
Dryocosmus kuriphilus Yasumatsu (Hymenoptera: Cynipidae) in Minho (Northern Portugal): Bioecology, Native Parasitoid Communities and Biological Control with Torymus sinensis Kamijo (Hymenoptera: Torymidae)
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Location of the Orchards under Study
2.2. Quantification of D. kuriphilus Populations
2.2.1. Framework
2.2.2. Quantification of Parasitoids and Global Rate of Parasitism
2.2.3. Identification of the Parasitoids
2.2.4. Indicators of the Parasitoids Population Structure
- n—number of individuals of a species;
- N—total number of organisms.
2.2.5. Quantification of Torimus Sinensis Populations and Effective Parasitism Rate
- no. of T. sinensis—number of T. sinensis parasitoids per sample;
- no. of galls—number of galls collected per site;
- MCG—average number of chambers per gall.
2.2.6. Assessment of the Existence of Statistically Significant Differences between Populations of D. kuriphilus
3. Results
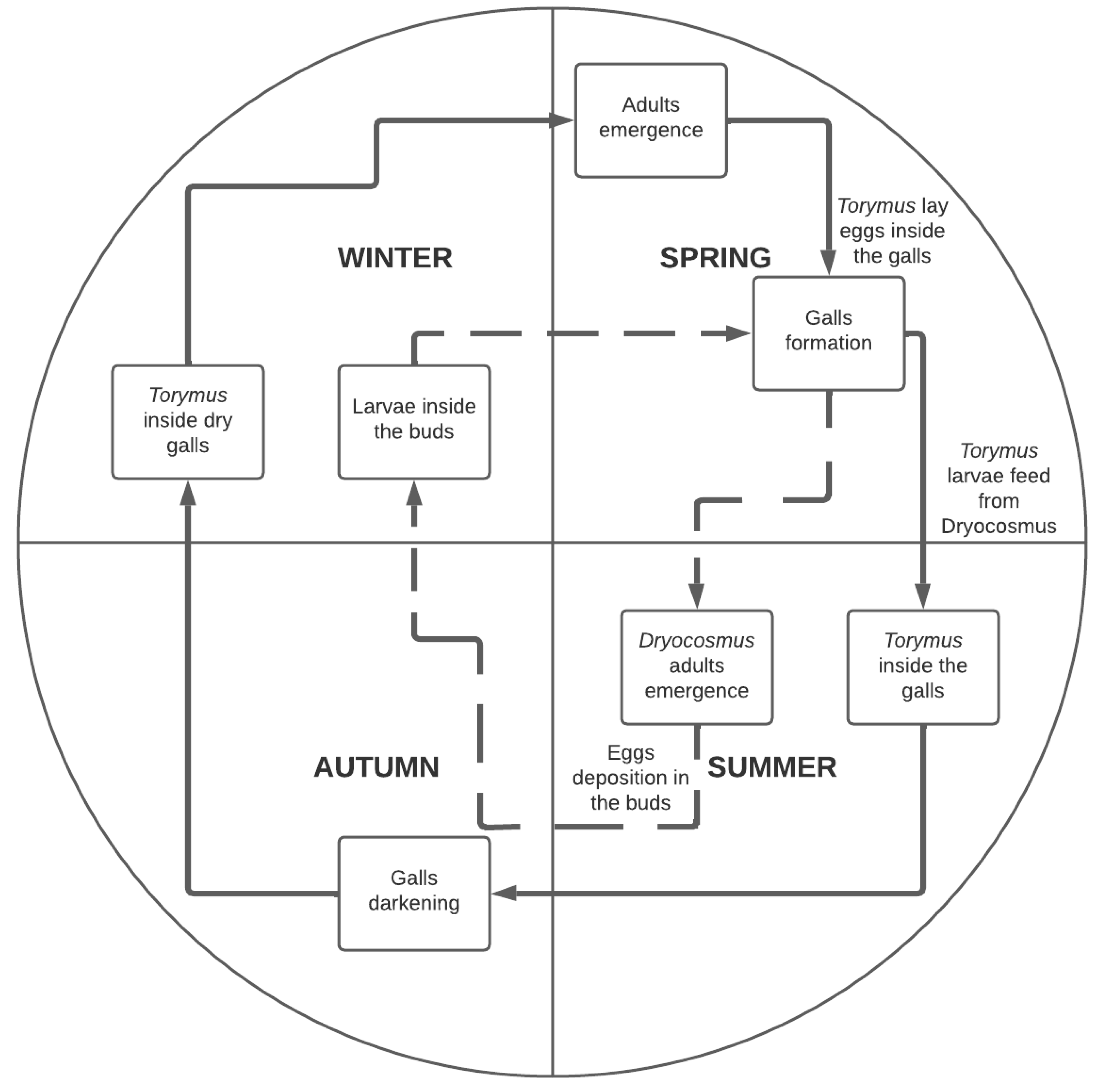
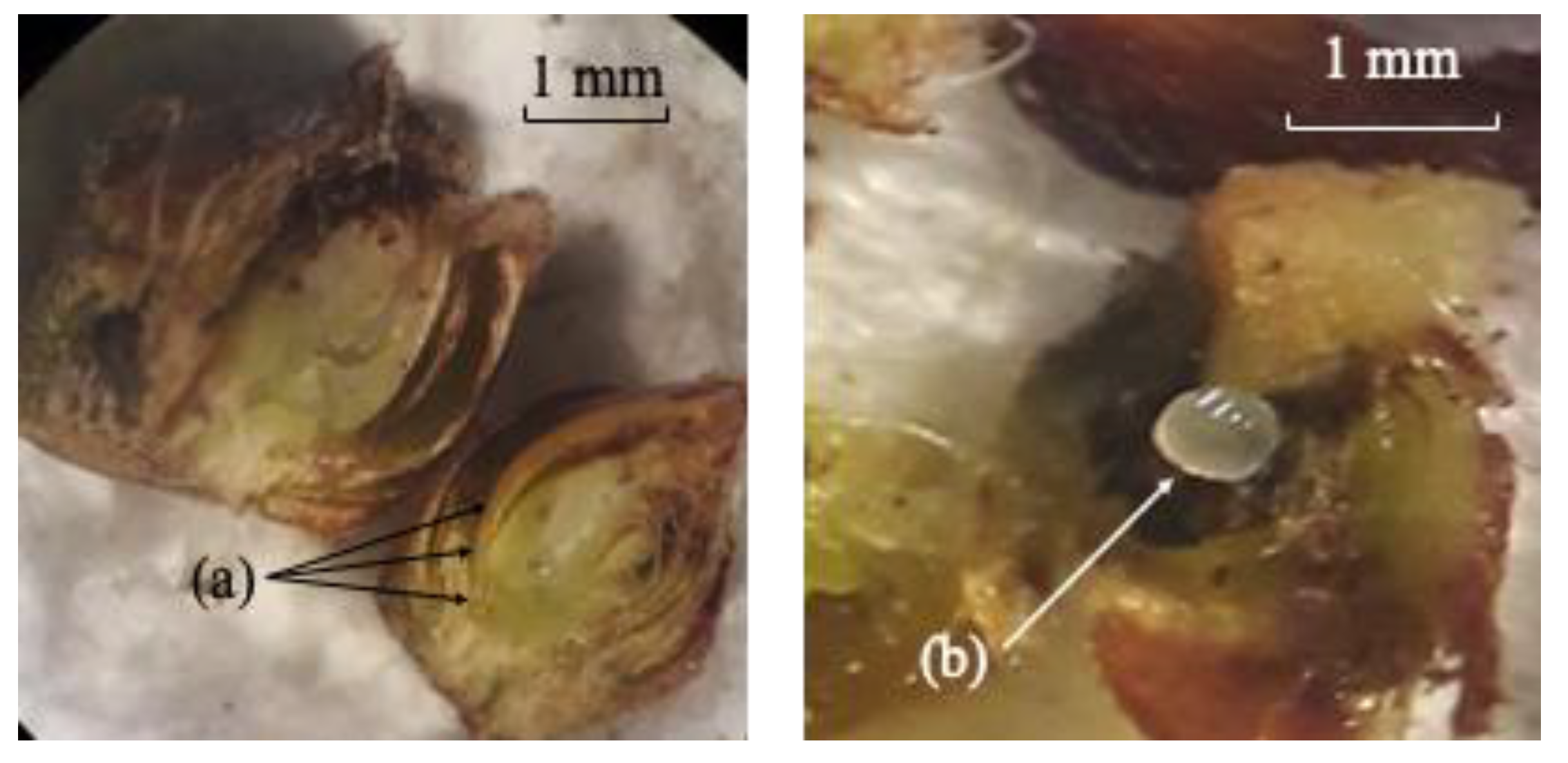
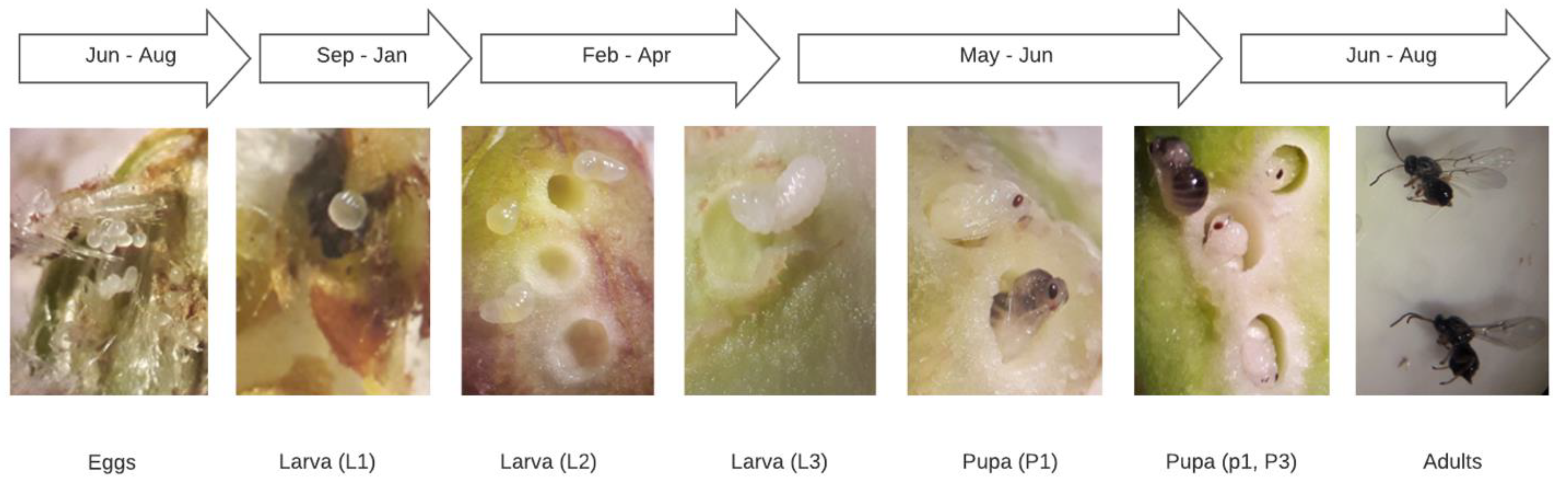
3.1. Bioecology of D. kuriphilus
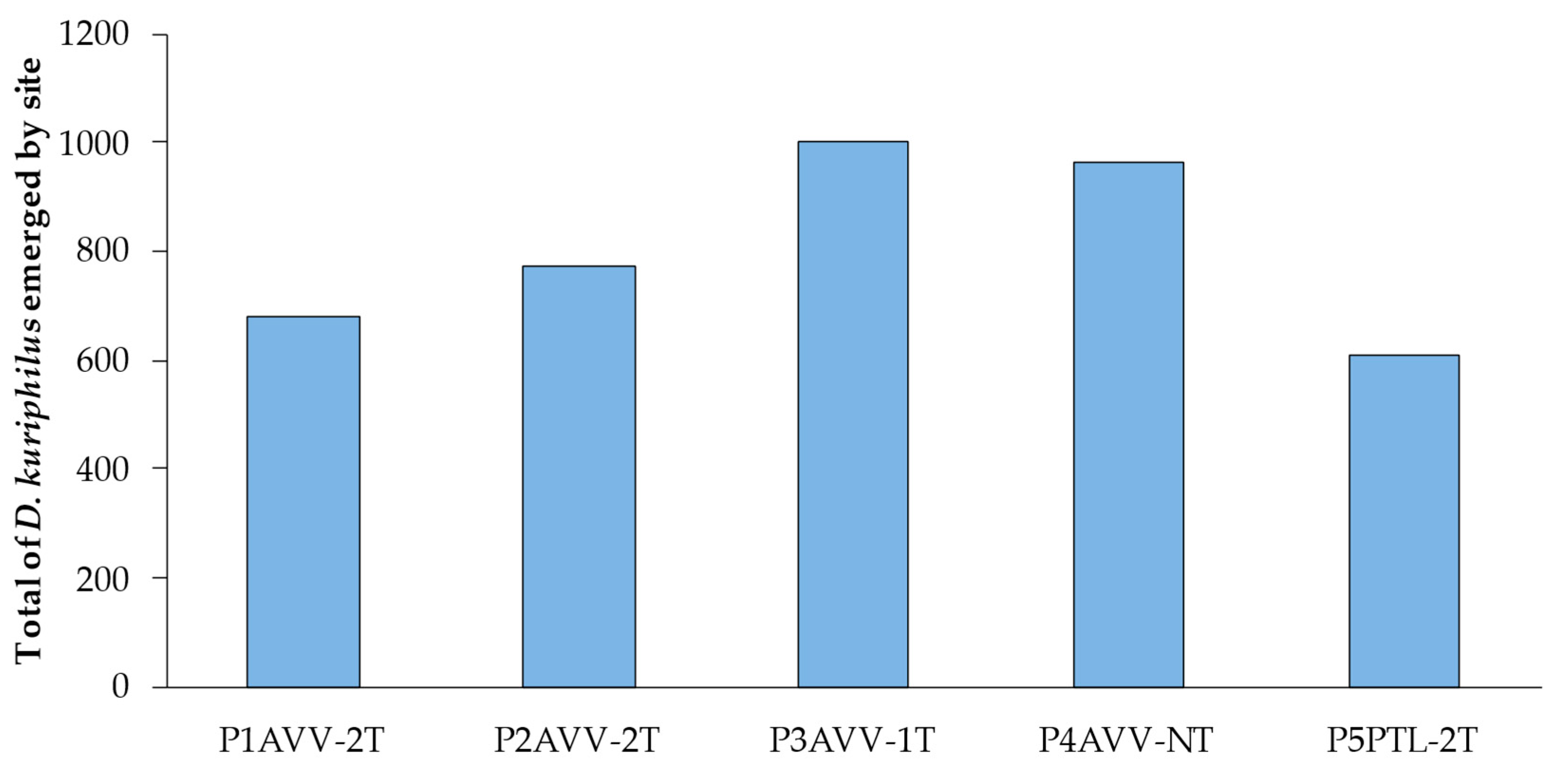
3.2. Quantification of Parasitoids and Rate of Natural Parasitism Associated with D. kuriphilus
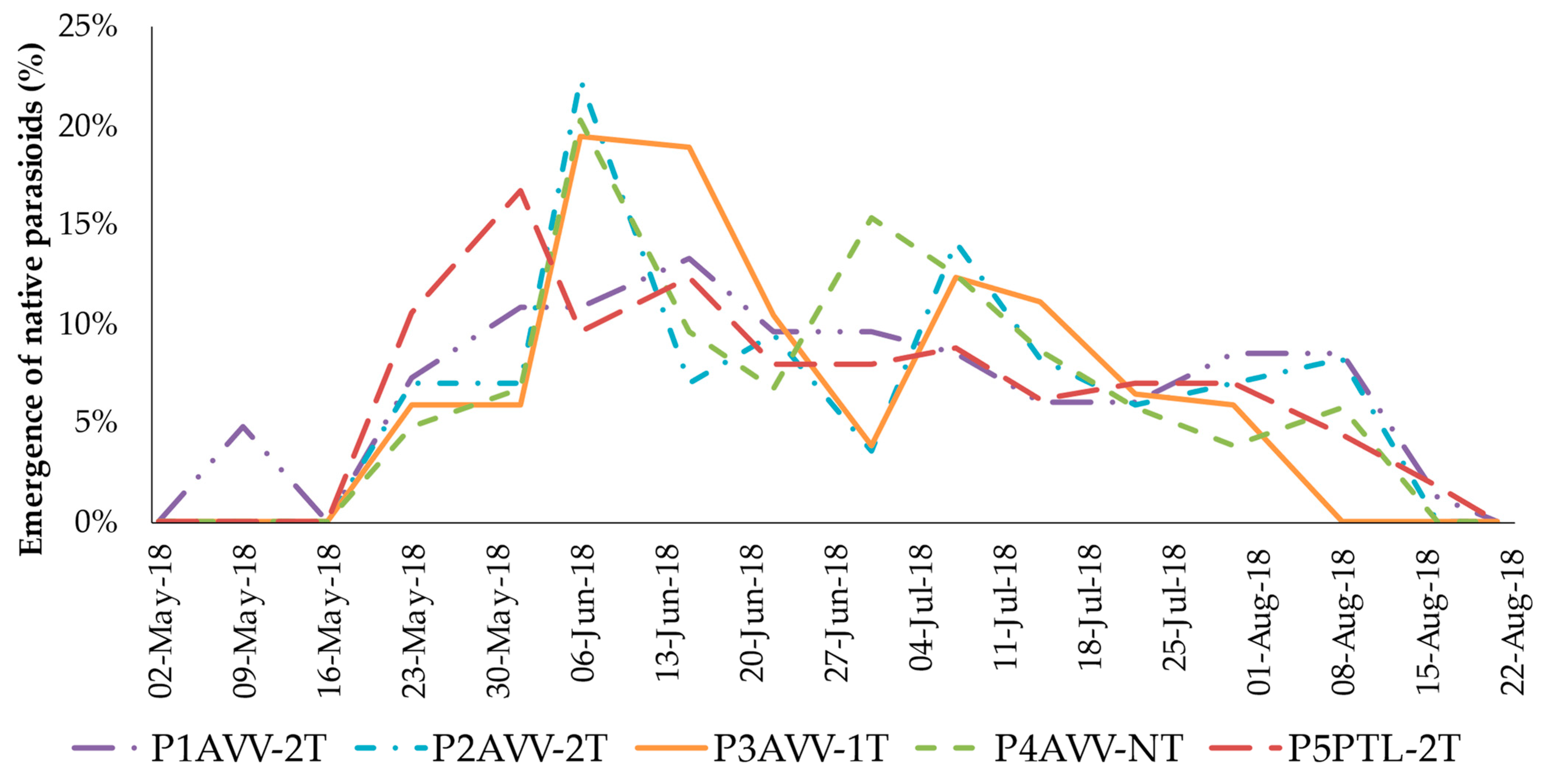
3.3. Identification and Population Dynamics of Native Parasitoids
3.4. Abundance and Specific Frequency of Native Parasitoids
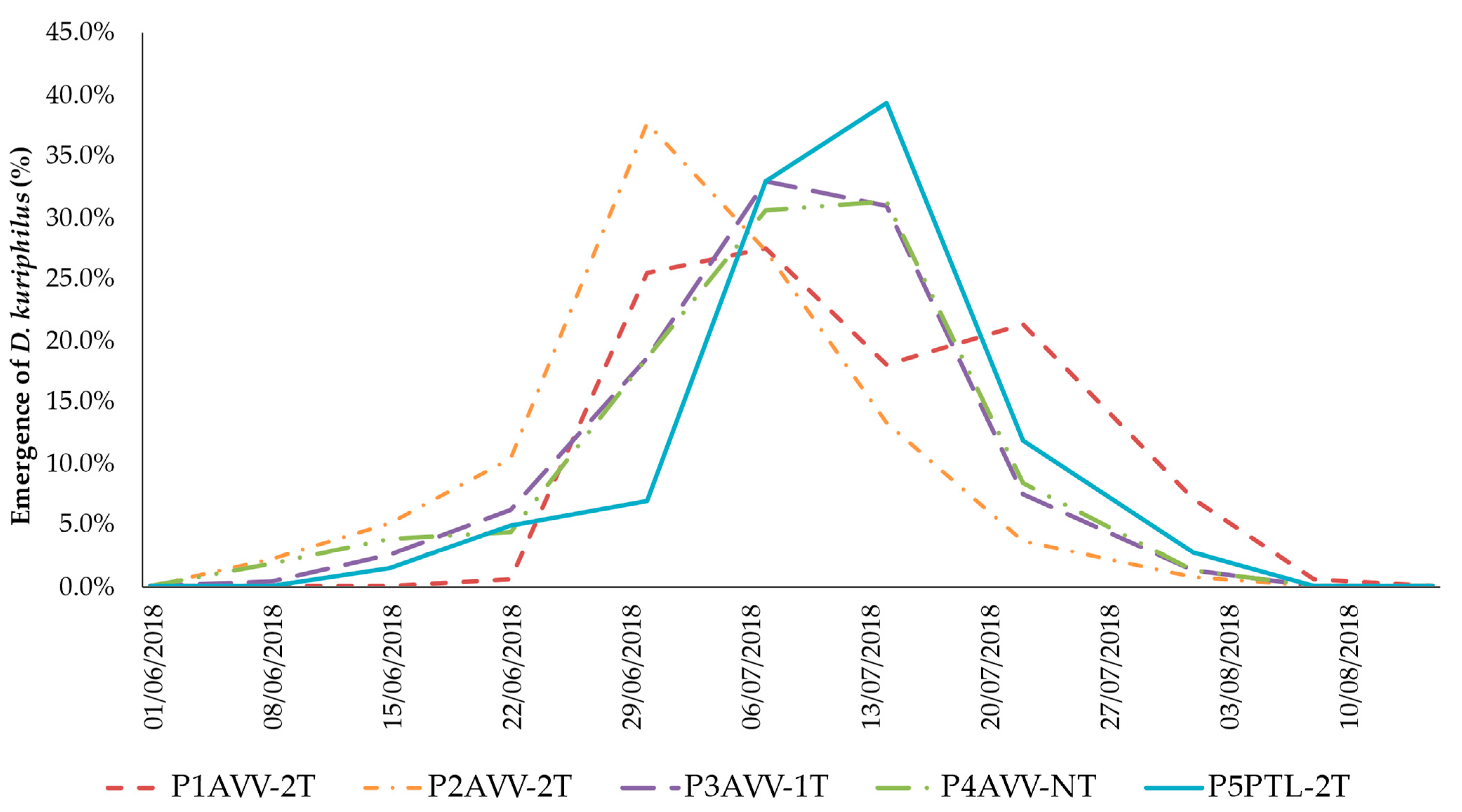
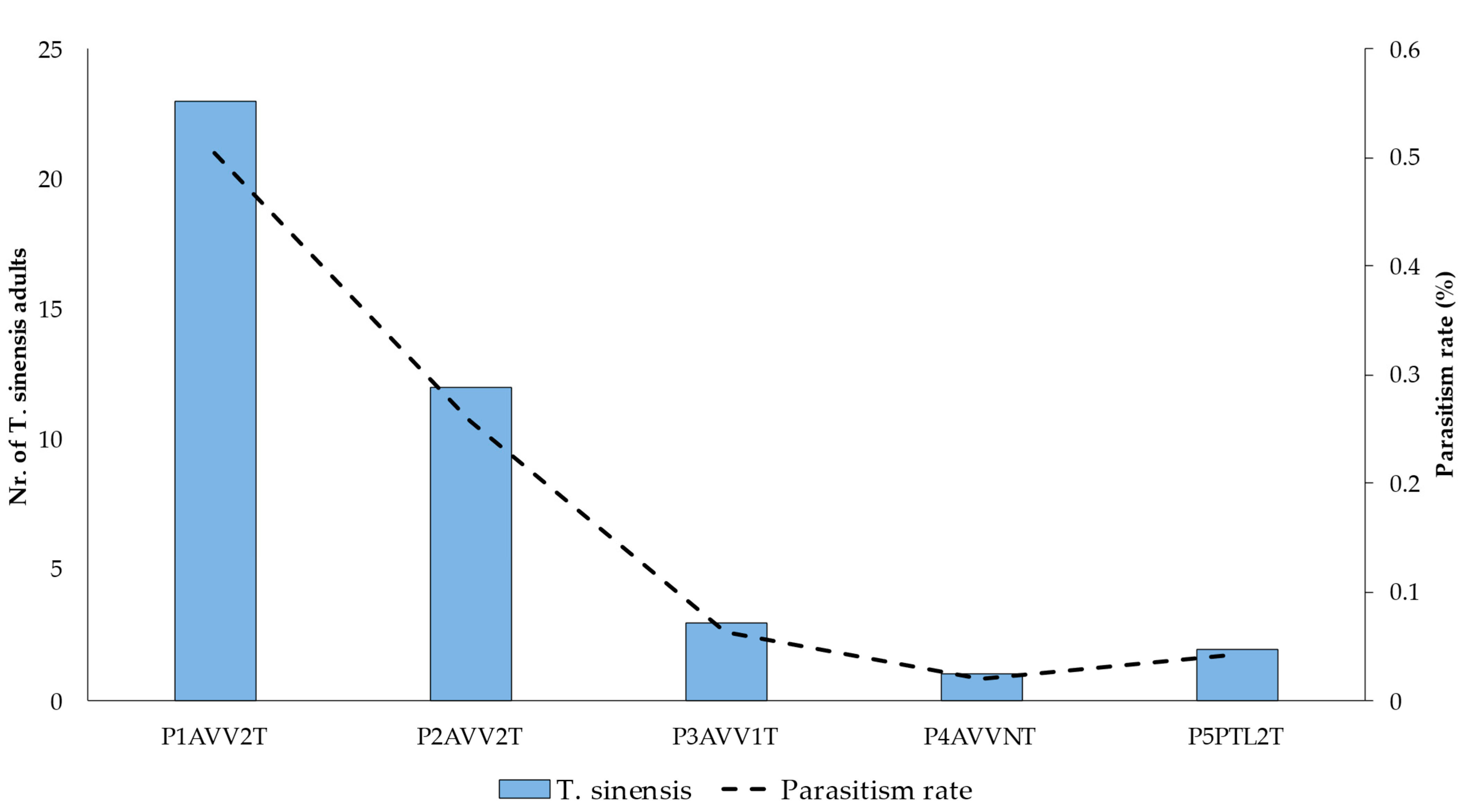
3.5. Establishment of T. sinensis Populations in Minho
4. Discussion
4.1. Bioecology of D. kuriphilus
4.2. Quantification of Parasitoids and Rate of Natural Parasitism Associated with D. kuriphilus
4.3. Identification and Population Dynamics of Native Parasitoids
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kremer, A.; Abbott, A.G.; Carlson, J.E.; Manos, P.S.; Plomion, C.; Sisco, P.; Staton, M.E.; Ueno, S.; Vendramin, G.G. Genomics of fagaceae. Tree Genet. Genomes 2012, 8, 583–610. [Google Scholar] [CrossRef]
- Barakat, A.; DiLoreto, D.S.; Zhang, Y.; Smith, C.; Baier, K.; Powell, W.A.; Wheeler, N.; Sederoff, R.; Carlson, J.E. Comparison of the transcriptomes of American chestnut (Castanea dentata) and Chinese chestnut (Castanea mollissima) in response to the chestnut blight infection. BMC Plant Biol. 2009, 9, 51. [Google Scholar] [CrossRef] [PubMed]
- Bounous, G.; Ertürk, U.; Akyuz, B.; Fulbright, D.; Serdar, U. Evaluation of the descriptive characteristics of chestnut. Acta Hortic. 2018, 1220, 35–44. [Google Scholar]
- Bounous, G.; Marinoni, D.T. Chestnut: Botany, horticulture, and utilization. Hortic. Rev. 2010, 31, 291–347. [Google Scholar]
- Pinto-Correia, T.; Vos, W. Multifunctionality in Mediterranean landscapes-past and. New Dimens. Eur. Landsc. 2004, 4, 135. [Google Scholar]
- Conedera, M.; Manetti, M.C.; Giudici, F.; Amorini, E. Distribution and economic potential of the Sweet chestnut (Castanea sativa Mill.) in Europe. Ecol. Mediterr. 2004, 30, 179–193. [Google Scholar] [CrossRef]
- Everard, J.; Christie, J. Sweet chestnut: Silviculture, timber quality and yield in the forest of Dean. For. Int. J. For. Res. 1995, 68, 133–144. [Google Scholar] [CrossRef]
- Freitas, T.R.; Santos, J.A.; Silva, A.P.; Fraga, H. Influence of climate change on chestnut trees: A review. Plants 2021, 10, 1463. [Google Scholar] [CrossRef]
- Baptista, P.; Martins, A.; Tavares, R.M.; Lino-Neto, T. Diversity and fruiting pattern of macrofungi associated with chestnut (Castanea sativa) in the Trás-os-Montes region (Northeast Portugal). Fungal Ecol. 2010, 3, 9–19. [Google Scholar] [CrossRef]
- Dinis, L.; Peixoto, F.; Pinto, T.; Costa, R.; Bennett, R.; Gomes-Laranjo, J. Study of morphological and phenological diversity in chestnut trees (‘Judia’variety) as a function of temperature sum. Environ. Exp. Bot. 2011, 70, 110–120. [Google Scholar] [CrossRef]
- Bragança, H.; Simões, S.; Onofre, N.; Santos, N. Factors influencing the incidence and spread of chestnut blight in northeastern Portugal. J. Plant Pathol. 2009, 91, 53–59. [Google Scholar]
- Franquinho Aguiar, A.M.; Cravo, D.; Dantas, L.M.; Vieira da Luz, T.M. The Oriental Chestnut Gall-wasp Dryocosmus Kuriphilus Yasumatsu, 1951,(Hymenoptera: Cynipidae) and Its Introduced and Native Parasitoids in Madeira Island, Portugal, with Notes on Some Other Parasitic Hymenoptera Reared from Gall Samples. Entomol. Mon. Mag. 2022, 158, 175–190. [Google Scholar] [CrossRef]
- Gomes-Laranjo, J.; Almeida, P.; Ferreira-Cardoso, J.; Peixoto, F. Ecophysiological characterization of C. sativa trees growing under different altitudes. Acta Hortic. 2009, 844, 119–126. [Google Scholar] [CrossRef]
- EFSA Panel on Plant Health. Risk assessment of the oriental chestnut gall wasp, Dryocosmus kuriphilus for the EU territory and identification and evaluation of risk management options. EFSA J. 2010, 8, 1619. [Google Scholar]
- Bonsignore, C.P.; Vizzari, G.; Vono, G.; Bernardo, U. Short-term cold stress affects parasitism on the Asian chestnut gall wasp Dryocosmus kuriphilus. Insects 2020, 11, 841. [Google Scholar] [CrossRef] [PubMed]
- Aebi, A.; Schönrogge, K.; Melika, G.; Alma, A.; Bosio, G.; Quacchia, A.; Picciau, L.; Abe, Y.; Moriya, S.; Yara, K. Parasitoid recruitment to the globally invasive chestnut gall wasp Dryocosmus kuriphilus. In Galling Arthropods and Their Associates; Springer: Tokyo, Japan, 2006; pp. 103–121. [Google Scholar]
- Brussino, G.; Bosio, G.; Baudino, M.; Giordano, R.; Ramello, F.; Melika, G. Pericoloso insetto esotico per il castagno europeo. Inf. Agrar. 2002, 58, 59–62. [Google Scholar]
- Quacchia, A.; Moriya, S.; Bosio, G.; Scapin, I.; Alma, A. Rearing, release and settlement prospect in Italy of Torymus sinensis, the biological control agent of the chestnut gall wasp Dryocosmus kuriphilus. BioControl 2008, 53, 829–839. [Google Scholar] [CrossRef]
- Aebi, A.; Schönrogge, K.; Melika, G.; Quacchia, A.; Alma, A.; Stone, G. Native and introduced parasitoids attacking the invasive chestnut gall wasp Dryocosmus kuriphilus. EPPO Bull. 2007, 37, 166–171. [Google Scholar] [CrossRef]
- Balsa, C.; Bento, A.; Paparella, F. Biological control of the Asian chestnut gall wasp in Portugal: Insights from a mathematical model. PLoS ONE 2021, 16, e0254193. [Google Scholar]
- Balsa, C.; Citerici, M.; Lopes, I.M.; Rufino, J. Numerical simulation of the biological control of the chestnut gall wasp with T. sinensis. In Proceedings of the ICAIW 2019 Workshops at the Second International Conference on Applies Informatics 2019, Madrid, Spain, 6–9 November 2019; Volume 2486, pp. 230–243. [Google Scholar]
- Bonal, R.; Vargas-Osuna, E.; Mena, J.D.; Aparicio, J.M.; Santoro, M.; Martín, A. Looking for variable molecular markers in the chestnut gall wasp Dryocosmus kuriphilus: First comparison across genes. Sci. Rep. 2018, 8, 5631. [Google Scholar] [CrossRef]
- Cardoso, J.F.S. The Endemic Parasitoids of the Chestnut Gall Wasp Dryocosmus kuriphilus Yasumatsu (Hymenoptera: Cynipidae) in Portugal and Biocontrol with Torymus Sinensis Kamijo (Hymenoptera: Torymidae). Master’s Thesis, Universidade do Minho, Braga, Portugal, 2020. [Google Scholar]
- Quacchia, A.; Moriya, S.; Askew, R.; Schönrogge, K. Torymus sinensis: Biology, host range and hybridization. Acta Hortic. 2014, 1043, 105–111. [Google Scholar] [CrossRef]
- Van Lenteren, J.C.; Bolckmans, K.; Köhl, J.; Ravensberg, W.J.; Urbaneja, A. Biological control using invertebrates and microorganisms: Plenty of new opportunities. BioControl 2018, 63, 39–59. [Google Scholar] [CrossRef]
- Piao, C.-S.; Moriya, S. Oviposition of Torymus sinensis Kamijo (Hymenoptera: Torymidae) under natural conditions. Entomol. Sci. 1999, 2, 329–334. [Google Scholar]
- Gibbs, M.; Schönrogge, K.; Alma, A.; Melika, G.; Quacchia, A.; Stone, G.N.; Aebi, A. Torymus sinensis: A viable management option for the biological control of Dryocosmus kuriphilus in Europe? BioControl 2011, 56, 527–538. [Google Scholar] [CrossRef]
- Borowiec, N.; Thaon, M.; Brancaccio, L.; Cailleret, B.; Ris, N.; Vercken, E. Early population dynamics in classical biological control: Establishment of the exotic parasitoid T orymus sinensis and control of its target pest, the chestnut gall wasp Dryocosmus kuriphilus, in F rance. Entomol. Exp. Appl. 2018, 166, 367–379. [Google Scholar] [CrossRef]
- Avtzis, D.N.; Melika, G.; Matošević, D.; Coyle, D.R. The Asian chestnut gall wasp Dryocosmus kuriphilus: A global invader and a successful case of classical biological control. J. Pest Sci. 2019, 92, 107–115. [Google Scholar] [CrossRef]
- Santos, A.L.; Santos, S.A.P.; Almeida, J.; Bento, A. Parasitoides autóctones associados a Dryocosmus kuriphilus Yasumatsu: Principais espécies e taxas de parasitismo. Rev. Ciências Agrárias 2018, 41, 141–144. [Google Scholar]
- Marsh, P.M. Hymenoptera of the world: An identification guide to families. Am. Entomol. 1994, 40, 115–116. [Google Scholar] [CrossRef]
- Morris, E.K.; Caruso, T.; Buscot, F.; Fischer, M.; Hancock, C.; Maier, T.S.; Meiners, T.; Müller, C.; Obermaier, E.; Prati, D. Choosing and using diversity indices: Insights for ecological applications from the German Biodiversity Exploratories. Ecol. Evol. 2014, 4, 3514–3524. [Google Scholar] [CrossRef] [Green Version]
- DRAP-Norte. Vespa das Galhas do Castanheiro (Dryocosmus kuriphilus) Luta Biológica Largadas de Torymus Sinensis; DRAP-Norte—Divisão de Apoio ao Setor Agroalimentar: Porto, Portugal, 2020.
- Nieves-Aldrey, J.L.; Gil-Tapetado, D.; Gavira, O.; Boyero, J.R.; Polidori, C.; Lombardero, M.J.; Blanco, D.; Rojo, P.R.; Vela, J.M.; Wong, E. Torymus sinensis Kamijo, a biocontrol agent against the invasive chestnut gall wasp Dryocosmus kuriphilus Yasumatsu in Spain: Its natural dispersal from France and the first data on establishment after experimental releases. For. Syst. 2019, 28, e001. [Google Scholar] [CrossRef]
- DGAV. Plano de Ação Nacional para Controlo do Inseto Dryocosmus Kuriphilus Yasumatsu (Vespa das Galhas do Castanheiro Relatório de Progresso Nº3); DGAV: Lisbon, Portugal, 2020; p. 30.
- Do Nascimento Lopes, N. A Vespa das Galhas dos Castanheiros-Relação com os Parâmetros Ambientais. Master’s Thesis, Faculdade de Ciências e Tecnologia, Universidade Nova de Lisboa, Lisbon, Portugal, 2018. [Google Scholar]
- DGAV. Plano de Ação Nacional para Controlo do Inseto Dryocosmus Kuriphilus Yasumatsu (Vespa das Galhas do Castanheiro); Direção Geral de Alimentação e Veterinária: Lisbon, Portugal, 2017; p. 26.
- Santos, A.; Pereira, J.; Santos, S.; Quacchia, A.; Bento, A. Biological control based on native parasitoids associated with Dryocosmus kuriphilus Yasumatsu in invaded countries. Nat. Enemies 2017, 1–23. [Google Scholar]
- Cardoso, J.; Almeida, M.T.; Bento, A. Biological control of Dryocosmus kuriphilus Yasumatsu with the parasitoid Torymus sinensis Kamijo. Millenium 2021, 14, 91–99. [Google Scholar]
- Santi, F.; Maini, S. New association between Dryocosmus kuriphilus and Torymus flavipes in chestnut trees in the Bologna area (Italy): First results. Bull. Insectology 2011, 64, 275–278. [Google Scholar]
- Murakami, Y.; Ohkubo, N.; Moriya, S.; Gyoutoku, Y.; Kim, H.C.; Kim, K.J. Parasitoids of Dryocosmus kuriphilus (Hymenoptera: Cynipidae) in South Korea with particular reference to ecologically different types of Torymus (Syntomaspis) sinensis (Hymenoptera: Torymidae). Appl. Entomol. Zool. 1995, 30, 277–284. [Google Scholar] [CrossRef]
- Stone, G.N.; Schönrogge, K.; Atkinson, R.J.; Bellido, D.; Pujade-Villar, J. The population biology of oak gall wasps (Hymenoptera: Cynipidae). Annu. Rev. Entomol. 2002, 47, 633–668. [Google Scholar] [CrossRef]
- Francati, S.; Alma, A.; Ferracini, C.; Pollini, A.; Dindo, M.L. Indigenous parasitoids associated with Dryocosmus kuriphilus in a chestnut production area of Emilia Romagna (Italy). Bull. Insectology 2015, 68, 127–134. [Google Scholar]
- Bosio, G.; Gerbaudo, C.; Piazza, E. Dryocosmus kuriphilus Yasumatsu: An outline seven years after the first report in Piedmont (Italy). Acta Hortic. 2009, 866, 341–348. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Orchard | Area (ha) | Location (Parish, Municipality) | Varieties | Nr. of Releases T. sinensis | Year of the Releases |
|---|---|---|---|---|---|
| P1AVV2T | 1.22 | Rio Cabrão (Arcos de Valdevez) | Amarelal | 2 | 2016 2017 |
| P2AVV2T | 0.29 | Távora S. Maria (Arcos de Valdevez) | Marigoule | 2 | 2016 2017 |
| P3AVV1T | 0.35 | Rio de Moinhos (Arcos de Valdevez) | Amarelal | 1 | 2017 |
| Marsol | |||||
| P4AVVNT | 1.10 | S. Paio (Arcos de Valdevez) | Amarelal | 0 | |
| Bouche Betizac | ----- | ||||
| Marigoule | |||||
| P5PTL2T | 1.83 | Gondufe (Ponte de Lima) | Marigoule | 2 | 2016 2017 |
| Location | No. of Buds | % of Infested Buds | Average No. of Larvae/Buds ± Standard Deviation |
|---|---|---|---|
| P1AVV2T | 200 | 78.0% | 2.54 ± 2.03 |
| P2AVV2T | 200 | 80.0% | 2.66 ± 2.02 |
| P3AVV1T | 200 | 74.0% | 2.90 ± 2.30 |
| P4AVVNT | 200 | 77.5% | 2.92 ± 2.33 |
| P5PTL2T | 300 | 67.0% | 2.04 ± 1.93 |
| Location | No. of Dissected Galls | No. of D. kuriphilus Chambers | Average No. of Chambers Per Gall ± Standard Deviation |
|---|---|---|---|
| P1AVV2T | 240 | 1514 | 6.31 ± 2.46 |
| P2AVV2T | 240 | 1544 | 6.43 ± 2.49 |
| P3AVV1T | 240 | 1541 | 6.42 ± 3.22 |
| P4AVVNT | 240 | 1676 | 6.98 ± 3.65 |
| P5PTL2T | 240 | 1487 | 6.20 ± 2.90 |
| Location | Total No. of Galls | No. of Parasitized Galls | % of Parasitized Galls |
|---|---|---|---|
| P1AVV2T | 240 | 89 | 37% |
| P2AVV2T | 240 | 123 | 51% |
| P3AVV1T | 240 | 82 | 34% |
| P4AVVNT | 240 | 103 | 43% |
| P5PTL2T | 240 | 100 | 42% |
| Location | Total Chambers | No. of Paratized Chambers | % Parasitized Chambers |
|---|---|---|---|
| P1AVV-2T | 1514 | 114 | 7.5% |
| P2AVV-2T | 1544 | 150 | 9.7% |
| P3AVV-1T | 1541 | 104 | 6.7% |
| P4AVV-NT | 1676 | 133 | 7.9% |
| P5AVV-2T | 1487 | 118 | 7.9% |
| Parasitoid Native Species | Chestnut Orchards | ||||
|---|---|---|---|---|---|
| P1AVV-2T | P2AVV-2T | P3AVV1T | P4AVV-NT | P5PTL-2T | |
| Eupelmus azureos (Ratzeburg, 1844) | A | A | P | P | P |
| Eupelmus uruzonus (Dalman, 1820) | P | P | P | P | P |
| Eurytoma brunnivientris (Ratzeburg, 1852) | P | P | P | P | P |
| Eurytoma pistaciae (Rondani, 1877) | A | A | A | P | A |
| Megastigmus dorsalis (Fabricius, 1798) | P | P | P | P | P |
| Mesopolobus sp. (Westwood, 1833) | P | P | P | P | P |
| Ormyrus pomaceus (Geoffroy, 1785) | P | P | P | P | P |
| Sycophila biguttata (Swederus, 1795) | A | P | A | P | A |
| Sycophila iracemae (Nieves Aldrey, 1984) | P | P | P | P | P |
| Sycophila variegatta (Curtis, 1831) | P | P | P | P | P |
| Torymus flavipes (Walker, 1833) | P | P | P | P | P |
| No. of species | 8 | 8 | 9 | 11 | 9 |
| Species | Family |
|---|---|
| E. azureos | Eupelmidae |
| E. urozonus | |
| Mesopolobus sp. | Pteromalidae |
| O. pomaceus | Ormyridae |
| E. brunnivientris | Eurytomidae |
| E. pistaciae | |
| S. biguttata | |
| S. iracemae | |
| S. variegatta | |
| M. dorsalis | Torymidae |
| T. flavipes |
| Parasitoid Native Species | No. of Insects | M | % |
|---|---|---|---|
| E. azureos | 5 | 3 | 60% |
| E. uruzonus | 14 | 5 | 100% |
| E. brunnivientris | 17 | 5 | 100% |
| E. pistaciae | 2 | 1 | 20% |
| M. dorsalis | 24 | 5 | 100% |
| Mesopolobus sp. | 44 | 5 | 100% |
| O. pomaceus | 39 | 5 | 100% |
| S. biguttata | 4 | 2 | 40% |
| S. iracemae | 190 | 5 | 100% |
| S. variegatta | 34 | 5 | 100% |
| T. flavipes | 57 | 5 | 100% |
| Native Species | P1AVV-2T | P2AVV-2T | P3AVV-1T | P4AVV-NT | P5PTL-2T | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| NoA | SDI | NoA | SDI | NoA | SDI | NoA | SDI | NoA | SDI | |
| Eupelmus azureos | 0 | 0.0000 | 0 | 0.0000 | 1 | 0.0000 | 2 | 0.0003 | 2 | 0.0003 |
| Eupelmus uruzonus | 3 | 0.0019 | 5 | 0.0028 | 1 | 0.0000 | 2 | 0.0003 | 3 | 0.0008 |
| Eurytoma brunnivientris | 1 | 0.0000 | 2 | 0.0003 | 7 | 0.0031 | 6 | 0.0045 | 1 | 0.0000 |
| Eurytoma pistacina | 0 | 0.0000 | 0 | 0.0000 | 0 | 0.0000 | 2 | 0.0003 | 0 | 0.0000 |
| Megastigmus dorsalis | 3 | 0.0019 | 4 | 0.0017 | 4 | 0.0009 | 5 | 0.0030 | 8 | 0.0072 |
| Mesopolobus sp. | 9 | 0.0226 | 4 | 0.0017 | 5 | 0.0015 | 10 | 0.0136 | 16 | 0.0306 |
| Ormyrus pomaceus | 1 | 0.0000 | 3 | 0.0008 | 20 | 0.0280 | 7 | 0.0063 | 8 | 0.0072 |
| Sycophila biguttata | 0 | 0.0000 | 3 | 0.0008 | 0 | 0.0000 | 1 | 0.0000 | 0 | 0.0000 |
| Sycophila iracemae | 18 | 0.0959 | 45 | 0.2773 | 63 | 0.2878 | 34 | 0.1689 | 29 | 0.1037 |
| Sycophila variegatta | 6 | 0.0094 | 9 | 0.0101 | 9 | 0.0053 | 6 | 0.0045 | 5 | 0.0026 |
| Torymus flavipes | 16 | 0.0752 | 10 | 0.0126 | 7 | 0.0031 | 7 | 0.0063 | 17 | 0.0347 |
| Total | 57 | 0.2068 | 85 | 0.3081 | 117 | 0.3296 | 82 | 0.208 | 89 | 0.187 |
| SDI | 0.793 | 0.692 | 0.670 | 0.792 | 0.813 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amorim, A.; Rodrigues, R.; Nunes, L.J.R.; Freitas, M.; Moura, L. Dryocosmus kuriphilus Yasumatsu (Hymenoptera: Cynipidae) in Minho (Northern Portugal): Bioecology, Native Parasitoid Communities and Biological Control with Torymus sinensis Kamijo (Hymenoptera: Torymidae). Agronomy 2022, 12, 2184. https://doi.org/10.3390/agronomy12092184
Amorim A, Rodrigues R, Nunes LJR, Freitas M, Moura L. Dryocosmus kuriphilus Yasumatsu (Hymenoptera: Cynipidae) in Minho (Northern Portugal): Bioecology, Native Parasitoid Communities and Biological Control with Torymus sinensis Kamijo (Hymenoptera: Torymidae). Agronomy. 2022; 12(9):2184. https://doi.org/10.3390/agronomy12092184
Chicago/Turabian StyleAmorim, Amaro, Raúl Rodrigues, Leonel J. R. Nunes, Mariano Freitas, and Luísa Moura. 2022. "Dryocosmus kuriphilus Yasumatsu (Hymenoptera: Cynipidae) in Minho (Northern Portugal): Bioecology, Native Parasitoid Communities and Biological Control with Torymus sinensis Kamijo (Hymenoptera: Torymidae)" Agronomy 12, no. 9: 2184. https://doi.org/10.3390/agronomy12092184