
Dwarf Tomato Plants Allow for Managing Agronomic Yield Gains with Fruit Quality and Pest Resistance through Backcrossing
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Experimental Design
2.2. Sample Collection and Evaluation
2.3. Statistical Analyses
3. Results
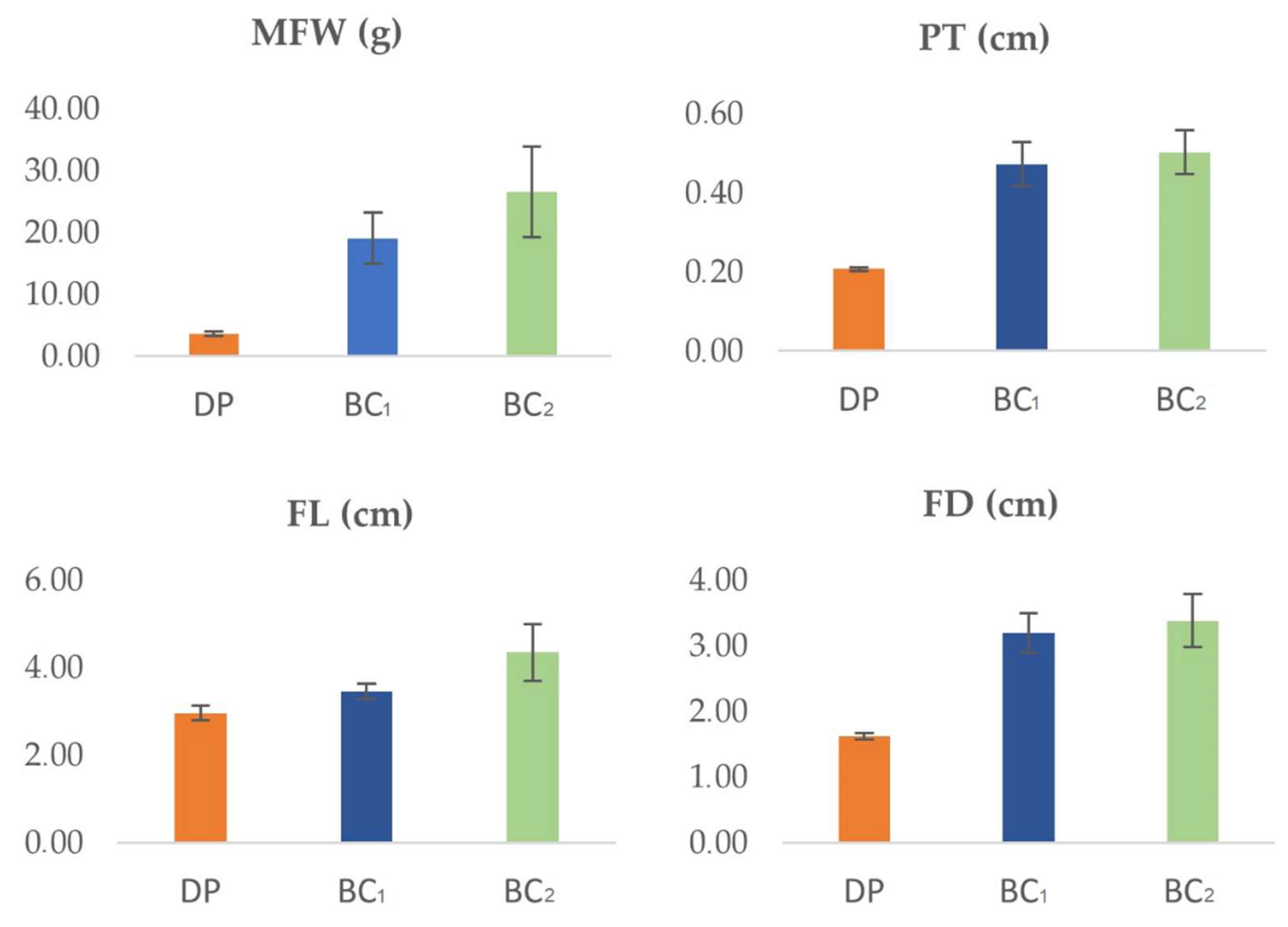
3.1. Agronomic Performance of Different Backcross Generations
3.2. Relative Superiority of BC1 and BC2 Generations
3.3. Fruit Quality Traits
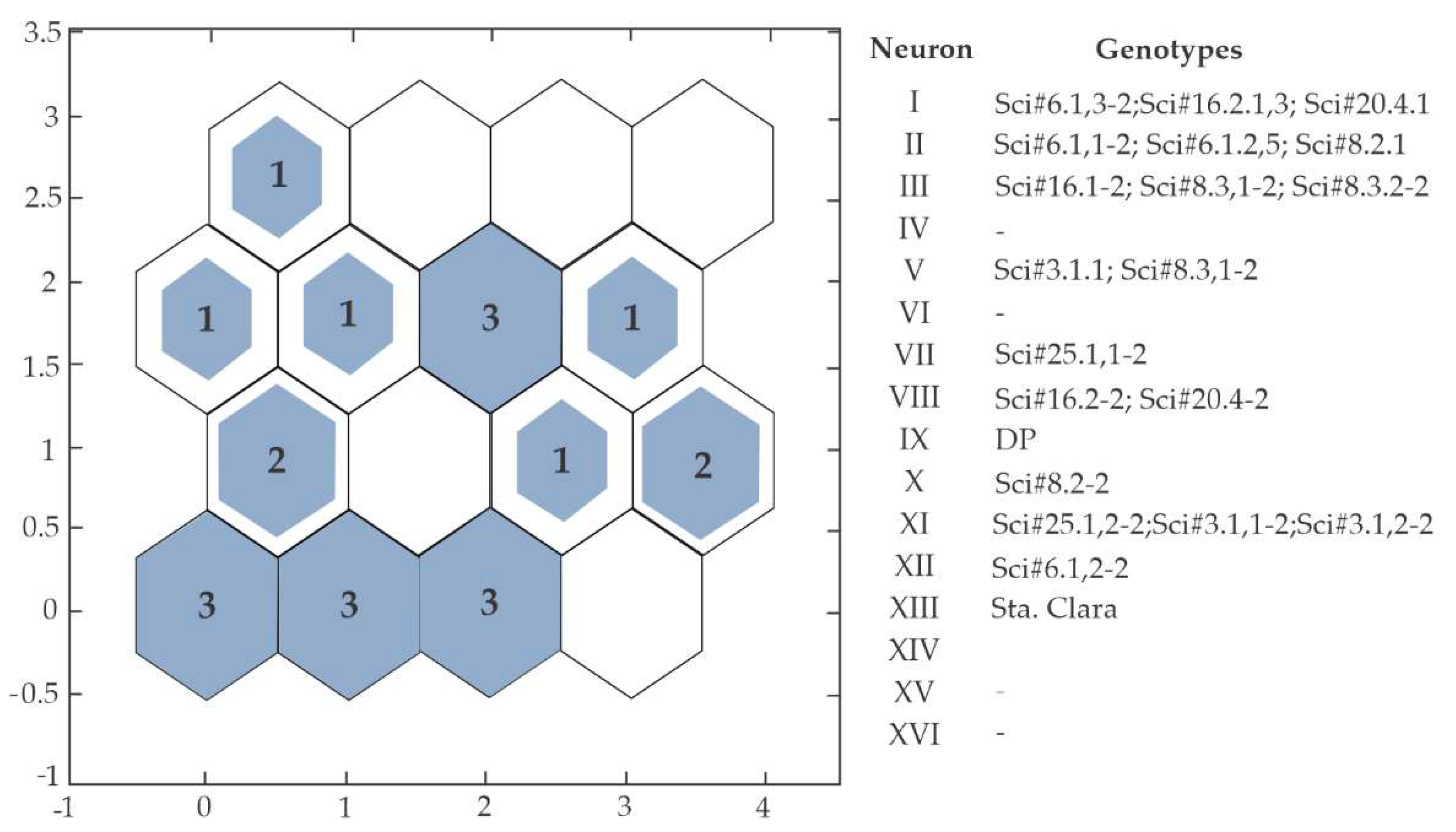
3.4. Genetic Dissimilarity between the Tested Genotypes
4. Discussion
4.1. Feasibility of the Method
4.2. Effect of Backcrossing on Agronomic Traits, Resistance, and Nutritional Quality of Tomato
4.3. Genetic Similarity Using Kohonen Self-Organizing Maps
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- IBGE. Levantamento Sistemático da Produção Agrícola. 2020. Available online: https://sidra.ibge.gov.br/home/lspa/brasil (accessed on 15 February 2021).
- Alvarenga, M.A.R. Tomate: Produção em Campo, em Casa-de-Vegetação e em Hidroponia; Editora UFLA: Lavras, Brazil, 2013; 455p. [Google Scholar]
- Shirahige, F.H.; de Melo, A.M.; Purquerio, L.F.V.; Carvalho, C.R.L.; de Melo, P.C.T. Yield and fruit quality of Santa Cruz and Italian tomatoes depending on fruit thinning. Hortic. Bras. 2010, 28, 292–298. [Google Scholar] [CrossRef] [Green Version]
- Deleo, J.P.B.; Brito Junior, J.S.; Paranhos, G.G. Especial tomate: Gestão sustentável. Hortifruti Bras. 2016, 15, 10–23. [Google Scholar]
- Osorio-Gracia, N.A.; Ortiz-Gonzalez, D.; Sandoval-Contreras, H.A.; Gomes, C.N.; Toledo-Picoli, E.A.; García-Parra, M.A. Produtividade de tomates oriundos da polinização natural e mecânica em duas épocas de plantio. Rev. Aronómica Noroeste Argent. 2020, 40, 51–61. [Google Scholar]
- Sestras, A.F.; Jidavu, M.; Sestras, R.E.; Apahidean, M.; Harsan, E.; Tamas, E.; Gao, Y. The response of several tomato cultivars for processing in central Transylvania conditions. II. Fruits quality. Not. Bot. Horti Agrobot. Cluj-Napoca 2006, 34, 62–68. [Google Scholar]
- Luz, J.M.; Bittar, C.A.; Oliveira, R.C.; Nascimento, A.R.; Nogueira, A.P. Performance and genetic divergence of tomato genotypes for industrial processing. Hortic. Bras. 2016, 34, 483–490. [Google Scholar] [CrossRef] [Green Version]
- Finzi, R.R.; Maciel, G.M.; da Silva, E.C.; Luz, J.M.Q.; Borba, M.E.A. Agronomic performance of mini-tomato hybrids from dwarf lines. Ciênc. Agrotecnol. 2017, 41, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Maciel, G.M.; Da Silva, E.C.; Fernandes, M.A.R. Ocorrência de nanismo em planta de tomateiro do tipo grape. Rev. Caatinga 2015, 28, 259–264. [Google Scholar] [CrossRef] [Green Version]
- Finzi, R.R.; Maciel, G.M.; Peres, H.G.; Silva, M.F.E.; Peixoto, J.V.M.; Gomes, D.A. Agronomic potential of BC1F2 dwarf round tomato populations. Ciênc. Agrotecnol. 2020, 44, 1–8. [Google Scholar] [CrossRef]
- de Oliveira, C.S.; Maciel, G.M.; Siquieroli, A.C.S.; Gomes, D.A.; Martins, M.P.D.C.; Finzi, R.R. Selection of F2RC1 saladette-type dwarf tomato plant populations for fruit quality and whitefly resistance. Rev. Bras. Eng. Agríc. Ambient. 2022, 26, 28–35. [Google Scholar] [CrossRef]
- Maluf, W.R.; Maciel, G.M.; Gomes, L.A.A.; Cardoso, M.D.G.; Gonçalves, L.D.; da Silva, E.C.; Knapp, M. Broad-Spectrum Arthropod Resistance in Hybrids between High- and Low-Acylsugar Tomato Lines. Crop Sci. 2010, 50, 439–450. [Google Scholar] [CrossRef]
- Farokhzadeh, S.; Alifakheri, B. Marker-assisted selection for disease resistance: Applications in breeding (Review). Int. J. Agri. Crop Sci. 2014, 7, 1392–1405. [Google Scholar]
- Maciel, G.M.; Silva, E.C. Proposta metodológica para quantificação de acilaçúcares em folíolos de tomateiro. Hortic. Bras. 2014, 32, 174–177. [Google Scholar] [CrossRef] [Green Version]
- Nagata, M.; Yamashita, I. Simple Method for Simultaneous Determination of Chlorophyll and Carotenoids in Tomato Fruit. Nippon. Shokuhin Kogyo Gakkaishi 1992, 39, 925–928. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Amaya, D.B. A Guide to Carotenoids Analysis in Food; International Life Sciences Institute Press: Washington, DC, USA, 2001; 64p. [Google Scholar]
- Rodriguez-Amaya, D.B.; Kimura, M. HarvestPlus Handbook for Carotenoid Analysis; IFPRI: Washington, DC, USA; CIAT: Cali, Colombia, 2004; 58p. [Google Scholar]
- Cruz, C.D. Genes Software—Extended and integrated with the R, Matlab and Selegen. Acta Sci. Agron. 2016, 38, 547–552. [Google Scholar] [CrossRef]
- Fiorini, C.V.A.; Marim, B.G.; Rodrigues, G.; Zaparoli, M.R.; Castro, J.P.A.; Silva, D.J.H. Cultivares. In Tomate: Tecnologia de Produção; Silva, D.J.H., Vale, F.X.R., Eds.; Editora UFV: Viçosa, Brazil, 2007; pp. 65–84. [Google Scholar]
- Zsögön, A.; Cermak, T.; Voytas, D.; Peres, L.E.P. Genome editing as a tool to achieve the crop ideotype and de novo domestication of wild relatives: Case study in tomato. Plant Sci. 2017, 256, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Naeem, M.; Waseem, M.; Zhu, Z.; Zhang, L. Downregulation of SlGRAS15 manipulates plant architecture in tomato (Solanum lycopersicum). Dev. Genes Evol. 2020, 230, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Borém, A.; Miranda, G.V.; Fritsche-Neto, R. Melhoramento de Plantas, 7th ed.; Editora UFV: Viçosa, Brazil, 2017; 543p. [Google Scholar]
- Melo, C.; Souza, M.; Sousa, A.; Viana, A.; Santos, E. Multivariate analysis of backcross progeny of Passiflora L. (Passifloraceae) for pre-breeding genotype selection. Genet. Mol. Res. 2015, 14, 15376–15389. [Google Scholar] [CrossRef] [PubMed]
- García-Fortea, E.; Gramazio, P.; Vilanova, S.; Fita, A.; Mangino, G.; Villanueva, G.; Arrones, A.; Knapp, S.; Prohens, J.; Plazas, M. First successful backcrossing towards eggplant (Solanum melongena) of a New World species, the silverleaf nightshade (S. elaeagnifolium), and characterization of interspecific hybrids and backcrosses. Sci. Hortic. 2019, 246, 563–573. [Google Scholar] [CrossRef]
- Silva, F.A.S.E.; Azevedo, C.A.V. Principal Components Analysis in the Software Assistat-Statistical Attendance. In Proceedings of the 7th World Congress on Computers in Agriculture, Reno, NV, USA, 22–24 June 2009. [Google Scholar]
- Lucini, T.; Faria, M.V.; Rohde, C.; Resende, J.T.V.; Oliveira, J.R.F. Acylsugar and the role of tricomes in tomato genotypes resistence to Tetranychus urticae. Arthopod-Plant Interact. 2015, 9, 45–53. [Google Scholar] [CrossRef]
- Dias, D.M.; Corte, L.E.-D.; Resende, J.T.V.; Zeffa, D.M.; Resende, N.C.V.; Zanin, D.S.; Filho, R.B.D.L. Acylsugars in tomato varieties confer resistance to the whitefly and reduce the spread of fumagine. Bragantia 2021, 80, e4421. [Google Scholar] [CrossRef]
- Dias, D.; Resende, J.; Faria, M.; Camargo, L.; Chagas, R.; Lima, I. Selection of processing tomato genotypes with high acyl sugar content that are resistant to the tomato pinworm. Genet. Mol. Res. 2013, 12, 381–389. [Google Scholar] [CrossRef]
- Martí, R.; Roselló, S.; Cebolla-Cornejo, J. Tomato as a source of carotenoids and polyphenols targeted to cancer prevention. Cancers 2016, 8, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, H.; Hiraga, M.; Shirasawa, K.; Nishiyama, M.; Kanahama, K.; Kanayama, Y. Analysis of a tomato introgression line, IL8-3, with increased Brix content. Sci. Hortic. 2013, 153, 103–108. [Google Scholar] [CrossRef]
- Schwarz, K.; De Resende, J.T.V.; Preczenhak, A.P.; De Paula, J.T.; Faria, M.V.; Dias, D.M. Desempenho agronômico e qualidade físico-química de híbridos de tomateiro em cultivo rasteiro. Hortic. Bras. 2013, 31, 410–418. [Google Scholar] [CrossRef]
- Bhattarai, K.; Sharma, S.; Panthee, D.R. Diversity among Modern Tomato Genotypes at Different Levels in Fresh-Market Breeding. Int. J. Agron. 2018, 2018, 4170432. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, O.; ElH, M.T.; Badr, A.; Wali, A.M. Evaluating the Performance of 16 Egyptian Wheat Varieties Using Self-Organizing Map (SOM) and Cluster Analysis. J. Appl. Sci. 2016, 16, 47–53. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, D.B.O.; Medeiros, L.A.; Carvalho, G.D.O.; Pimentel, I.M.; Rojas, G.X.; Sousa, L.A.; Souza, G.M.; De Sousa, L.B. Use of computational intelligence in the genetic divergence of colored cotton plants. Biosci. J. 2021, 37, e37007. [Google Scholar] [CrossRef]
- de Oliveira, C.S.; Maciel, G.M.; Siquieroli, A.C.S.; Gomes, D.A.; Diniz, N.M.; Luz, J.M.Q.; Yada, R.Y. Artificial neural networks and genetic dissimilarity among saladette type dwarf tomato plant populations. Food Chem. Mol. Sci. 2021, 3, 100056. [Google Scholar] [CrossRef] [PubMed]
- Gomes, D.A.; Maciel, G.M.; Siquieroli, A.C.S.; de Oliveira, C.S.; Finzi, R.R.; Marques, D.J. Selection of BC1F3 populations of Santa Cruz type dwarf tomato plant by computational intelligence techniques. Bragantia 2021, 80, e4821. [Google Scholar] [CrossRef]
- Sant’Anna, I.D.C.; Gouvêa, L.R.L.; Martins, M.A.; Junior, E.J.S.; de Freitas, R.S.; Gonçalves, P.D.S. Genetic diversity associated with natural rubber quality in elite genotypes of the rubber tree. Sci. Rep. 2021, 11, 1081. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Genotype | Generation | MFW | PT | FL | FD | FS | NL | IL | AS |
|---|---|---|---|---|---|---|---|---|---|
| Donor parent | - | 3.61 c | 0.20 d | 2.95 b | 1.61 c | 1.83 d | 2.00 a | 1.26 a | 36.28 b |
| Santa Clara | - | 26.59 a | 0.55 a | 3.84 b | 3.62 a | 1.05 a | 2.05 a | 7.27 b | 25.25 b |
| Sci#6.1,1-2 | BC2 | 20.32 b | 0.47 b | 3.77 b | 3.26 b | 1.15 a | 2.75 c | 1.99 a | 36.63 b |
| Sci#6.1,2-2 | BC2 | 25.73 a | 0.49 b | 3.90 b | 3.51 a | 1.10 a | 2.91 c | 1.70 a | 37.83 b |
| Sci#6.1,3-2 | BC2 | 20.36 b | 0.48 b | 3.98 b | 3.09 b | 1.29 b | 2.44 b | 2.33 a | 47.81 a |
| Sci#16.2-2 | BC2 | 17.41 b | 0.40 c | 3.71 b | 2.87 b | 1.29 b | 2.97 c | 1.84 a | 41.80 a |
| Sci#16.1-2 | BC2 | 24.11 a | 0.50 a | 4.16 a | 3.37 a | 1.23 a | 2.86 c | 1.95 a | 36.55 b |
| Sci#25.1,1-2 | BC2 | 30.81 a | 0.54 a | 4.67 a | 3.45 a | 1.35 b | 3.00 c | 1.70 a | 35.30 b |
| Sci#25.1,2-2 | BC2 | 28.82 a | 0.52 a | 4.64 a | 3.48 a | 1.33 b | 2.66 c | 1.54 a | 37.35 b |
| Sci#3.1,1-2 | BC2 | 26.12 a | 0.51 a | 4.62 a | 3.35 a | 1.39 b | 2.67 c | 2.03 a | 32.19 b |
| Sci#3.1,2-2 | BC2 | 31.59 a | 0.54 a | 4.81 a | 3.81 a | 1.28 b | 2.80 c | 1.60 a | 41.10 a |
| Sci#8.2-2 | BC2 | 25.01 a | 0.47 b | 4.63 a | 2.92 b | 1.58 c | 2.60 c | 1.97 a | 43.16 a |
| Sci#20.4-2 | BC2 | 32.72 a | 0.54 a | 4.76 a | 3.31 b | 1.44 b | 2.77 c | 1.65 a | 47.45 a |
| Sci#8.3,1-2 | BC2 | 29.61 a | 0.50 a | 4.08 a | 3.71 a | 1.10 a | 3.00 c | 1.76 a | 40.40 a |
| Sci#8.3,2-2 | BC2 | 32.56 a | 0.56 a | 4.69 a | 3.76 a | 1.24 a | 2.73 c | 1.94 a | 41.86 a |
| Sci#6.1.2,5 | BC1 | 19.77 b | 0.52 a | 3.36 b | 3.28 b | 1.02 a | 2.41 b | 1.71 a | 39.53 b |
| Sci#16.2.1,3 | BC1 | 17.93 b | 0.47 b | 3.60 b | 3.05 b | 1.18 a | 2.22 a | 1.57 a | 39.26 b |
| Sci#3.1.1 | BC1 | 14.93 b | 0.38 c | 3.27 b | 3.05 b | 1.08 a | 2.92 c | 1.83 a | 42.81 a |
| Sci#8.2.1 | BC1 | 25.13 a | 0.52 a | 3.46 b | 3.62 a | 0.95 a | 2.75 c | 1.74 a | 39.91 b |
| Sci#20.4.1 | BC1 | 18.64 b | 0.47 b | 3.50 b | 3.12 b | 1.12 a | 2.33 b | 1.82 a | 31.91 b |
| Sci#8.3,1.2 | BC1 | 17.98 b | 0.45 b | 3.53 b | 3.02 b | 1.18 a | 2.49 b | 2.02 a | 44.21 a |
| Solanum pennellii | - | - | - | - | - | - | - | - | 50.57 a |
| KS 1 | - | 0.043 | 0.646 | 0.011 | 0.040 | 0.010 | 0.329 | 0.037 | 0.839 |
| OM 2 | - | 0.014 | 0.658 | 0.056 | 0.014 | 0.184 | 0.021 | 0.782 | 0.414 |
| F (Tukey) 3 | - | 0.9380 | 0.443 | 0.827 | 0.528 | 0.878 | 0.986 | 0.982 | 0.123 |
| Genotype 1 | Generation | TSS | CC | LC |
|---|---|---|---|---|
| Donor parent | Donor parent | 6.93 a | 1.74 a | 2.94 c |
| Santa Clara | Check | 5.37 b | 0.87 b | 5.07 a |
| Sci#6.1,1-2 | BC2 | 4.90 c | 1.34 a | 2.89 c |
| Sci#6.1,2-2 | BC2 | 4.80 c | 0.88 b | 3.08 c |
| Sci#6.1,3-2 | BC2 | 4.74 c | 1.37 a | 2.72 c |
| Sci#16.2-2 | BC2 | 5.74 b | 0.48 b | 4.04 b |
| Sci#16.1-2 | BC2 | 4.37 c | 1.40 a | 2.72 c |
| Sci#25.1,1-2 | BC2 | 4.77 c | 1.10 b | 2.42 d |
| Sci#25.1,2-2 | BC2 | 5.51 b | 1.27 a | 2.55 c |
| Sci#3.1,1-2 | BC2 | 5.03 c | 0.87 b | 2.40 d |
| Sci#3.1,2-2 | BC2 | 5.02 c | 0.91 b | 2.00 d |
| Sci#8.2-2 | BC2 | 5.72 b | 1.31 a | 2.42 d |
| Sci#20.4-2 | BC2 | 4.99 c | 0.96 b | 3.73 b |
| Sci#8.3,1-2 | BC2 | 4.58 c | 1.20 a | 2.67 c |
| Sci#8.3,2-2 | BC2 | 4.42 c | 1.38 a | 3.36 b |
| Sci#6.1.2,5 | BC1 | 4.68 c | 1.31 a | 2.57 c |
| Sci#16.2.1,3 | BC1 | 4.15 c | 1.11 b | 1.81 d |
| Sci#3.1.1 | BC1 | 5.46 b | 1.41 a | 2.15 d |
| Sci#8.2.1 | BC1 | 4.88 c | 1.11 b | 2.15 d |
| Sci#20.4.1 | BC1 | 5.03 c | 1.06 b | 2.75 c |
| Sci#8.3,1.2 | BC1 | 5.03 c | 1.24 a | 2.05 d |
| KS 2 | - | 0.079 | 0.025 | 0.046 |
| OM 3 | - | 0.101 | 0.353 | 0.480 |
| F (Tukey) 4 | - | 0.487 | 0.460 | 0.323 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomes, D.A.; Machado, T.G.; Maciel, G.M.; Siquieroli, A.C.S.; de Oliveira, C.S.; de Sousa, L.A.; da Silva, H.P. Dwarf Tomato Plants Allow for Managing Agronomic Yield Gains with Fruit Quality and Pest Resistance through Backcrossing. Agronomy 2022, 12, 3087. https://doi.org/10.3390/agronomy12123087
Gomes DA, Machado TG, Maciel GM, Siquieroli ACS, de Oliveira CS, de Sousa LA, da Silva HP. Dwarf Tomato Plants Allow for Managing Agronomic Yield Gains with Fruit Quality and Pest Resistance through Backcrossing. Agronomy. 2022; 12(12):3087. https://doi.org/10.3390/agronomy12123087
Chicago/Turabian StyleGomes, Danilo Araújo, Tardele Gomes Machado, Gabriel Mascarenhas Maciel, Ana Carolina Silva Siquieroli, Camila Soares de Oliveira, Luciana Alves de Sousa, and Humberto Pereira da Silva. 2022. "Dwarf Tomato Plants Allow for Managing Agronomic Yield Gains with Fruit Quality and Pest Resistance through Backcrossing" Agronomy 12, no. 12: 3087. https://doi.org/10.3390/agronomy12123087