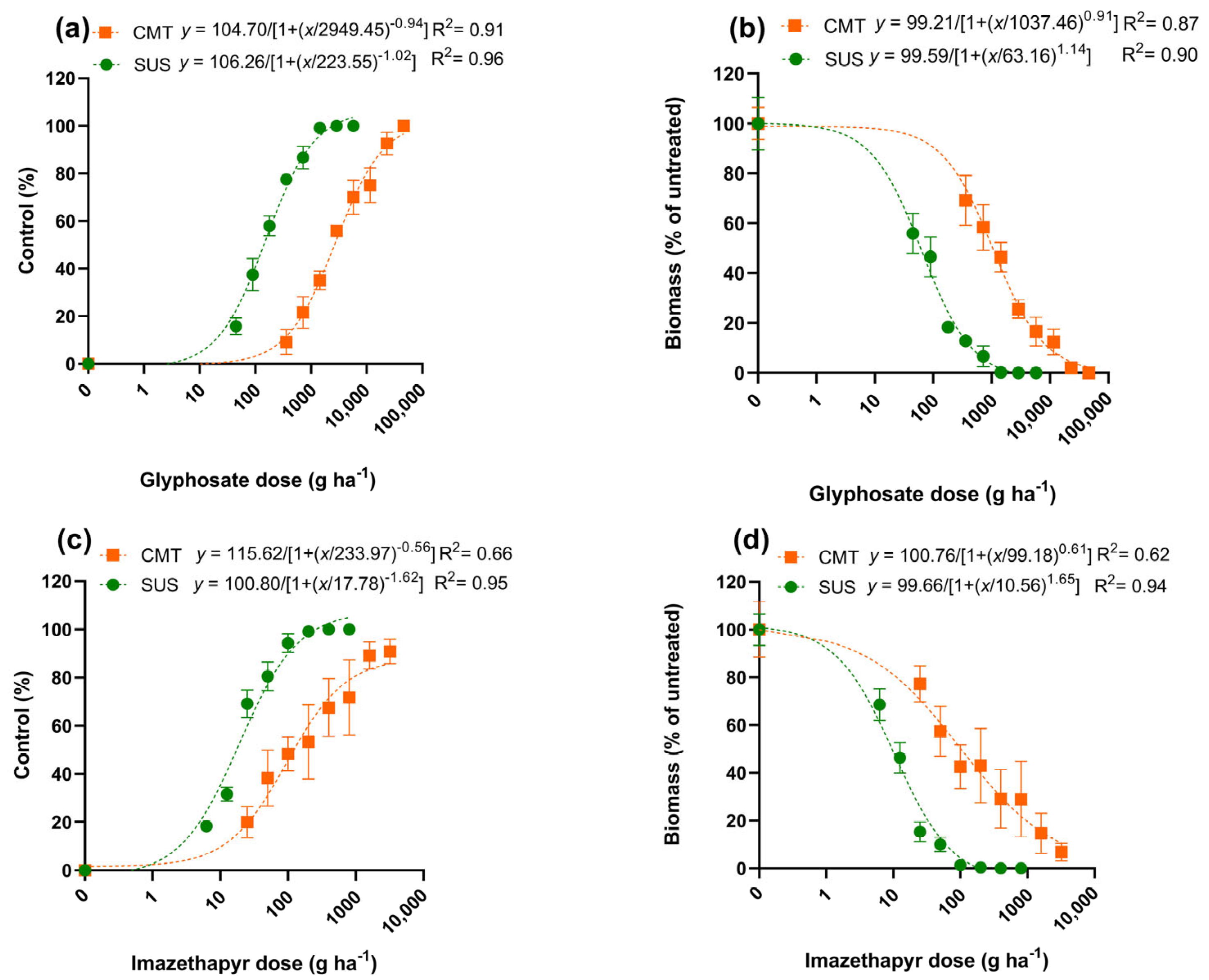
3.2. Plant Dose–Response Curve
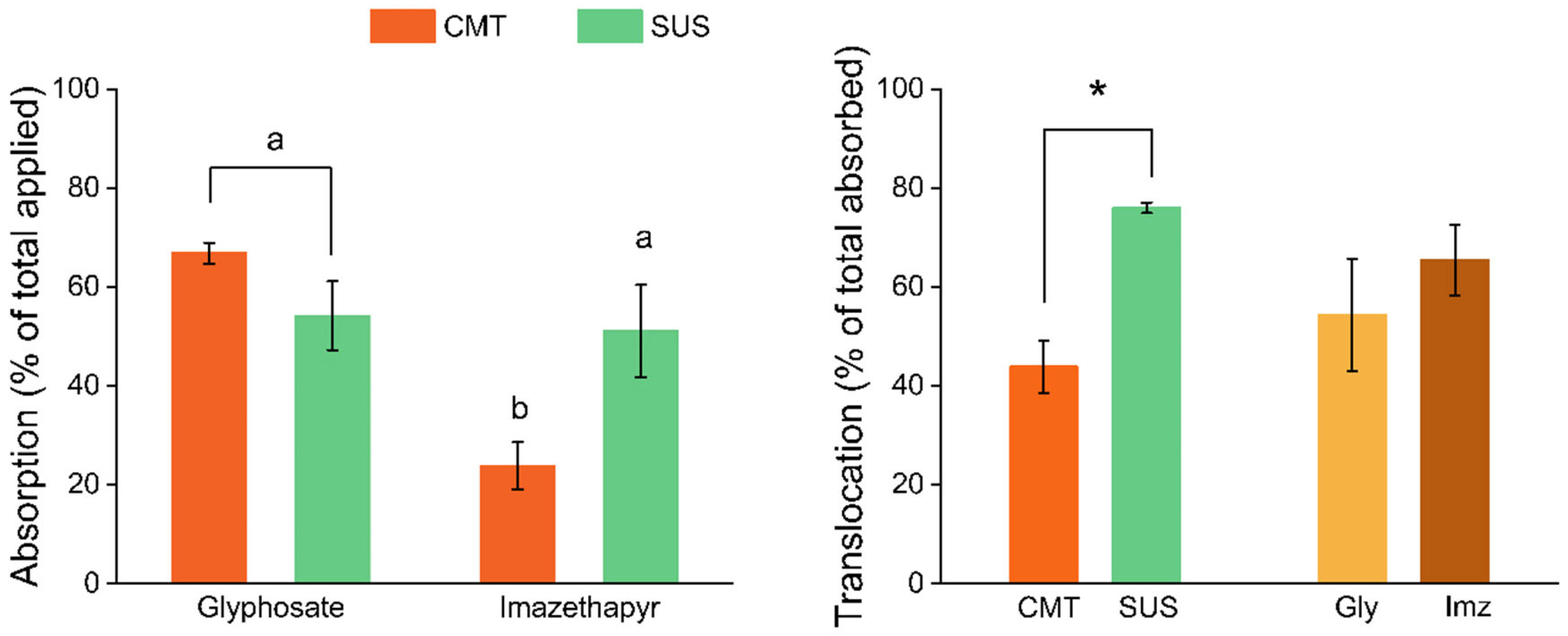
3.3. Absorption and Translocation of 14C-Glyphosate and 14C-Imazethapyr
3.4. Metabolism of 14C-Glyphosate e 14C-Imazethapyr in Plants
4. Discussion
4.1. Plant Dose-Response Curve
4.2. Absorption and Translocation of 14C-Glyphosate and 14C-Imazethapyr
4.3. 14C-Glyphosate and 14C-Imazethapyr Metabolism
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heap, I. The International Herbicide-Resistant Weed Database. Online. Available online: https://www.weedscience.org/Home.aspx (accessed on 17 April 2023).
- Shetty, M.J.; Chandan, K.; Krishna, H.C.; Aparna, G.S. Genetically Modified Crops: An overview. J. Pharmacogn. Phytochem. 2018, 7, 2405–2410. [Google Scholar]
- García, M.J.; Palma-Bautista, C.; Vazquez-Garcia, J.G.; Rojano-Delgado, A.M.; Osuna, M.D.; Torra, J.; De Prado, R. Multiple Mutations in the EPSPS and ALS Genes of Amaranthus hybridus Underlie Resistance to Glyphosate and ALS Inhibitors. Sci. Rep. 2020, 10, 17681. [Google Scholar] [CrossRef] [PubMed]
- Duke, S.O. The History and Current Status of Glyphosate. Pest Manag. Sci. 2018, 74, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- USDA. United States Department of Agriculture. Online. Available online: https://apps.fas.usda.gov/psdonline/app/index.html#/app/compositeViz (accessed on 17 April 2023).
- ISAAA. International Service for the Acquisition of Agri-Biotech Applications. Online. 2018. Available online: https://www.isaaa.org/resources/infographics/wherearebiotechcropsgrown/default.asp (accessed on 18 April 2023).
- Coble, H.D.; Schroeder, J. Call to Action on Herbicide Resistance Management. Weed Sci. 2016, 64, 661–666. [Google Scholar] [CrossRef]
- Wechsler, S.J.; McFadden, J.R.; Smith, D.J. What Do Farmers’ Weed Control Decisions Imply about Glyphosate Resistance? Evidence from Surveys of US Corn Fields. Pest Manag. Sci. 2018, 74, 1143–1154. [Google Scholar] [CrossRef]
- EMBRAPA. Brazilian Agricultural Research Company. Online. 2017. Available online: https://www.embrapa.br/busca-de-noticias/-/noticia/29194891/plantas-daninhas-resistentes-aumentam-custo-de-producao-desoja#:~:text=dos%20produtores%20brasileiros.%E2%80%9D-,Segundo%20o%20pesquisador%2C%20o%20custo%20m%C3%A9dio%20no%20Brasil%20para%20o,glifosato%20para%20controle%20da%20infestante (accessed on 18 April 2023).
- Gonçalves Netto, A.; Nicolai, M.; Carvalho, S.J.P.; Borgato, E.A.; Christoffoleti, P.J. Resistência Múltipla de Amaranthus Palmeri Aos Herbicidas Inibidores Da ALS e EPSPS No Estado Do Mato Grosso, Brasil. Planta Daninha 2016, 34, 581–587. [Google Scholar] [CrossRef]
- Faccini, D.; Vitta, J.I. Germination Characteristics of Amaranthus quitensis as Affected by Seed Production Date and Duration of Burial. Weed Res. 2005, 45, 371–378. [Google Scholar] [CrossRef]
- Dellaferrera, I.; Cortés, E.; Panigo, E.; De Prado, R.; Christoffoleti, P.; Perreta, M. First Report of Amaranthus hybridus with Multiple Resistance to 2,4-D, Dicamba, and Glyphosate. Agronomy 2018, 8, 140. [Google Scholar] [CrossRef]
- Barroso, A.A.M.; Yamauti, M.S.; Nepomuceno, M.P.; Alves, P.L.C.A. Efeito da densidade e da distância de caruru-de-mancha e amendoim-bravo na cultura do feijoeiro. Planta Daninha 2012, 30, 47–53. [Google Scholar] [CrossRef]
- Chaudhari, S.; Jordan, D.L.; York, A.C.; Jennings, K.M.; Cahoon, C.W.; Chandi, A.; Inman, M.D. Biology and Management of Glyphosate-Resistant and Glyphosate-Susceptible Palmer Amaranth (Amaranthus palmeri) Phenotypes from a Segregating Population. Weed Sci. 2017, 65, 755–768. [Google Scholar] [CrossRef]
- Küpper, A.; Borgato, E.A.; Patterson, E.L.; Netto, A.G.; Nicolai, M.; Carvalho, S.J.P.; Nissen, S.J.; Gaines, T.A.; Christoffoleti, P.J. Multiple Resistance to Glyphosate and Acetolactate Synthase Inhibitors in Palmer Amaranth (Amaranthus palmeri) Identified in Brazil. Weed Sci. 2017, 65, 317–326. [Google Scholar] [CrossRef]
- Milani, A.; Panozzo, S.; Pinton, S.; Danielis, R.A.; Sattin, M.; Scarabel, L. Diversity and Spread of Acetolactate Synthase Allelic Variants at Position 574 Endowing Resistance in Amaranthus hybridus in Italy. Plants 2023, 12, 332. [Google Scholar] [CrossRef] [PubMed]
- Resende, L.S.; Christoffoleti, P.J.; Netto, A.G.; Presoto, J.C.; Nicolai, M.; Maschietto, E.H.G.; Borsato, E.F.; Penckowski, L.H. Glyphosate-Resistant Smooth-Pigweed (Amaranthus hybridus) in Brazil. Adv. Weed Sci. 2022, 40, e20210022. [Google Scholar] [CrossRef] [PubMed]
- Dayan, F.E.; Owens, D.K.; Tranel, P.J.; Preston, C.; Duke, S.O. Evolution of Resistance to Phytoene Desaturase and Protoporphyrinogen Oxidase Inhibitors—State of Knowledge. Pest Manag. Sci. 2014, 70, 1358–1366. [Google Scholar] [CrossRef]
- Kaundun, S.S. Resistance to Acetyl-CoA Carboxylase-Inhibiting Herbicides. Pest Manag. Sci. 2014, 70, 1405–1417. [Google Scholar] [CrossRef]
- Gaines, T.A.; Duke, S.O.; Morran, S.; Rigon, C.A.G.; Tranel, P.J.; Küpper, A.; Dayan, F.E. Mechanisms of Evolved Herbicide Resistance. J. Biol. Chem. 2020, 295, 10307–10330. [Google Scholar] [CrossRef]
- Délye, C.; Jasieniuk, M.; Le Corre, V. Deciphering the Evolution of Herbicide Resistance in Weeds. Trends Genet. 2013, 29, 649–658. [Google Scholar] [CrossRef]
- Gaines, T.A.; Zhang, W.; Wang, D.; Bukun, B.; Chisholm, S.T.; Shaner, D.L.; Nissen, S.J.; Patzoldt, W.L.; Tranel, P.J.; Culpepper, A.S.; et al. Gene Amplification Confers Glyphosate Resistance in Amaranthus palmeri. Proc. Natl. Acad. Sci. USA 2010, 107, 1029–1034. [Google Scholar] [CrossRef]
- Nandula, V.K.; Ray, J.D.; Ribeiro, D.N.; Pan, Z.; Reddy, K.N. Glyphosate Resistance in Tall Waterhemp (Amaranthus tuberculatus) from Mississippi Is Due to Both Altered Target-Site and Nontarget-Site Mechanisms. Weed Sci. 2013, 61, 374–383. [Google Scholar] [CrossRef]
- Lorentz, L.; Gaines, T.A.; Nissen, S.J.; Westra, P.; Strek, H.J.; Dehne, H.W.; Ruiz-Santaella, J.P.; Beffa, R. Characterization of Glyphosate Resistance in Amaranthus tuberculatus Populations. J. Agric. Food Chem. 2014, 62, 8134–8142. [Google Scholar] [CrossRef]
- Perotti, V.E.; Larran, A.S.; Palmieri, V.E.; Martinatto, A.K.; Alvarez, C.E.; Tuesca, D.; Permingeat, H.R. A Novel Triple Amino Acid Substitution in the EPSPS Found in a High-Level Glyphosate-Resistant Amaranthus hybridus Population from Argentina. Pest Manag. Sci. 2019, 75, 1242–1251. [Google Scholar] [CrossRef] [PubMed]
- García, M.J.; Palma-Bautista, C.; Rojano-Delgado, A.M.; Bracamonte, E.; Portugal, J.; Alcántara-De la Cruz, R.; De Prado, R. The Triple Amino Acid Substitution Tap-Ivs in the Epsps Gene Confers High Glyphosate Resistance to the Superweed Amaranthus hybridus. Int. J. Mol. Sci. 2019, 20, 2396. [Google Scholar] [CrossRef]
- Sibony, M.; Rubin, B. Molecular Basis for Multiple Resistance to Acetolactate Synthase-Inhibiting Herbicides and Atrazine in Amaranthus blitoides (Prostrate Pigweed). Planta 2003, 216, 1022–1027. [Google Scholar] [CrossRef] [PubMed]
- Mcnaughton, K.E.; Letarte, J.; Lee, E.A.; François, J. Mutations in ALS Confer Herbicide Resistance in Redroot Pigweed (Amaranthus retroflexus) and Powell Amaranth (Amaranthus powellii). Weed Sci. 2005, 53, 17–22. [Google Scholar] [CrossRef]
- Chen, J.; Huang, Z.; Zhang, C.; Huang, H.; Wei, S.; Chen, J.; Wang, X. Molecular Basis of Resistance to Imazethapyr in Redroot Pigweed (Amaranthus retroflexus L.) Populations from China. Pestic. Biochem. Physiol. 2015, 124, 43–47. [Google Scholar] [CrossRef]
- Huang, Z.; Chen, J.; Zhang, C.; Huang, H.; Wei, S.; Zhou, X.; Chen, J.; Wang, X. Target-Site Basis for Resistance to Imazethapyr in Redroot Amaranth (Amaranthus retroflexus L.). Pestic. Biochem. Physiol. 2016, 128, 10–15. [Google Scholar] [CrossRef]
- Larran, A.S.; Lorenzetti, F.; Tuesca, D.; Perotti, V.E.; Permingeat, H.R. Molecular Mechanisms Endowing Cross-Resistance to ALS-Inhibiting Herbicides in Amaranthus hybridus from Argentina. Plant Mol. Biol. Rep. 2018, 36, 907–912. [Google Scholar] [CrossRef]
- Nol, N.; Tsikou, D.; Eid, M.; Livieratos, I.C.; Giannopolitis, C.N. Shikimate Leaf Disc Assay for Early Detection of Glyphosate Resistance in Conyza canadensis and Relative Transcript Levels of EPSPS and ABC Transporter Genes. Weed Res. 2012, 52, 233–241. [Google Scholar] [CrossRef]
- Moretti, M.L.; Van Horn, C.R.; Robertson, R.; Segobye, K.; Weller, S.C.; Young, B.G.; Johnson, W.G.; Douglas Sammons, R.; Wang, D.; Ge, X.; et al. Glyphosate Resistance in Ambrosia trifida: Part 2. Rapid Response Physiology and Non-Target-Site Resistance. Pest Manag. Sci. 2018, 74, 1079–1088. [Google Scholar] [CrossRef]
- Van Horn, C.R.; Moretti, M.L.; Robertson, R.R.; Segobye, K.; Weller, S.C.; Young, B.G.; Johnson, W.G.; Schulz, B.; Green, A.C.; Jeffery, T.; et al. Glyphosate Resistance in Ambrosia trifida: Part 1. Novel Rapid Cell Death Response to Glyphosate. Pest Manag. Sci. 2018, 74, 1071–1078. [Google Scholar] [CrossRef]
- Dominguez-Valenzuela, J.A.; Gherekhloo, J.; Fernández-Moreno, P.T.; Cruz-Hipolito, H.E.; Alcántara-de la Cruz, R.; Sánchez-González, E.; De Prado, R. First Confirmation and Characterization of Target and Non-Target Site Resistance to Glyphosate in Palmer Amaranth (Amaranthus palmeri) from Mexico. Plant Physiol. Biochem. 2017, 115, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Larran, A.S.; Palmieri, V.E.; Tuesca, D.; Permingeat, H.R.; Perotti, V.E. Coexistence of Target-Site and Non-Target-Site Mechanisms of Glyphosate Resistance in Amaranthus palmeri Populations from Argentina. Acta Sci.-Agron. 2022, 44, e55183. [Google Scholar] [CrossRef]
- Ghanizadeh, H.; Harrington, K.C. Non-Target Site Mechanisms of Resistance to Herbicides. CRC. Crit. Rev. Plant Sci. 2017, 36, 24–34. [Google Scholar] [CrossRef]
- Wright, A.A.; Molin, W.T.; Nandula, V.K. Distinguishing between Weedy Amaranthus Species Based on Intron 1 Sequences from the 5-Enolpyruvylshikimate-3-Phosphate Synthase Gene. Pest Manag. Sci. 2016, 72, 2347–2354. [Google Scholar] [CrossRef] [PubMed]
- Gazziero, D.L.P. Procedures for Installation, Evaluation and Analysis of Experiments with Herbicides. Braz. Weed Sci. Soc. 2005, 1, 1–42. [Google Scholar]
- Xu, Y.; Li, S.; Hao, L.; Li, X.; Zheng, M. Tribenuron-Methyl-Resistant Descurainia sophia L. Exhibits Negative Cross-Resistance to Imazethapyr Conferred by a Pro197Ser Mutation in Acetolactate Synthase and Reduced Metabolism. Pest Manag. Sci. 2022, 78, 1467–1473. [Google Scholar] [CrossRef]
- Monquero, P.A.; Christoffoleti, P.J.; Osuna, M.D.; De Prado, R.A. Absorção, Translocação e Metabolismo Do Glyphosate Por Plantas Tolerantes e Suscetíveis a Este Herbicida. Planta Daninha 2004, 22, 445–451. [Google Scholar] [CrossRef]
- Sprankle, P.; Sandberg, C.L.; Meggitt, W.F.; Penner, D. Separation of Glyphosate and Possible Metabolites by Thin-Layer Chromatograpy. Weed Sci. 1978, 6, 673–674. [Google Scholar] [CrossRef]
- Streibig, J.C. Herbicide Bioassay. Weed Res. 1988, 28, 479–484. [Google Scholar] [CrossRef]
- Ritz, C.; Baty, F.; Streibig, J.C.; Gerhard, D. Dose-Response Analysis Using R. PLoS ONE 2015, 10, e0146021. [Google Scholar] [CrossRef]
- Owen, M.D.K. Diverse Approaches to Herbicide-Resistan Weed Management. Weed Sci. 2016, 64, 570–584. [Google Scholar] [CrossRef]
- Sammons, R.D.; Gaines, T.A. Glyphosate Resistance: State of Knowledge. Pest Manag. Sci. 2014, 70, 1367–1377. [Google Scholar] [CrossRef]
- Kleinman, Z.; Rubin, B. Non-Target-Site Glyphosate Resistance in Conyza bonariensis Is Based on Modified Subcellular Distribution of the Herbicide. Pest Manag. Sci. 2017, 73, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Han, Y.; Sun, Y.; Huang, H.; Wei, S.; Huang, Z. Growth and Competitiveness of ALS-Inhibiting Herbicide-Resistan Amaranthus retroflexus L. Plants 2022, 11, 2639. [Google Scholar] [CrossRef] [PubMed]
- Mendes, R.R.; Silva, V.F.V.; Ferreira, L.A.I.; de Oliveira, R.S. Sulfonylurea Resistance in Amaranthus hybridus from Southern Brazil. Rev. Ceres 2022, 69, 374–378. [Google Scholar] [CrossRef]
- Goggin, D.E.; Cawthray, G.R.; Powles, S.B. 2,4-D Resistance in Wild Radish: Reduced Herbicide Translocation via Inhibition of Cellular Transport. J. Exp. Bot. 2016, 67, 3223–3235. [Google Scholar] [CrossRef]
- De Carvalho, L.B.; Alves, P.L.D.C.A.; González-Torralva, F.; Cruz-Hipolito, H.E.; Rojano-Delgado, A.M.; De Prado, R.; Gil-Humanes, J.; Barro, F.; Luque De Castro, M.D. Pool of Resistance Mechanisms to Glyphosate in Digitaria insularis. J. Agric. Food Chem. 2012, 60, 615–622. [Google Scholar] [CrossRef]
- Adu-Yeboah, P.; Malone, J.M.; Gill, G.; Preston, C. Reduced Glyphosate Translocation in Two Glyphosate-Resistant Populations of Rigid Ryegrass (Lolium rigidum) from Fence Lines in South Australia. Weed Sci. 2014, 62, 4–10. [Google Scholar] [CrossRef]
- Palma-Bautista, C.; Torra, J.; Garcia, M.J.; Bracamonte, E.; Rojano-Delgado, A.M.; Alcántara-De La Cruz, R.; De Prado, R. Reduced Absorption and Impaired Translocation Endows Glyphosate Resistance in Amaranthus palmeri Harvested in Glyphosate-Resistant Soybean from Argentina. J. Agric. Food Chem. 2019, 67, 1052–1060. [Google Scholar] [CrossRef]
- Ghanizadeh, H.; Harrington, K.C.; James, T.K.; Woolley, D.J.; Ellison, N.W. Mechanisms of Glyphosate Resistance in Two Perennial Ryegrass (Lolium perenne) Populations. Pest Manag. Sci. 2015, 71, 1617–1622. [Google Scholar] [CrossRef]
- Alcántara de la Cruz, R.; Barro, F.; Domínguez-Valenzuela, J.A.; De Prado, R. Physiological, Morphological and Biochemical Studies of Glyphosate Tolerance in Mexican Cologania (Cologania broussonetii (Balb.) DC.). Plant Physiol. Biochem. 2016, 98, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; D’Avignon, D.A.; Ackerman, J.J.H.; Collavo, A.; Sattin, M.; Ostrander, E.L.; Hall, E.L.; Sammons, R.D.; Preston, C. Vacuolar Glyphosate-Sequestration Correlates with Glyphosate Resistance in Ryegrass (Lolium Spp.) from Australia, South America, and Europe: A 31P NMR Investigation. J. Agric. Food Chem. 2012, 60, 1243–1250. [Google Scholar] [CrossRef] [PubMed]
- D’Avignon, D.A.; Ge, X. In Vivo NMR Investigations of Glyphosate Influences on Plant Metabolism. J. Magn. Reson. 2018, 292, 59–72. [Google Scholar] [CrossRef]
- Ge, X.; D’avignon, D.A.; Ackerman, J.J.H.; Sammons, R.D. In Vivo 31p-Nuclear Magnetic Resonance Studies of Glyphosate Uptake, Vacuolar Sequestration, and Tonoplast Pump Activity in Glyphosate-Resistant Horseweed. Plant Physiol. 2014, 166, 1255–1268. [Google Scholar] [CrossRef]
- Fernández-Moreno, P.T.; Bastida, F.; De Prado, R. Evidence, Mechanism and Alternative Chemical Seedbank-Level Control of Glyphosate Resistance of a Rigid Ryegrass (Lolium rigidum) Biotype from Southern Spain. Front. Plant Sci. 2017, 8, 450. [Google Scholar] [CrossRef]
- Lonhienne, T.; Garcia, M.D.; Low, Y.S.; Guddat, L.W. Herbicides That Inhibit Acetolactate Synthase. Front. Agric. Sci. Eng. 2022, 9, 155–160. [Google Scholar] [CrossRef]
- Murphy, B.P.; Tranel, P.J. Target-Site Mutations Conferring Herbicide Resistance. Plants 2019, 8, 382. [Google Scholar] [CrossRef] [PubMed]
- Bracamonte, E.; Fernández-Moreno, P.T.; Barro, F.; de Prado, R. Glyphosate-Resistant Parthenium hysterophorus in the Caribbean Islands: Non-Target Site Resistance and Target Site Resistance in Relation to Resistance Levels. Front. Plant Sci. 2016, 7, 1845. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.S.; Tranel, P.J.; Stewart, C.N. Non-Target-Site Herbicide Resistance: A Family Business. Trends Plant Sci. 2007, 12, 6–13. [Google Scholar] [CrossRef]
- González-Torralva, F.; Rojano-Delgado, A.M.; Luque de Castro, M.D.; Mülleder, N.; De Prado, R. Two Non-Target Mechanisms Are Involved in Glyphosate-Resistant Horseweed (Conyza canadensis L. Cronq.) Biotypes. J. Plant Physiol. 2012, 169, 1673–1679. [Google Scholar] [CrossRef]
- Chen, R.-F.; Wang, H.-H.; Wang, C.-Y. Translocation and Metabolism of Injected Glyphosate in Lead Tree (Leucaena leucocephala). Weed Sci. 2009, 57, 229–234. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Herbicide | Population | Field Dose | Dose–Response (g a.i. or e.a. ha−1) |
|---|---|---|---|
| Glyphosate | CMT | 1440 a | 0, 360, 720, 1440, 2880, 5760, 11,520, 23,040, 46,080 |
| SUS | 1440 a | 0, 45, 90, 180, 360, 720, 1440, 2880, 5760 | |
| Imazethapyr | CMT | 100 b | 0, 25, 50, 100, 200, 400, 800, 1600, 3200 |
| SUS | 100 b | 0, 6.25, 12.5, 25, 50, 100, 200, 400, 800 |
| Glyphosate | Population | |
|---|---|---|
| SUS | CMT | |
| LD50 | 223.5 ± 40 | 2949.4 ± 677 |
| RF 1 (LD50) | - | 13.1 |
| GR50 | 63.1 ± 13 | 1037.4 ± 233 |
| RF 1 (GR50) | - | 16.4 |
| LD90 | 1909.7 ± 895 | 29,910.5 ± 19,623 |
| GR90 | 428 ± 135 | 11,318.4 ± 3789 |
| Imazethapyr | ||
| LD50 | 17.7 ± 2.4 | 233.9 ± 258.1 ns |
| RF 1 (LD50) | - | 13.2 |
| GR50 | 10.5 ± 1.8 | 99.1 ± 36.2 |
| RF 1 (GR50) | - | 9.4 |
| LD90 | 68.8 ± 24.0 | 11,812.1 ± 29,221.3 ns |
| GR90 | 39.9 ± 12.4 | 3493.3 ± 2095.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nalin, D.; Munhoz-Garcia, G.V.; Witter, A.P.W.; Takeshita, V.; Oliveira, C.d.; Adegas, F.S.; Tornisielo, V.L.; Oliveira Junior, R.S.d.; Constantin, J. Absorption, Translocation, and Metabolism of Glyphosate and Imazethapyr in Smooth Pigweed with Multiple Resistance. Agronomy 2023, 13, 1720. https://doi.org/10.3390/agronomy13071720
Nalin D, Munhoz-Garcia GV, Witter APW, Takeshita V, Oliveira Cd, Adegas FS, Tornisielo VL, Oliveira Junior RSd, Constantin J. Absorption, Translocation, and Metabolism of Glyphosate and Imazethapyr in Smooth Pigweed with Multiple Resistance. Agronomy. 2023; 13(7):1720. https://doi.org/10.3390/agronomy13071720
Chicago/Turabian StyleNalin, Daniel, Gustavo Vinícios Munhoz-Garcia, Ana Paula Werkhausen Witter, Vanessa Takeshita, Claudia de Oliveira, Fernando Storniolo Adegas, Valdemar Luiz Tornisielo, Rubem Silvério de Oliveira Junior, and Jamil Constantin. 2023. "Absorption, Translocation, and Metabolism of Glyphosate and Imazethapyr in Smooth Pigweed with Multiple Resistance" Agronomy 13, no. 7: 1720. https://doi.org/10.3390/agronomy13071720