
Differential Responses of Soil Microbial N-Cycling Functional Genes to 35 yr Applications of Chemical Fertilizer and Organic Manure in Wheat Field Soil on Loess Plateau


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design
2.3. Soil Sampling
2.4. Laboratory Analysis
2.5. Soil DNA Extraction and Sequencing
2.6. Metagenomic Analysis
2.7. Statistical Analysis
3. Results
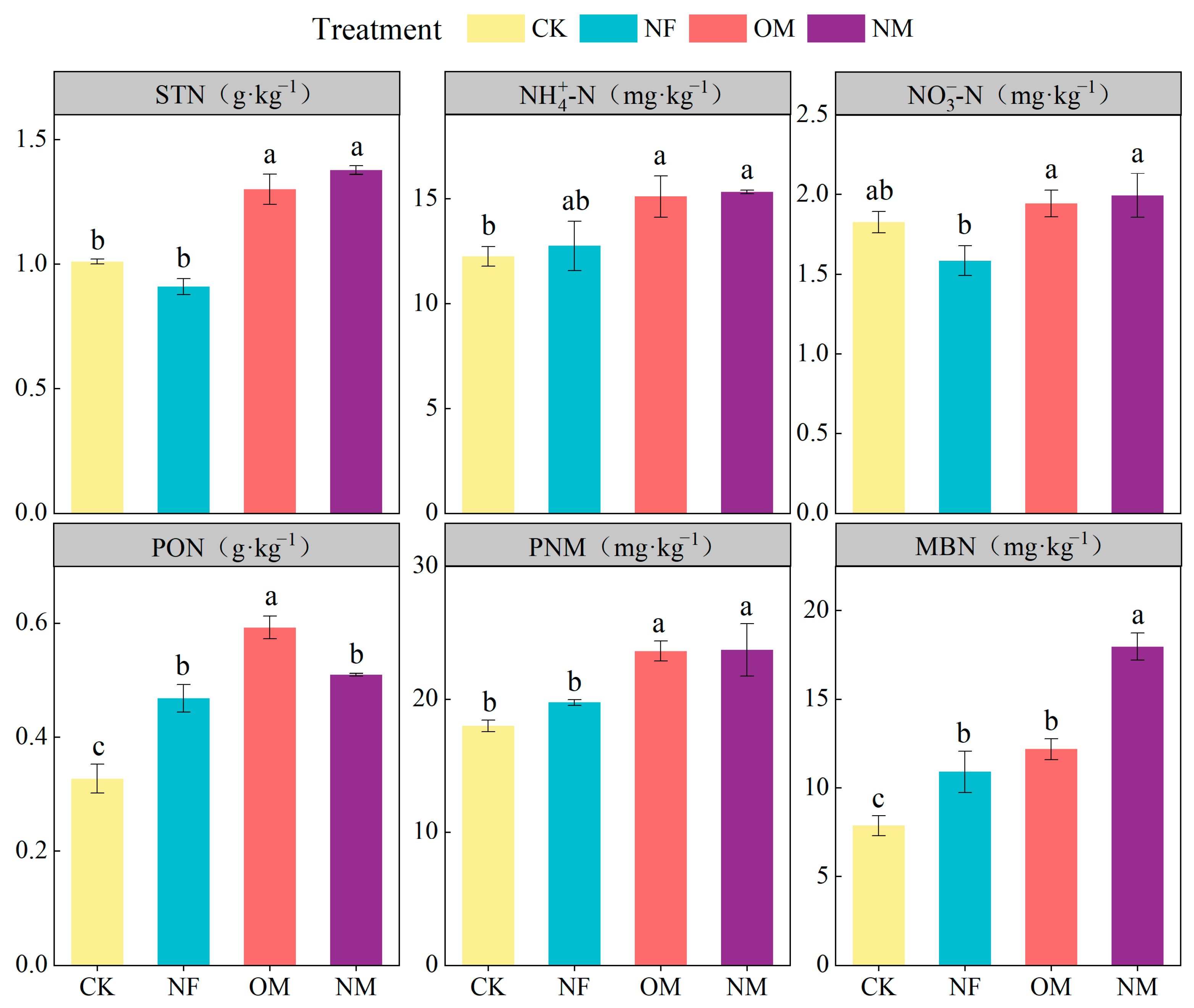
3.1. Soil N Fractions
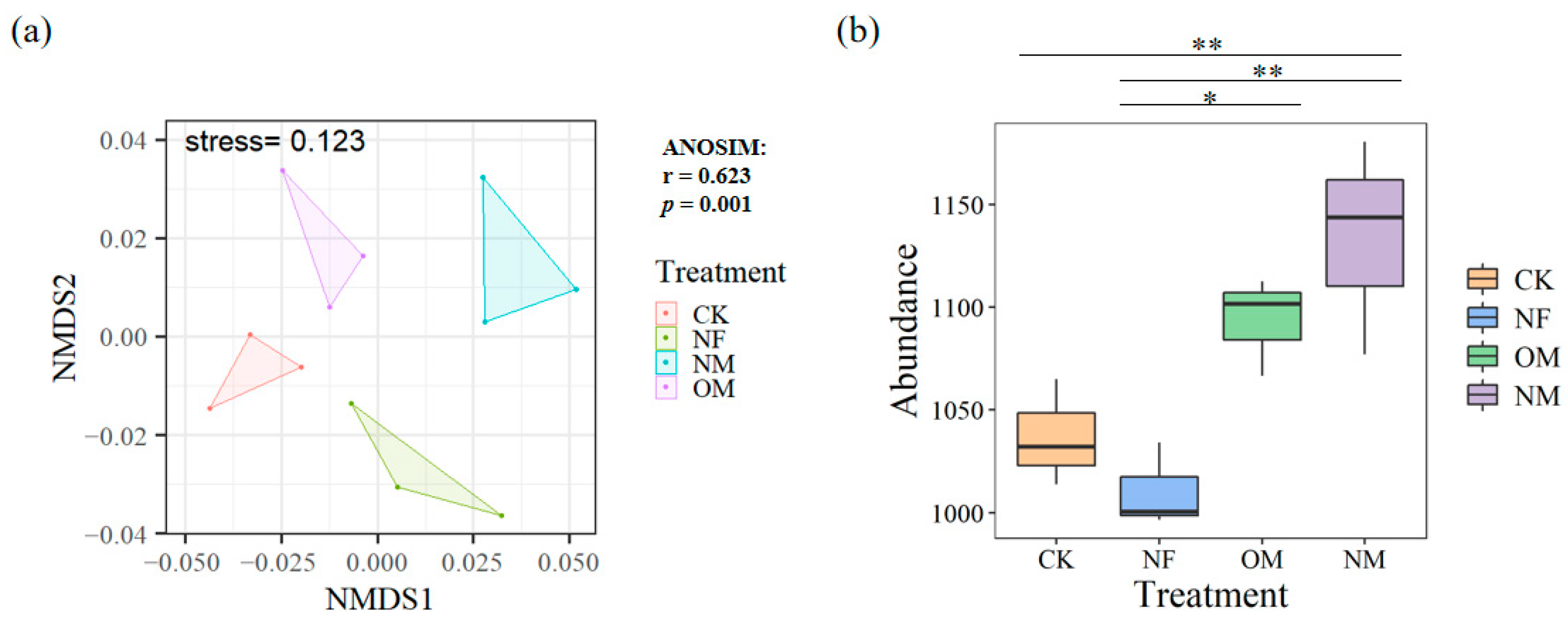
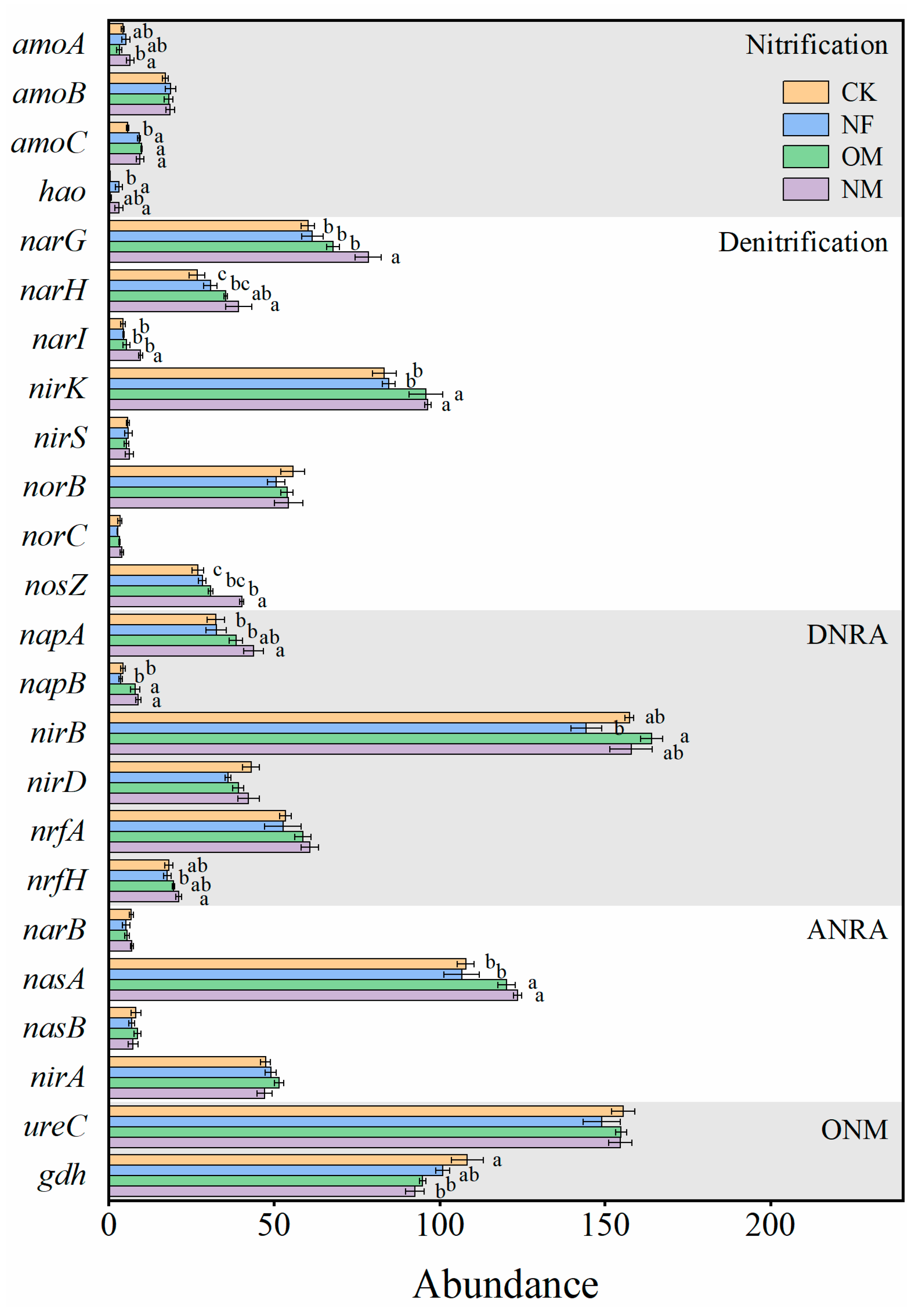
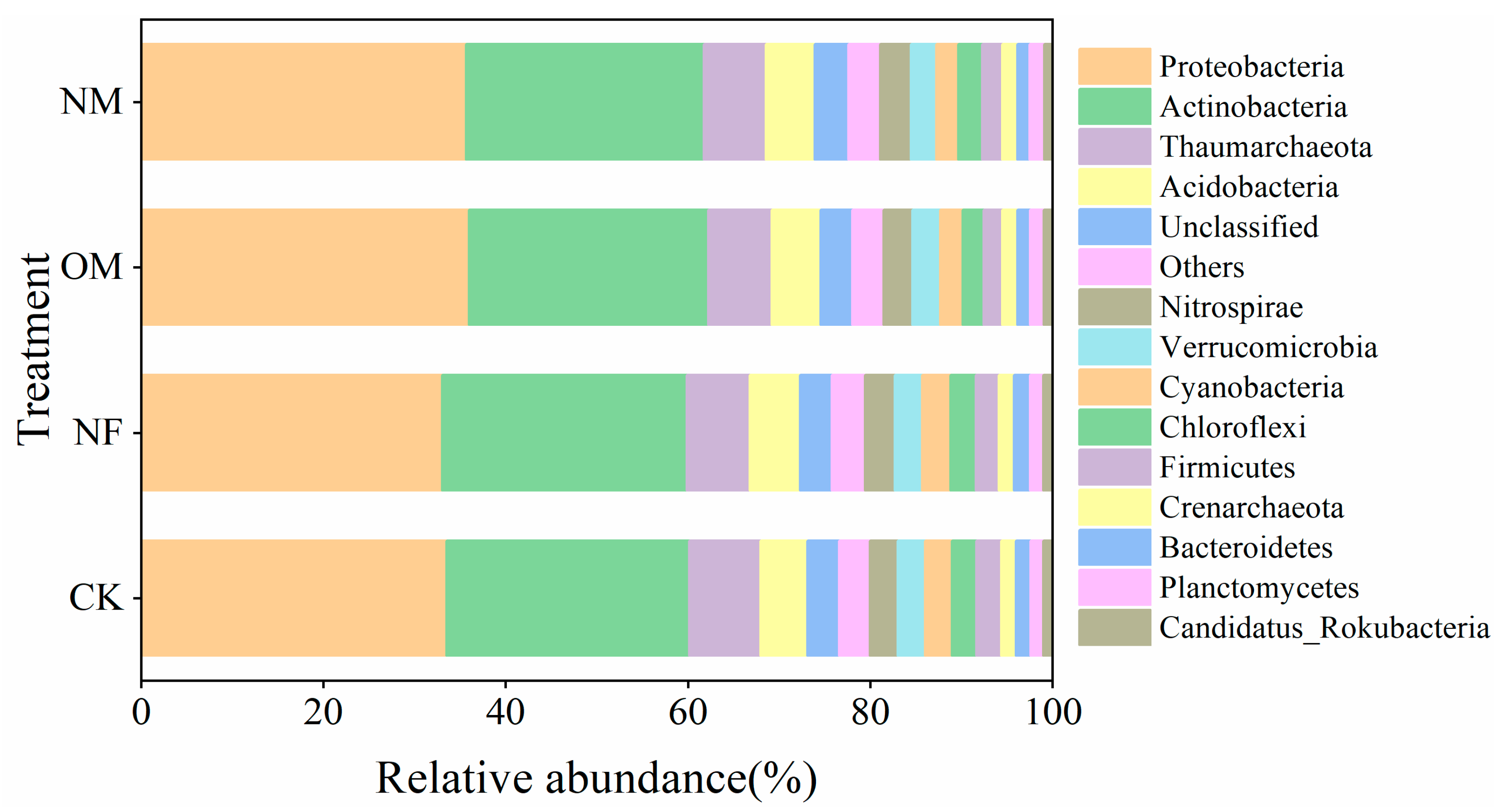
3.2. The Abundances of N-Cycling Functional Genes
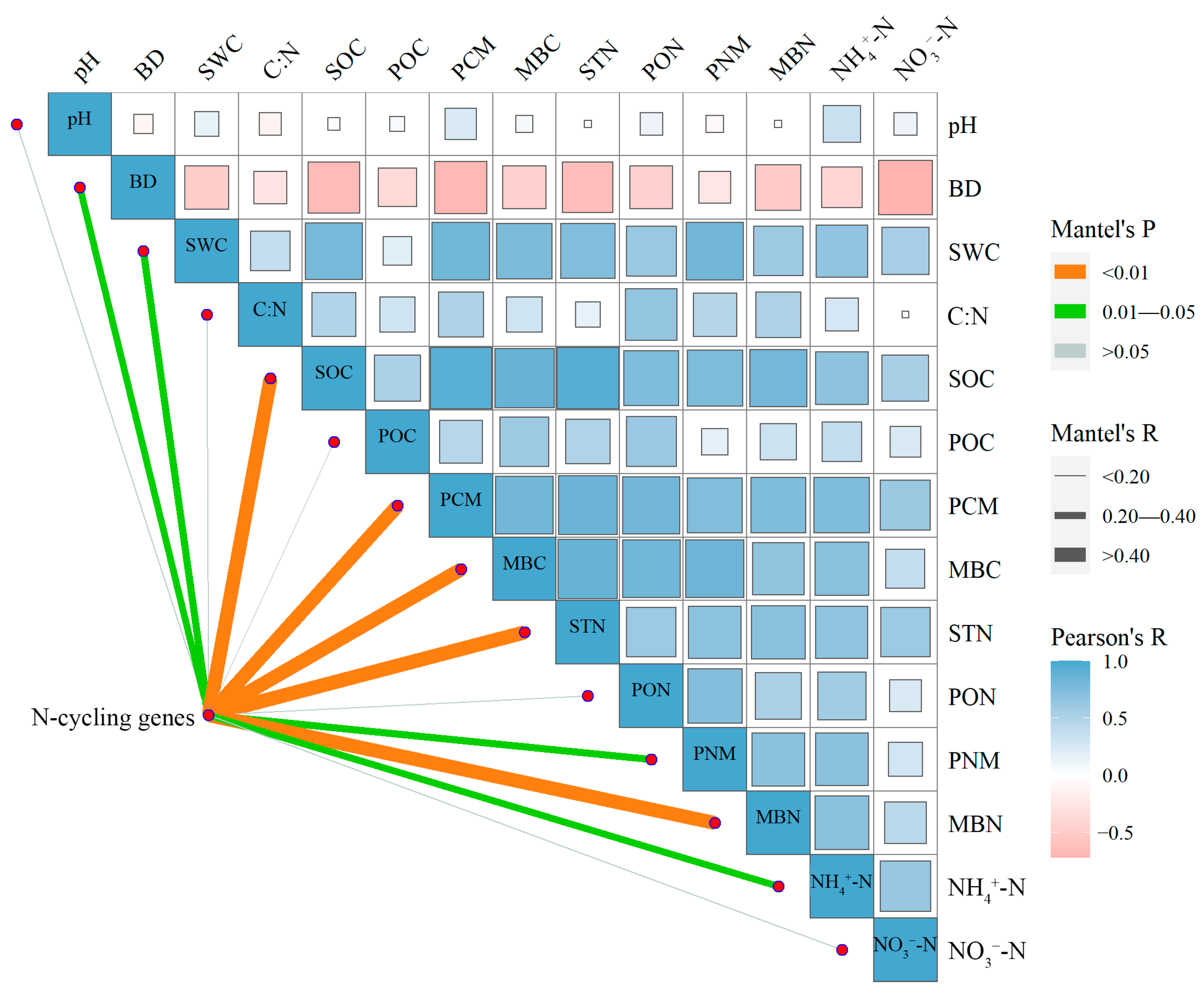
3.3. Linkages between Soil Properties and N-Cycling Functional Genes
4. Discussion
4.1. Soil N Fractions Response to Long-Term Fertilization
4.2. N-Cycling Functional Genes Response to Long-Term Fertilization
4.3. Driving Factors of N-Cycling Functional Genes
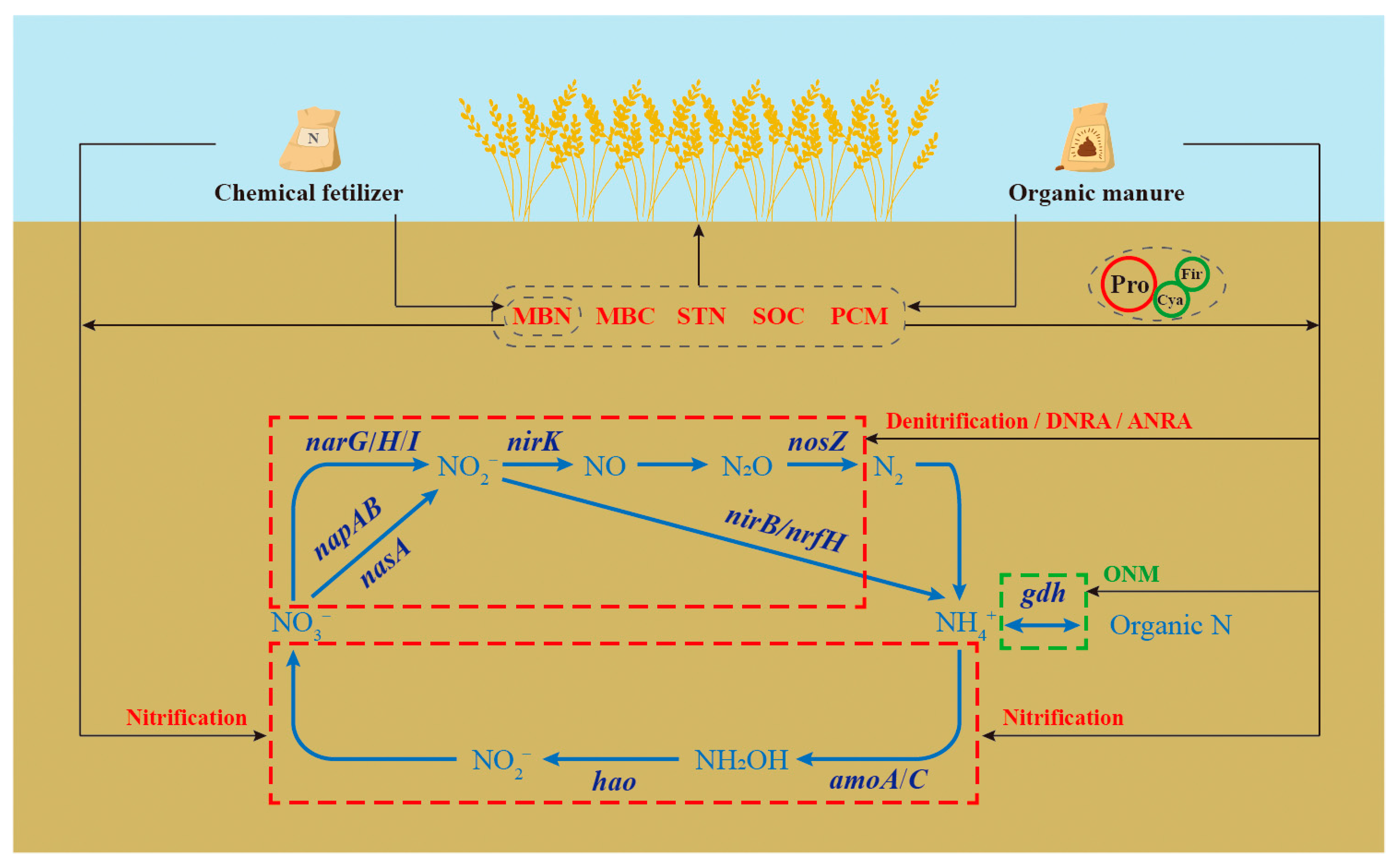
4.4. A Model of N-Cycling Genes Induced by Long-Term Fertilization
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ollivier, J.; Toewe, S.; Bannert, A.; Hai, B.; Kastl, E.-M.; Meyer, A.; Su, M.X.; Kleineidam, K.; Schloter, M. Nitrogen turnover in soil and global change. FEMS Microbiol. Ecol. 2011, 78, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Stein, L.Y.; Klotz, M.G. The nitrogen cycle. Curr. Biol. 2016, 26, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Kuypers, M.M.M.; Marchant, H.K.; Kartal, B. The microbial nitrogen-cycling network. Nat. Rev. Microbiol. 2018, 16, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Hallin, S.; Jones, C.M.; Schloter, M.; Philippot, L. Relationship between N-cycling communities and ecosystem functioning in a 50-year-old fertilization experiment. ISME J. 2009, 3, 597–605. [Google Scholar] [CrossRef]
- Sun, R.; Guo, X.; Wang, D.; Chu, H. Effects of long-term application of chemical and organic fertilizers on the abundance of microbial communities involved in the nitrogen cycle. Appl. Soil Ecol. 2015, 95, 171–178. [Google Scholar] [CrossRef]
- Wang, J.; Cheng, Y.; Cai, Z.; Zhang, J. Effects of long-term fertilization on key processes of soil nitrogen cycling in agricultural soil: A review. Acta Pedol. Sin. 2016, 53, 292–304. (In Chinese) [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, X.; Zhang, L.; Zeng, L.; Liu, Y.; Wang, X.; He, P.; Li, S.; Liang, G.; Zhou, W.; et al. The stronger impact of inorganic nitrogen fertilization on soil bacterial community than organic fertilization in short-term condition. Geoderma 2021, 382, 114752. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, C.; Wang, Y.; Hu, Y.; Christie, P.; Zhang, J.L.; Li, X. Maize yield and soil fertility with combined use of compost and inorganic fertilizers on a calcareous soil on the North China Plain. Soil Till. Res. 2016, 155, 85–94. [Google Scholar] [CrossRef]
- Ouyang, Y.; Evans, S.E.; Friesen, M.L.; Tiemann, L.K. Effect of nitrogen fertilization on the abundance of nitrogen cycling genes in agricultural soils: A meta-analysis of field studies. Soil Biol. Biochem. 2018, 127, 71–78. [Google Scholar] [CrossRef]
- Li, C.; Ma, S.; Shao, Y.; Ma, S.; Zhang, L. Effects of long-term organic fertilization on soil microbiologic characteristics, yield and sustainable production of winter wheat. J. Integr. Agr. 2018, 17, 210–219. [Google Scholar] [CrossRef]
- Zhou, Z.; Gan, Z.; Shangguan, Z.; Zhang, F. Effects of long-term repeated mineral and organic fertilizer applications on soil organic carbon and total nitrogen in a semi-arid cropland. Eur. J. Agron. 2013, 45, 20–26. [Google Scholar] [CrossRef]
- Li, W.X.; Wang, C.; Zheng, M.M.; Cai, Z.J.; Wang, B.R.; Shen, R.F. Fertilization strategies affect soil properties and abundance of N-cycling functional genes in an acidic agricultural soil. Appl. Soil Ecol. 2020, 156, 103704. [Google Scholar] [CrossRef]
- Cui, X.; Zhang, Y.; Gao, J.; Peng, F.; Gao, P. Long-term combined application of manure and chemical fertilizer sustained higher nutrient status and rhizospheric bacterial diversity in reddish paddy soil of Central South China. Sci. Rep. 2018, 8, 16554. [Google Scholar] [CrossRef]
- Gai, X.; Liu, H.; Liu, J.; Zhai, L.; Yang, B.; Wu, S.; Ren, T.; Lei, Q.; Wang, H. Long-term benefits of combining chemical fertilizer and manure applications on crop yields and soil carbon and nitrogen stocks in North China Plain. Agric. Water Manag. 2018, 208, 384–392. [Google Scholar] [CrossRef]
- Nelson, M.B.; Martiny, A.C.; Martiny, J.B.H. Global biogeography of microbial nitrogen-cycling traits in soil. Proc. Natl. Acad. Sci. USA 2016, 113, 8033–8040. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, S.; Liu, Y.; Feng, K.; Deng, Y. The applications of metagenomics in the detection of environmental microbes involving in nitrogen cycle. Biotech. Bull. 2018, 34, 1–14. (In Chinese) [Google Scholar] [CrossRef]
- Guo, S.; Wu, J.; Dang, T. Effects of crop rotation and fertilization on aboveground biomass and soil organic C in semi-arid region. Sci. Agric. Sin. 2008, 41, 744–751. (In Chinese) [Google Scholar]
- Wang, J.; Fu, X.; Ghimire, R.; Sainju, U.M.; Jia, Y.; Zhao, F. Responses of soil bacterial community and enzyme activity to organic matter components under long-term fertilization on the Loess Plateau of China. Appl. Soil Ecol. 2021, 166, 103992. [Google Scholar] [CrossRef]
- Cambardella, C.A.; Elliott, E.T. Particulate soil organic-matter changes across a grassland cultivation sequence. Soil Sci. Soc. Am. J. 1992, 56, 777–783. [Google Scholar] [CrossRef]
- Haney, R.L.; Franzluebbers, A.J.; Porter, E.B.; Hons, F.M.; Zuberer, D.A. Soil carbon and nitrogen mineralization: Influence of drying temperature. Soil Sci. Soc. Am. J. 2004, 68, 489–492. [Google Scholar] [CrossRef]
- Franzluebbers, A.J.; Hons, F.M.; Zuberer, D.A. Soil organic-carbon, microbial biomass, and mineralizable carbon and nitrogen in sorghum. Soil Sci. Soc. Am. J. 1995, 59, 460–466. [Google Scholar] [CrossRef]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D.; et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Fan, T.; Zhao, G.; Dang, Y.; Wang, L.; Li, S.; Cheng, W. Effect of long-term fixed application of different material replacing partly chemical nitrogenous fertilizer on soil nutrition in dry land of eastern Gansu province. China Soils Fert. 2018, 3, 85–91. (In Chinese) [Google Scholar] [CrossRef]
- Gai, X.; Liu, H.; Zhai, L.; Yang, B.; Ren, T.; Wang, H.; Wu, S.; Lei, Q. Effects of long-term additional application of organic manure or straw incorporation on soil nitrogen leaching risk. Sci. Agric. Sin. 2018, 51, 2336–2347. (In Chinese) [Google Scholar] [CrossRef]
- Yan, D.; Wang, D.; Yang, L. Long-term effect of chemical fertilizer, straw, and manure on labile organic matter fractions in a paddy soil. Biol. Fert. Soils 2007, 44, 93–101. [Google Scholar] [CrossRef]
- Zhao, D.; Wang, J.; Fu, X. Effect of long-term fertilization on soil total nitrogen and its fractions in dryland farming system. J. Soil Water Conserv. 2016, 30, 303–307. (In Chinese) [Google Scholar] [CrossRef]
- Chen, J.; Liang, G.; Zhou, W.; Wang, X.; Sun, J.; Liu, D.; Hu, C. Responses of soil organic carbon and nitrogen fractions to long-term organic fertilization under rice-wheat rotation. Plant Nutr. Fert. Sci. 2019, 25, 36–44. (In Chinese) [Google Scholar] [CrossRef]
- Zang, Y.; Hao, M.; Zhang, L.; Zhang, H. Effects of wheat cultivation and fertilization on soil microbial biomass carbon, soil microbial biomass nitrogen and soil basal respiration in 26 years. Acta Ecol. Sin. 2015, 35, 1445–1451. (In Chinese) [Google Scholar] [CrossRef]
- Zhang, Q.; Miao, F.; Wang, Z.; Shen, Y.; Wang, G. Effects of long-term fertilization management practices on soil microbial biomass in China’s cropland: A meta-analysis. Agron. J. 2017, 109, 1183–1195. [Google Scholar] [CrossRef]
- Gong, W.; Yan, X.; Cai, Z.; Wang, J.; Hu, T.; Gong, Y.; Ran, H. Effects of long-term fertilization on soil particulate organic carbon and nitrogen in a wheat-maize cropping system. Chin. J. Appl. Ecol. 2008, 19, 2375–2381. (In Chinese) [Google Scholar]
- Wang, H.; Xu, M.; Ma, X.; Duan, Y. Research advances of microorganism and ammonia oxidizing bacteria under long-term fertilization in Chinese typical cropland. China Soils Fert. 2018, 2, 1–12. (In Chinese) [Google Scholar] [CrossRef]
- Mueller, C.; Laughlin, R.J.; Christie, P.; Watson, C.J. Effects of repeated fertilizer and cattle slurry applications over 38 years on N dynamics in a temperate grassland soil. Soil Biol. Biochem. 2011, 43, 1362–1371. [Google Scholar] [CrossRef]
- Zhang, J.B.; Zhu, T.B.; Cai, Z.C.; Qin, S.W.; Mueller, C. Effects of long-term repeated mineral and organic fertilizer applications on soil nitrogen transformations. Eur. J. Soil Sci. 2012, 63, 75–85. [Google Scholar] [CrossRef]
- Sun, R.; Wang, F.; Hu, C.; Liu, B. Metagenomics reveals taxon-specific responses of the nitrogen-cycling microbial community to long-term nitrogen fertilization. Soil Biol. Biochem. 2021, 156, 108214. [Google Scholar] [CrossRef]
- Ouyang, Y.; Norton, J.M.; Stark, J.M.; Reeve, J.R.; Habteselassie, M.Y. Ammonia-oxidizing bacteria are more responsive than archaea to nitrogen source in an agricultural soil. Soil Biol. Biochem. 2016, 96, 4–15. [Google Scholar] [CrossRef]
- Luo, G.; Friman, V.P.; Chen, H.; Liu, M.; Wang, M.; Guo, S.; Ling, N.; Shen, Q. Long-term fertilization regimes drive the abundance and composition of N-cycling-related prokaryotic groups via soil particle-size differentiation. Soil Biol. Biochem. 2018, 116, 213–223. [Google Scholar] [CrossRef]
- Mooshammer, M.; Wanek, W.; Haemmerle, I.; Fuchslueger, L.; Hofhansl, F.; Knoltsch, A.; Schnecker, J.; Takriti, M.; Watzka, M.; Wild, B.; et al. Adjustment of microbial nitrogen use efficiency to carbon: Nitrogen imbalances regulates soil nitrogen cycling. Nat. Commun. 2014, 5, 3694. [Google Scholar] [CrossRef]
- Wang, J.; Lin, S.; Li, B. Nitrogen cycling and management strategies in chinese agriculture. Sci. Agric. Sin. 2016, 49, 503–517. (In Chinese) [Google Scholar] [CrossRef]
- Chu, C.; Wu, Z.; Huang, Q.; Han, C.; Zhong, W. Effect of organic matter promotion on nitrogen-cycling genes and functional microorganisms in acidic red soils. Chin. J. Environ. Sci. 2020, 41, 2468–2475. (In Chinese) [Google Scholar] [CrossRef]
- Guo, J.; Zhu, C.; Liu, W.; Wang, J.; Ling, N.; Guo, S. Effects of different fertilization managements on functional microorganisms involved in nitrogen cycle. Plant Nutr. Fert. Sci. 2021, 27, 751–759. (In Chinese) [Google Scholar] [CrossRef]
- O’Sullivan, C.A.; Fillery, I.R.P.; Roper, M.M.; Richards, R.A. Identification of several wheat landraces with biological nitrification inhibition capacity. Plant Soil 2016, 404, 61–74. [Google Scholar] [CrossRef]
- Wang, F.; Chen, S.; Wang, Y.; Zhang, Y.; Hui, C.; Liu, B. Long-term nitrogen fertilization elevates the activity and abundance of nitrifying and denitrifying microbial communities in an upland soil: Implications for nitrogen loss from intensive agricultural systems. Front. Microbiol. 2018, 9, 02424. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ji, H.; Gao, C. Differential responses of soil bacterial taxa to long-term P, N, and organic manure application. J. Soil Sediment 2016, 16, 1046–1058. [Google Scholar] [CrossRef]
- Li, W.; Zhang, Y.; Mao, W.; Wang, C.; Yin, S. Functional potential differences between Firmicutes and Proteobacteria in response to manure amendment in a reclaimed soil. Can. J. Microbiol. 2020, 66, 689–697. [Google Scholar] [CrossRef]
- Li, Y.; Tremblay, J.; Bainard, L.D.; Cade-Menun, B.; Hamel, C. Long-term effects of nitrogen and phosphorus fertilization on soil microbial community structure and function under continuous wheat production. Environ. Microbiol. 2020, 22, 1066–1088. [Google Scholar] [CrossRef] [PubMed]
- Fan, K.; Delgado-Baquerizo, M.; Guo, X.; Wang, D.; Zhu, Y.; Chu, H. Biodiversity of key-stone phylotypes determines crop production in a 4-decade fertilization experiment. ISME J. 2021, 15, 550–561. [Google Scholar] [CrossRef]
- Li, J.; Wang, G.; Yan, B.; Liu, G. The responses of soil nitrogen transformation to nitrogen addition are mainly related to the changes in functional gene relative abundance in artificial Pinus tabulaeformis forests. Sci. Total Environ. 2020, 723, 1376779. [Google Scholar] [CrossRef]
- An, T.; Wang, F.; Ren, L.; Ma, S.; Li, S.; Liu, L.; Wang, J. Ratio of nitrate to ammonium mainly drives soil bacterial dynamics involved in nitrate reduction processes. Appl. Soil Ecol. 2022, 169, 104164. [Google Scholar] [CrossRef]
- Finzi, A.C.; Abramoff, R.Z.; Spiller, K.S.; Brzostek, E.R.; Darby, B.A.; Kramer, M.A.; Phillips, R.P. Rhizosphere processes are quantitatively important components of terrestrial carbon and nutrient cycles. Global Chang. Biol. 2015, 21, 2082–2094. [Google Scholar] [CrossRef]
- Sorensen, P. Immobilisation, remineralisation and residual effects in subsequent crops of dairy cattle slurry nitrogen compared to mineral fertiliser nitrogen. Plant Soil 2004, 267, 285–296. [Google Scholar] [CrossRef]
- Chen, L.; Hao, M.; Li, Z. Effects of long-term application of fertilizers on nitrate accumulation in the Loess Plateau dryland. Res. Soil Water Conserv. 2014, 21, 43–46. (In Chinese) [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, Y.; Yang, C.; Sainju, U.M.; Zhang, N.; Zhao, F.; Wang, W.; Wang, J. Differential Responses of Soil Microbial N-Cycling Functional Genes to 35 yr Applications of Chemical Fertilizer and Organic Manure in Wheat Field Soil on Loess Plateau. Agronomy 2023, 13, 1516. https://doi.org/10.3390/agronomy13061516
Liang Y, Yang C, Sainju UM, Zhang N, Zhao F, Wang W, Wang J. Differential Responses of Soil Microbial N-Cycling Functional Genes to 35 yr Applications of Chemical Fertilizer and Organic Manure in Wheat Field Soil on Loess Plateau. Agronomy. 2023; 13(6):1516. https://doi.org/10.3390/agronomy13061516
Chicago/Turabian StyleLiang, Yinyan, Caidi Yang, Upendra M. Sainju, Nannan Zhang, Fazhu Zhao, Weizhen Wang, and Jun Wang. 2023. "Differential Responses of Soil Microbial N-Cycling Functional Genes to 35 yr Applications of Chemical Fertilizer and Organic Manure in Wheat Field Soil on Loess Plateau" Agronomy 13, no. 6: 1516. https://doi.org/10.3390/agronomy13061516