
Seed Priming and Foliar Application with Ascorbic Acid and Salicylic Acid Mitigate Salt Stress in Wheat



1
Crop Physiology Research Department, Field Crops Research Institute, Agricultural Research Center, Giza 12619, Egypt
2
Department of Agronomy, Faculty of Agriculture, Sher-e-Bangla Agricultural University, Dhaka 1207, Bangladesh
*
Author to whom correspondence should be addressed.
Agronomy 2023, 13(2), 493; https://doi.org/10.3390/agronomy13020493
Submission received: 29 December 2022
/
Revised: 20 January 2023
/
Accepted: 6 February 2023
/
Published: 8 February 2023
(This article belongs to the Special Issue New Insight into Crops Defense Response)
Abstract
:Ascorbic acid (AA) and salicylic acid (SA) are naturally active antioxidants that protect against plant stresses, including salinity. We studied the physiological response of wheat to AA and SA (100, 200 ppm) as well as the combined treatment of AA and SA (100 ppm) through application as both priming and foliar spray treatments under saline conditions. The results showed that wheat plants under salt-affected soils exhibited numerous physiological effects in plant metabolism, which subsequently affected the qualitative and quantitative parameters of growth and yield. Moreover, the photosynthetic pigments, antioxidant content, and yield are significantly enhanced under the combined treatment of AA and SA. In contrast, the application of AA and SA lowered the osmolytes and lipid peroxidation content under saline conditions. Accordingly, the enhancement of the mentioned parameter was related to the scavenging of the reactive oxygen species and decreasing the oxidative stress on the plant under the salinity stress. Our results explore the significance of applied AA and SA as efficacious compounds in wheat farming under saline conditions. The combined application of (100 ppm) AA with (100 ppm) SA using priming or a foliar spray can be a promising treatment for beneficent wheat growth and productivity improvement under salt-affected soil conditions.
1. Introduction
The salinity problem worldwide increases due to marginal lands being brought into production, unsustainable irrigation and land clearing, and the management practices of salinity [1,2]. In general, plants’ physiological processes are disrupted by salt stress as a result of increasing osmotic pressure, nutritional imbalance, salt toxicity, and oxidative stress [3] and consequently decrease the yield [4,5]. Seed germination significantly decreased under salinity stress, reducing the vegetative growth and reproductive behavior, resulting in yield losses [6,7]. Osmotic stress induced by salinity and ionic stress causes the increase in reactive oxygen species (ROS) production in the plant, which causes damage to cell organelles and membrane components and causes cell and plant death at severe levels of salinity [8].
Wheat (Triticum aestivum) is the most important crop among other cereals due to its domestication and contribution as an important part of daily man food calories and protein intake [9,10,11].
Despite the fact that breeders significantly improve wheat grain yield potential, progress in increasing wheat yield in less favorable environments, such as salt-affected soils, is difficult to achieve [12,13]. Numerous stress elicitor applications ameliorate the plant response to salinity stress, subsequently decreasing ROS accumulation [14,15,16]. Scientists have used various techniques to overcome salinity, and one of them is using certain eco-friendly compounds to overcome various stresses on plants [17,18]. Ascorbic and salicylic acids are antioxidant compounds produced endogenously in plant cells as signal molecules [19]. They have been investigated as the most cost-efficient and efficacious compounds as they enhance tolerance to salinity for many plants, such as wheat [20,21], mung bean [19], and maize [22]. Under different abiotic stresses, the antioxidants positively detoxify ROS by triggering H2O2 scavenging enzymes (catalase, peroxidase, etc.) in whole plant cells [23,24].
Ascorbic acid (an antioxidant with low molecular weight) acts as a scavenger of ROS in plants, improving growth and increasing stress tolerance [25,26]. It has a vital role in plant cell division and expansion, photosynthesis, flowering regulation, senility in leaves, apical meristem formation, and detoxification and stress defense by neutralizing the ROS [27], in addition to being a cofactor for enzyme activity [28]. Exogenous applied AA enhances the production of ascorbic acid endogenously in plant cells, which mitigates the adverse effects of salt stress in many crops [29,30].
The application of exogenous SA decreased the adverse impacts of salinity in plants and promoted the physiological and biochemical parameters of wheat and several crops under abiotic stress conditions [31,32,33,34].
Priming seeds enhance seed germination, as well as seedling development, by expediting the pre-occurrence of metabolic events and overcoming adverse conditions by activating a stress–responsive system [35,36].
Therefore, this study was carried out to study the growth, productivity, and physiological response of wheat under salt affected-soil conditions, the practicability of priming seeds or foliar application with ascorbic and salicylic acids, and the different doses and their synergetic impact on plant growth and yield improvement for wheat varieties.
2. Materials and Methods
Two field experiments were conducted at South El Husainia Plain, El Sharkia Gov., Egypt (31°00′15″ N 32°08′15″ E) through two consecutive winter seasons 2018/2019 and 2019/2020. To investigate the physiological effects and productivity of wheat under salt-affected soil conditions and its response to seed priming and foliar application with ascorbic and salicylic acids, all the growth parameters, photosynthetic pigments, soluble sugars, antioxidant enzymes, total phenols, and lipid peroxidation were investigated. Moreover, the osmolytes, yield, Na+/K+ ratio, and protein content were also assessed in the seeds of wheat plants.
The trial was sown in a randomized complete block design (RCBD), with four replications, while the plot size of each treatment was 10.5 m2. The experiment was achieved in the same place in the two seasons and irrigated using the flood irrigation method. The experimental site soil’s physical and chemical properties are shown in Table 1. In addition, the meteorological conditions of the experimental site in seasons 2018/2019 and 2019/2020 are shown in Table 2.
Wheat grains cv. Masr 1 were sown on the 12 and 14 November in the 2018/2019 and 2019/2020 seasons, respectively. In total, 35 kg P2O5 ha−1 in the form of calcium super phosphate (15.5% P2O5) was added during soil preparation. At the same time, 215 kg nitrogen ha−1 as urea (46% N) was added before irrigation in three recommended doses.
The treatments were as follows: AA (0, 100, 200 ppm), SA (0, 100, 200 ppm) and combined treatment of 100 ppm AA plus 100 ppm SA. Those treatments were applied as seed priming and foliar spray application. Priming treatments were achieved by soaking wheat seeds for 8 h and then air dried before sowing. The foliar application was undertaken 35 and 50 days after sowing (DAS).
All other cultural practices were achieved as recommended by Egyptian Field Crops Research Institute for wheat cultivation in the region.
Growth characters were measured, and samples of six average plants were taken, at 65, 80, and 95 DAS, from each plot at random, and at the same time, other plant samples were taken and dried at 70 °C in the oven to a constant weight. According to Hunt [37] formulas, leaf area index (LAI), net assimilation rate (NAR), and crop growth rate (CGR) were calculated:
where:
LAI = leaf area of plant (cm2)/land area occupied by plant (cm2).
CGR (g m−2 day−1) = (W2 − W1)/(t2 − t1).
NAR (g m−2 day−1) = (W2 − W1)(loge A2 − loge A1)/(A2 − A1)(t2 − t1).
A2 − A1= the difference in leaf area (cm2) between two taken samples.
W2 − W1= the difference between two samples in accumulated dry weight (g) of whole plants.
t2 − t1= number of days between two consecutive samples (day).
Loge = natural logarithm.
Photosynthetic pigments (chl a, chl b, and carotenoids) content of leaves were estimated at 80 DAS, according to Lichtenthaler and Buschmann [38].
2.1. Assay of Antioxidant Enzymatic Activities
Fresh wheat leaf samples were homogenized in a chilled mortar along with liquid nitrogen. Next, 0.1 mM disodium EDTA and 0.1 g polyvinylpyrrolidone were mixed with 5 mL of 100 mM potassium–phosphate (K-P) buffer (pH 7.0). Then, the samples were filtered, and the enzyme assays were achieved from the supernatants. Determination of superoxide dismutase activity was performed according to Scebba et al. [39]. A 2 mL leaf extract was mixed with a 3 mL solution mixture consisting of 50 mM K-P buffer (pH 7.8), 13 mM L-methionine, 0.1 mM EDTA, 75 µM nitroblue tetrazolium, and 2 µM riboflavin. The reaction was initiated by exposing the mixtures to fluorescent light (cool white) for 15 min, and the resulting blue reaction color was spectrophotometrically determined at 560 nm. Determining catalase activity was performed according to Aebi [40]. The, 3 mL of the reaction solution containing leaf extract, 50 mM K-P buffer (pH 7.0), and 30% H2O2 (w/v), was used for assaying CAT activity. The changes in the absorbance of H2O2 at 240 nm was measured as the activity of the CAT enzyme. Determination of peroxidase enzymatic activity was achieved using guaiacol according to Maehly and Chance [41] by adding 0.5 mL leaf extract to 3 mL reaction mixture containing 10 mM K-P buffer (pH 7.0), 10 mM H2O2, and 20 mM guaiacol. As a result of tetraguaiacol production, the increase in absorbance was determined at 470 nm. Determination of APX activity was achieved according to method of Chen and Asada [42].
2.2. Soluble Sugar Determination
Soluble sugar content in leaves was determined following the procedures modified by Yemm and Willis [43]. Extraction of soluble sugars occurred by dipping in 10 mL 80% ethanol (v/v) at 25 °C overnight with periodical shaking. Then, 3.0 mL of anthrone reagent (150 mg anthrone + 100 mL 72% H2SO4 (v/v)) freshly prepared as reacting reagent was added to 0.1 mL of the extract in a boiling water bath for 10 min, and the contents of the soluble sugars analyzed in leaf extracts were determined as glucose equivalent.
2.3. Determination of Lipid Peroxidation
Malondialdehyde (MDA) content in wheat leaves in the form of lipid peroxidation level was assayed according to the standard method of Heath and Packer [44]. The absorption of the supernatant was determined at 532 and 600 nm wavelengths, respectively. Using an absorption coefficient of 155 mM−1 cm−1, the MDA content was calculated.
2.4. Determination of Glycine Betaine, Total Phenols, and Proline
Glycine betaine (GB) content in wheat plant leaves was determined according to Grieve and Grattan [45]. Next, 125 mg of dried finely powdered leaf material was mechanically shaken in 5 mL of distilled water at 25 °C for 24 h. The filtrates of the samples were diluted in 1:1 ratio with 2 N HCl. Then, the mixed solution was cooled for 1 h on ice with interrupted vortexing; after that, 0.2 mL of cold potassium iodine reagent was added, and the mixture was softly vortexed. Samples were stored at 4 °C for 16 h and centrifuged for 15 min at 12,000× g. One mL micropipette was used to separate the supernatant. After that, the crystals of the peridotite complex were dissolved in 9 mL reagent grade 1, 2-dichloro ethane. The absorbance was measured after 2 h at 365 nm.
The Folin–Ciocalteu method was achieved to determine the total phenolic contents in the plant leaf extract according to Kaur and Kapoor [46]. Next, 0.2 mL of plant extract samples was completed to 3 mL with distilled water, then mixed with 0.5 mL of Folin–Ciocalteu reagent for 3 min. Then, 2 mL of 20% (w/v) sodium carbonate was added. After one hour of allowing the mixture to stand in the dark, absorbance was measured at 650 nm.
The content of proline in the wheat leaves was assayed according to Bates et al. [47]. Then, 0.25 g fresh leaves were homogenized in 2.5 mL of 3% sulfosalicylic acid and centrifugated for 10 min at 10,000× g. Next, 2.5 mL supernatant was relocated to test tubes containing 2.5 mL sulfosalicylic acid and 1 mL each of acid–ninhydrin solution and glacial acetic acid and incubated at 100 °C for 1 h. Then, 2 mL toluene was added to the incubated mixture to terminate the reaction with continuous stirring for 15–20 s. Chromophore containing toluene layer was separated, and absorbance was determined at 528 nm.
2.5. Determination of Na+ and K+ Ion Content
Sodium and potassium ion content in wheat plant were determined as mmole kg−1 dry weight, according to Allen et al. [48].
2.6. Determination of Yield and Yield Attributes
At harvesting time, plant height (cm), spike length (cm) and spike weight (g), 1000-grain weight (g), grain and straw yields (t ha−1) as well as harvest index were determined.
2.7. Determination of Nitrogen and Protein Content in Wheat Seeds
Using Micro-Kjeldahl method, nitrogen content in wheat seeds was determined following Bremner and Mulvaney [49]. The values of nitrogen content in the wheat seeds were determined from the standard curve and expressed as a percentage of dry weight according to the methods of Thaltooth et al. [50] and Yagoob and Yagoob [51].
Protein content percentage = total nitrogen (%) × 6.25
2.8. Statistical Analysis
Statistical analysis of variance was performed for taken data of the two seasons according to Steel and Torrie [52]. The average for treatments was compared using Fisher’s least significant difference (LSD) test at 0.05 level of significance.
3. Results
3.1. Growth Parameters
The foliar spraying and priming with AA and SA significantly increased the height of the wheat plant, crop growth rate, and net assimilation rate compared with the control (Table 3). In general, priming with AA and SA was more significant than foliar spraying on plant height, CGR, and NAR for the two studied growth periods. However, the highest values of plant height, CGR, and NAR were observed as a result of priming wheat seeds with the combined treatment (100 ppm) in both growing seasons. The increase in plant height due to priming with 100 ppm AA and 100 ppm SA was 5% in the two seasons compared to priming with water. At the same time, the CGR increased as a result of the same treatment by 16% for the first period and 13% for the second period in the two seasons. In addition, the increase in NAR was 15% for the first period and 14% for the second period.
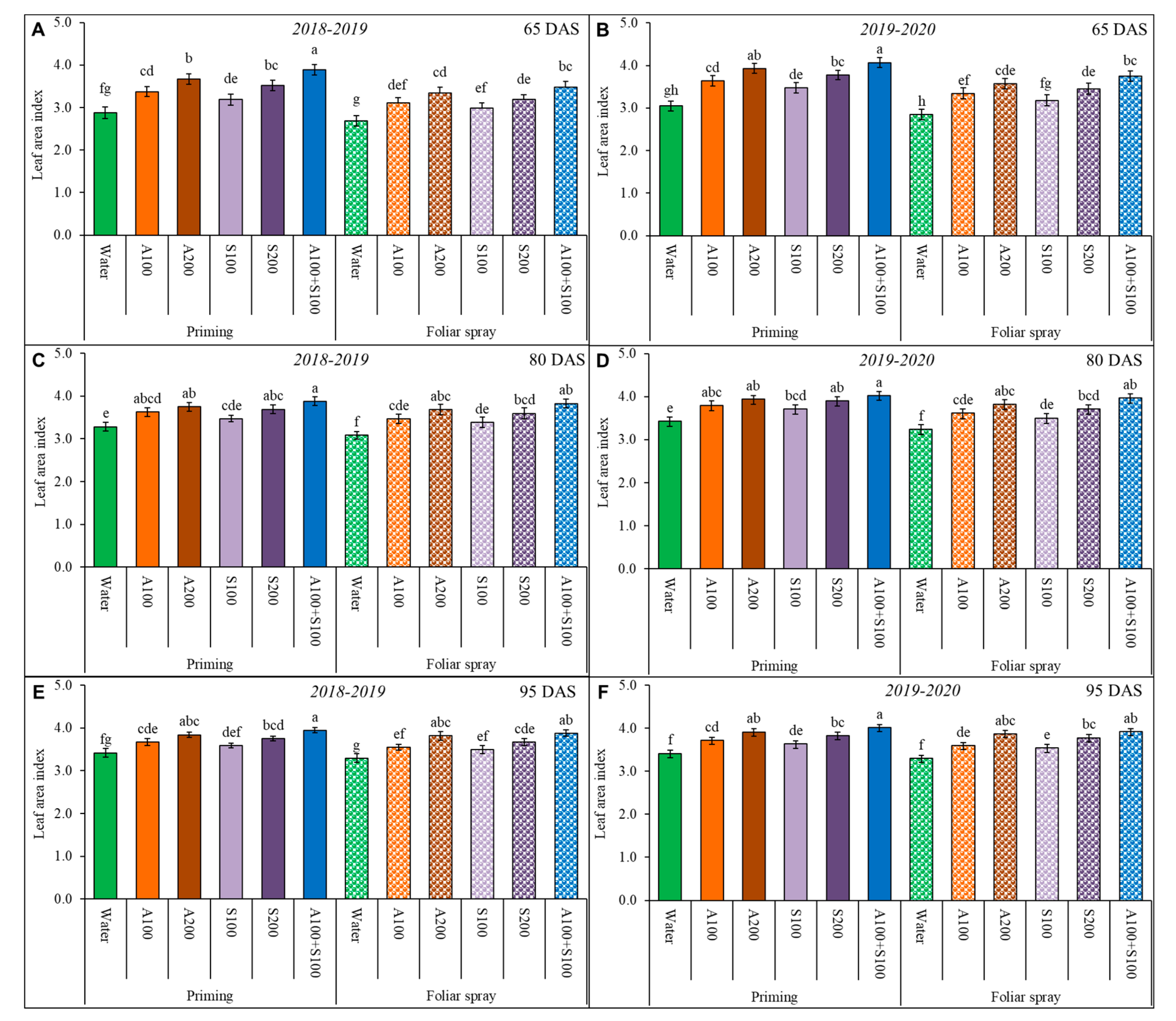
The leaf area index of the wheat plants was significantly affected under the saline condition in the two seasons during the three periods of vegetative growth (Figure 1). However, the LAI of wheat plants treated with AA and SA as a priming application was more significant than that treated with the foliar application, especially in the first period at 65 DAS, in the two studied seasons compared to control treatments. The most significant LAI was achieved in priming wheat seeds with 100 ppm AA and SA followed by priming with 200 ppm AA in the three periods and the two seasons.
3.2. Photosynthetic Pigments
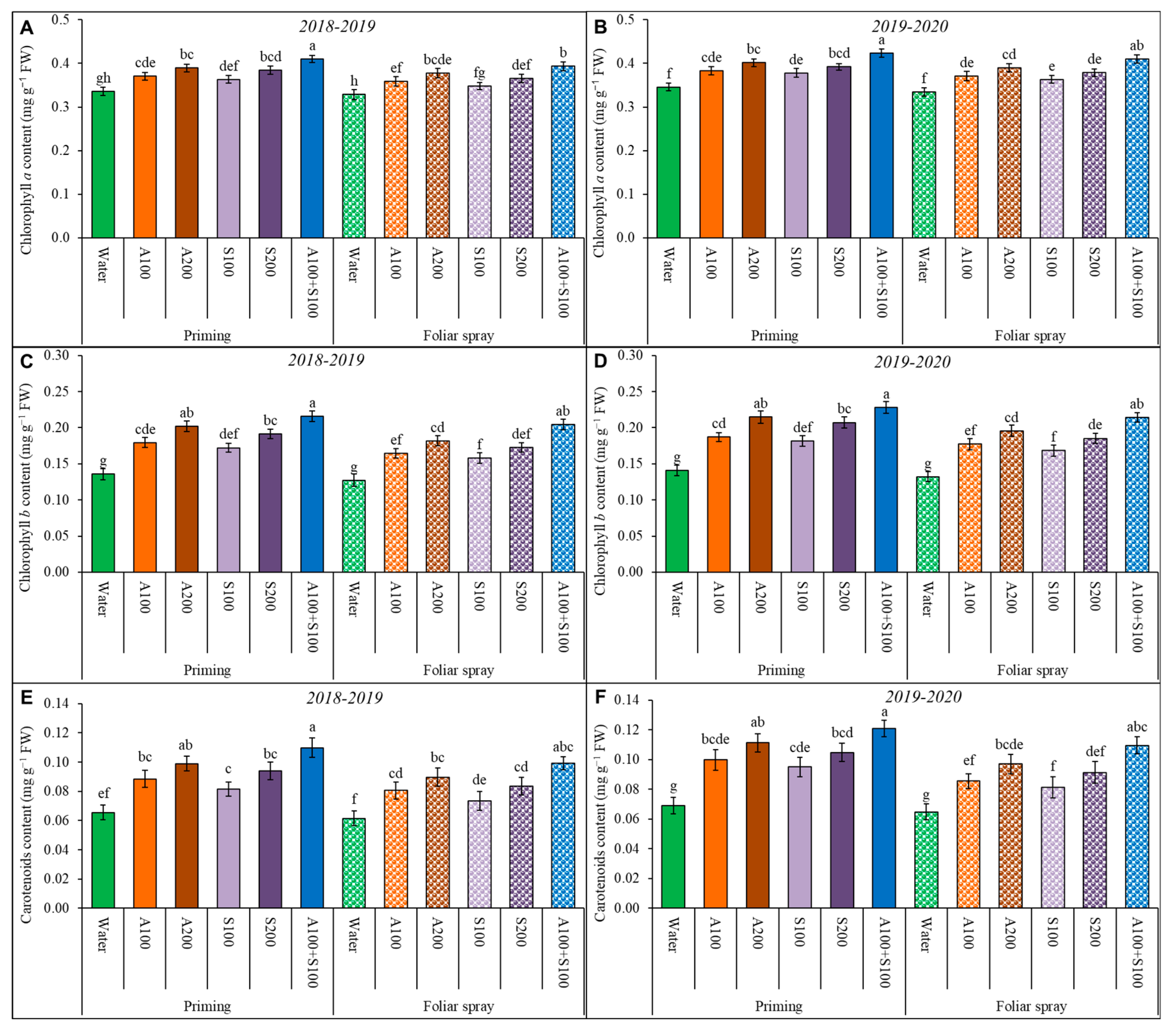
Photosynthetic pigments (chl a, chl b, and carotenoids) in wheat leaves were significantly affected under the saline conditions at 80 DAS (Figure 2). The contents of chl a, chl b, and carotenoids were significantly increased in treated plants (priming and foliar spray) compared to control treatments. The highest significant values of chl a, chl b, and carotenoids were achieved as a result of priming wheat seeds with 100 ppm AA and SA, followed by foliar spray treatment with 100 ppm AA and SA.
3.3. Antioxidant Enzyme Activities
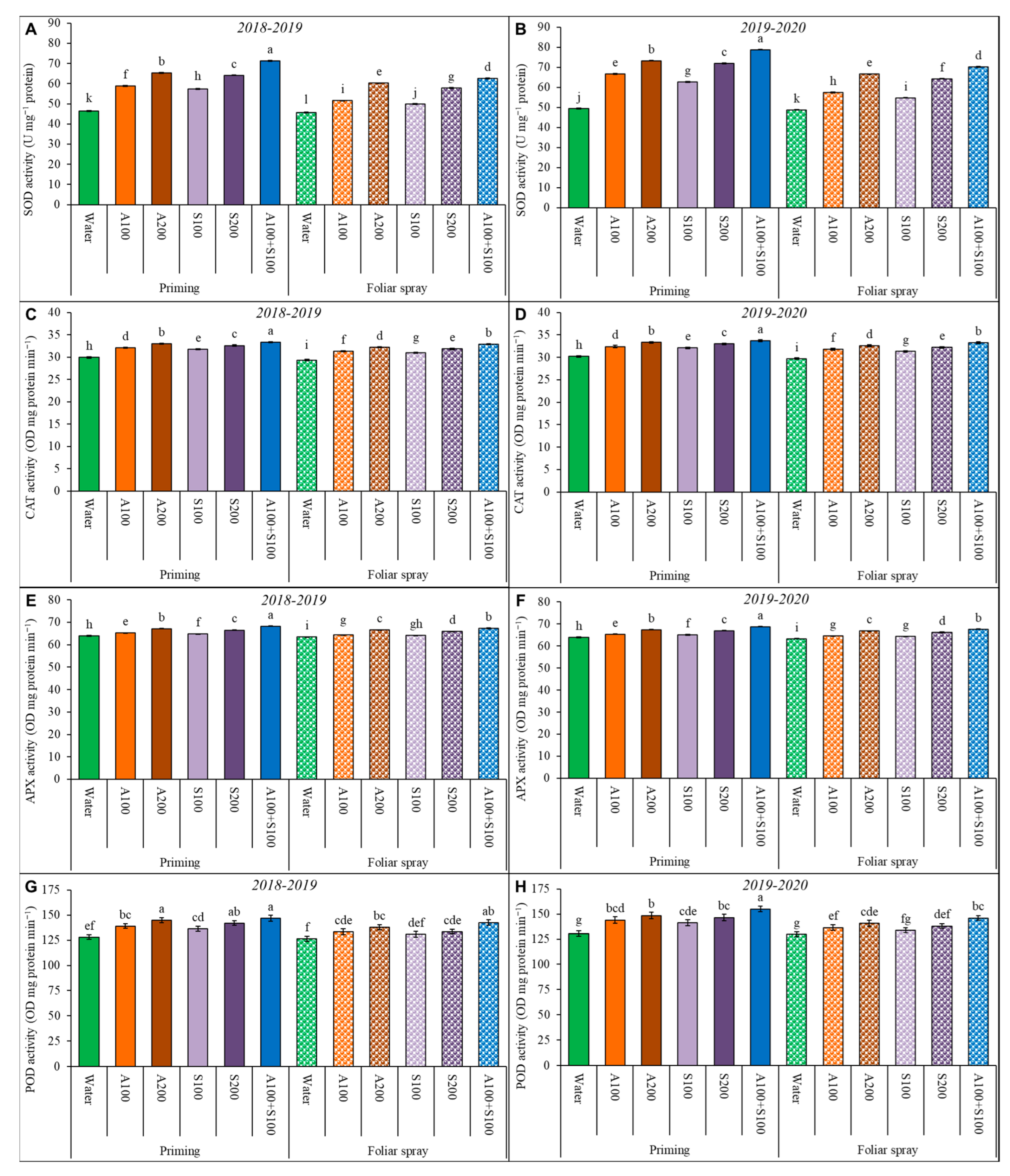
Antioxidant enzymes SOD, CAT, APX, and POD activities in untreated plants were significantly decreased under saline conditions (Figure 3). Conversely, the antioxidant enzyme activities in the exogenous application treatments of AA and/or SA as priming or a foliar spray significantly increased the antioxidant enzyme activities under saline conditions in the two growing seasons. The highest activity of enzymes was achieved by the combined treatment of AA and SA as priming with 100 ppm. It significantly increased SOD by 59%; CAT by 12%; APX by 8%; and POD by 19%, compared to the control.
3.4. Soluble Sugar Content
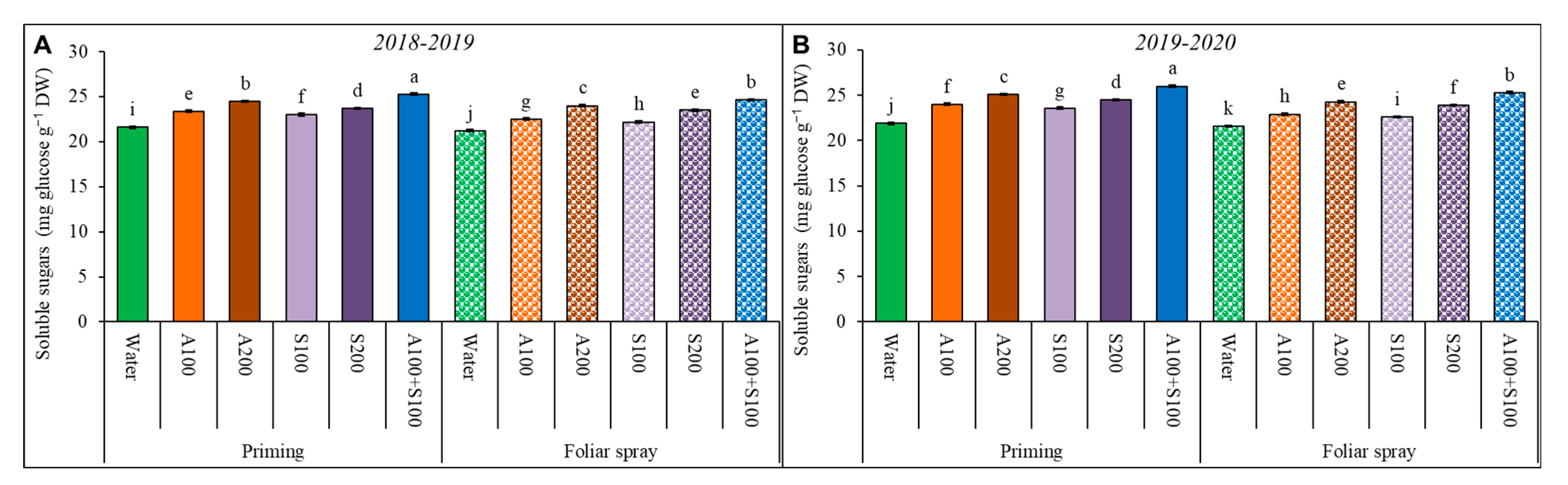
The total soluble sugars content in wheat plant leaves increased under salt-affected soil conditions. The content increased further upon foliar and priming application of AA and SA in the two seasons (Figure 4). The increase in soluble sugar content was positively correlated with increasing the concentration of the AS and SA treatments. However, priming treatments produced more soluble sugars than foliar treatments. The priming with combined 100 ppm AA and SA treatment achieved the highest significant values of the soluble sugars compared to control treatments. This treatment increased the soluble sugar level significantly by 19% compared to the control.
3.5. Lipid Peroxidation
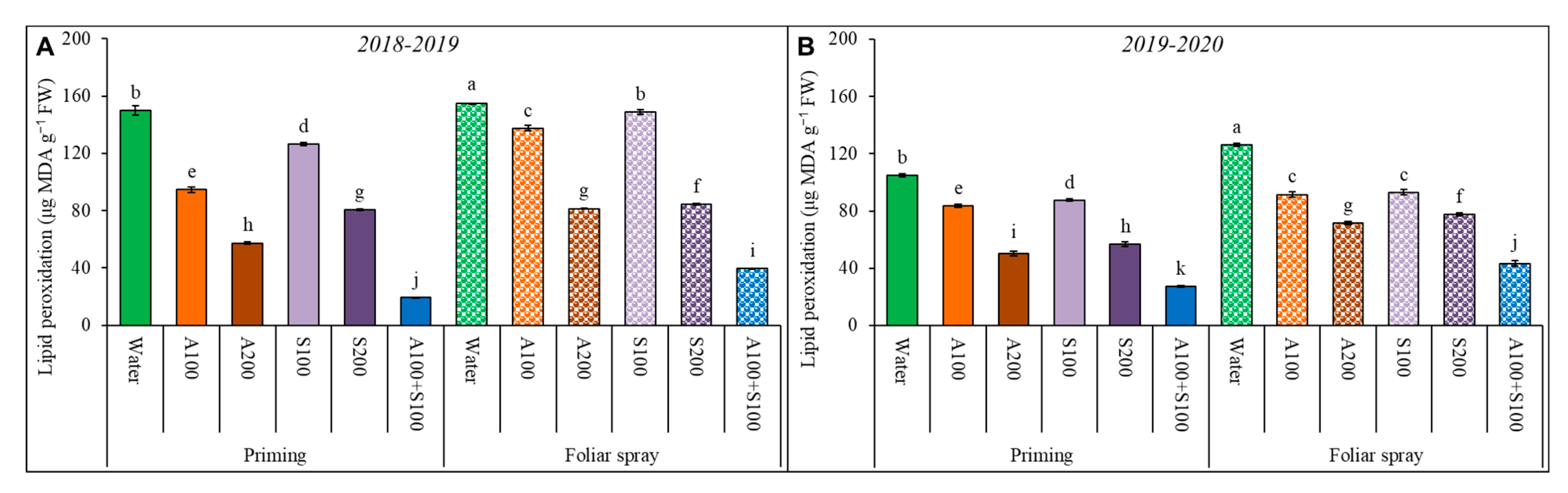
Lipid peroxidation in wheat plant leaves increased under salt-affected soil conditions and decreased due to the applications of AA and SA as priming and/or foliar in the two seasons (Figure 5). The decrease in lipid peroxidation with priming treatments was higher than with foliar treatments in the two seasons. The highest decrease in lipid peroxidation was achieved with the combined treatment of 100 ppm AA and 100 ppm SA as priming, then followed by the foliar spray with 100 ppm AA and 100 ppm SA compared with the control. The combined treatment of 100 ppm AA and 100 ppm SA as priming significantly decreased lipid peroxidation in the two successive seasons.
3.6. Glycine Betaine, Total Phenols, and Proline
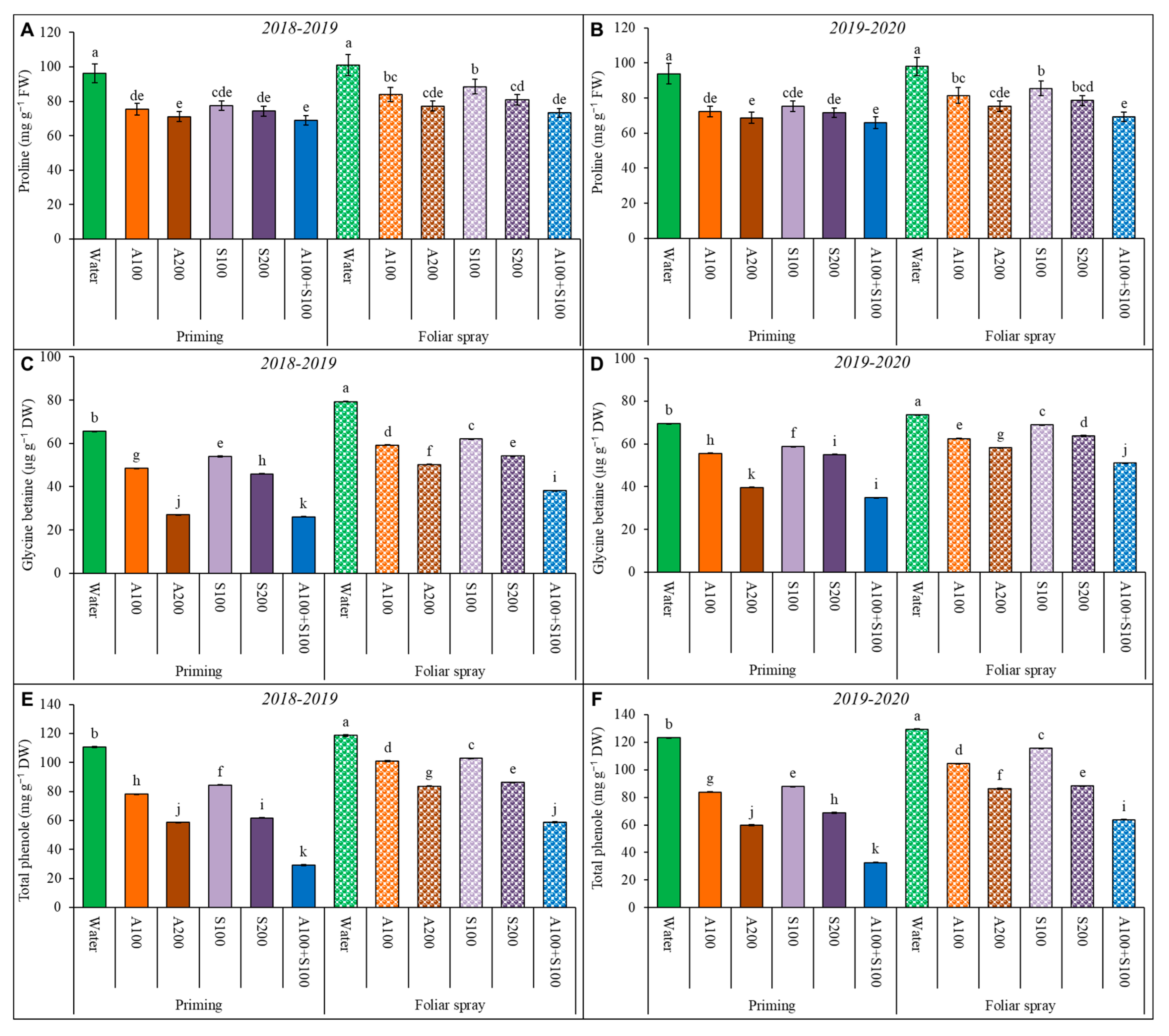
Glycine betaine, total phenols, and proline concentrations in wheat leaves increased under salt-affected soil conditions in the two seasons (Figure 6). Applying AA and SA as priming or a foliar spray decreased glycine betaine, total phenols, and the proline content in wheat plant leaves. The reduction rate increases with a higher concentration of AA and SA. The combined priming treatment resulted in the lowest values among all treatments. It significantly lowered the glycine betaine level by 60%, total phenols by 74%, and proline by 30%, compared with the control.
3.7. Ion Homeostasis
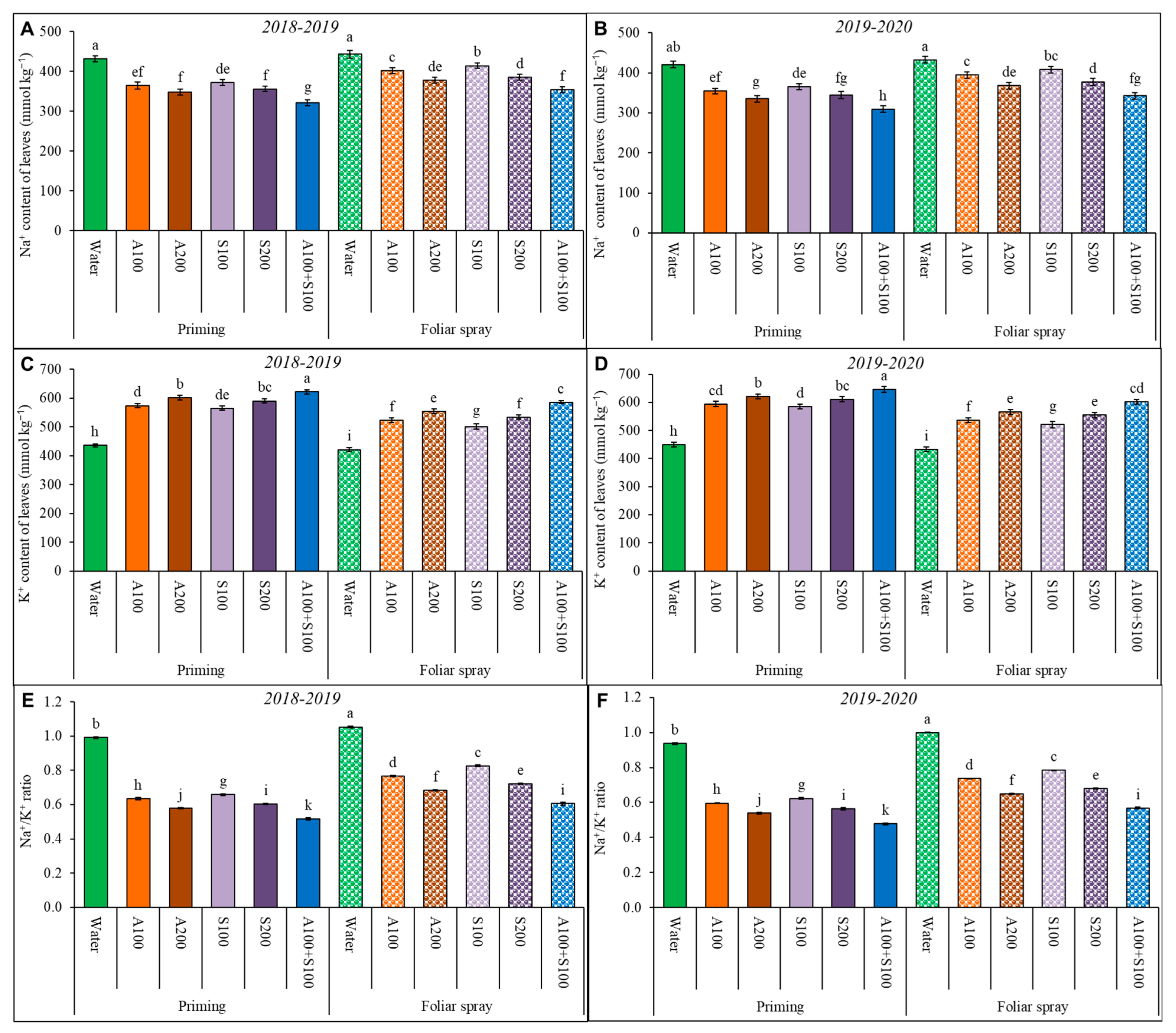
The sodium and potassium content, as well as the Na+/K+ ratio, in the wheat plants were affected by salt stress, as shown in Figure 7. However, priming or a foliar spray of AA and SA decreased the Na+ content and Na+/K+ ratio in wheat leaves. The reduction rate increases with a higher concentration of AA and SA. The combined AA and SA priming application resulted in the lowest values among all treatments. It significantly lowered the Na+ content by 27% and Na+/K+ ratio by 49% compared to the control. At the same time, the application of AA and SA increased the K+ content in wheat leaves. The rate of enhancement is significantly triggered by a higher concentration of AA and SA. The 100 ppm AA and 100 ppm SA combined priming treatment resulted in the highest values for the K+ content among all treatments. It significantly increased the K+ content level by 44% compared to the control.
3.8. Yield and Yield Components
The wheat yield and yield attributes were significantly affected by salt stress under salt-affected soil conditions in the two growing seasons (Table 4). However, applying antioxidants increased the spike length, spike weight, 1000-grain weight, grain yield, straw yield, and harvest index in wheat. Those increases increased with increasing the concentration of antioxidants compared with untreated plants in the two growing seasons. The priming treatments gave values for yield and yield components higher than the foliar spray treatments. At the same time, the application of the 100 ppm AA and 100 ppm SA combined priming treatment resulted in the significantly highest values for yield. Then, in second came the priming with 200 ppm AA and combined foliar spray with 100 ppm AA and SA compared with the control.
3.9. Protein Content in Seeds
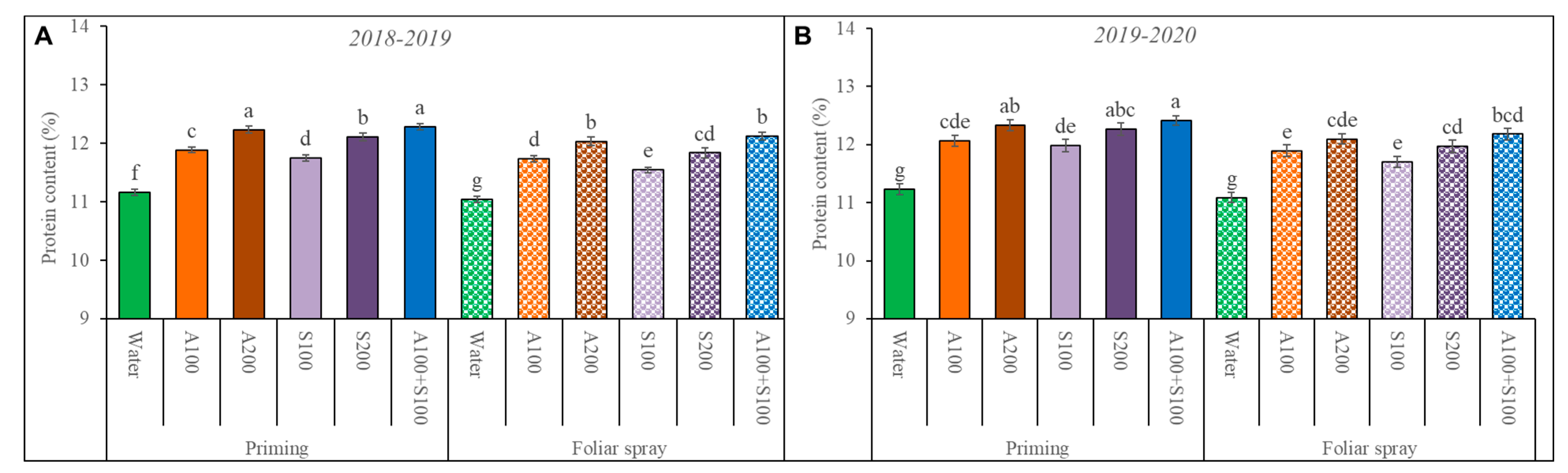
As shown in Figure 8, the wheat treated with AA and SA increased the protein content in seeds. A higher concentration of AA and SA directly resulted in a higher level of protein content in the seeds. Furthermore, the combined priming with 100 ppm AA and SA treatment resulted in the highest values for protein content in seeds among all treatments. It significantly increased the protein content level by 11% in the two seasons compared to the control.
3.10. Correlation of Grain Yield with Growth, Physiological Parameters, and Yield Attributes
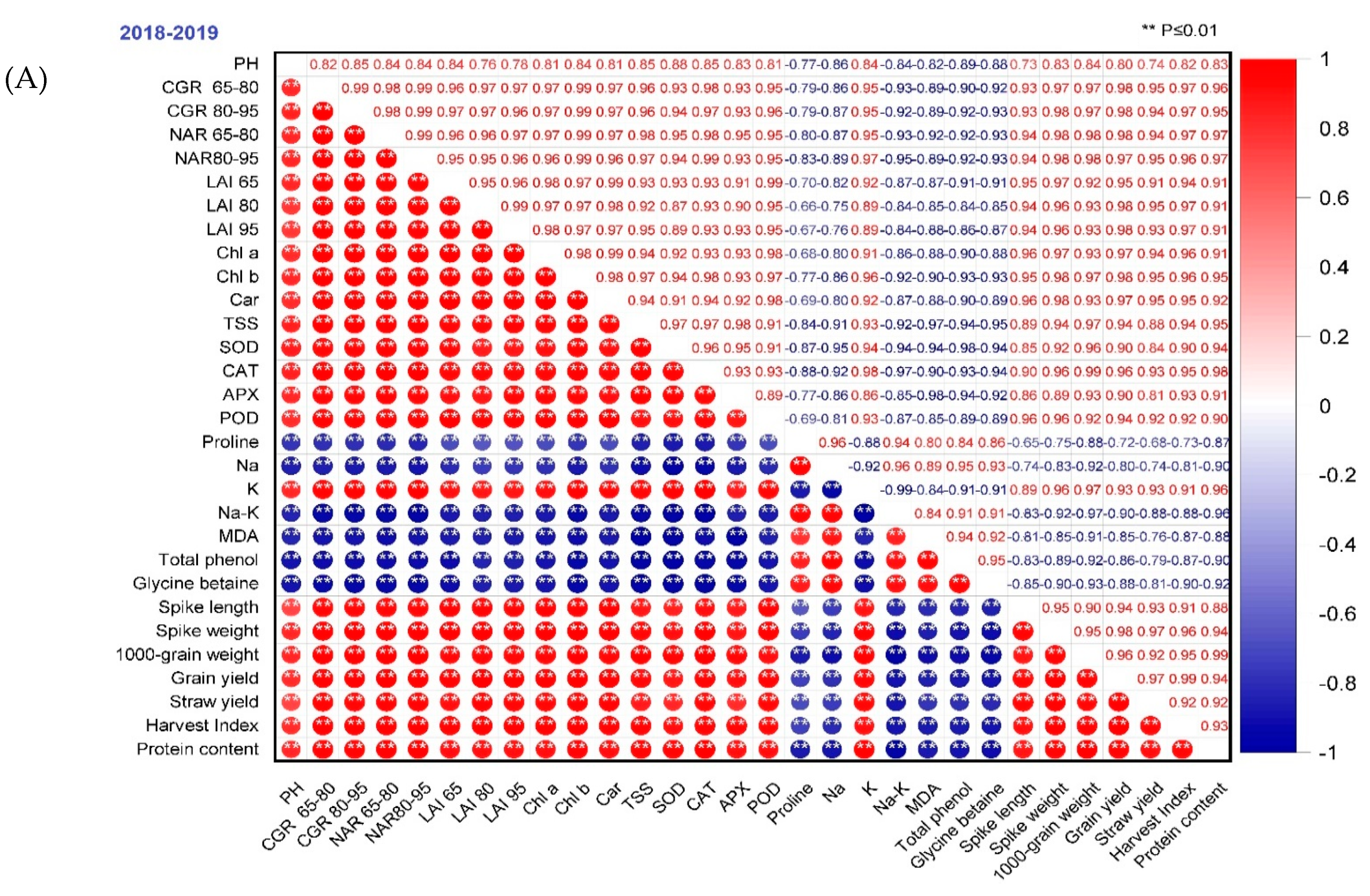
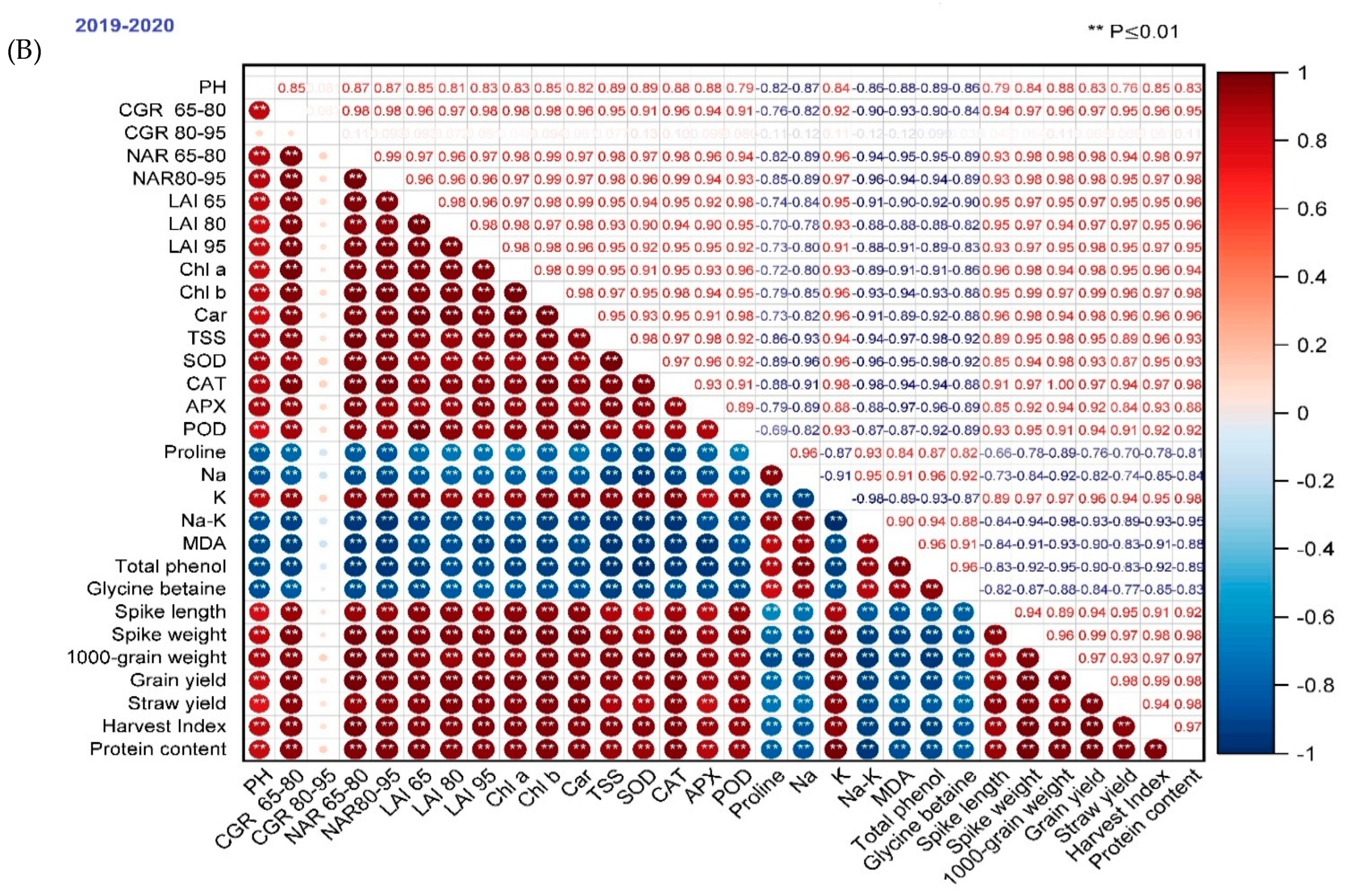
The association between the two traits represents the performance of treatments under such situations for which the data were recorded. PH showed a significant relationship with CGR, LAI, NAR, chl a, chl b, carotenoids, TSS, SOD, CAT, APX, POD, K, spike length, spike weight, 1000-grain weight, grain yield, straw yield, and the harvest index as well as protein content. Other traits exhibited a significant negative correlation in the two seasons, except that those parameters exhibited non-significant relationships in the 2019/2020 season (Figure 9). The results indicated that the yield increased with higher PH, LAI, CGR, NAR, chl a, chl b, carotenoids, TSS, SOD, CAT, POD, APX, and decreasing proline, MDA, total phenols, glycine betaine, Na and Na/K ratio. The studied parameters showed more significant positive and negative correlations in the 2019/2020 season than in 2018/2019.
4. Discussion
Salinity stress caused a significant reduction in the LAI, CGR, and NAR of wheat plants under saline soil conditions, as shown in the results in the two seasons. The priming and foliar application of AA and SA enhanced the LAI, CGR and NAR, especially AA and SA priming. The AA and SA priming improved dehydrogenase and α–amylase activities compared to the control treatment. The α–amylase enzyme in seeds hydrolyses starch to maltodextrin, sucrose, glucose, etc., to supply metabolites and energy to initiate seed germination and activate the growth of seedlings [53]. Moreover, AA and SA priming lowers the stress effect on seeds, which helps them to overcome adverse abiotic stress at the germination phase [54]. Subsequently, the increased tillers count leads to more vigorous seedlings at the beginning of the wheat plant growth phase [55,56]. At the same time, the foliar application of AA and SA mitigates the salinity effect by stimulating growth parameters that subsequently lead to a significant increase in the LAI, CGR, and NAR. The enhancement of the plant growth parameters reflects the effect of AA and SA signal molecules responsible for stimulating tolerance in plants to abiotic stress. The improvement of plant growth may be due to the inhibition of Na+ and Cl- harmful effects in the saline condition. In addition, it may be due to triggering the antioxidant machinery and its effect on membrane permeability and lipid peroxidation. Furthermore, it may prevent the lowering of cytokinin and IAA content, which subsequently reduces stress-induced wheat growth inhibition [57,58].
According to our study, photosynthetic pigments were reduced under saline soil conditions. The reduction in photosynthetic pigments may be due to the minimized absorption rate of light required for the photosynthesis process [59]. The application of AA and SA reduces the deleterious effect of salinity by increasing and upregulating the synthesis of photosynthetic pigments. However, photosynthetic pigment content is considered a significant parameter for crop salt tolerance [60]. The application of AA and SA improved photosynthetic pigments by inhibiting the chlorophyllase effect in plants [61]. Exogenous AA and SA application is predictable to control stomatal opening under stress. Then, it subsequently lowers the transpiration, maintains turgor, and improves the growth and productivity of plants under stress conditions [62,63,64]. The AA and SA reduce the salinity harmful effect by elevating the antioxidant machinery and decreasing the ROS. This, in turn, stimulates cell division and elongation, prevents chlorophyll breakdown, and enhances photosynthesis by inhibiting chlorophyll oxidase enzymes [65,66].
To overcome ROS’s harmful effect on susceptible macromolecules, plants increase the antioxidant enzyme activities for mediating the fast scavenging of toxic ROS. Increasing antioxidant activities overcomes the toxic levels of ROS [67]. The antioxidant enzymes SOD, CAT, POD, and APX are involved in H2O2 detoxification. In the current study, those enzymes were decreased under salinity stress, and this may be due to the increasing ROS-mediated oxidative stress and cell injury in wheat plants. The application AA and SA as priming and/or foliar spray increased the activities of those enzymes. The suggested mechanism of those enzymes is to scavenge H2O2 by converting it into H2O and O2 and cease its toxicity [4].
Under soil saline conditions, the glycine betaine, total phenols, and proline contents increase, and those increases have an important role to overcome the undesirable effect of salinity stress and ameliorate plant growth [68]. Proline, a water-soluble amino acid and useful solute, is considered an osmoprotectant and preserves the pressure of cell turgor [69]. However, the application of AA and SA under the conditions of this study lowers the undesirable effect of salinity on wheat plant growth and decreases the glycine betaine, total phenols, and proline accumulations. These results are supported by other studies where AA and SA reduced total phenols under salinity stress in wheat [57], as well as the proline content under salinity stress in tomatoes [70], sweet peppers [71], and soybeans [72].
In this study, the results show that under salt-affected soil, the K+ content decreased, and the Na+ content increased in the wheat leaves. This may be due to an ionic imbalance [73] as a result of the cells’ plasma membrane disorder in the integrity as well as a negative correlation among the sodium elements and other nutrients [74,75]. Moreover, AA and SA applications affect many fundamental physiological actions in plants, such as rising nutrient uptake and lowering Na+ contents under salinity stress [76,77]. In addition, AA and SA applications switched the selectivity of the Na+ and K+ uptake and lowered the Na+/K+ ratio, which protected the cell membrane from harm [78].
The decreases in yield and its components under saline soil conditions may be a result of the reduction in photoassimilation assembly, as well as photoassimilation motivation, which achieved the lowest values in the harvest index [79]. Furthermore, the yield reduction may be a result of the reverse effect of salinity stress on plant growth parameters and physiological processes such as photosynthesis, water absorbance, and grain filling [80]. Nevertheless, in this study, AA and SA improved tolerance to salinity in wheat plants and promoted yield, especially, with combined treatments. The function of AA and SA on yield is maybe due to their important role in tolerance to stress. In general, AA and SA decrease oxidative stress and increase plant growth and yield under salt stress as described earlier by Li et al. [81], Hafez and Gharib [82], El-Hawary and Nashed [22], and El-Hawary et al. [83].
The protein content in wheat seeds was affected by salinity, and this may be due to the inhibitory effect on plant growth and its chemical contents. Those variations were reflected in productivity and its components as well as the protein percentage content in seeds. The application of AA and SA, especially with the combined treatments, significantly increased the protein content in wheat seeds. However, it can be concluded that AA and SA enhance protein synthesis and delay senescence [84].
5. Conclusions
As meteorological conditions during vegetation growth influenced plant growth and grain filling in the first season as the temperature was high, the study represented the important role of ascorbic and salicylic acids in intermediating the reverse effects of salt stress on growth, yield, and physiological traits. Priming and foliar spray application of AA and SA significantly ameliorated the growth and yield of wheat under saline soil conditions by stimulating stress tolerance and increasing the activity of plant enzymes. Moreover, based on the present findings, it can be concluded that ascorbic acid at 100 ppm with salicylic acid at 100 ppm as priming or a foliar spray could be recommended for obtaining better growth and yield of wheat plants under salt-affected soil conditions. However, for more application of ascorbic and salicylic acids to different wheat genotypes under salt-affected soil conditions, more studies should be undertaken to optimize the application. Information in this regard would also help wheat producers, as well as in breeding programs, to select the appropriate wheat cultivars that can be successfully exploited under salt-affected soil. The estimation of ascorbic and salicylic acid application on wheat in different growth stages of wheat could provide the possibility of selecting salt tolerant wheat genotypes at early growth stages.
Author Contributions
Conceptualization, M.M.E.-H. and M.H.; methodology, O.S.M.H.; formal analysis, M.H.; investigation, M.M.E.-H. and O.S.M.H.; writing—original draft preparation, M.M.E.-H. and O.S.M.H.; writing—review and editing, M.H.; visualization, M.M.E.-H. and M.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
All data are available in this manuscript.
Acknowledgments
We acknowledge Md. Rakib Hossain Raihan for the critical check and formatting.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Munns, R.; Gilliham, M. Salinity Tolerance of Crops—What Is the Cost? New Phytol. 2015, 208, 668–673. [Google Scholar] [CrossRef] [PubMed]
- Kopittke, P.M.; Menzies, N.W.; Wang, P.; McKenna, B.A.; Lombi, E. Soil and the Intensification of Agriculture for Global Food Security. Environ. Int. 2019, 132, 105078. [Google Scholar] [CrossRef] [PubMed]
- Alnusairi, G.S.H.; Mazrou, Y.S.A.; Qari, S.H.; Elkelish, A.A.; Soliman, M.H.; Eweis, M.; Abdelaal, K.; El-Samad, G.A.; Ibrahim, M.F.M.; ElNahhas, N. Exogenous Nitric Oxide Reinforces Photosynthetic Efficiency, Osmolyte, Mineral Uptake, Antioxidant, Expression of Stress-Responsive Genes and Ameliorates the Effects of Salinity Stress in Wheat. Plants 2021, 10, 1693. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive Oxygen Species and Antioxidant Defense in Plants under Abiotic Stress: Revisiting the Crucial Role of a Universal Defense Regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef] [PubMed]
- Taha, R.S.; Seleiman, M.F.; Alhammad, B.A.; Alkahtani, J.; Alwahibi, M.S.; Mahdi, A.H.A. Activated Yeast Extract Enhances Growth, Anatomical Structure, and Productivity of Lupinus termis L. Plants under Actual Salinity Conditions. Agronomy 2021, 11, 74. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Kheir, A.M.S. Saline Soil Properties, Quality and Productivity of Wheat Grown with Bagasse Ash and Thiourea in Different Climatic Zones. Chemosphere 2018, 193, 538–546. [Google Scholar] [CrossRef]
- Hussain, S.; Hussain, S.; Ali, B.; Ren, X.; Chen, X.; Li, Q.; Saqib, M.; Ahmad, N. Recent Progress in Understanding Salinity Tolerance in Plants: Story of Na+/K+ Balance and Beyond. Plant Physiol. Biochem. 2021, 160, 239–256. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Raihan, M.R.H.; Masud, A.A.C.; Rahman, K.; Nowroz, F.; Rahman, M.; Nahar, K.; Fujita, M. Regulation of Reactive Oxygen Species and Antioxidant Defense in Plants under Salinity. Int. J. Mol. Sci. 2021, 22, 9326. [Google Scholar] [CrossRef]
- Giraldo, P.; Benavente, E.; Manzano-Agugliaro, F.; Gimenez, E. Worldwide Research Trends on Wheat and Barley: A Bibliometric Comparative Analysis. Agronomy 2019, 9, 352. [Google Scholar] [CrossRef]
- Iqbal, M.A.; Rahim, J.; Naeem, W.; Hassan, S.; Khattab, Y.; El-Sabagh, A. Rainfed Winter Wheat (Triticum aestivum L.) Cultivars Respond Differently to Integrated Fertilization in Pakistan. Fresenius Environ. Bull. 2021, 30, 3115–3121. [Google Scholar]
- Kizilgeci, F.; Yildirim, M.; Islam, M.S.; Ratnasekera, D.; Iqbal, M.A.; Sabagh, A.E. Normalized Difference Vegetation Index and Chlorophyll Content for Precision Nitrogen Management in Durum Wheat Cultivars under Semi-Arid Conditions. Sustainability 2021, 13, 3725. [Google Scholar] [CrossRef]
- Pessoa, L.G.M.; Freire, M.D.S.; de Araújo Filho, J.C.; dos Santos, P.R.; Miranda, M.F.A.; Freire, F.J. Characterization and Classification of Halomorphic Soils in the Semiarid Region of Northeastern Brazil. J. Agric. Sci. 2019, 11, 405. [Google Scholar] [CrossRef]
- Ljubičić, N.; Popović, V.; Ćirić, V.; Kostić, M.; Ivošević, B.; Popović, D.; Pandžić, M.; El Musafah, S.; Janković, S. Multivariate Interaction Analysis of Winter Wheat Grown in Environment of Limited Soil Conditions. Plants 2021, 10, 604. [Google Scholar] [CrossRef]
- Hernández, J.A. Salinity Tolerance in Plants: Trends and Perspectives. Int. J. Mol. Sci. 2019, 20, 2408. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Huang, R. Modulation of Ethylene and Ascorbic Acid on Reactive Oxygen Species Scavenging in Plant Salt Response. Front. Plant Sci. 2019, 10, 319. [Google Scholar]
- Yu, Z.; Duan, X.; Luo, L.; Dai, S.; Ding, Z.; Xia, G. How Plant Hormones Mediate Salt Stress Responses. Trends Plant Sci. 2020, 25, 1117–1130. [Google Scholar] [CrossRef] [PubMed]
- Godoy, F.; Olivos-Hernández, K.; Stange, C.; Handford, M. Abiotic Stress in Crop Species: Improving Tolerance by Applying Plant Metabolites. Plants 2021, 10, 186. [Google Scholar] [CrossRef] [PubMed]
- Mir, R.A.; Somasundaram, R. Salicylic Acid and Salt Stress Tolerance in Plants: A Review. J. Stress Physiol. Biochem. 2021, 17, 32–50. [Google Scholar]
- Nawaz, M.; Ashraf, M.Y.; Khan, A.; Nawaz, F. Salicylic Acid– and Ascorbic Acid–Induced Salt Tolerance in Mung Bean (Vigna radiata (L.) Wilczek) Accompanied by Oxidative Defense Mechanisms. J. Soil Sci. Plant Nutr. 2021, 21, 2057–2071. [Google Scholar] [CrossRef]
- Abdullah, H.A.; Faysal, M.S.; Al-Rashedy, H.S.M. Role of Ascorbic Acid in Reducing the Harmful Effects of Sodium Chloride in Triticum aestivum L. and Trigonella foenum-Graecum L. Plants. Egyptian J. Bot. 2021, 61, 901–910. [Google Scholar] [CrossRef]
- Ahmad, Z.; Waraich, E.A.; Tariq, R.M.S.; Iqbal, M.A.; Ali, S.; Soufan, W.; Hassan, M.M.; Islam, M.S.; El Sabagh, A. Foliar Applied Salicylic Acid Ameliorates Water and Salt Stress by Improving Gas Exchange and Photosynthetic Pigments in Wheat. Pak. J. Bot 2021, 53, 1553–1560. [Google Scholar] [CrossRef] [PubMed]
- El-Hawary, M.M.; Nashed, M.E. Effect of Foliar Application by Some Antioxidants on Growth and Productivity of Maize under Saline Soil Conditions. J. Plant Prod. Mansoura Univ. 2019, 10, 93–99. [Google Scholar]
- Hasanuzzaman, M.; Nahar, K.; Bhuiyan, T.F.; Anee, T.I.; Inafuku, M.; Oku, H.; Fujita, M. Salicylic Acid: An All-Rounder in Regulating Abiotic Stress Responses in Plants. In Phytohormones–Signaling Mechanisms and Crosstalk in Plant Development and Stress Responses; El-Esawi, M., Ed.; InTech: London, UK, 2017; ISBN 978-953-51-3411-4. [Google Scholar]
- Zandi, P.; Schnug, E. Reactive Oxygen Species, Antioxidant Responses and Implications from a Microbial Modulation Perspective. Biology 2022, 11, 155. [Google Scholar] [CrossRef]
- Akram, N.A.; Shafiq, F.; Ashraf, M. Ascorbic Acid-A Potential Oxidant Scavenger and Its Role in Plant Development and Abiotic Stress Tolerance. Front. Plant Sci. 2017, 8, 613. [Google Scholar] [CrossRef] [PubMed]
- El-Beltagi, H.S.; Mohamed, H.I.; Sofy, M.R. Role of Ascorbic Acid, Glutathione and Proline Applied as Singly or in Sequence Combination in Improving Chickpea Plant through Physiological Change and Antioxidant Defense under Different Levels of Irrigation Intervals. Molecules 2020, 25, 1702. [Google Scholar] [CrossRef] [PubMed]
- Dolatabadian, A.; Modarres Sanavy, S.a.M.; Sharifi, M. Alleviation of Water Deficit Stress Effects by Foliar Application of Ascorbic Acid on Zea mays L. J. Agron. Crop Sci. 2009, 195, 347–355. [Google Scholar] [CrossRef]
- Xu, Y.; Xu, Q.; Huang, B. Ascorbic Acid Mitigation of Water Stress-Inhibition of Root Growth in Association with Oxidative Defense in Tall Fescue (Festuca arundinacea Schreb.). Front. Plant Sci. 2015, 6, 807. [Google Scholar]
- Billah, M.; Rohman, M.M.; Hossain, N.; Uddin, M.S. Exogenous Ascorbic Acid Improved Tolerance in Maize (Zea mays L.) by Increasing Antioxidant Activity under Salinity Stress. AJAR 2017, 12, 1437–1446. [Google Scholar] [CrossRef]
- Wang, Y.-H.; Zhang, G.; Chen, Y.; Gao, J.; Sun, Y.-R.; Sun, M.-F.; Chen, J.-P. Exogenous Application of Gibberellic Acid and Ascorbic Acid Improved Tolerance of Okra Seedlings to NaCl Stress. Acta Physiol. Plant. 2019, 41, 93. [Google Scholar] [CrossRef]
- Mandavia, C.; Cholke, P.; Mandavia, M.K.; Raval, L.; Gursude, A. Influence of Brassinolide and Salicylic Acid on Biochemical Parameters of Wheat (Triticum aestivum L.) under Salinity Stress. Indian J. Agric. Biochem. 2014, 27, 73–76. [Google Scholar]
- Jayakannan, M.; Bose, J.; Babourina, O.; Rengel, Z.; Shabala, S. Salicylic Acid in Plant Salinity Stress Signalling and Tolerance. Plant Growth Regul. 2015, 76, 25–40. [Google Scholar] [CrossRef]
- Yadav, T.; Kumar, A.; Yadav, R.K.; Yadav, G.; Kumar, R.; Kushwaha, M. Salicylic Acid and Thiourea Mitigate the Salinity and Drought Stress on Physiological Traits Governing Yield in Pearl Millet-Wheat. Saudi J. Biol. Sci. 2020, 27, 2010–2017. [Google Scholar] [CrossRef] [PubMed]
- El-Taher, A.M.; Abd El-Raouf, H.S.; Osman, N.A.; Azoz, S.N.; Omar, M.A.; Elkelish, A.; Abd El-Hady, M.A.M. Effect of Salt Stress and Foliar Application of Salicylic Acid on Morphological, Biochemical, Anatomical, and Productivity Characteristics of Cowpea (Vigna unguiculata L.) Plants. Plants 2022, 11, 115. [Google Scholar] [CrossRef]
- Bhanuprakash, K.; Yogeesha, H.S. Seed Priming for Abiotic Stress Tolerance: An Overview. In Abiotic Stress Physiology of Horticultural Crops; Rao, N.K.S., Shivashankara, K.S., Laxman, R.H., Eds.; Springer: New Delhi, India, 2016; pp. 103–117. [Google Scholar]
- Johnson, R.; Puthur, J.T. Seed Priming as a Cost Effective Technique for Developing Plants with Cross Tolerance to Salinity Stress. Plant Physiol. Biochem. 2021, 162, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Hunt, R. Basic Growth Analysis; Springer Netherlands: Dordrecht, 1990. [Google Scholar]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and Carotenoids: Measurement and Characterization by UV-VIS Spectroscopy. Curr. Protoc. Food anal. Chem. 2001, 1, F4.3.1–F4.3.8. [Google Scholar] [CrossRef]
- Scebba, F.; Sebastiani, L.; Vitagliano, C. Protective Enzymes Against Activated Oxygen Species in Wheat (Triticum aestivum L.) Seedlings: Responses to Cold Acclimation. J. Plant Physiol. 1999, 155, 762–768. [Google Scholar] [CrossRef]
- Aebi, H. [13] Catalase in Vitro. In Methods in Enzymology; Oxygen Radicals in Biological Systems; Academic Press: Cambridge, MA, USA, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Maehly, A.C.; Chance, B. The Assay of Catalases and Peroxidases. In Methods of Biochemical Analysis; Glick, D., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2006; pp. 357–424. [Google Scholar]
- Chen, G.-X.; Asada, K. Inactivation of Ascorbate Peroxidase by Thiols Requires Hydrogen Peroxide. Plant Cell Physiol. 1992, 33, 117–123. [Google Scholar] [CrossRef]
- Yemm, E.W.; Willis, A.J. The Respiration of Barley Plants IX. The Metabolism of Roots During the Assimilation of Nitrogen. New Phytol. 1956, 55, 229–252. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in Isolated Chloroplasts: I. Kinetics and Stoichiometry of Fatty Acid Peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Grieve, C.M.; Grattan, S.R. Rapid Assay for Determination of Water Soluble Quaternary Ammonium Compounds. Plant Soil 1983, 70, 303–307. [Google Scholar] [CrossRef]
- Kaur, C.; Kapoor, H.C. Anti-Oxidant Activity and Total Phenolic Content of Some Asian Vegetables. Int. J. Food Sci. 2002, 37, 153–161. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Chemical Analysis of Ecological Materials; Allen, S.E. (Ed.) Blackwell Scientific: Oxford, UK, 1974. [Google Scholar]
- Bremner, J.M. Nitrogen-Total. In Methods of Soil Analysis: Part 3 Chemical Methods; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpour, P.N., Tabatabai, M.A., Johnston, C.T., Sumner, M.E., Eds.; SSSA Book Series; Soil Science Society of America, American Society of Agronomy: Madison, WI, USA, 2018; pp. 1085–1121. [Google Scholar]
- Thalooth, A.T.; Tawfik, M.M.; Mohamed, H.M. A Comparative Study on the Effect of Foliar Application of Zinc, Potassium and Magnesium on Growth, Yield and Some Chemical Constituents of Mungbean Plants Grown under Water Stress Conditions. World J. Agric. Sci. 2006, 2, 37–46. [Google Scholar]
- Yagoob, H.; Yagoob, M. The Effects of Water Deficit Stress on Protein Yield of Mung Bean Genotypes. J. Agric. Sci. 2014, 2, 30–35. [Google Scholar]
- Steel, R.G.D.; Torrie, J.H. Principles and Procedures of Statistics: A Biometrical Approach, 2nd ed.; McGraw-Hill: New York, NY, USA, 1980. [Google Scholar]
- Li, Z.; Xu, J.; Gao, Y.; Wang, C.; Guo, G.; Luo, Y.; Huang, Y.; Hu, W.; Sheteiwy, M.S.; Guan, Y.; et al. The Synergistic Priming Effect of Exogenous Salicylic Acid and H2O2 on Chilling Tolerance Enhancement during Maize (Zea mays L.) Seed Germination. Front. Plant Sci. 2017, 8, 1153. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, K.; Job, C.; Groot, S.P.C.; Puype, M.; Demol, H.; Vandekerckhove, J.; Job, D. Proteomic Analysis of Arabidopsis Seed Germination and Priming. Plant Physiol. 2001, 126, 835–848. [Google Scholar] [CrossRef]
- Kaur, S.; Gupta, A.K.; Kaur, N. Seed Priming Increases Crop Yield Possibly by Modulating Enzymes of Sucrose Metabolism in Chickpea. J. Agron. Crop Sci. 2005, 191, 81–87. [Google Scholar] [CrossRef]
- Farooq, M.; Hussain, M.; Imran, M.; Ahmad, I.; Atif, M.; Alghamdi, S.S. Improving the Productivity and Profitability of Late Sown Chickpea by Seed Priming. Int. J. Plant Prod. 2019, 13, 129–139. [Google Scholar] [CrossRef]
- Desoky, E.-S.M.; Merwad, A.-R.M. Improving the Salinity Tolerance in Wheat Plants Using Salicylic and Ascorbic Acids. J. Agric. Sci. 2015, 7, 203. [Google Scholar] [CrossRef]
- EL-Hadidi, E.; El-Zehery, T.; AL-Wetwat, R. Response of Wheat Plant to Foliar Application of Organic Acids under Saline Conditions. JSSAE 2018, 9, 135–143. [Google Scholar] [CrossRef]
- Pastenes, C.; Pimentel, P.; Lillo, J. Leaf Movements and Photoinhibition in Relation to Water Stress in Field-Grown Beans. J. Exp. Bot. 2005, 56, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, B.; Yaşar, F.; Oezpay, T.; Tuerkoezue, D.; Terziodlu, O.; Tamkoc, A. Variations in Response to Salt Stress among Field Pea Genotypes (Pisum sativum Sp. Arvense L.). J. Anim. Vet. Adv. 2008, 7, 907–910. [Google Scholar]
- Karim, F.M.; Khursheed, M.Q. Effect of Foliar Application of Salicylic Acid on Growth, Yield Components and Chemical Constituents of Wheat (Triticum aestivum L. Var. Cham 6); Tikrit University: Tikrit, Iraq, 2011. [Google Scholar]
- He, Y.; Liu, Y.; Cao, W.; Huai, M.; Xu, B.; Huang, B. Effects of Salicylic Acid on Heat Tolerance Associated with Antioxidant Metabolism in Kentucky Bluegrass. Crop Sci. 2005, 45, 988–995. [Google Scholar] [CrossRef]
- Hasan, M.A.; Al-Taweel, S.K.; Alamrani, H.A.; Al-Naqeeb, M.A.; Al-Baldawwi, M.H.K.; Hamza, J.H. Anatomical and Physiological Traits of Broad Bean (Vicia faba L.) Seedling Affected by Salicylic Acid and Salt Stress. Indian J. Agric. Res. 2018, 52, 368–373. [Google Scholar] [CrossRef]
- El-Beltagi, H.S.; Ahmad, I.; Basit, A.; Shehata, W.F.; Hassan, U.; Shah, S.T.; Haleema, B.; Jalal, A.; Amin, R.; Khalid, M.A.; et al. Ascorbic Acid Enhances Growth and Yield of Sweet Peppers (Capsicum annum) by Mitigating Salinity Stress. Gesunde Pflanz. 2022, 74, 423–433. [Google Scholar] [CrossRef]
- Hayat, Q.; Hayat, S.; Mohd, I.; Ahmad, A. Effect of Exogenous Salicylic Acid under Changing Environment: A Review. Environ. Exp. Bot. 2010, 68, 14–25. [Google Scholar] [CrossRef]
- Kang, G.; Li, G.; Guo, T. Molecular Mechanism of Salicylic Acid-Induced Abiotic Stress Tolerance in Higher Plants. Acta Physiol. Plant 2014, 36, 2287–2297. [Google Scholar] [CrossRef]
- Hashem, A.; Abd_Allah, E.F.; Alqarawi, A.A.; Aldubise, A.; Egamberdieva, D. Arbuscular mycorrhizal Fungi Enhances Salinity Tolerance of Panicum turgidum Forssk by Altering Photosynthetic and Antioxidant Pathways. J. Plant Interact. 2015, 10, 230–242. [Google Scholar] [CrossRef]
- Nathalie, V.; Christian, H. Proline Accumulation in Plants: A Review. Amino Acids 2008, 35, 753–759. [Google Scholar]
- Zulfiqar, F.; Akram, N.A.; Ashraf, M. Osmoprotection in Plants under Abiotic Stresses: New Insights into a Classical Phenomenon. Planta 2020, 251, 3. [Google Scholar] [CrossRef] [Green Version]
- Mimouni, H.; Wasti, S.; Manaa, A.; Gharbi, E.; Chalh, A.; Vandoorne, B.; Lutts, S.; Ahmed, H.B. Does Salicylic Acid (SA) Improve Tolerance to Salt Stress in Plants? A Study of SA Effects on Tomato Plant Growth, Water Dynamics, Photosynthesis, and Biochemical Parameters. OMICS J. Integr. Biol. 2016, 20, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Abdelaal, K.A.; EL-Maghraby, L.M.; Elansary, H.; Hafez, Y.M.; Ibrahim, E.I.; El-Banna, M.; El-Esawi, M.; Elkelish, A. Treatment of Sweet Pepper with Stress Tolerance-Inducing Compounds Alleviates Salinity Stress Oxidative Damage by Mediating the Physio-Biochemical Activities and Antioxidant Systems. Agronomy 2020, 10, 26. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Ahmed, N.; Saha, T.; Rahman, M.; Rahman, K.; Alam, M.M.; Rohman, M.M.; Nahar, K. Exogenous Salicylic Acid and Kinetin Modulate Reactive Oxygen Species Metabolism and Glyoxalase System to Confer Waterlogging Stress Tolerance in Soybean (Glycine max L.). Plant Stress 2022, 3, 100057. [Google Scholar] [CrossRef]
- Roshdy, A.E.-D.; Alebidi, A.; Almutairi, K.; Al-Obeed, R.; Elsabagh, A. The Effect of Salicylic Acid on the Performances of Salt Stressed Strawberry Plants, Enzymes Activity, and Salt Tolerance Index. Agronomy 2021, 11, 775. [Google Scholar] [CrossRef]
- Guo, Q.; Liu, L.; Barkla, B.J. Membrane Lipid Remodeling in Response to Salinity. Int. J. Mol. Sci. 2019, 20, 4264. [Google Scholar] [CrossRef]
- Wakeel, A.; Farooq, M.; Qadir, M.; Schubert, S. Potassium Substitution by Sodium in Plants. Crit Rev. Plant Sci. 2011, 30, 401–413. [Google Scholar] [CrossRef]
- Per, T.S.; Fatma, M.; Mohd., A.; Javied, S.; Khan, N.A. Salicylic Acid and Nutrients Interplay in Abiotic Stress Tolerance. In Salicylic Acid: A Multifaceted Hormone; Nazar, R., Iqbal, N., Khan, N.A., Eds.; Springer: Singapore, 2017; pp. 221–237. [Google Scholar]
- Tohma, O.; Esitken, A. Response of Salt Stressed Strawberry Plants to Foliar Salicylic Acid Pre-Treatments. J. Plant Nutr. 2011, 34, 590–599. [Google Scholar] [CrossRef]
- El-Nasharty, A.B.; El-Nwehy, S.S.; Abou El-Nour, E.-Z.A.E.-M.A.; Rezk, A.I. Impact of Salicylic Acid Foliar Application on Two Wheat Cultivars Grown under Saline Conditions. Pak. J. Bot 2019, 51, 1939–1944. [Google Scholar] [CrossRef]
- Taha, R.S.; Seleiman, M.F.; Shami, A.; Alhammad, B.A.; Mahdi, A.H.A. Integrated Application of Selenium and Silicon Enhances Growth and Anatomical Structure, Antioxidant Defense System and Yield of Wheat Grown in Salt-Stressed Soil. Plants 2021, 10, 1040. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Aslam, M.T.; Alhammad, B.A.; Hassan, M.U.; Maqbool, R.; Chattha, M.U.; Khan, I.; Gitari, H.I.; Uslu, O.S.; Roy, R.; et al. Salinity Stress in Wheat: Effects, Mechanisms and Management Strategies. Phyton 2022, 91, 667–694. [Google Scholar]
- Li, G.; Peng, X.; Wei, L.; Kang, G. Salicylic Acid Increases the Contents of Glutathione and Ascorbate and Temporally Regulates the Related Gene Expression in Salt-Stressed Wheat Seedlings. Gene 2013, 529, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Hafez, E.M.; Gharib, H.S. Effect of Exogenous Application of Ascorbic Acid on Physiological and Biochemical Characteristics of Wheat under Water Stress. Int J. Plant Prod. 2016, 10, 579–596. [Google Scholar]
- El-Hawary, M.M.; Gad, K.I.; Osman, A.; Ismail, A. Physiological Response of Some Wheat Varieties to Foliar Application with Yeast, Potassium and Ascorbic Acid under Salt Affected Soil Conditions. Biosci. Res. 2019, 16, 1009–1027. [Google Scholar]
- Sadak, M.S.; Mahmod, M.H.; Dawood, M.G. Response of Growth, Yield and Yield Quality of Wheat (Triticum aestivum L.) to Ascorbic Acid or Nicotinamide Application in Newly Reclaimed Soil. J. Plant Prod. 2008, 33, 1783–1797. [Google Scholar] [CrossRef]
Figure 1.
Leaf area index (LAI) of wheat affected by AA and SA treatments in two winter seasons 2018/2019 and 2019/2020; (A,B) LAI at 65 DAS, (C,D) LAI at 80 DAS, (E,F) LAI at 95 DAS. Data are mean ± standard error (n = 4). Letters represent significant differences between treatments at p < 0.05 level according to the LSD test.
Figure 1.
Leaf area index (LAI) of wheat affected by AA and SA treatments in two winter seasons 2018/2019 and 2019/2020; (A,B) LAI at 65 DAS, (C,D) LAI at 80 DAS, (E,F) LAI at 95 DAS. Data are mean ± standard error (n = 4). Letters represent significant differences between treatments at p < 0.05 level according to the LSD test.

Figure 2.
Photosynthetic pigments of wheat leaves affected by AA and SA treatments in the two winter seasons 2018/2019 and 2019/2020. (A,B): chlorophyll a, (C,D): chlorophyll b, and (E,F): carotenoids. Data are mean ± standard error (n = 4). Letters represent significant differences between treatments at p < 0.05 level according to LSD test.
Figure 2.
Photosynthetic pigments of wheat leaves affected by AA and SA treatments in the two winter seasons 2018/2019 and 2019/2020. (A,B): chlorophyll a, (C,D): chlorophyll b, and (E,F): carotenoids. Data are mean ± standard error (n = 4). Letters represent significant differences between treatments at p < 0.05 level according to LSD test.

Figure 3.
Antioxidant enzyme activities affected by AA and SA treatments in wheat leaves at 80 DAS during the two winter seasons 2018/2019 and 2019/2020. (A,B): SOD, (C,D): CAT, (E,F): APX, (G,H): POD. Data are mean ± standard error (n = 4). Letters represent significant differences between treatments at p < 0.05 level according to LSD test.
Figure 3.
Antioxidant enzyme activities affected by AA and SA treatments in wheat leaves at 80 DAS during the two winter seasons 2018/2019 and 2019/2020. (A,B): SOD, (C,D): CAT, (E,F): APX, (G,H): POD. Data are mean ± standard error (n = 4). Letters represent significant differences between treatments at p < 0.05 level according to LSD test.

Figure 4.
Wheat leaf soluble sugar content affected by AA and SA treatments at 80 DAS during the two winter seasons (A) 2018/2019 and (B) 2019/2020. Data are mean ± standard error (n = 4). Letters represent significant differences between treatments at p < 0.05 level according to LSD test.
Figure 4.
Wheat leaf soluble sugar content affected by AA and SA treatments at 80 DAS during the two winter seasons (A) 2018/2019 and (B) 2019/2020. Data are mean ± standard error (n = 4). Letters represent significant differences between treatments at p < 0.05 level according to LSD test.

Figure 5.
Lipid peroxidation in wheat affected by AA and SA treatments at 80 DAS during the two winter seasons (A) 2018/2019 and (B) 2019/2020. Data are mean ± standard error (n = 4). Letters represent significant differences between treatments at p < 0.05 level according to LSD test.
Figure 5.
Lipid peroxidation in wheat affected by AA and SA treatments at 80 DAS during the two winter seasons (A) 2018/2019 and (B) 2019/2020. Data are mean ± standard error (n = 4). Letters represent significant differences between treatments at p < 0.05 level according to LSD test.

Figure 6.
Effect of foliar spray and priming with antioxidant AA and SA on proline (A,B); glycine betaine (C,D); and total phenols (E,F) contents of wheat leaves in the two winter seasons 2018/2019 and 2019/2020. Data are mean ± standard error (n = 4). Letters represent significant differences between treatments at p < 0.05 level according to the LSD test.
Figure 6.
Effect of foliar spray and priming with antioxidant AA and SA on proline (A,B); glycine betaine (C,D); and total phenols (E,F) contents of wheat leaves in the two winter seasons 2018/2019 and 2019/2020. Data are mean ± standard error (n = 4). Letters represent significant differences between treatments at p < 0.05 level according to the LSD test.

Figure 7.
Effect of antioxidant AA and SA as a foliar spray and priming on sodium content (A,B); potassium content (C,D); and Na+/K+ ratio (E,F) of wheat leaves in the two winter seasons 2018/2019 and 2019/2020. Data are mean ± standard error (n = 4). Letters represent significant differences between treatments at p < 0.05 level according to the LSD test.
Figure 7.
Effect of antioxidant AA and SA as a foliar spray and priming on sodium content (A,B); potassium content (C,D); and Na+/K+ ratio (E,F) of wheat leaves in the two winter seasons 2018/2019 and 2019/2020. Data are mean ± standard error (n = 4). Letters represent significant differences between treatments at p < 0.05 level according to the LSD test.

Figure 8.
Effect of foliar spray and priming with antioxidant AA and SA on the protein content of wheat seeds in the two winter seasons (A) 2018/2019 and (B) 2019/2020. Data are mean ± standard error (n = 4). Letters represent significant differences between treatments at p < 0.05 level according to the LSD test.
Figure 8.
Effect of foliar spray and priming with antioxidant AA and SA on the protein content of wheat seeds in the two winter seasons (A) 2018/2019 and (B) 2019/2020. Data are mean ± standard error (n = 4). Letters represent significant differences between treatments at p < 0.05 level according to the LSD test.

Figure 9.
Correlation coefficients of wheat grain yield on growth, physiological traits, and yield components in the two seasons (A) 2018–2019 and (B) 2019–2020. According to the color scale, the darker positive scale represents most treatment responses, while the darker negative stripes show the least response. The dark red color shows a high positive association, and the dark blue shows a high negative correlation between traits. Similarly, as the color intensity decreases, the treatments show moderate performance in both the positive and negative ranges.
Figure 9.
Correlation coefficients of wheat grain yield on growth, physiological traits, and yield components in the two seasons (A) 2018–2019 and (B) 2019–2020. According to the color scale, the darker positive scale represents most treatment responses, while the darker negative stripes show the least response. The dark red color shows a high positive association, and the dark blue shows a high negative correlation between traits. Similarly, as the color intensity decreases, the treatments show moderate performance in both the positive and negative ranges.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The soil physical and chemical properties of the experimental site in seasons 2018/2019 and 2019/20.
Table 1.
The soil physical and chemical properties of the experimental site in seasons 2018/2019 and 2019/20.
| Season | EC (dS m−1) in (1:5) Extract | pH (1:2.5) | Texture | Cations (mL Equivalent L−1) | Anions (mL Equivalent L−1) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Ca2+ | Mg2+ | Na+ | K+ | HCO3− | Cl− | SO42− | ||||
| 2018/2019 | 6.5 | 8.20 | Clay | 10.00 | 17.91 | 54.93 | 1.54 | 1.68 | 60.75 | 21.95 |
| 2019/2020 | 6.2 | 8.20 | Clay | 10.00 | 17.71 | 51.93 | 1.55 | 1.68 | 57.75 | 21.76 |
Table 2.
Meteorological conditions of the experimental site in seasons 2018/2019 and 2019/2020.
| Month | Temperature °C | Precipitation (mm) | Perennial Average | |
|---|---|---|---|---|
| Max | Min | |||
| 11/2018 | 26.4 | 17.9 | 0 | 54 |
| 12/2018 | 21.7 | 13 | 11 | |
| 1/2019 | 19.9 | 11.4 | 20 | |
| 2/2019 | 22.7 | 13.6 | 5 | |
| 3/2019 | 25.8 | 15.5 | 2 | |
| 4/2019 | 26.6 | 16.4 | 16 | |
| 11/2019 | 24.7 | 17 | 12 | 73 |
| 12/2019 | 21 | 12.7 | 16 | |
| 1/2020 | 18.8 | 9.9 | 2 | |
| 2/2020 | 19.9 | 10.5 | 26 | |
| 3/2020 | 21.7 | 12.1 | 8 | |
| 4/2020 | 24.8 | 14.3 | 9 | |
Table 3.
Growth parameters (plant height, CGR, and NAR) of wheat plants treated with AA and SA as priming and foliar application at different concentrations in the two winter seasons 2018/2019 and 2019/2020.
Table 3.
Growth parameters (plant height, CGR, and NAR) of wheat plants treated with AA and SA as priming and foliar application at different concentrations in the two winter seasons 2018/2019 and 2019/2020.
| Treatments | Plant Height (cm) | CGR (g m−2 Day−1) | NAR (g m−2 Day−1) | |||
|---|---|---|---|---|---|---|
| (65–80 Day) | (80–95 Day) | (65–80 Day) | (80–95 Day) | |||
| 2018/2019 | ||||||
| Priming | Water | 90.75 f | 14.12 h | 17.05 g | 10.27 f | 11.37 g |
| A 100 | 92.75 bcde | 15.38 de | 18.22 cd | 10.98 d | 12.24 d | |
| A 200 | 94.25 b | 16.04 abc | 18.84 b | 11.37 b | 12.59 b | |
| S 100 | 92.25 cdef | 15.19 ef | 17.99 de | 10.82 d | 12.08 e | |
| S 200 | 93.50 bc | 15.78 bcd | 18.55 bc | 11.23 bc | 12.43 c | |
| A 100 + S 100 | 95.50 a | 16.41 a | 19.34 a | 11.62 a | 12.80 a | |
| Foliar spray | Water | 89.50 g | 13.45 i | 16.31 h | 10.09 g | 11.13 h |
| A 100 | 91.50 def | 14.80 fg | 17.72 ef | 10.81 d | 11.97 e | |
| A 200 | 92.25 cdef | 15.61 cde | 18.34 cd | 11.17 c | 12.34 cd | |
| S 100 | 91.00 ef | 14.57 g | 17.42 fg | 10.63 e | 11.80 f | |
| S 200 | 92.00 cdef | 15.23 ef | 17.99 de | 10.98 d | 12.04 e | |
| A 100 + S 100 | 93.25 bcd | 16.17 ab | 18.94 b | 11.41 b | 12.66 ab | |
| LSD 0.05 | 1.23 | 0.38 | 0.36 | 0.16 | 0.15 | |
| 2019/2020 | ||||||
| Priming | Water | 92.3 f | 14.74 f | 17.26 g | 10.32 g | 11.42 i |
| A 100 | 94.3 cd | 15.87 cd | 18.63 cde | 11.14 cd | 12.39 ef | |
| A 200 | 96.0 ab | 16.44 b | 19.30 b | 11.48 b | 12.71 bc | |
| S 100 | 93.5 cdef | 15.54 de | 18.45 cde | 10.96 de | 12.24 fg | |
| S 200 | 95.0 bc | 16.28 bc | 18.85 bc | 11.44 b | 12.63 cd | |
| A 100 + S 100 | 97.0 a | 17.08 a | 19.93 a | 11.88 a | 13.07 a | |
| Foliar spray | Water | 90.5 g | 13.85 g | 16.84 h | 10.17 g | 11.22 j |
| A 100 | 93.0 def | 15.59 de | 18.15 ef | 10.89 e | 12.10 g | |
| A 200 | 94.8 bc | 16.25 bc | 18.74 cd | 11.25 c | 12.49 de | |
| S 100 | 92.5 ef | 15.17 e | 17.83 f | 10.72 f | 11.93 h | |
| S 200 | 94.0 cde | 15.84 cd | 18.29 def | 11.13 cd | 12.18 g | |
| A 100 + S 100 | 95.3 bc | 16.88 a | 19.24 b | 11.55 b | 12.82 b | |
| LSD 0.05 | 1.22 | 0.40 | 0.39 | 0.15 | 0.15 | |
Letters represent significant differences between treatments at p < 0.05 level according to the LSD test.
Table 4.
Yield and yield attributes of wheat affected by foliar and priming application with various rates of antioxidants during 2018/2019 and 2019/2020 seasons.
Table 4.
Yield and yield attributes of wheat affected by foliar and priming application with various rates of antioxidants during 2018/2019 and 2019/2020 seasons.
| Treatments | Spike Length (cm) | Spike Weight (g) | 1000-Grain Weight (g) | Grain Yield (t ha−1) | Straw Yield (t ha−1) | Harvest Index | |
|---|---|---|---|---|---|---|---|
| 2018/2019 | |||||||
| Priming | Water | 11.2 ef | 4.12 f | 48.4 h | 6.99 e | 11.4 c | 38.1 e |
| A 100 | 12.3 bcd | 4.51 bcd | 53.0 d | 7.59 bcd | 11.9 ab | 38.9 d | |
| A 200 | 12.8 ab | 4.67 b | 54.3 b | 7.93 ab | 12.0 a | 39.7 ab | |
| S 100 | 11.9 cd | 4.46 cde | 51.7 f | 7.51 cd | 11.9 ab | 38.8 d | |
| S 200 | 12.4 abc | 4.56 bcd | 53.9 c | 7.79 bc | 12.0 ab | 39.4 bc | |
| A 100 + S 100 | 13.0 a | 4.82 a | 55.1 a | 8.13 a | 12.1 a | 40.2 a | |
| Foliar spray | Water | 11.0 f | 4.00 f | 47.6 i | 6.70 f | 11.3 c | 37.2 f |
| A 100 | 12.0 cd | 4.41 de | 51.4 f | 7.51 cd | 11.8 ab | 38.8 d | |
| A 200 | 12.4 abc | 4.53 bcd | 53.2 d | 7.75 bc | 11.9 ab | 39.4 bc | |
| S 100 | 11.6 def | 4.32 e | 50.1 g | 7.28 d | 11.7 b | 38.3 e | |
| S 200 | 11.8 cde | 4.44 cde | 52.5 e | 7.64 bc | 11.9 ab | 39.2 bcd | |
| A 100 + S 100 | 12.8 ab | 4.61 bc | 54.2 bc | 7.95 ab | 12.1 a | 39.7 ab | |
| LSD 0.05 | 0.504 | 0.122 | 0.377 | 0.242 | 0.185 | 0.420 | |
| 2019/2020 | |||||||
| Priming | Water | 11.3 d | 4.18 f | 48.6 i | 7.07 f | 11.5 c | 38.1 f |
| A 100 | 12.5 abc | 4.66 bcd | 53.2 d | 7.89 bcd | 12.0 ab | 39.6 bc | |
| A 200 | 13.0 ab | 4.81 b | 54.7 b | 8.16 b | 12.2 a | 40.1 b | |
| S 100 | 12.1 c | 4.61 cde | 52.1 f | 7.79 cde | 12.0 ab | 39.5 cd | |
| S 200 | 12.6 abc | 4.72 bc | 54.2 c | 8.05 bc | 12.1 ab | 40.0 b | |
| A 100 + S 100 | 13.3 a | 5.01 a | 55.4 a | 8.42 a | 12.2 a | 40.9 a | |
| Foliar spray | Water | 11.1 d | 4.08 f | 47.7 j | 6.98 f | 11.4 c | 38.0 f |
| A 100 | 12.3 bc | 4.50 de | 51.6 g | 7.65 de | 11.9 b | 39.2 de | |
| A 200 | 12.6 abc | 4.68 bc | 53.4 d | 7.99 bc | 12.0 ab | 39.9 b | |
| S 100 | 11.9 c | 4.46 e | 50.3 h | 7.55 e | 11.8 b | 38.9 e | |
| S 200 | 12.0 c | 4.61 cde | 52.8 e | 7.90 bcd | 11.9 ab | 39.8 bc | |
| A 100 + S 100 | 13.0 ab | 4.78 bc | 54.4 bc | 8.13 b | 12.2 a | 40.1 b | |
| LSD 0.05 | 0.535 | 0.120 | 0.344 | 0.219 | 0.176 | 0.349 | |
Letters represent significant differences between treatments at p < 0.05 level according to the LSD test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
El-Hawary, M.M.; Hashem, O.S.M.; Hasanuzzaman, M. Seed Priming and Foliar Application with Ascorbic Acid and Salicylic Acid Mitigate Salt Stress in Wheat. Agronomy 2023, 13, 493. https://doi.org/10.3390/agronomy13020493
AMA Style
El-Hawary MM, Hashem OSM, Hasanuzzaman M. Seed Priming and Foliar Application with Ascorbic Acid and Salicylic Acid Mitigate Salt Stress in Wheat. Agronomy. 2023; 13(2):493. https://doi.org/10.3390/agronomy13020493
Chicago/Turabian StyleEl-Hawary, Mohammed Mohammed, Omnia S. M. Hashem, and Mirza Hasanuzzaman. 2023. "Seed Priming and Foliar Application with Ascorbic Acid and Salicylic Acid Mitigate Salt Stress in Wheat" Agronomy 13, no. 2: 493. https://doi.org/10.3390/agronomy13020493
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.