
Dung Beetle Activity Is Soil-Type-Dependent and Modulates Pasture Growth and Associated Soil Microbiome
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Facility Construction, Preparation of Lysimeter Columns, and Experimental Design
2.2. Establishment of Pasture
2.3. Introduction of Dung Beetles and Cattle Dung
2.4. Irrigation of Soil Columns
2.5. Collection of Soil Leachates
2.6. Optimisation Protocols for Testing for E. coli and Coliform Bacteria
2.7. Enumeration of Total Coliforms and E. coli in Soil Leachates
2.8. Collection of Pasture Biomass
2.9. Analysis of Total N and P in Soil Leachates
2.10. Microbial DNA Extraction from Soils and Marker Gene Sequencing
2.11. Analysis of Quantitative Data
2.12. Analysis of Marker Gene Sequence Data
3. Results
3.1. Soil Water Content
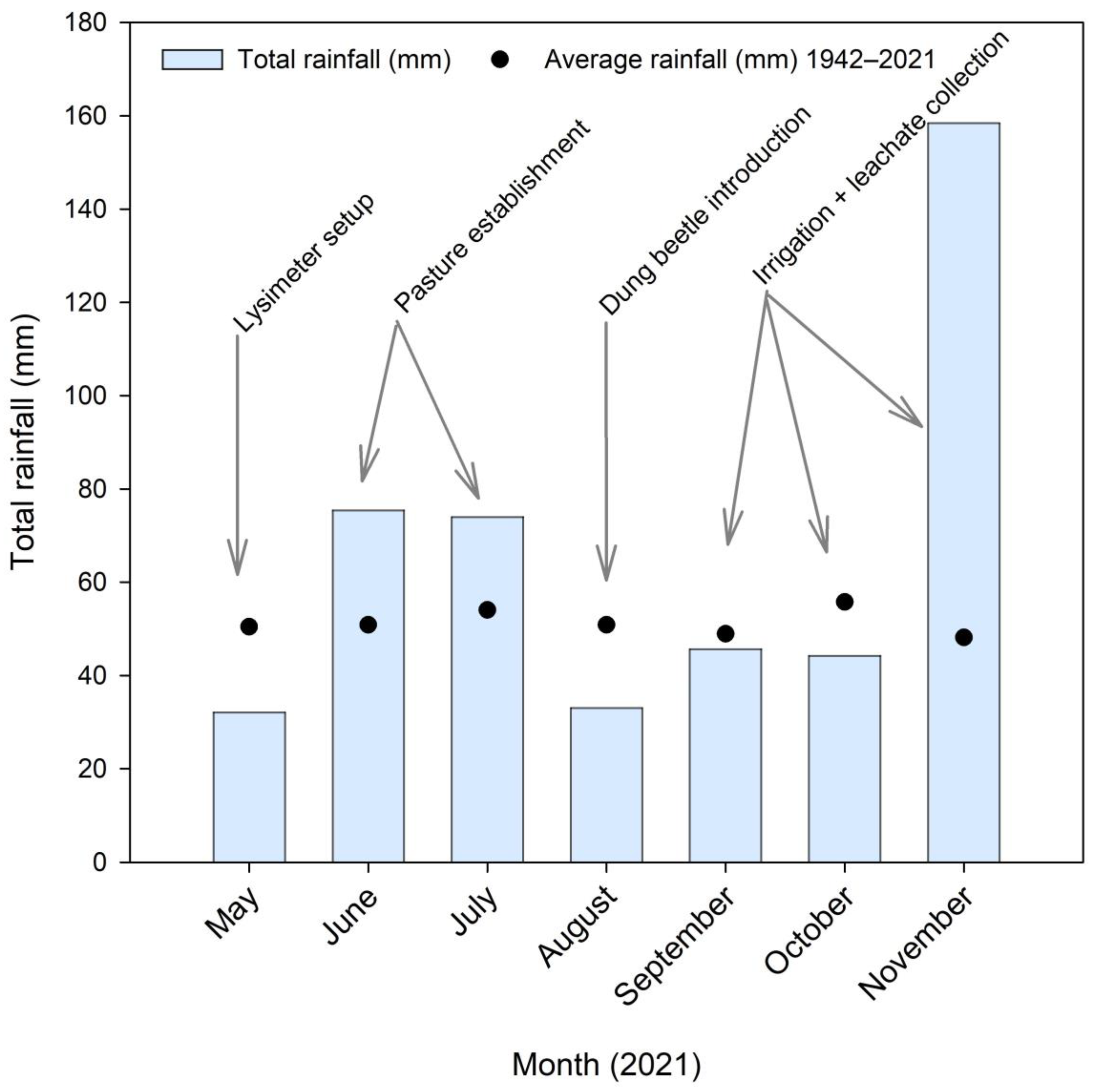
3.2. Rainfall
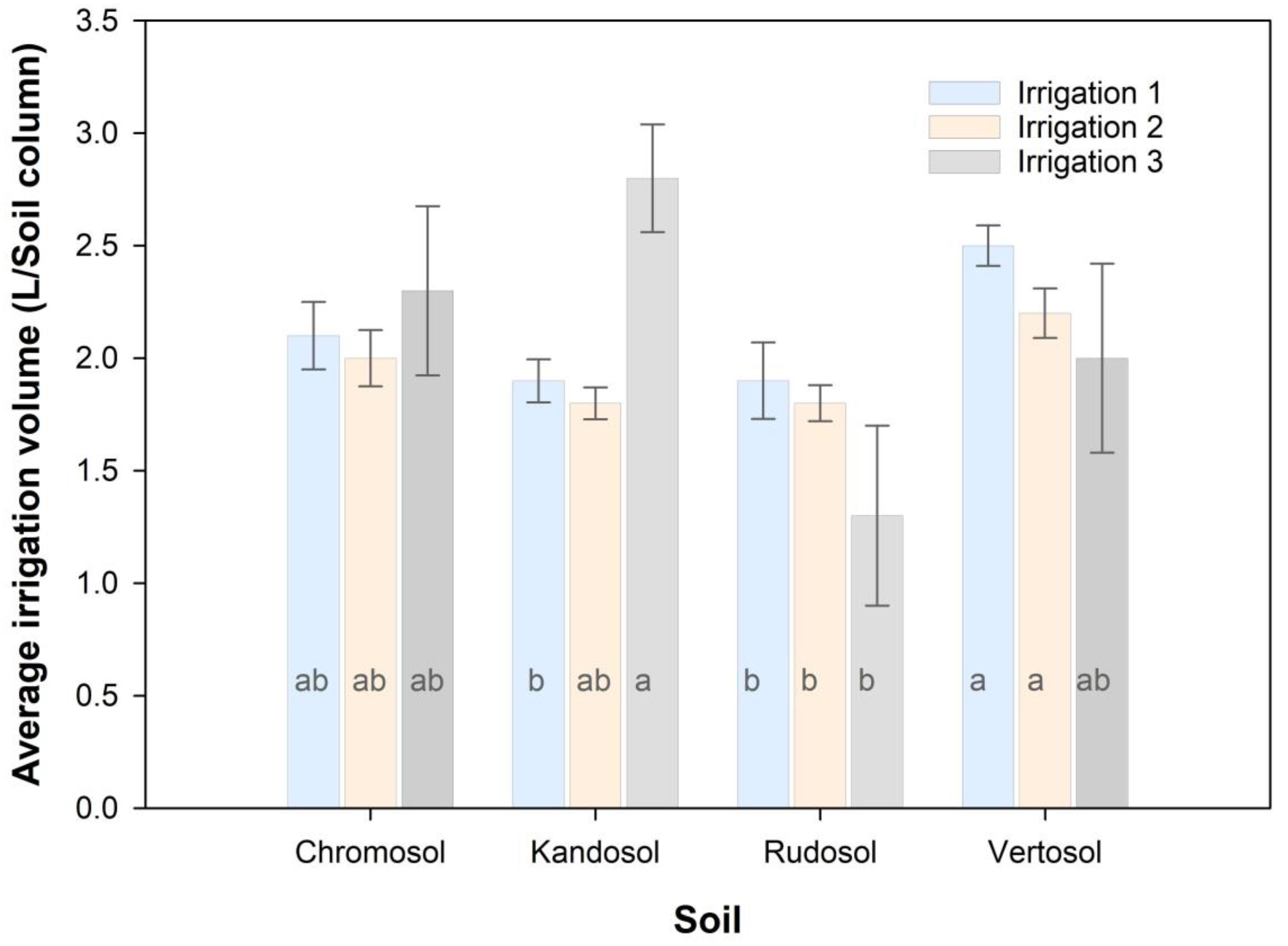
3.3. Irrigation
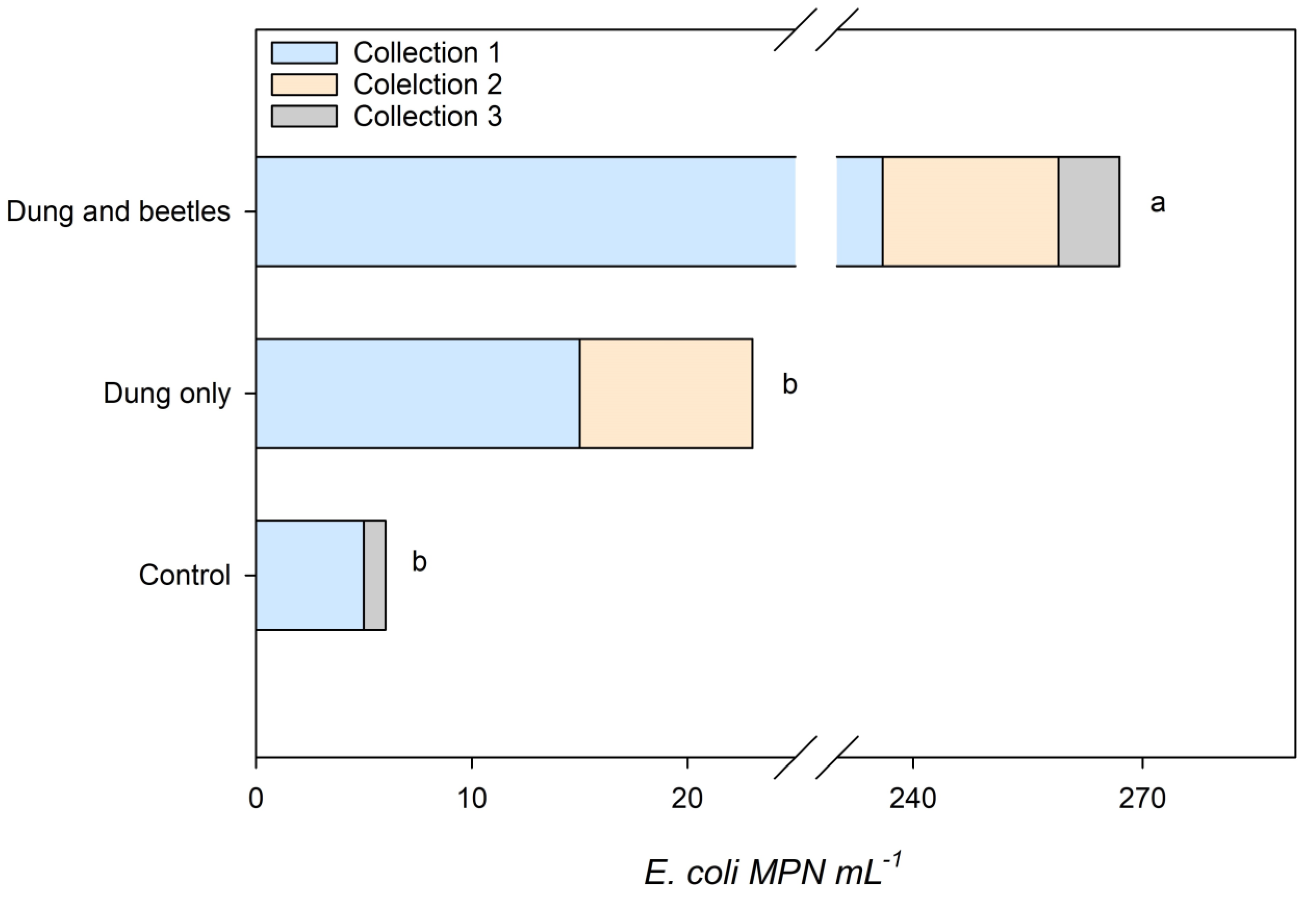
3.4. Microbial Contamination in Leachates
3.5. Total N, Total P, and NOx in Soil Leachates
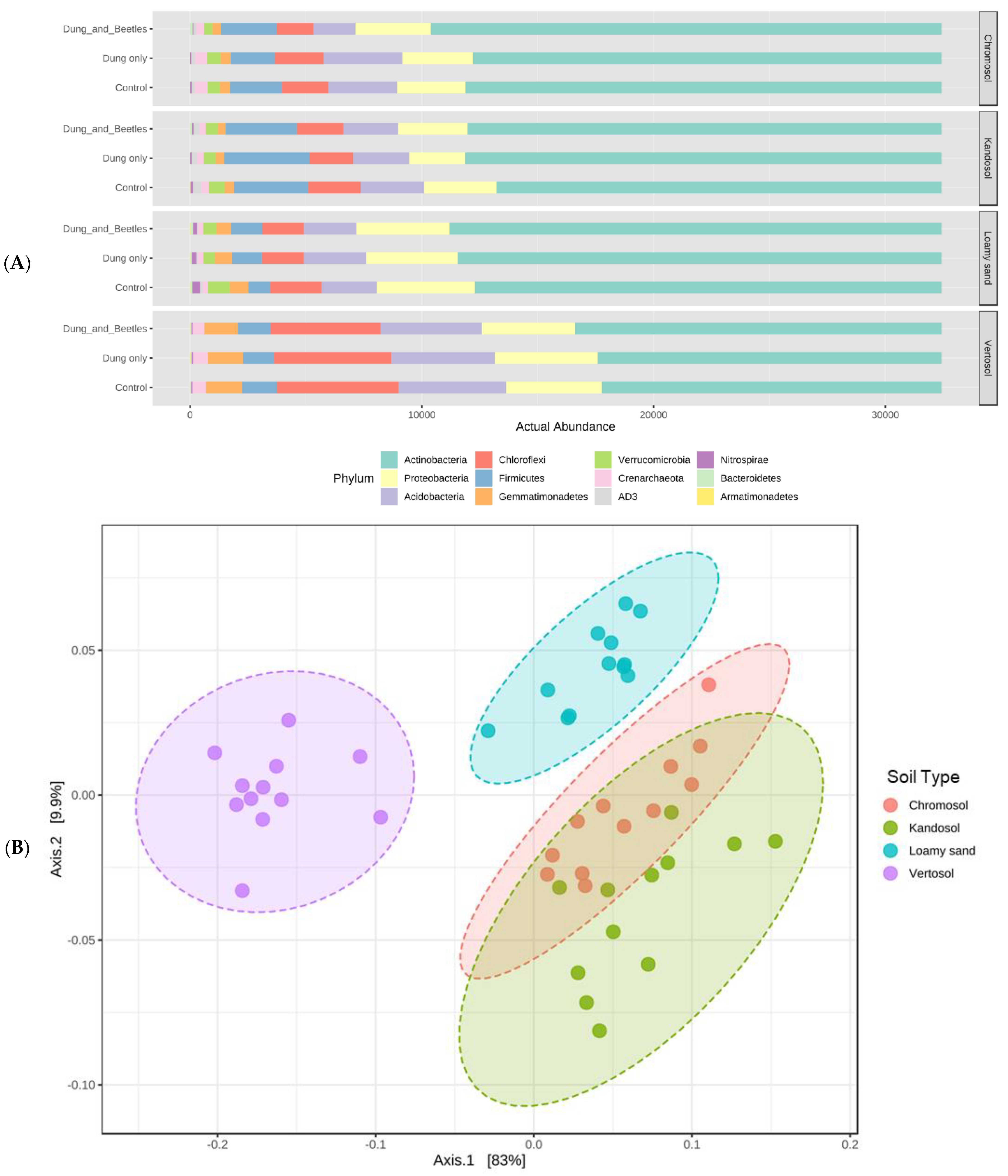
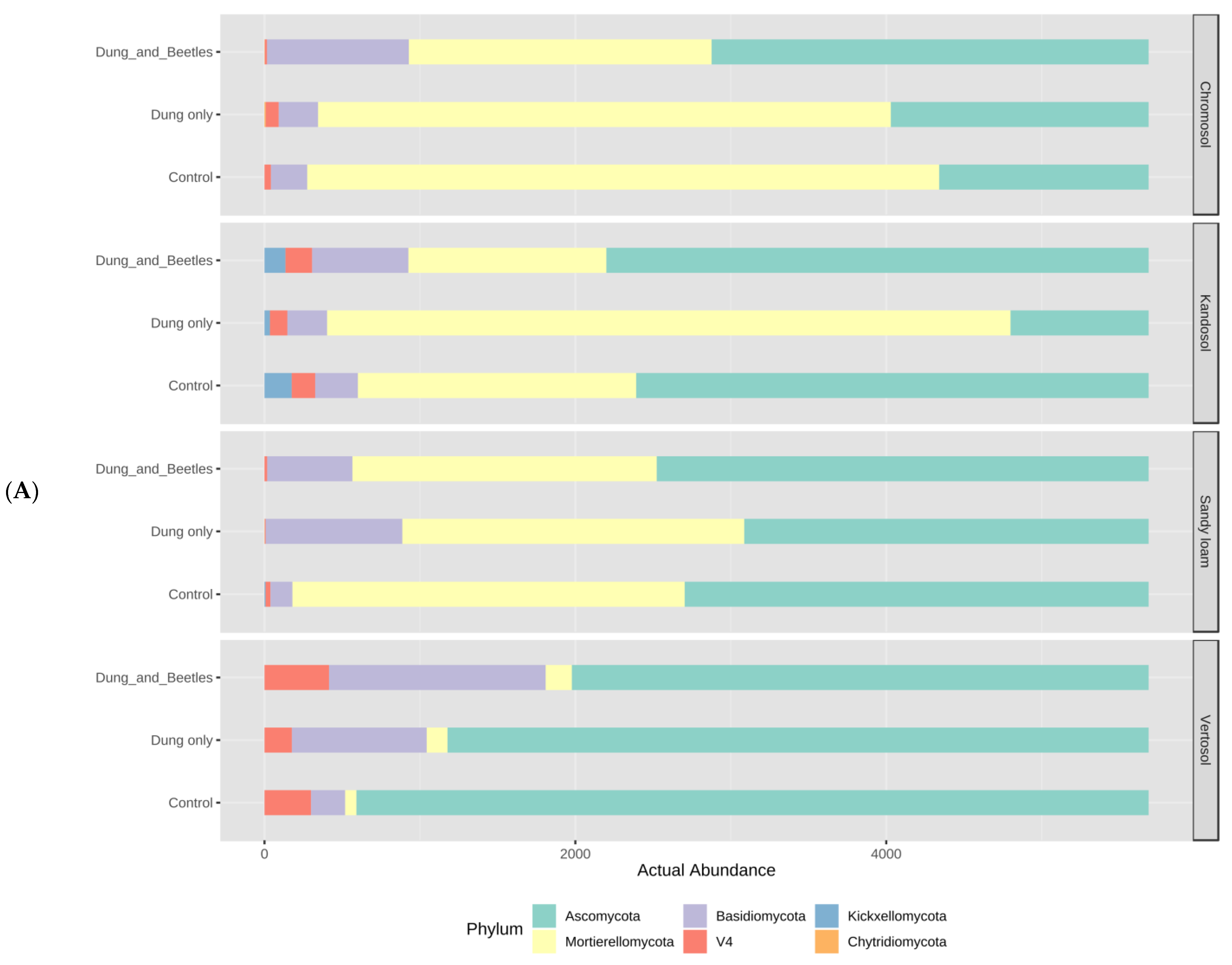
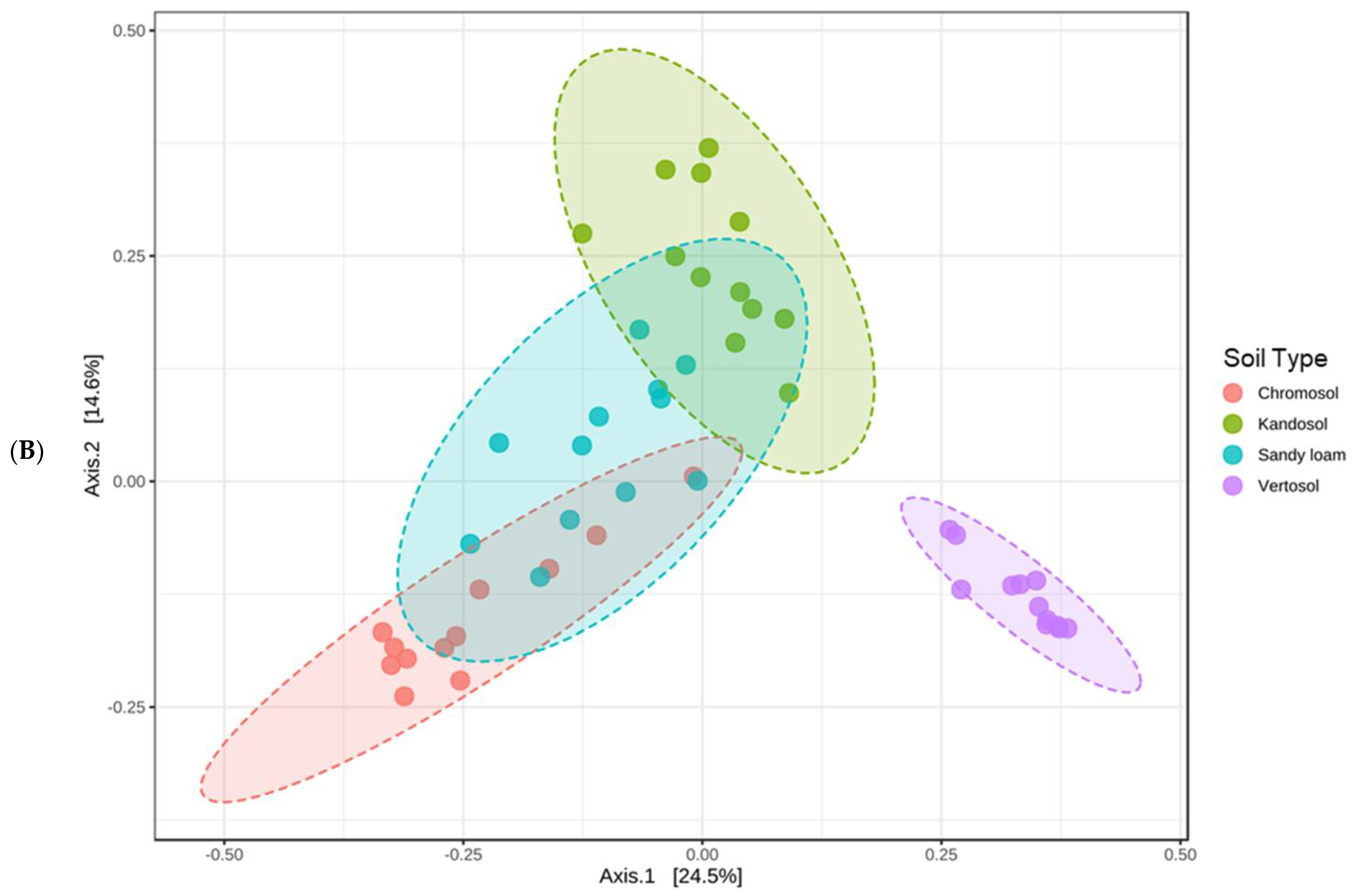
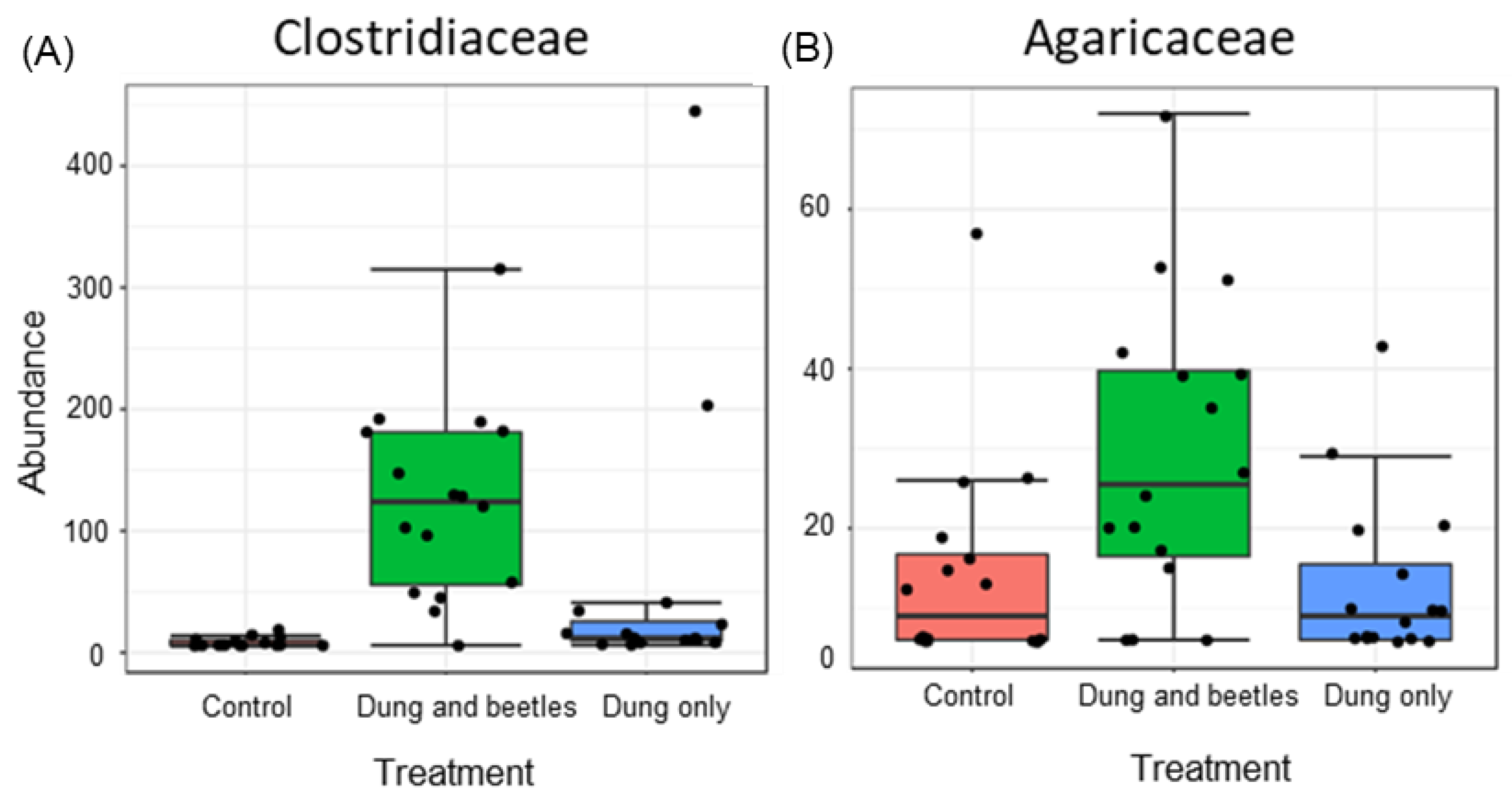
3.6. Soil Microbiome Assessment
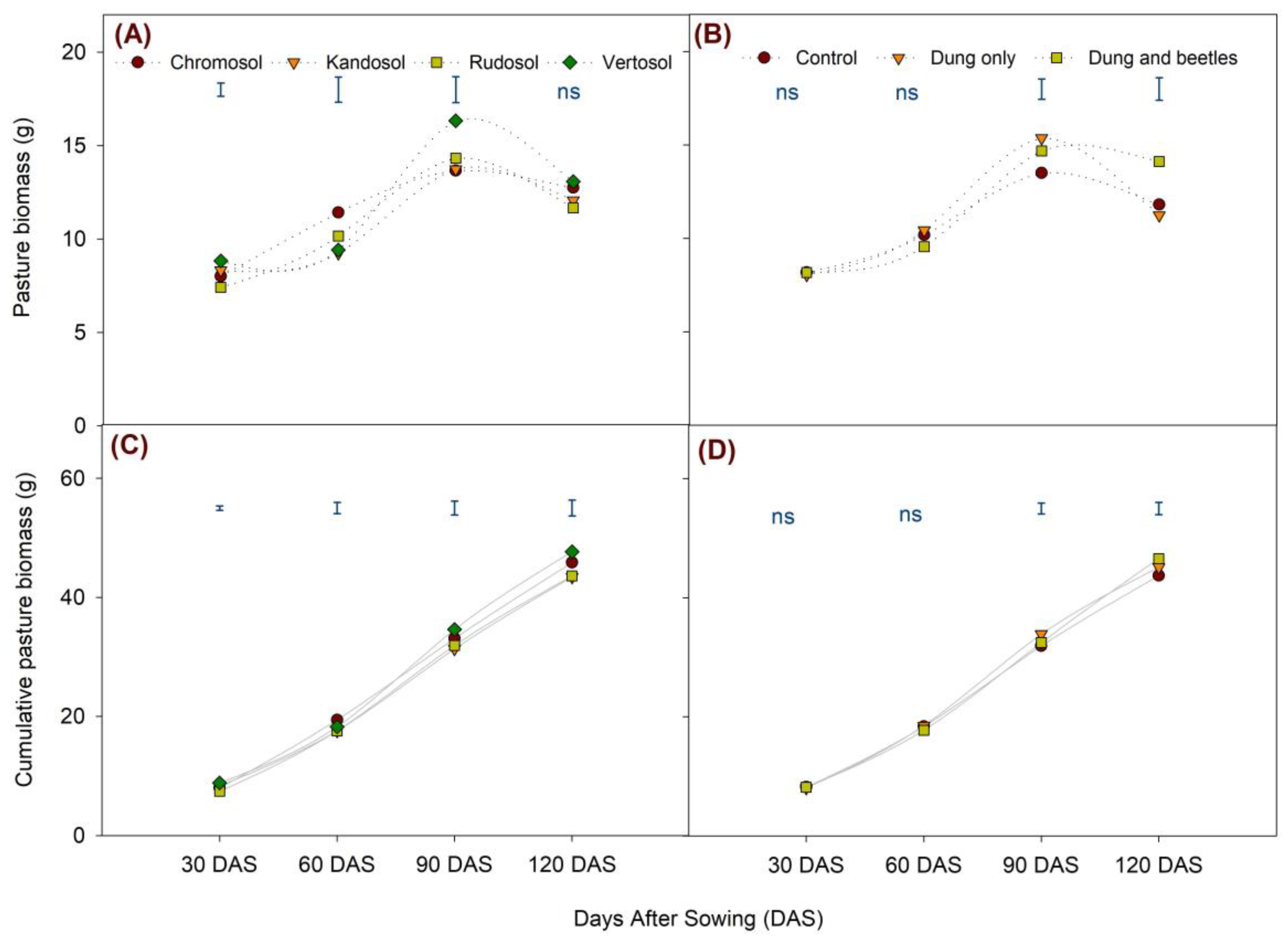
3.7. Pasture Biomass
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Doube, B.M. Ecosystem services provided by dung beetles in Australia. Basic Appl. Ecol. 2018, 26, 35–49. [Google Scholar] [CrossRef]
- Bornemissza, G. Australian dung beetle project 1965–1975. AMRC Rev. Aust. Meat Res. Commun. 1976, 30, 1–30. [Google Scholar]
- Pokhrel, M.R.; Cairns, S.C.; Hemmings, Z.; Floate, K.D.; Andrew, N.R. A Review of Dung Beetle Introductions in the Antipodes and North America: Status, Opportunities, and Challenges. Environ. Entomol. 2021, 50, 762–780. [Google Scholar] [CrossRef]
- Vernes, K.; Pope, L.C.; Hill, C.J.; Bärlocher, F. Seasonally, dung specificity and competition in dung beetle assemblages in the Australian Wet Tropics, north-eastern Australia. J. Trop. Ecol. 2005, 21, 1–8. [Google Scholar] [CrossRef]
- Beynon, S.A.; Wainwright, W.A.; Christie, M. The application of an ecosystem services framework to estimate the economic value of dung beetles to the U.K. cattle industry. Ecol. Entomol. 2015, 40, 124–135. [Google Scholar] [CrossRef] [Green Version]
- Johnson, S.N.; Lopaticki, G.; Barnett, K.; Facey, S.L.; Powell, J.R.; Hartley, S.E.; Rasmann, S. An insect ecosystem engineer alleviates drought stress in plants without increasing plant susceptibility to an above-ground herbivore. Funct. Ecol. 2016, 30, 894–902. [Google Scholar] [CrossRef] [Green Version]
- Edwards, P. Introduced Dung Beetles in Australia 1967–2007: Current Status and Future Directions; Dung Beetles for Landcare Farming Committee: Sinnamon Park, Australia, 2009. [Google Scholar]
- Doube, B.M.; Macqueen, A.; Ridsdill-Smith, T.J.; Weir, T.A. Native and Introduced Dung Beetles in Australia; Princeton University Press: Princeton, NJ, USA, 2014; p. 255. [Google Scholar]
- Edwards, P.; Wilson, P.K.; Wright, J. Introduced Dung Beetles in Australia: A Pocket Field Guide, 3rd ed.; CSIRO Publishing: Clayton, Australia, 2015. [Google Scholar]
- Doube, B. The Pasture Growth and Environmental Benefits of Dung Beetles to the Southern Australian Cattle Industry; Meat & Livestock Industry: North Sydney, Australia, 2008; pp. 85–93. [Google Scholar]
- Doube, B.; Dalton, G. Dung Beetles: Transform a Pollutant into an Environmental and Agricultural Benefit: A Practical Guide to the Benefits, Behaviour, Species, Establishment, Management and Monitoring of Dung Beetles; Fleurieu Beef Group: Strathalbyn, Australia, 2003. [Google Scholar]
- Ryan, U.; Yang, R.; Gordon, C.; Doube, B. Effect of dung burial by the dung beetle Bubas bison on numbers and viability of Cryptosporidium oocysts in cattle dung. Exp. Parasitol. 2011, 129, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tixier, T.; Bloor, J.M.G.; Lumaret, J.-P. Species-specific effects of dung beetle abundance on dung removal and leaf litter decomposition. Acta Oecol. 2015, 69, 31–34. [Google Scholar] [CrossRef]
- Keller, N.; van Meerveld, I.; Ghazoul, J.; Chiew, L.Y.; Philipson, C.D.; Godoong, E.; Slade, E.M. Dung beetles as hydrological engineers: Effects of tunnelling on soil infiltration. Ecol. Entomol. 2022, 47, 84–94. [Google Scholar] [CrossRef]
- Nichols, E.; Spector, S.; Louzada, J.; Larsen, T.; Amezquita, S.; Favila, M.E. Ecological functions and ecosystem services provided by Scarabaeinae dung beetles. Biol. Conserv. 2008, 141, 1461–1474. [Google Scholar] [CrossRef]
- Li, Y.; Li, M.; Liu, H.; Qin, W. Influence of soil texture on the process of subsurface drainage in saturated-unsaturated zones. Int. J. Agric. Biol. Eng. 2021, 14, 82–89. [Google Scholar] [CrossRef]
- Gladish, D.W.; He, D.; Wang, E. Pattern analysis of Australia soil profiles for plant available water capacity. Geoderma 2021, 391, 114977. [Google Scholar] [CrossRef]
- Salomão, R.P.; Pires, D.d.A.; Baccaro, F.B.; Schietti, J.; Vaz-de-Mello, F.Z.; Lima, A.P.; Magnusson, W.E. Water table level and soil texture are important drivers of dung beetle diversity in Amazonian lowland forests. Appl. Soil Ecol. 2022, 170, 104260. [Google Scholar] [CrossRef]
- Silva, T.R.; Medeiros, M.B.; Noronha, S.E.; Pinto, J.R.R. Species distribution models of rare tree species as an evaluation tool for synergistic human impacts in the Amazon rainforest. Braz. J. Bot. 2017, 40, 963–971. [Google Scholar] [CrossRef] [Green Version]
- Scholtz, C.H.; Davis, A.L.V.; Kryger, U. Evolutionary Biology and Conservation of Dung Beetles; Pensoft Pub.: Sofia, Bulgaria; Moscow, Russia, 2009. [Google Scholar]
- Farias, P.M.d.; Hernández, M.I.M. Dung Beetles Associated with Agroecosystems of Southern Brazil: Relationship with Soil Properties. Rev. Bras. Cienc. Solo 2017, 41, e0160248. [Google Scholar] [CrossRef]
- Isbell, R.F. The Australian Soil Classification; CSIRO Publishing: Clayton, Australia, 2021; p. 192. [Google Scholar]
- Aislabie, J.; McLeod, M.; McGill, A.; Rhodes, P.; Forgie, S. Impact of dung beetle activity on the quality of water percolating through Allophanic soil. Soil Res. 2021, 59, 266. [Google Scholar] [CrossRef]
- Jones, M.S.; Tadepalli, S.; David, F.B.; Wu, V.C.H.; Drummond, F. Suppression of Escherichia coli O157:H7 by dung beetles (Coleoptera: Scarabaeidae) using the lowbush blueberry agroecosystem as a model system. PLoS ONE 2015, 10, e0120904. [Google Scholar] [CrossRef] [PubMed]
- Barragán, F.; Douterlungne, D.; Ramírez-Hernández, A.; Gelviz-Gelvez, S.M.; Guzmán Miranda, A.V.; Rodas Ortíz, J.P. The rolling dung master: An ecosystem engineer beetle mobilizing soil nutrients to enhance plant growth across a grassland management intensity gradient in drylands. J. Arid Environ. 2022, 197, 104673. [Google Scholar] [CrossRef]
- Evans, K.S.; Mamo, M.; Wingeyer, A.; Schacht, W.H.; Eskridge, K.M.; Bradshaw, J.; Ginting, D. Soil Fauna Accelerate Dung Pat Decomposition and Nutrient Cycling into Grassland Soil. Rangel. Ecol. Manag. 2019, 72, 667–677. [Google Scholar] [CrossRef] [Green Version]
- Slade, E.M.; Roslin, T.; Santalahti, M.; Bell, T. Disentangling the ‘brown world’ faecal–detritus interaction web: Dung beetle effects on soil microbial properties. Oikos 2016, 125, 629–635. [Google Scholar] [CrossRef] [Green Version]
- Kaleri, A.R.; Ma, J.; Abro, S.A.; Faqir, Y.; Nabi, F.; Hakeem, A.; Ahmed, A.; Ahmed, S.; Jakhar, A.M.; Shah, S.M.; et al. Dung Beetle Improves soil Bacterial Diversity and Enzyme Activity and Enhances Growth and Antioxidant Content of Chinese Cabbage (Brassica rapa ssp. pekinensis). J. Soil Sci. Plant Nutr. 2021, 21, 3387–3401. [Google Scholar] [CrossRef]
- Manning, P.; Slade, E.M.; Beynon, S.A.; Lewis, O.T. Effect of dung beetle species richness and chemical perturbation on multiple ecosystem functions. Ecol. Entomol. 2017, 42, 577–586. [Google Scholar] [CrossRef]
- Dung Beetle Ecosystem Engineers. Project Objectives in About Dung Beetle Ecosystem Engineers Project. Available online: https://www.dungbeetles.com.au/about/project-objectives (accessed on 15 November 2022).
- Ma, L.; Haque, K.; Weston, P.; Gurr, G.; Eberbach, P.; Weston, L. Evaluating the effects of tunnelling dung beetle species on multiple soil functions. In Proceedings of the SSA/NZSSS Joint Conference 2021, Cairns, Australia, 27 June–2 July 2021. [Google Scholar]
- Bureau of Meteorology. Summary Statistics Wagga Wagga AMO in Climate Statistics for Australian Locations; Bureau of Meteorology: Melbourne, Australia, 2022. [Google Scholar]
- Isbell, R. The Australian Soil Classification; CSIRO Publishing: Clayton, Australia, 2016. [Google Scholar]
- GRDC. Southern GrowNotes Oats; GRDC: Barton, Australia, 2017. [Google Scholar]
- GRDC. Southern GrowNotes Vetch; GRDC: Barton, Australia, 2018. [Google Scholar]
- Hall, M.; Beiko, R.G. 16S rRNA gene analysis with QIIME2. In Microbiome Analysis; Springer: Berlin/Heidelberg, Germany, 2018; pp. 113–129. [Google Scholar]
- Edgar, R. Usearch; Lawrence Berkeley National Lab. (LBNL): Berkeley, CA, USA, 2010. [Google Scholar]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Abarenkov, K.; Nilsson, R.H.; Larsson, K.-H.; Alexander, I.J.; Eberhardt, U.; Erland, S.; Høiland, K.; Kjøller, R.; Larsson, E.; Pennanen, T. The UNITE database for molecular identification of fungi–recent updates and future perspectives. New Phytol. 2010, 186, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J. MicrobiomeAnalyst: A web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 2017, 45, W180–W188. [Google Scholar] [CrossRef] [PubMed]
- Yu, O.-Y.; Raichle, B.; Sink, S. Impact of biochar on the water holding capacity of loamy sand soil. Int. J. Energy Environ. Eng. 2013, 4, 44. [Google Scholar] [CrossRef] [Green Version]
- Donn, M.; Reed, D.; Vanderzalm, J.; Page, D. Assessment of E. coli Attenuation during Infiltration of Treated Wastewater: A Pathway to Future Managed Aquifer Recharge. Water 2020, 12, 173. [Google Scholar] [CrossRef] [Green Version]
- Dworkin, M.; Falkow, S.; Rosenberg, E.; Schleifer, K.-H.; Stackebrandt, E. The Prokaryotes: Volume 4: Bacteria: Firmicutes, Cyanobacteria; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Vellinga, E.C. Ecology and distribution of lepiotaceous fungi (Agaricaceae)—A review. Nova Hedwig. 2004, 78, 273–300. [Google Scholar] [CrossRef]
- Meissner, R.; Rupp, H.; Haselow, L. Use of lysimeters for monitoring soil water balance parameters and nutrient leaching. In Climate Change and Soil Interactions; Elsevier: Amsterdam, The Netherlands, 2020; pp. 171–205. [Google Scholar]
- Abdou, H.; Flury, M. Simulation of water flow and solute transport in free-drainage lysimeters and field soils with heterogeneous structures. Eur. J. Soil Sci. 2004, 55, 229–241. [Google Scholar] [CrossRef]
- Eberbach, P.L.; Hoffmann, J.; Moroni, S.J.; Wade, L.J.; Weston, L.A. Rhizo-lysimetry: Facilities for the simultaneous study of root behaviour and resource use by agricultural crop and pasture systems. Plant Methods 2013, 9, 3. [Google Scholar] [CrossRef] [Green Version]
- Odonkor, S.T.; Mahami, T. Escherichia coli as a tool for disease risk assessment of drinking water sources. Int. J. Microbiol. 2020, 2020, 2534130. [Google Scholar] [CrossRef] [PubMed]
- Crampton, A.; Ragusa, A. The E. coli load in self-managed rural water in Australia. Internet J. Microbiol. 2010, 9, 1. [Google Scholar]
- King, T. Effect of Solid Cattle Manure and Liquid HOG Manure Application on Phosphorus and Nitrogen in Soil, Run-Off and Leachate in Saskatchewan Soil; University of Saskatchewan: Saskatoon, SK, Canada, 2015. [Google Scholar]
- King, T.; Schoenau, J.; Elliott, J. Relationship between manure management application practices and phosphorus and nitrogen export in snowmelt run-off water from a Black Chernozem Saskatchewan soil. Sustain. Agric. Res. 2017, 6, 93–114. [Google Scholar] [CrossRef] [Green Version]
- Reid-Soukup, D.A.; Ulery, A.L. Smectites. Soil Mineral. Environ. Appl. 2002, 7, 467–499. [Google Scholar]
- Zhou, H.; Chen, C.; Wang, D.; Arthur, E.; Zhang, Z.; Guo, Z.; Peng, X.; Mooney, S.J. Effect of long-term organic amendments on the full-range soil water retention characteristics of a Vertisol. Soil Tillage Res. 2020, 202, 104663. [Google Scholar] [CrossRef]
- Murphy, S.; Lodge, G. Soil water characteristics of a red chromosol and brown vertosol and pasture growth. In Proceedings of the 10th Australian agronomy conference, Hobart’, Hobart, Australia, 29 January–1 February 2001. [Google Scholar]
- Maldonado, M.B.; Aranibar, J.N.; Serrano, A.M.; Chacoff, N.P.; Vázquez, D.P. Dung beetles and nutrient cycling in a dryland environment. Catena 2019, 179, 66–73. [Google Scholar] [CrossRef]
- Kirk, A.A. The biology of Bubas bison (L.) (Coleoptera: Scarabaeidae) in southern France and its potential for recycling dung in Australia. Bull. Entomol. Res. 1983, 73, 129–136. [Google Scholar] [CrossRef]
- Senechkin, I.V.; Speksnijder, A.G.; Semenov, A.M.; Van Bruggen, A.H.; Van Overbeek, L.S. Isolation and partial characterization of bacterial strains on low organic carbon medium from soils fertilized with different organic amendments. Microb. Ecol. 2010, 60, 829–839. [Google Scholar] [CrossRef]
- Blagodatskaya, E.; Kuzyakov, Y. Active microorganisms in soil: Critical review of estimation criteria and approaches. Soil Biol. Biochem. 2013, 67, 192–211. [Google Scholar] [CrossRef]
- Semenov, M.V.; Krasnov, G.S.; Semenov, V.M.; Ksenofontova, N.; Zinyakova, N.B.; van Bruggen, A.H. Does fresh farmyard manure introduce surviving microbes into soil or activate soil-borne microbiota? J. Environ. Manag. 2021, 294, 113018. [Google Scholar] [CrossRef]
- Sun, M.; Liu, X.; Shi, K.; Peng, F.; Xiao, Y. Effects of Root Zone Aeration on Soil Microbes Species in a Peach Tree Rhizosphere and Root Growth. Microorganisms 2022, 10, 1879. [Google Scholar] [CrossRef]
- Li, Y.; Niu, W.; Wang, J.; Liu, L.; Zhang, M.; Xu, J. Effects of artificial soil aeration volume and frequency on soil enzyme activity and microbial abundance when cultivating greenhouse tomato. Soil Sci. Soc. Am. J. 2016, 80, 1208–1221. [Google Scholar] [CrossRef] [Green Version]
- Schlatter, D.C.; Kahl, K.; Carlson, B.; Huggins, D.R.; Paulitz, T. Fungal community composition and diversity vary with soil depth and landscape position in a no-till wheat-based cropping system. FEMS Microbiol. Ecol. 2018, 94, fiy098. [Google Scholar] [CrossRef]
- Sipilä, T.P.; Yrjälä, K.; Alakukku, L.; Palojärvi, A. Cross-site soil microbial communities under tillage regimes: Fungistasis and microbial biomarkers. Appl. Environ. Microbiol. 2012, 78, 8191–8201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, A.; Xu, M.; Wang, B.; Zhang, W.; Liang, G.; Hou, E.; Luo, Y. Manure acts as a better fertilizer for increasing crop yields than synthetic fertilizer does by improving soil fertility. Soil Tillage Res. 2019, 189, 168–175. [Google Scholar] [CrossRef]
- Bertone, M.A.; Green, J.T.; Washburn, S.P.; Poore, M.H.; Watson, D.W. The Contribution of Tunneling Dung Beetles to Pasture Soil Nutrition. Forage Grazingl. 2006, 4, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Yoshitake, S.; Soutome, H.; Koizumi, H. Deposition and decomposition of cattle dung and its impact on soil properties and plant growth in a cool-temperate pasture. Ecol. Res. 2014, 29, 673–684. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Chromosol | Kandosol | Rudosol | Vertosol |
|---|---|---|---|---|
| Phosphorus (mg/kg P) | 7.22 | 12.79 | 11.26 | 87.03 |
| Nitrate Nitrogen (mg/kg N) | 4.35 | 2.26 | 0.49 | 12.74 |
| Ammonium Nitrogen (mg/kg N) | 6.64 | 7.45 | 5.80 | 12.05 |
| Sulphur (mg/kg S) | 4.64 | 6.94 | 4.17 | 22.29 |
| pH (1:5 water) | 6.91 | 6.80 | 7.00 | 7.66 |
| Electrical Conductivity (dS/m) | 0.04 | 0.04 | 0.03 | 0.11 |
| Estimated Organic Matter (% OM) | 1.33 | 1.40 | 0.62 | 1.61 |
| Effective Cation Exchange Capacity (ECEC) (cmol+/kg) | 3.71 | 6.87 | 2.97 | 29.46 |
| Calcium (%) | 70.31 | 62.13 | 69.61 | 66.16 |
| Magnesium (%) | 14.27 | 21.73 | 16.48 | 28.56 |
| Potassium (%) | 13.60 | 15.09 | 10.94 | 4.43 |
| Sodium—ESP (%) | 1.63 | 0.94 | 2.83 | 0.82 |
| Aluminium (%) | 0.20 | 0.11 | 0.13 | 0.03 |
| Zinc (mg/kg) | 0.60 | 0.65 | <0.5 | 0.82 |
| Manganese (mg/kg) | 37.22 | 27.27 | 24.17 | 11.02 |
| Iron (mg/kg) | 29.47 | 38.05 | 14.57 | 39.67 |
| Copper (mg/kg) | 0.45 | 1.01 | 0.31 | 2.00 |
| Boron (mg/kg) | 0.51 | 0.82 | 0.33 | 1.22 |
| Silicon (mg/kg Si) | 42.45 | 74.65 | 36.98 | 46.31 |
| Total Carbon (%) | 0.76 | 0.80 | 0.35 | 0.92 |
| Total Nitrogen (%) | 0.11 | 0.11 | 0.07 | 0.14 |
| Carbon/Nitrogen Ratio | 6.86 | 7.24 | 4.68 | 6.54 |
| Primer Name | Primer Sequence |
|---|---|
| 341F—Universal 16S | 5′-CCTAYGGGRBGCASCAG-3′ |
| 806R—Universal 16S | 5′-GGACTACNNGGGTATCTAAT-3′ |
| 1F—Universal ITS | 5′-CTTGGTCATTTAGAGGAAGTAA-3′ |
| 2R—Universal ITS | 5′-GCTGCGTTCTTCATCGATGC-3′ |
| Soil Type | Packed Soil Water Content (L/L) | Gravitational Water Content w/w | Bulk Density Gcm−3 |
|---|---|---|---|
| Chromosol | 0.090 | 0.109 | 1.187 |
| Kandosol | 0.073 | 0.093 | 1.165 |
| Rudosol | 0.065 | 0.067 | 1.142 |
| Vertosol | 0.087 | 0.276 | 1.337 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, L.; Weeraratne, N.; Gurusinghe, S.; Aktar, J.; Haque, K.M.S.; Eberbach, P.; Gurr, G.G.; Weston, L.A. Dung Beetle Activity Is Soil-Type-Dependent and Modulates Pasture Growth and Associated Soil Microbiome. Agronomy 2023, 13, 325. https://doi.org/10.3390/agronomy13020325
Ma L, Weeraratne N, Gurusinghe S, Aktar J, Haque KMS, Eberbach P, Gurr GG, Weston LA. Dung Beetle Activity Is Soil-Type-Dependent and Modulates Pasture Growth and Associated Soil Microbiome. Agronomy. 2023; 13(2):325. https://doi.org/10.3390/agronomy13020325
Chicago/Turabian StyleMa, Long, Nirodha Weeraratne, Saliya Gurusinghe, Jesmin Aktar, K. M. Shamsul Haque, Philip Eberbach, Geoff G. Gurr, and Leslie A. Weston. 2023. "Dung Beetle Activity Is Soil-Type-Dependent and Modulates Pasture Growth and Associated Soil Microbiome" Agronomy 13, no. 2: 325. https://doi.org/10.3390/agronomy13020325