

Biocontrol Potential of Endophytic Fungi for the Eco-Friendly Management of Root Rot of Cuminum cyminum Caused by Fusarium solani

 ,
,  ,
, 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Morphological Characterization of Pathogen
2.2. Pathogenicity Screening and Selection of Virulent Isolate
2.3. Isolation of Fungal Endophytes
2.4. Screening of Endophytes for Biocontrol Potential
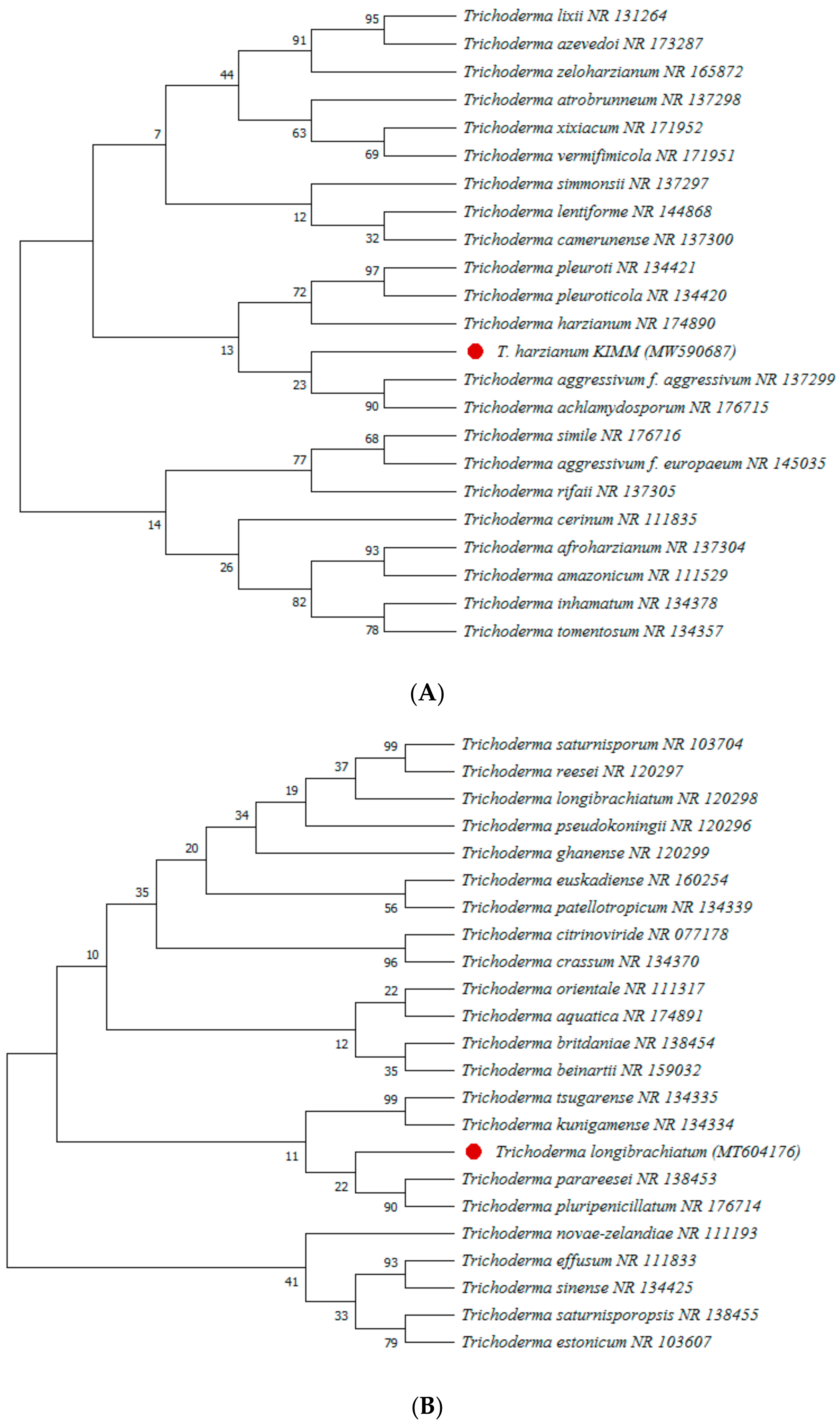
2.5. Molecular Identification of Selected Pathogen and Endophyte
2.6. In Vitro Quantification of Siderophore and Indole Acetic Acid Production by Bioagents
2.6.1. Siderophore Detection
2.6.2. Indole-3-Acetic Acid Quantification
2.7. In Vitro Culture Filtrate Assay
2.7.1. Preparation of Culture Filtrates
2.7.2. Effect of Endophyte Culture Filtrates on Cumin Seed Germination
2.7.3. In Vivo Effects of Culture Filtrates
2.8. Effect of Culture Filtrates on Defense Enzymes
- Protein Assay
- Estimation of Defense Enzymes
2.8.1. Peroxidase (PO) Assay
2.8.2. Phenylalanine Ammonia-Lyase (PAL) Assay
2.8.3. Polyphenol Oxidase (PPO) Assay
2.8.4. Total Phenolic Assay
2.9. Statistical Analysis
3. Results
3.1. Isolation and Morphological Characterization of Pathogen
3.2. In Vivo Screening, Selection, and Identification of Pathogen
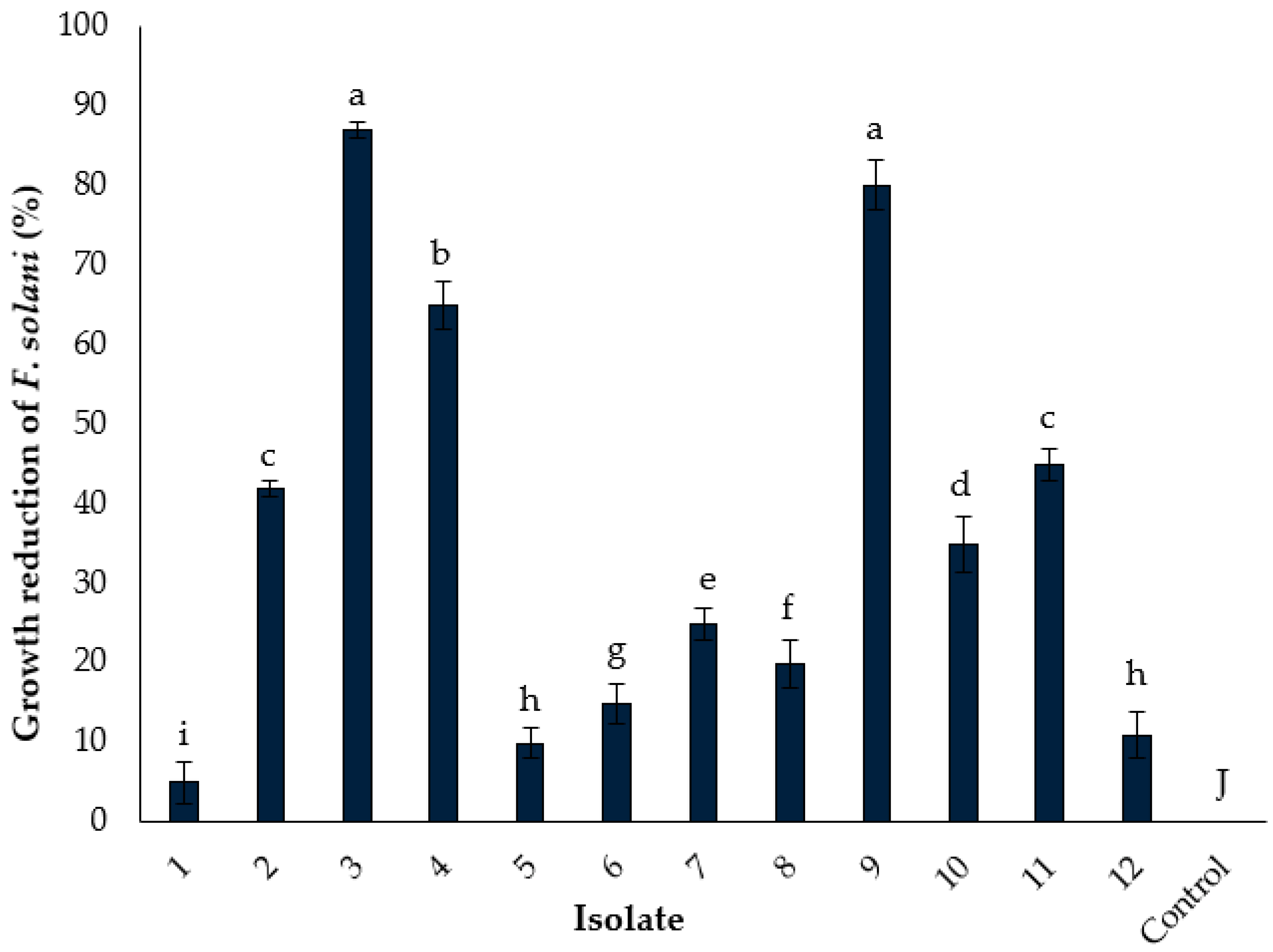
3.3. Isolation and In Vitro Screening of Fungal Endophytes for Biocontrol Potential
3.4. In Vitro Quantification of Siderophore and Indole-3-Acetic Acid Production by Endophyte Antagonists
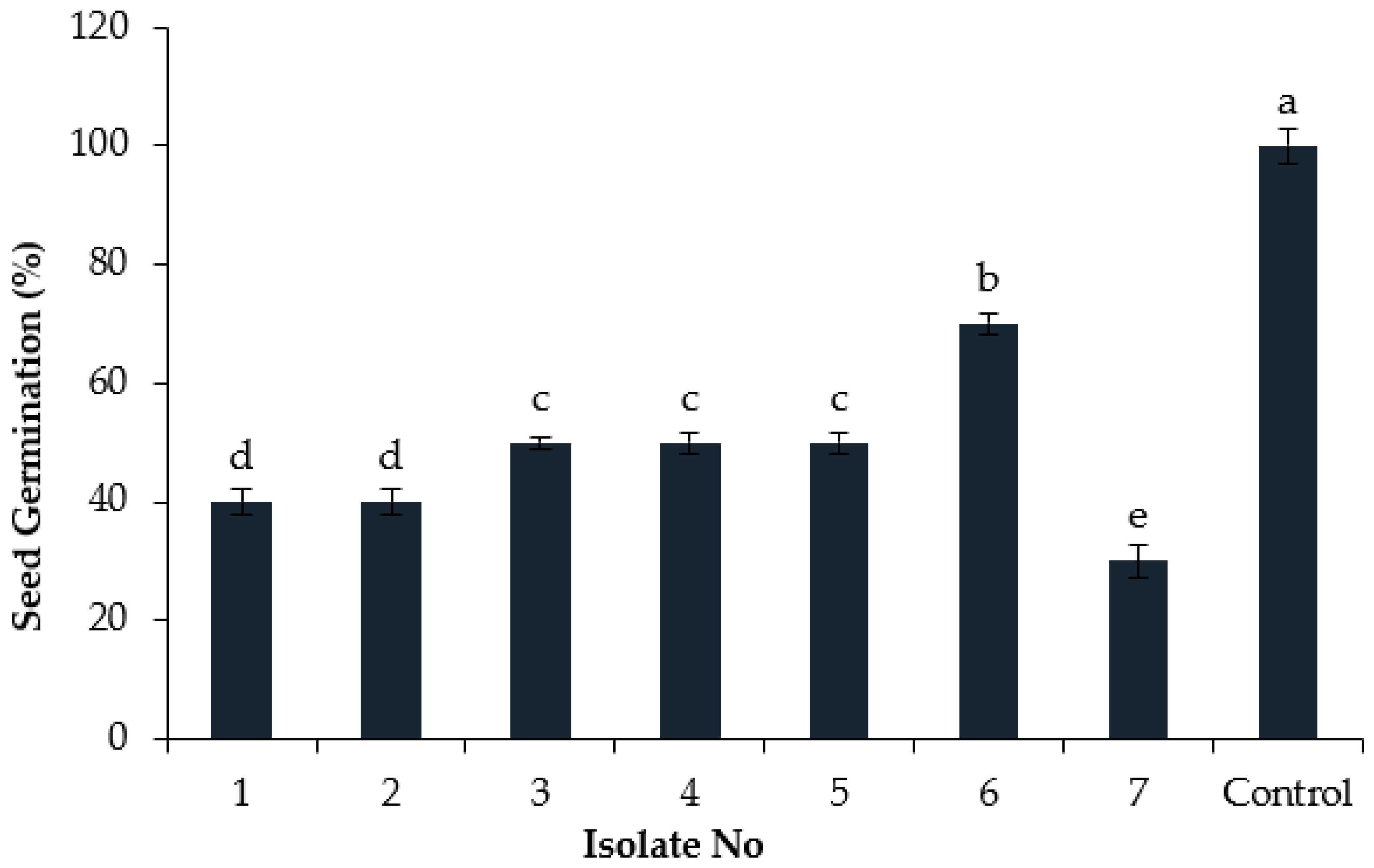
3.5. Effect of Culture Filtrates on Cumin Seed Germination
3.6. In Vivo Effect of Culture Filtrates
3.7. Estimation of Defense-Related Enzymes
3.7.1. Peroxidase (PO)
3.7.2. Phenylalanine Ammonia-Lyase (PAL)
3.7.3. Polyphenol Oxidase (PPO)
3.7.4. Phenolic Content
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rebey, I.B.; Kefi, S.; Bourgou, S.; Ouerghemmi, I.; Ksouri, R.; Tounsi, M.S.; Marzouk, B. Ripening stage and extraction method effects on physical properties, polyphenol composition and antioxidant activities of cumin (Cuminumcyminum L.) seeds. Plant Foods Hum. Nutrit. 2014, 69, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Mnif, S.; Aifa, S. Cumin (Cuminum cyminum L.) from traditional uses to potential biomedical applications. Chem. Biodivers 2015, 12, 733–742. [Google Scholar] [CrossRef] [PubMed]
- Rakesh, P.; Misra, A.K.; Singh, H.B.; Alok, K.; Dinesh, S. Diseases of Medicinal and Aromatic Plants and Their Management; Today and Tomorrow Printers and Publisher: New Delhi, India, 2019; pp. 339–352. [Google Scholar]
- Mehl, H.L.; Epstein, L. Identification of Fusarium solani f. sp. cucurbitae race 1 and race 2 with PCR and production of disease free pumpkin seeds. Plant Dis. 2007, 91, 1288–1292. [Google Scholar] [PubMed] [Green Version]
- Amer, M.A.; Salem, S.S.; Ahmed, M.A.K.; Deiaa, A.E.; Hossam, M.F.; Amr, H.H. Potential of biosynthesized zinc oxide nanoparticles to control Fusarium wilt disease in eggplant (Solanum melongena) and promote plant growth. Biometals 2022, 35, 601–616. [Google Scholar]
- Chanon, S.; Rungthip, S.; Toan, L.T.; Piyaporn, P.; Natthiya, B. Induced resistance against Fusarium solani root rot disease in cassava plant (Manihot esculenta Crantz) promoted by salicylic acid and Bacillus subtilis. Acta Agr. Sca. Section B-Soil Plant Sci. 2022, 72, 516–526. [Google Scholar]
- Yaojun, D.; Binbin, T.; Mimi, H.; Lili, W.; Kai, W.; Sixia, Y.; Jianfeng, L.; Hong, Y.; Changmei, W.; Fang, Y.; et al. High concentrations of antagonistic bacterial strains from diseased sanqi ginseng rhizosphere suppressed Fusarium root rot. Eur. J. Plant Pathol. 2022, 163, 143–153. [Google Scholar]
- Romberg, M.K.; Davis, R.M. Host Range and Phylogeny of Fusarium solani f. sp. eumartii from Potato and Tomato in California. Plant Dis. 2007, 91, 585–592. [Google Scholar] [CrossRef] [Green Version]
- Tuncturk, R.; Tuncturk, M. Effects of different phosphorus levels on the yield and quality components of cumin (Cuminum cyminum L.). Res. J. Agr. Biol. Sci. 2006, 2, 336–340. [Google Scholar]
- Imran, M.; Esmat, F.A.; Abo-Elyousr, K.A.M.; Sallam, N.M.A.; Khan, M.M.M.; Younas, M.W. Characterization and sensitivity of Botrytis cinerea to Benzimidazole and SDHI fungicides and illustration of resistance profile. Aus. Plant Pathol. 2021, 50, 589–601. [Google Scholar] [CrossRef]
- Can, Z.; Imran, M.; Xiao, L.; Hu, Z.; Li, G.; Zhang, F.; Liu, X. Difenoconazole Resistance Shift in Botrytis cinereal From Tomato in China Associated with Inducible Expression of CYP51. Plant Dis. 2021, 105, 400–407. [Google Scholar]
- Zhang, C.; Diao, Y.; Wang, W.; Hao, J.; Imran, M.; Duan, H.; Liu, X. Assessing the Risk for Resistance and Elucidating the Genetics of Colletotrichum truncatum That Is Only Sensitive to Some DMI Fungicides. Front. Microbiol. 2017, 8, 1779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imran, M.; Abo-Elyousr, K.A.M.; El-Sharnouby, M.E.; Ali, E.F.; Sallam, N.M.A.; Bagy, H.M.M.K.; Abdel-Rahim, I.R. Biocontrol Potential of Trichoderma harzianum and Zinc Nanoparticles to Mitigate Gray Mold Disease of Tomato. Gesunde Pflanz. 2022. [CrossRef]
- Zhang, W.-l.; Li, L.-I.; Zu, Y.-G.; Sonia, P. Effect of priming on the germination of Peltophorumdubium seeds under water stress. J. For. Res. 2004, 15, 287–290. [Google Scholar]
- Imran, M.; Abo-Elyousr, K.A.M.; Mousa, M.A.; Saad, M.M. A study on the synergetic effect of Bacillus amyloliquefaciens and dipotassium phosphate on Alternaria solani causing early blight disease of tomato. Eur. J. Plant Pathol. 2022, 162, 63–77. [Google Scholar] [CrossRef]
- Stappler, E.; Dattenböck, C.; Tisch, D.; Schmoll, M. Analysis of light-and carbon-specific transcriptomes implicates a class of G-protein-coupled receptors in cellulose sensing. Msphere 2017, 2, e00089-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Medina, A.; VanWees, S.C.M.; Pieterse, C.M.J. Airborne signals from Trichoderma fungi stimulate iron uptake responses in roots resulting in priming of jasmonic acid-dependent defences in shoots of Arabidopsis thaliana and Solanum lycopersicum. Plant Cell Environ. 2017, 40, 2691–2705. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Li, H.Q.; Zong, S.C.; Gao, J.M.; Zhang, A.L. Chemical and bioactive diversities of the genus Chaetomium secondary metabolites. Mini-Rev. Med. Chem. 2012, 12, 127–148. [Google Scholar] [CrossRef]
- Sibounnavong, P.; Sibounnavong, P.S.; Kanokmedhakul, S.; Soytong, K. Antifungal activities of Chaetomium brasilense CB01 and Chaetomium cupreum CC03 against Fusarium oxysporum f.sp. lycopersici race 2. J. Agric. Technol. 2012, 8, 1029–1038. [Google Scholar]
- Li, H.; Xiao, J.; Gao, Y.; Tang, J.; Zhang, A.; Gao, J. Chaetoglobosins from Chaetomium globosum, an endophytic fungus in Ginkgo biloba, and their phytotoxic and cytotoxic activities. J. Agric. Food Chem. 2014, 62, 3734–3741. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, Y.; Yan, W.; Cao, L.; Xiao, Y.; Ye, Y. Chaetomium globosum CDW7, a potential biological control strain and its antifungal metabolites. FEMS Microbiol. Lett. 2017, 364, fnw287. [Google Scholar] [CrossRef] [Green Version]
- Hung, P.M.; Pongnak, W.; Soytong, K.; Supatta, P. Efficacy of Chaetomium species as biological control agents against Phytophthora nicotianae root rot in citrus. Mycobiology 2015, 43, 288–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, H.-M.; Wang, M.; Gong, W.-F.; Zhang, L.-Q. The screening and identification of the biological control fungi Chaetomium spp. against wheat common root rot. FEMS Microbiol. Lett. 2018, 365, fny242. [Google Scholar] [CrossRef] [PubMed]
- Arunkumar, K.; Parthasarathy, S.; Harish, S.; Lingan, R.; Raguchander, T. Evaluating an isotonic aqueous formulation of Chaetomium globosum Kunze for the management of potato black scurf disease caused by Rhizoctonia solani Kuhn in India. J. Plant Pathol. 2022, 104, 191–202. [Google Scholar]
- Tian, Y.; Fu, X.; Zhang, G.; Zhang, R.; Kang, Z.; Gao, K.; Mendgen, K. Mechanisms in Growth-Promoting of Cucumber by the Endophytic Fungus Chaetomium globosum Strain ND35. J. Fungi 2022, 8, 180. [Google Scholar] [CrossRef]
- Yun, Z.; Heqin, Z.; Yonghao, Y.; Canming, T. Antifungal Activity of Chaetoviridin A from Chaetomium globosum CEF-082 Metabolites Against Verticillium dahliae in Cotton. Mol. Plant-Microbe Inter. 2021, 34, 758–769. [Google Scholar]
- Monika, S.; Dhriti, K.; Vipul, K.; Sheteiwy, M.S.; Ramakrishnan, M.; Landi, M.; Araniti, F.; Anket, S. Trichoderma: The “Secrets” of a Multitalented Biocontrol Agent. Plants 2020, 9, 762. [Google Scholar]
- Kredics, L.; Chen, L.; Kedves, O.; Büchner, R.; Hatvani, L.; Allaga, H.; Nagy, V.D.; Khaled, J.M.; Alharbi, N.S.; Vágvölgyi, C. Molecular tools for monitoring Trichoderma in agricultural environments. Front. Microbiol. 2018, 9, 1599. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, G.Y.; Li, X.Z.; Hu, M.; Wang, B.Y.; Ruan, B.H.; Zhou, H.; Zhao, L.X.; Zhou, J.; Ding, Z.T.; et al. Phytotoxic, antibacterial, and antioxidant activities of mycotoxins and other metabolites from Trichoderma sp. Nat. Prod. Res. 2017, 31, 2745–2752. [Google Scholar] [CrossRef]
- Kumar, S. Trichoderma: A biological weapon for managing plant diseases and promoting sustainability. Int. J. Agric. Sci. Med. Vet. 2013, 1, 106–121. [Google Scholar]
- Nandini, M.L.N.; Srinivasulu, B.; Gopal, K.; Ruth, C.; Devi, P.R.; Babu, M.R.; Padmaja, V.V. Induction of induced systemic resistance in capsicum chilli against Colletotrichum truncatum through fungal biocontrol agents. J. Agr. Ecol. 2022, 13, 99–111. [Google Scholar] [CrossRef]
- Candra, R.T.; Prasasty, V.D.; Karmawan, L.U. Biochemical Analysis of Banana Plants in Interaction between Endophytic Bacteria Kocuriarhi zophila and the Fungal Pathogen Fusarium oxysporum f. sp. cubense Tropical Race (Foc TR4). Biol. Life Sci. Forum. 2022, 11, 84. [Google Scholar]
- Vidhyasekaran, P. Fungal Pathogenesis in Plants and Crops; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar]
- Prasannath, K.; Dharmadasa, K.N.P.; De Costa, D.M.; Hemachandra, K.S. Variations of Incidence, Types of Virus Diseases and Insect Vector Populations of Tomato (Solanum lycopersicum L.), Grown in Different Agroecological Regions of Sri Lanka under Two Crop Management Systems. Trop. Agr. Res. 2015, 25, 376–395. [Google Scholar] [CrossRef]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Woo, S.L.; Lorito, M. Trichoderma-plant-pathogen interactions. Soil Biol. Biochem. 2008, 40, 1–10. [Google Scholar] [CrossRef]
- Li, L.; Steffens, J.C. Over expression of polyphenol oxidase in transgenic tomato plants results in enhanced bacterial disease resistance. Planta 2002, 215, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Kamran, M.A.; Javed, M.T.; Hayat, K.; Farooq, M.A.; Ali, N.; Ali, M.; Manghwar, H.; Jan, F.; Chaudhary, H.J. Individual and combinatorial application of Kocuriarhi zophila and citric acid on phytoextraction of multi-metal contaminated soils by Glycine max L. Environ. Exp. Bot. 2019, 159, 23–33. [Google Scholar] [CrossRef]
- Al-Jaradi, A.; Al-Mahmooli, I.; Janke, R.; Maharachchikumbura, S.; Al-Saady, N.; Al-Sadi, A.M. Isolation and identification of pathogenic fungi and oomycetes associated with beans and cowpea root diseases in Oman. PeerJ 2018, 6, e6064. [Google Scholar] [CrossRef] [Green Version]
- Booth, C. The Genus Fusarium Commonwealth Mycological Institute; CAB International: Wallingford, UK, 1971. [Google Scholar]
- Nelson, P.E.; Toussoun, T.A.; Marasas, W.F.O. Fusarium Species: An Illustrated Manual for Identification; Pennsylvania State University Press: University Park, PA, USA, 1983. [Google Scholar]
- Burgess, L.W.; Summerell, B.A.; Bullock, S.; Gott, K.P.; Backhouse, D. Laboratory Manual for Fusarium Research, 3rd ed.; University of Sydney and Botanic Garden: Sydney, Australia, 1994. [Google Scholar]
- Heidari, M.; Sadeghi, H. Germination and emergence of primed cumin (Cuminum cyminum L.) seeds with GA3 under different temperature regimes. Int. J. Biosci. 2014, 5, 266–272. [Google Scholar]
- Gams, W.; Bissett, J. Morphology and identification of Trichoderma. Basic biology, taxonomy and genetics. In Trichoderma and Glicladium; Kubicek, C.P., Harman, G.E., Eds.; Taylor & Francis Ltd.: London, UK, 1998; Volume 1, pp. 1–34. [Google Scholar]
- Rifai, E. A revision of the genus Trichoderma. Mycologia 1969, 116, 1–56. [Google Scholar]
- Onions, A.H.S.; Barron, G.L. Monophialidic Species of Paecilomyces; Commonwealth Mycological Institute: Richmond, UK, 1967. [Google Scholar]
- Bekker, T.F.; Kaiser, C.; Merwe, R.V.D.; Labuschagne, N. In-vitro inhibition of mycelial growth of several phytopathogenic fungi by soluble potassium silicate. S. Afr. J. Plant Soil 2006, 23, 169–172. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Wang, J.L.; Chen, J.; Mao, L.J.; Feng, X.X.; Zhang, C.L. Trichoderma Biodiversity of Agricultural Fields in East China Reveals a Gradient Distribution of Species. PLoS ONE 2016, 11, e0160613. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among Ascomycetes: Evidence from an RNA Polymerse II Subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. Mycologia 2013, 64, 315–322. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evolution 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Bano, N.; Musarrat, J. Characterization of a new Pseudomonas aeruginosa strain NJ-1 5 as a potential biocontrol agent. Curr. Microbiol. 2003, 46, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Gour, H.N.; Agrawal, S. A wilt toxin from Fusarium oxysporum f.sp. cumini Patel and Prasad. Curr. Sci. 1988, 57, 849–851. [Google Scholar]
- Bradford, M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Anal. Biochem. 1976, 72, 248–250. [Google Scholar] [CrossRef]
- Pütter, J. Peroxidases. In Methods of Enzymatic Analysis; Academic Press: Cambridge, MA, USA, 1974; pp. 685–690. [Google Scholar]
- Zucker, M. Sequential induction of phenylalanine ammonia-lyase and a lyase-inactivating system in potato tuber disks. Plant Physiol. 1968, 43, 365–374. [Google Scholar] [CrossRef]
- Jockusch, H. Induction of defence responses against Erwinia soft rot by an endogenous pectatelyase in potatoes. Physiol. Mol. Plant Pathol. 2002, 60, 91–100. [Google Scholar]
- Şahin, F.; Güllüce, M.; Daferera, D.; Sökmen, A.; Sökmen, M.; Polissiou, M.; Agar, G.; Özer, H. Biological activities of the essential oils and methanol extract of Origanum vulgare ssp. vulgare in the Eastern Anatolia region of Turkey. Food Control 2004, 15, 549–557. [Google Scholar] [CrossRef]
- Steel, R.G.D.; Torrie, J.H.; Dickey, D.A. Principles and Procedures of Statistics, A Biometrical Approach, 3rd ed.; McGraw Hill Book Co. Inc.: New York, NY, USA, 1996. [Google Scholar]
- Garima, B.; Indu, R.; Sanjay, K.; Vinay, S. Induction of β-1,3-glucanase and chitinase activity, cloning and their characterization in the defense response of Cuminum cyminum plant against the fungal pathogen Fusarium oxysporum. Arch. Phytopathol. Plant Prot. 2021, 54, 1617–1632. [Google Scholar]
- Mohammadi, A.; Shams-Ghahfarokhi, M.; Nazarian-Firouzabadi, F.; Kachuei, R.; Gholami-Shabani, M.; Razzaghi-Abyaneh, M. Giberellafujikuroi species complex isolated from maize and wheat in Iran: Distribution, molecular identification and fumonisin B1 in vitro biosynthesis. J. Sci. Food Agric. 2016, 96, 1333–1340. [Google Scholar] [CrossRef] [PubMed]
- Nurbailis, A.D.; Haliatur, R.; Yenny, L. Potential of culture filtrate from Trichoderma spp. as biofungicide to Colletotrichum gloeosporioides causing anthracnose disease in chili. Biodiversitas 2019, 20, 2915–2920. [Google Scholar] [CrossRef]
- Leslie, J.F.; Summerell, B.A. Fusarium laboratory workshops—A recent history. Mycotoxin Res. 2006, 22, 73. [Google Scholar] [CrossRef]
- Hider, R.C.; Kong, X. Chemistry and biology of siderophores. Nat. Prod. Rep. 2010, 27, 637–657. [Google Scholar] [CrossRef] [PubMed]
- Debbi, A.; Boureghda, H.; Monte, E.; Hermosa, R. Distribution and genetic variability of Fusariumoxysporum associated with tomato diseases in Algeria and biocontrol strategy with indigenous Trichoderma spp. Front. Microbiol. 2018, 9, 282. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.N.; Son, S.H.; Jordan, M.C.; Levin, D.B.; Ayele, B.T. Lignin biosynthesis in wheat (Triticumaestivum L.): Its response to water logging and association with hormonal levels. BMC Plant Biol. 2016, 16, 28. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, S.; Serrano, M.; Haridon, F.L.; Tjamos, S.E.; Metraux, J.P. Reactive oxygen species and plant resistance to fungal pathogens. Phytochem 2015, 112, 54–62. [Google Scholar] [CrossRef]
- Okey, E.N.; Duncan, E.J.; Sirju-charran, G.; Sreenivasan, T.N. Phytopthora canker resistance in cacao: Role of peroxidase, polyphenol oxidase and phenylalanine ammonia lyase. J. Phytopathol. 1997, 145, 295–299. [Google Scholar] [CrossRef]
- Fotoohiyan, Z.; Rezaee, S.; Bonjar, G.H.S.; Mohammadi, A.H.; Moradi, M. Induction of systemic resistance by Trichoderma harzianum isolates in pistachio plants infected with Verticillium dahliae. J. Nuts 2015, 6, 95–111. [Google Scholar]
- La Camera, S.; Gouzerh, G.; Dhondt, S.; Hoffmann, L.; Fritig, B.; Legrand, M.; Heitz, T. Metabolic reprogramming in plant innate immunity: The contributions of phenylpropanoid and oxylipin pathways. Immunol. Rev. 2004, 198, 267–284. [Google Scholar] [CrossRef] [PubMed]
- Pawel, S.; Urszula, K.; Bederska-Błaszczyk, M.; Agnieszka, G. Canavanine Increases the Content of Phenolic Compounds in Tomato (Solanum lycopersicum L.) Roots. Plants 2020, 9, 1595. [Google Scholar]
- Abo-Elyousr, K.A.M.; Ibrahim, Y.E.; Balabel, N.M. Induction of disease defensive enzymes in response to treatment with acibenzolar-S-methyl (ASM) and Pseudomonas fluorescens Pf2 and inoculated with Ralstonia solanacearum race 3, biovar2 (phylotype II). J. Phytopathol. 2012, 160, 382–389. [Google Scholar] [CrossRef]
- Youssef, K.; Roberto, S.R.; Tiepo, A.N.; Constantino, L.V.; de Resende, J.T.V.; Abo-Elyousr, K.A.M. Salt Solution Treatments Trigger Antioxidant Defense Response against Gray Mold Disease in Table Grapes. J. Fungi 2020, 6, 179. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
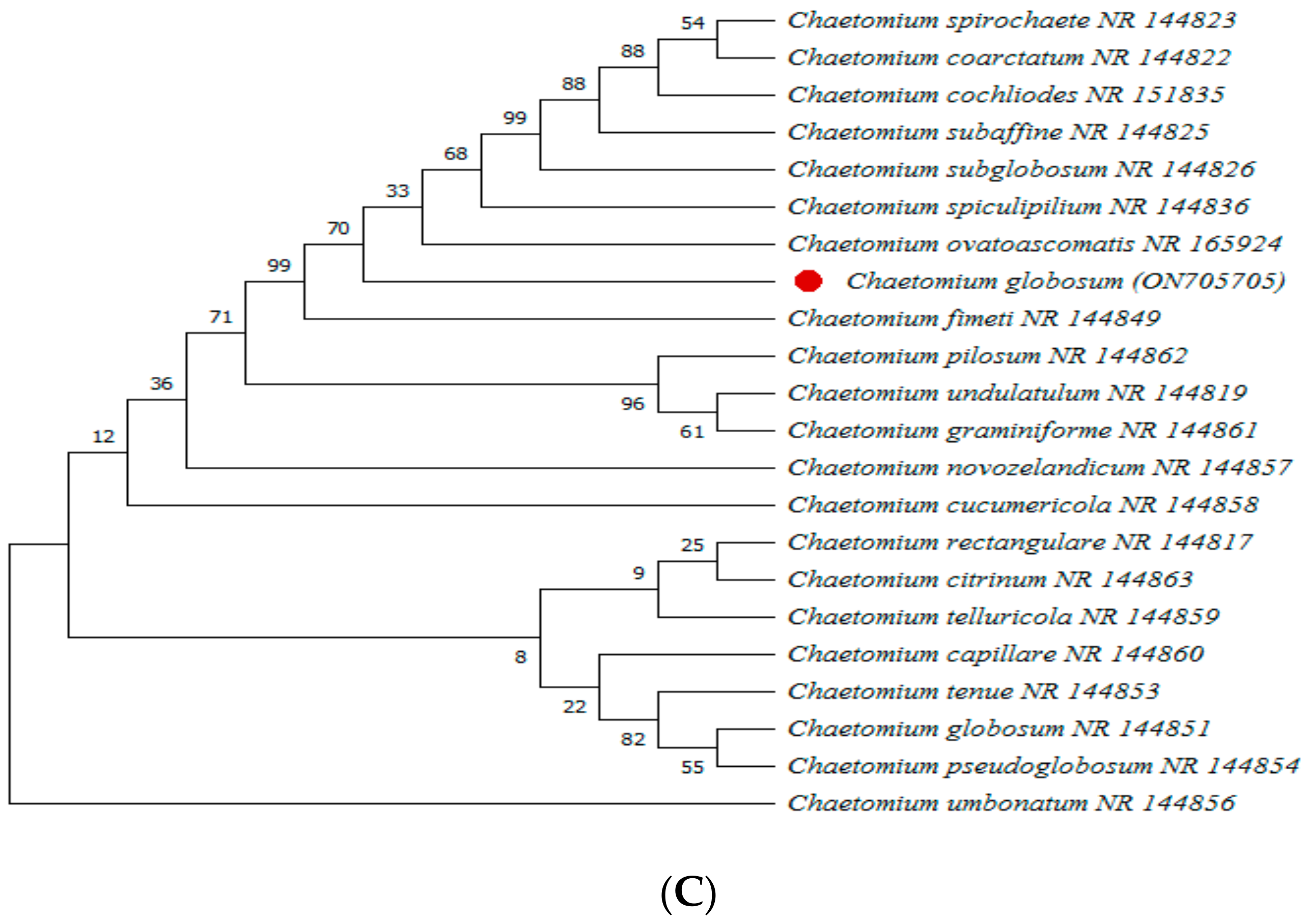{kind=link}
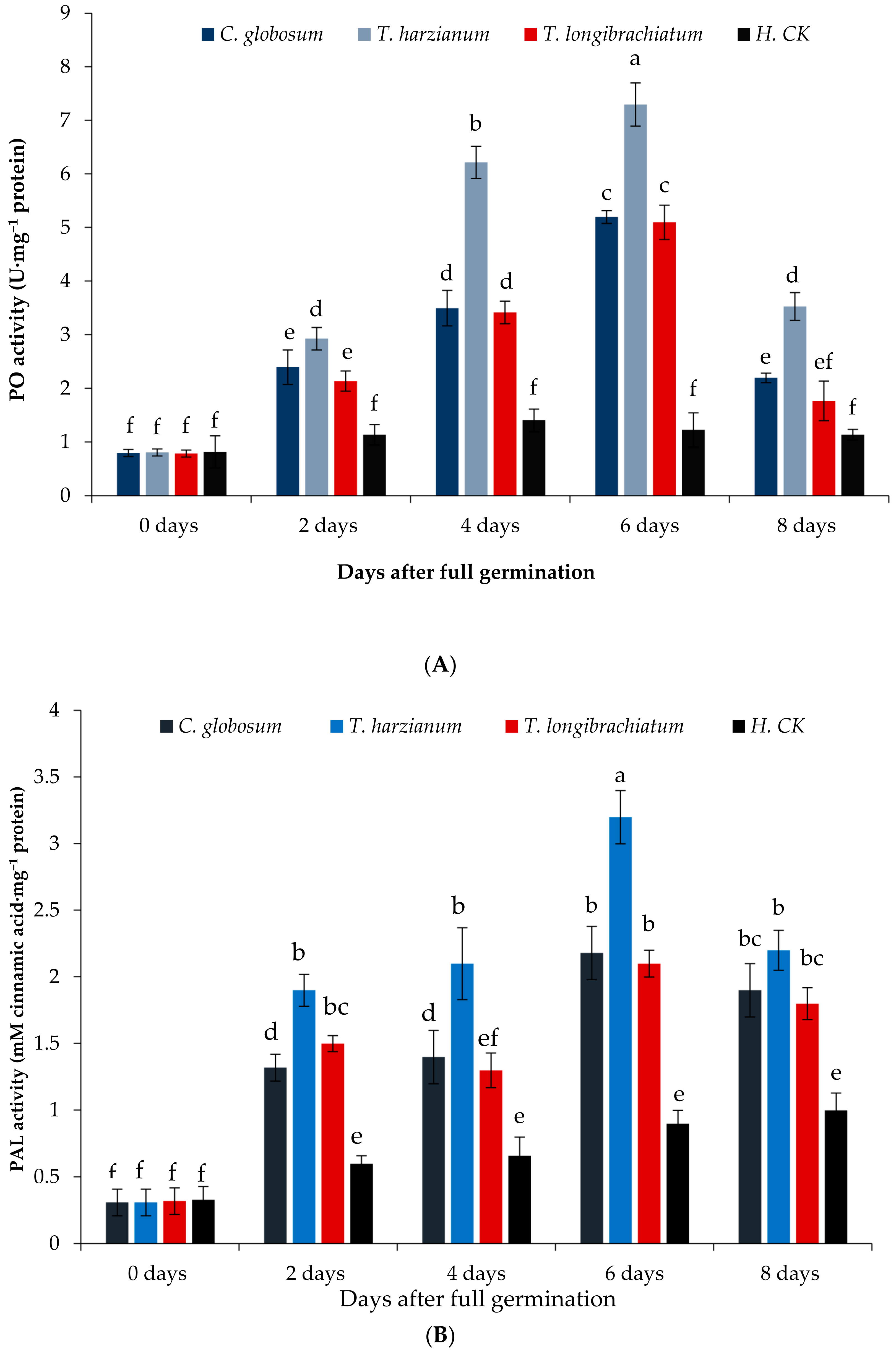{kind=link}
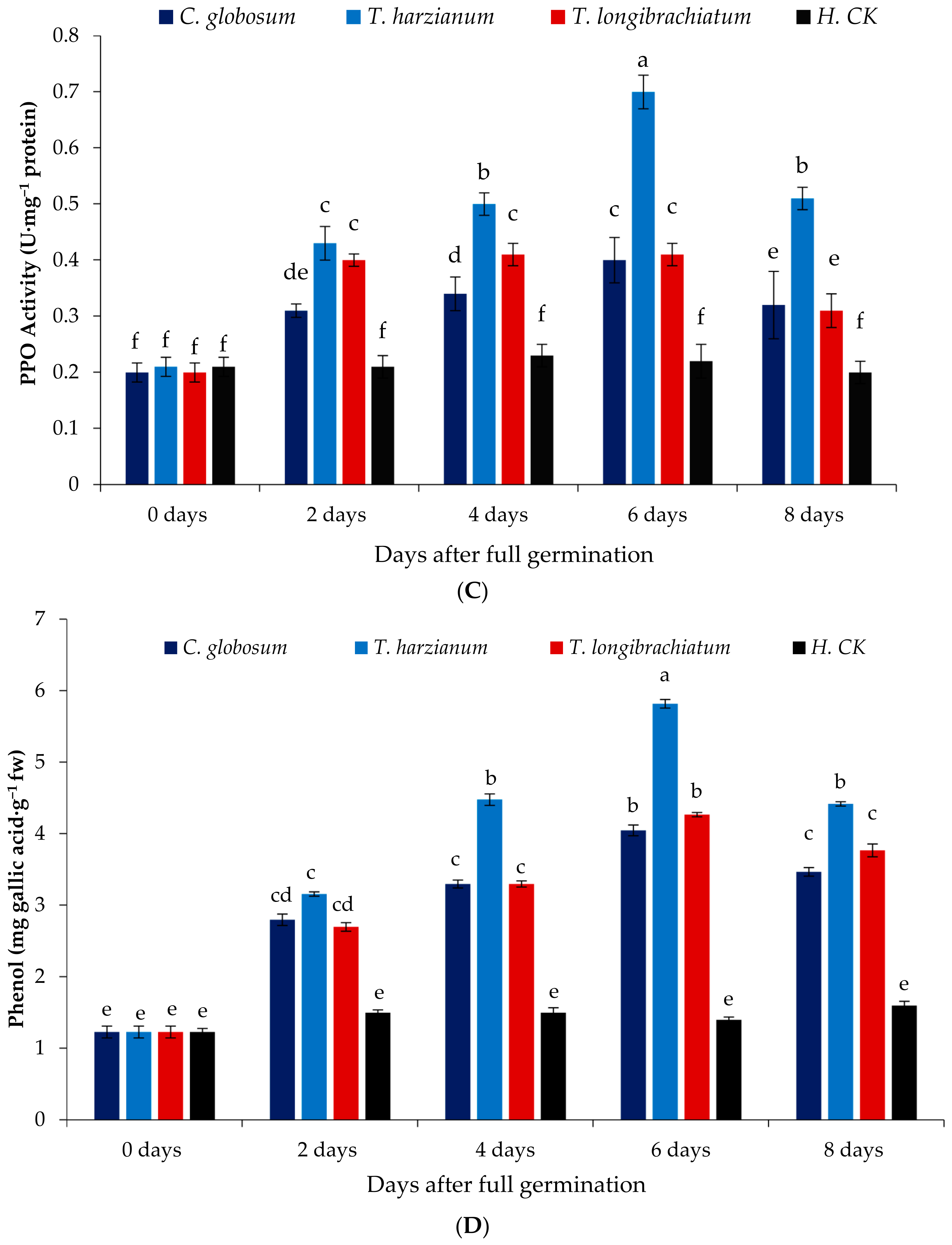{kind=link}
| Isolates | Halo Zone (mm) | IAA (μg/mL) |
|---|---|---|
| Chaetomium globosum | 3.6 ± 0.67 b | 2.5 ± 1.02 b |
| Trichoderma harzianum | 4.3 ± 1.03 a | 3.6 ± 0.48 a |
| Trichoderma longibrachiatum | 3.5 ± 0.96 b | 2.4 ± 0.87 b |
| Treatment of Culture Filtrates | Seed Germination (%) |
|---|---|
| Chaetomium globosum | 80 ± 0.46 b |
| Trichoderma harzianum | 90 ± 1.93 a |
| Trichoderma longibrachiatum | 80 ± 0.63 b |
| Infected control | 40 ± 0.39 d |
| Isolates | DS (%) | DR (%) |
|---|---|---|
| Chaetomium globosum | 25 ± 0.56 b | 59.3 |
| Trichoderma harzianum | 20 ± 0.21 c | 67.7 |
| Trichoderma longibrachiatum | 26 ± 0.87 b | 58.1 |
| Infected control | 62 ± 0.47 a | 0 |
| Healthy control | 00 ± 0.00 d | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abo-Elyousr, K.A.M.; Ibrahim, O.H.M.; Al-Qurashi, A.D.; Mousa, M.A.A.; Saad, M.M. Biocontrol Potential of Endophytic Fungi for the Eco-Friendly Management of Root Rot of Cuminum cyminum Caused by Fusarium solani. Agronomy 2022, 12, 2612. https://doi.org/10.3390/agronomy12112612
Abo-Elyousr KAM, Ibrahim OHM, Al-Qurashi AD, Mousa MAA, Saad MM. Biocontrol Potential of Endophytic Fungi for the Eco-Friendly Management of Root Rot of Cuminum cyminum Caused by Fusarium solani. Agronomy. 2022; 12(11):2612. https://doi.org/10.3390/agronomy12112612
Chicago/Turabian StyleAbo-Elyousr, Kamal A. M., Omer H. M. Ibrahim, Adel D. Al-Qurashi, Magdi A. A. Mousa, and Maged M. Saad. 2022. "Biocontrol Potential of Endophytic Fungi for the Eco-Friendly Management of Root Rot of Cuminum cyminum Caused by Fusarium solani" Agronomy 12, no. 11: 2612. https://doi.org/10.3390/agronomy12112612