
Responses of Cell Wall Components to Low Nitrogen in Rapeseed Roots


1
College of Resources and Environmental Sciences, Hunan Agricultural University, Changsha 410128, China
2
College of Resources and Environmental Sciences, Qingdao Agricultural University, Qingdao 266109, China
*
Author to whom correspondence should be addressed.
Agronomy 2022, 12(5), 1044; https://doi.org/10.3390/agronomy12051044
Submission received: 15 February 2022
/
Revised: 7 March 2022
/
Accepted: 2 April 2022
/
Published: 27 April 2022
(This article belongs to the Special Issue Breeding for Climate Change Adaptation through Tolerance to Abiotic Stresses)
Abstract
:Rapeseed (Brassica napus L.) is a major oil crop in China, with the world’s largest planted area and total yield. Rapeseed has a high demand for nitrogen (N), and nitrogen deficiency in soil is an important limiting factor for rapeseed production. However, rapeseed responds to N deprivation by regulating its own morphology, structure, and physiology. We carried out the current experiment by utilizing low N (LN: 0.3 mM NO3−) and normal N (CK: 6.0 mM NO3−) treatments using Brassica napus as the experimental material. The study results showed that low N induced root elongation in rapeseed, and the root length of LN treatment was 2.37 times that of HN treatment. The dry matter of roots also significantly increased due to low N treatment. Meanwhile, low N treatment decreased photosynthetic pigment (including chlorophyll a, chlorophyll b, and carotenoids) contents and dry mass accumulation of leaves. A higher root/shoot ratio and N physiological efficiency were observed under low N treatment. The changes in cell wall components (pectin, cellulose, hemicellulose, and lignin), related enzymes, and genes’ transcription levels in roots were determined and the results suggested that low N promoted the demethylation of ion-bound pectin (ISP) and covalently bound pectin (CSP), the content of CSP and cellulose. The promoted pectin methylesterase (PME) activity, inhibited pectin and cellulose degradation enzymes, and up/downregulation of related genes also confirming the results of cell wall components. The low N-increased demethylation degree of pectin and content of pectin and cellulose in cell walls was conducive to cell wall loosening and cell wall synthesis during cell division and elongation, ultimately promoting root-adaptive elongation. The study revealed a possible mechanism in which the alteration of cell wall component content and structure participates in cell elongation and expansion, which directly induces root elongation under N deficiency. The successful implementation of this research may be conducive to facilitating the development of rapeseed cultivars with high N use efficiency through root-based genetic improvements and improving plant adaptability to low N.
1. Introduction
As an important oil crop with the highest oil production, rapeseed (Brassica napus L.) plays an important role in the supply of edible oil in China. Nitrogen (N), also known as “bioelement”, is an essential macroelement for plant growth and development. Nitrogen participates in many important physiological metabolic processes in plants [1]. In agricultural production, rapeseed has a large demand for N fertilizer, and the N deficiency in soil is an important limiting factor for rapeseed production. The N use efficiency of rapeseed is also at a low level [2,3]. Therefore, farmers often increase the application of N fertilizer to obtain a high yield in actual production, while the application of N fertilizer not only promotes agricultural production but also causes many problems for the environment and crops. Excessive N fertilizer causes soil acidification, hardening, and water eutrophication through surface runoff and leaching loss. At the same time, excess N induces crop overgrowth, aggravates crop diseases and lodging, and reduces grain yield and quality. Plant adaptation to N deficiency is crucial for improving crop N efficiency and reducing chemical N fertilization.
Under low N stress, plant roots make morphological, structural, and physiological adaptation responses, which could be a compensatory effect of plants, hunting for N sources as much as feasible. Nitrogen deficiency-induced changes in root architecture are very important for enhancing the N acquisition of plants [4,5]. A lot of previous studies have focused on the adaptive response of plant roots to low N and found that low N increases root dry weight, total root length, and the change of root structure [6], and significantly increases root/shoot ratio [7,8,9]. The morphological and physiological responses of roots to changes in N application have been widely reported [10,11]. The length of a root is determined by the combination of cell division, cell differentiation, and the extent of expansion and elongation of cells [12,13]. The plant cell wall is a highly dynamic, complex network structure and is involved in various cell metabolic activities, including maintaining cell adhesion, cell elongation, and expansion [14]. Under biotic and abiotic stresses, cell wall is differently modified in structure and composition by the activation of a wide range of enzymes [15,16,17], and the modification in cell wall structure and composition is closely related to cell wall loosening, which is a direct cause of cells expansion and plant growth [18]. A lot of studies have focused on revealing the mechanism of roots with low N in many plants, with known responses of auxin transporter and auxin signaling, cell division and expansion, and the abundance of proteins involved in cell differentiation, cell wall modification, and phenylpropanoid biosynthesis [19,20,21,22]. However, how low N affects the structure and components of cell walls in rapeseed roots remains largely unknown.
Therefore, in the present experiment, Brassica napus (L.) was used as the research material, and we aimed to suggest the compositional response of rapeseed root cell walls to low N and reveal the relationship of cell wall components and root elongation under N deficiency. The results provide scientific and theoretical guidance for breeding rapeseed varieties with high N use efficiency and benefits to realizing “zero growth” of chemical fertilizer.
2. Results
2.1. Effects of Low N on the Growth and Photosynthetic Pigment Content in Rapeseed
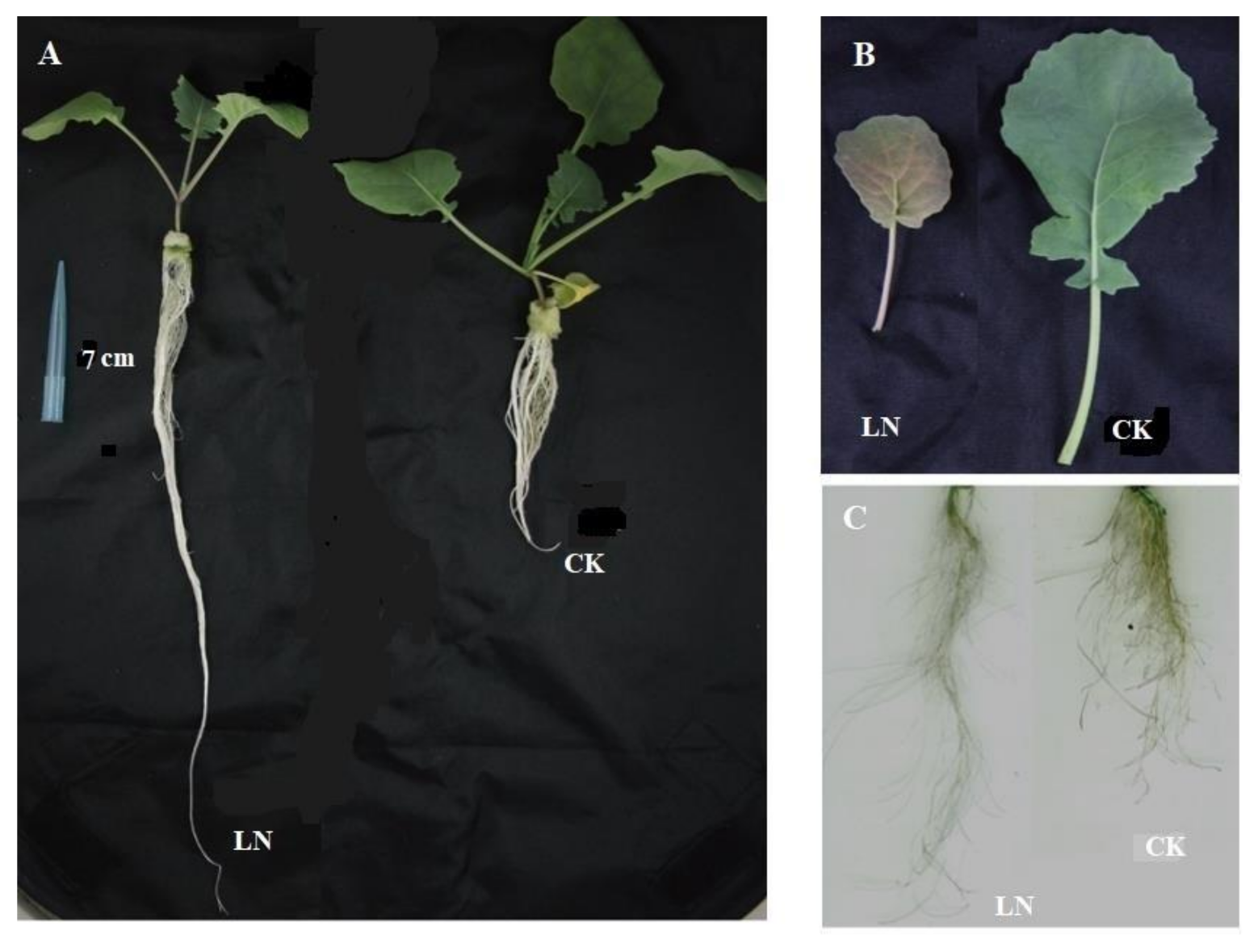
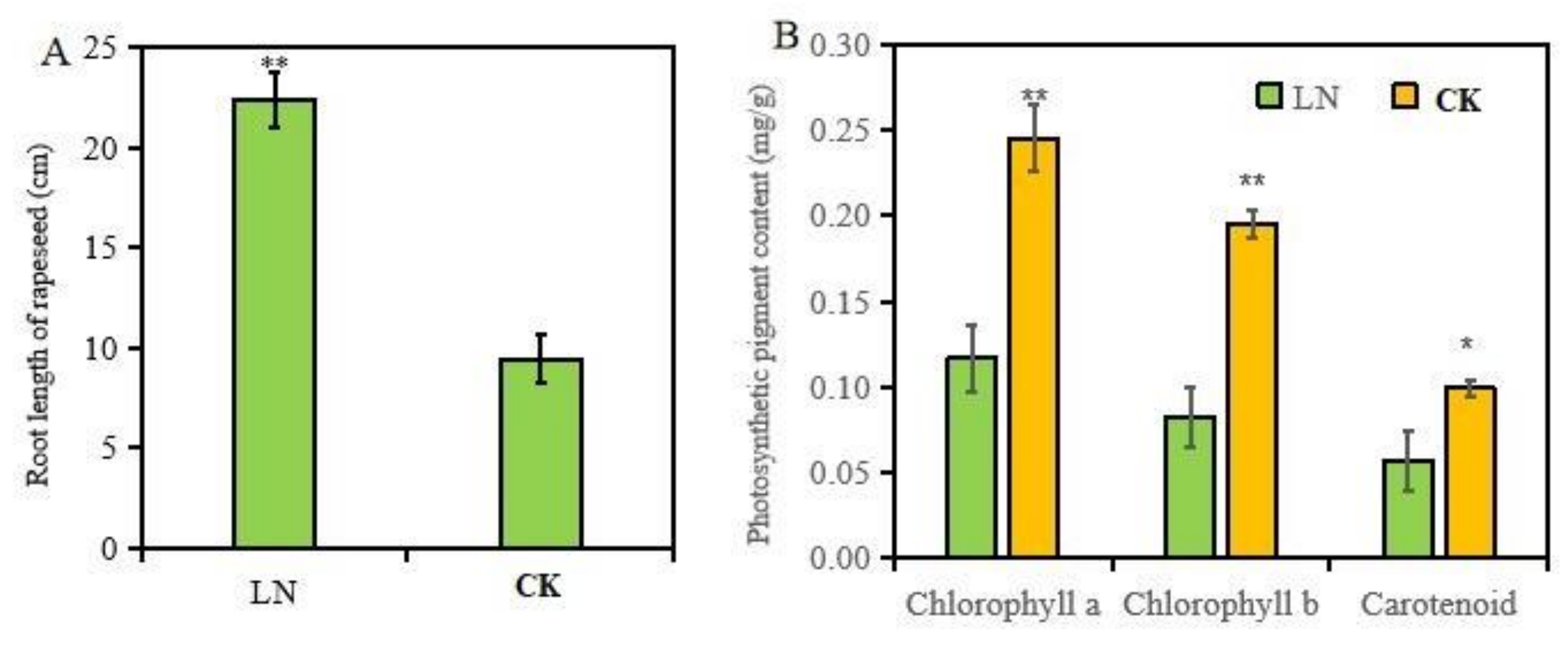
As shown in Figure 1, after 10 days of treatments with low N (0.3 mM NO3−) or normal N (0.6 mM NO3−), the roots and leaves of rapeseed displayed significantly N-deficient symptoms. The leaves of N-starved rapeseed were significantly smaller than those treated with high N, and low N induced large amounts of anthocyanin accumulation in leaves. The results in Figure 2 show that low N significantly decreased the content of photosynthetic pigments (chlorophyll a, b, and carotenoids) in leaves, while the roots showed adaptive elongation in rapeseed under low N treatment.
2.2. Effects of Low N on N Content, Root Shoot/Ratio, and N Physiological Efficiency of Rapeseed
Dry matter accumulation of plants reflects the growth status of plants to a certain extent. Low N stress remarkably reduced the dry matter accumulation of rapeseed leaves, while it significantly promoted the dry matter accumulation of roots (Table 1), which echoed the results of root morphology. The root/shoot ratio can be used to characterize the distribution characteristics of photosynthetic products in plants. Under N starvation conditions, plants distribute more photosynthetic products to roots to promote the preferential growth of roots. As shown in Table 1, the root/shoot ratio of rapeseed treated with low N was significantly higher than that of rapeseed treated with normal N. In the meantime, low N decreased the total N content and N accumulation in rapeseed leaves and roots at a significant level. Nitrogen physiological use efficiency is an index reflecting the N use efficiency of plants, which represents the production of dry matter per unit of N accumulation. Low N treatment of rapeseed has a higher N physiological efficiency, which indicates that N-deficient rapeseed can produce more dry matter with less N accumulation, and this phenomenon is also a low N adaptive response of plants.
2.3. Differences in Cell Wall Components of Rapeseed Roots under Different N Treatments
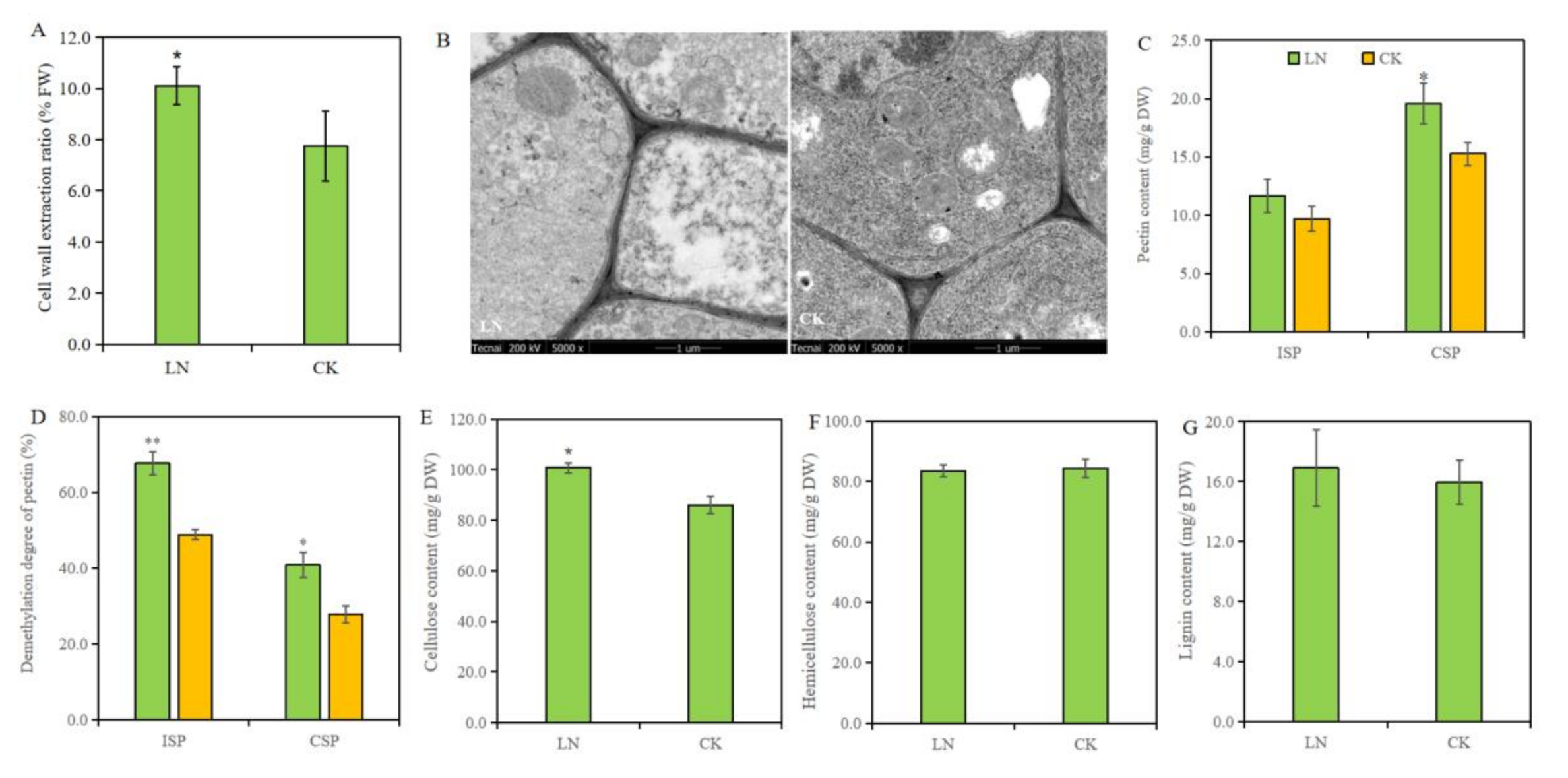
The extraction rate of root cell walls was significantly increased by low N treatment, which was 30.58% higher than that of CK treatment (Figure 3A). TEM images were used to show the difference of cell wall and our result suggested no obvious change in cell wall thickness and morphological structure (Figure 3B).
In order to more accurately study the responses of main chemical components in root cell walls to low N treatments, we quantitatively measured the content of different types of pectin, cellulose, hemicellulose, and lignin in cell walls. Compared to normal N treatment, low N stress significantly increased the content of CSP and cellulose in root cell walls by 17.81% and 17.44%, respectively, while having no obvious effect on ISP, hemicellulose, and lignin content (Figure 3C,E–G). Furthermore, the pectin demethylation was analyzed and the results showed that low N increased the demethylation degree of ISP and CPS (Figure 3D), which was beneficial to cell wall loosing.
2.4. Differences in Degradation of Pectin and Cellulose in Roots under Different N Treatments
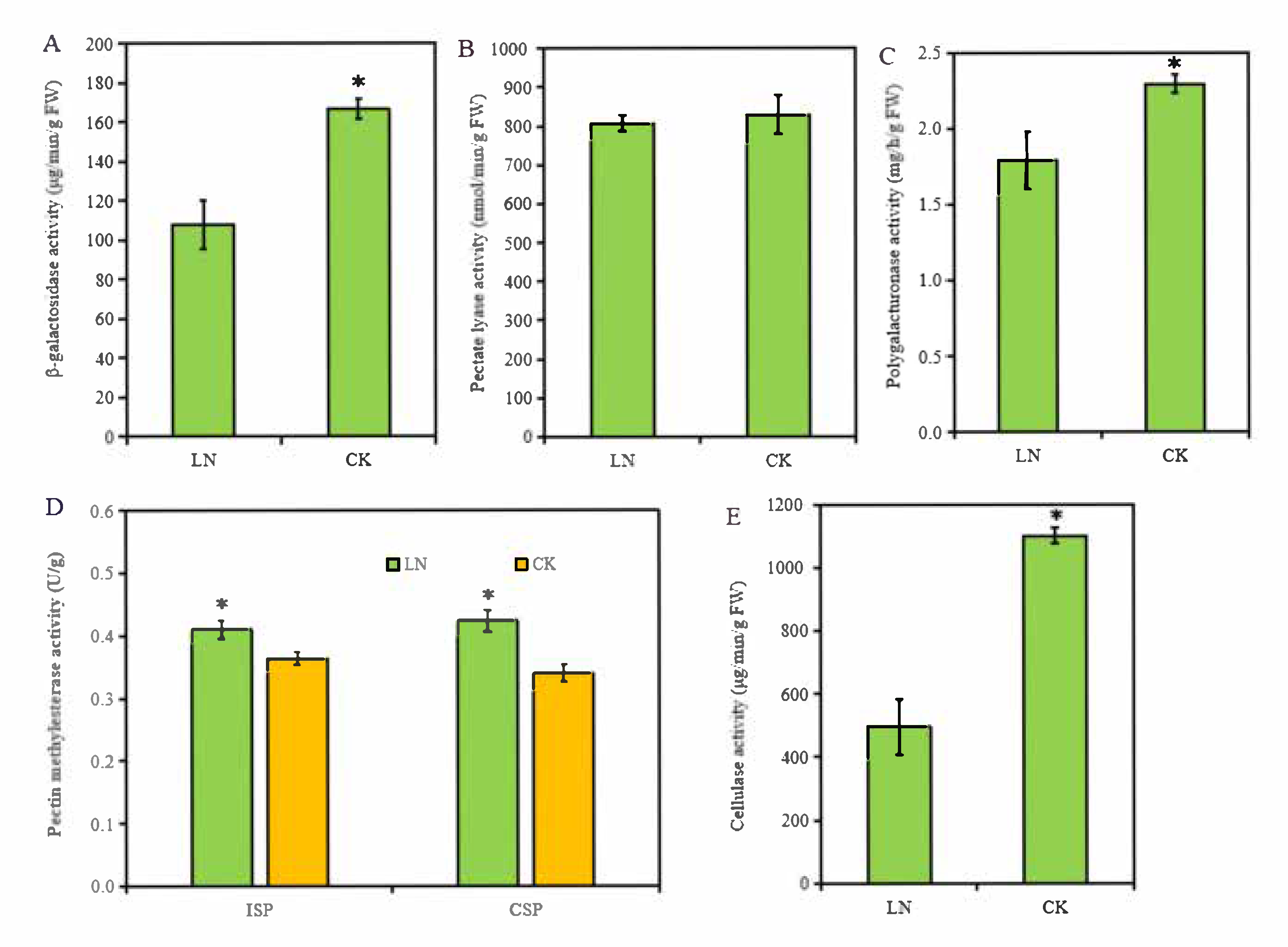
Our analyses of cell wall components in rapeseed roots suggested that low N not only increased CSP and cellules content but also promoted the demethylation of ISP and CSP. To further confirm this, we determined the activity of enzymes involved in the pectin demethylation, and degradation of pectin and cellulose in root cell walls. Using the chemical colorimetry method, we determined the activity of the PME and three pectinases, pectic lyase, polygalacturonase, and β-galactosidase, in the root cell walls. We found that the activity of β-galactosidase and polygalacturonase was remarkably inhibited by N deficiency (Figure 4B,D). Thus, the lower β-galactosidase and polygalacturonase activity may be involved in inhibited pectin degradation in roots under low N condition. Low N increased the PME activity of ISP and CSP from 0.36 to 0.41 U/g and 0.33 to 0.42 U/g, respectively (Figure 4E), which corresponding to the result of pectin demethylation degree (Figure 3D).
2.5. Responses of Genes Involved in Pectin Demethylation and Degradation of Pectin and Cellulose in Roots to Low N Treatment
Our physiological analyses indicated that low N promoted pectin demethylation and increased ISP and cellulose content in root cell walls. The transcriptional profiling of the pectinases and cellulose further verified the above results. The comparison of low N and CK treatments identified five DEGs that were involved in pectate lyase and were both downregulated by N deficiency; two DEGs involved in polygalacturonase were significantly downregulated; 5/7 DEGs involved in β-galactosidase were significantly downregulated; and 7/9 DEGs related to cellulose degradation were significantly downregulated (Table 2). In the meantime, between low N and CK treatment, we identified 15 and 2 DEGs of PME family and PME inhibitor (PMEI) family, respectively; the transcription level of 12 DEGs related to PME was improved, and 2 PMEI genes all were down-expressed by low N.
3. Discussions
Nitrogen starvation is a major limiting factor for rapeseed production. The root system is the most important organ for nutrient acquisition from the soil. Growing evidence has indicated that the availability of N can modulate root growth and architecture in plants. However, most previous studies on the response of rapeseed to N starvation have mainly focused on leaves. Our study suggested that low N significantly decreased the photosynthetic pigments content in leaves (Figure 2B), and thereby inhibited the progress of photosynthesis and the accumulation of dry matter in leaves (Table 1). Under low N stress, plant roots make morphological, structural, and physiological adaptive responses, such as increased root/shoot ratio [9,23], promoted root elongation and root weight, and changes in root architecture [6,24,25]. In a recent study, a 3D in situ quantification analysis system was used to reveal the architectural response of rapeseed roots to N deficiency and found that N deficiency not only stimulated the elongation of the main root and lateral root, but also increased the number of root tips [26]. In our study, the root images obtained from the root scanner and root length showed that low N stress changed the root morphology with promoted root elongation and increased root surface area (Figure 1C and Figure 2A), which is conducive to more effective contact with N in soil and improving absorption efficiency in an N-limited environment. Furthermore, low N treatment significantly increased the dry matter accumulation of rapeseed roots (Table 1), which is closely related to root morphology alteration.
A lot of studied have focused on revealing the modulation mechanism of root architecture alteration, and Giehl et al. noted that the most plausible mechanistic view is that once perceived nutrient signals, a set of genes/proteins in the root developmental program were affected in order to modify specific root developmental processes [13]. Root system architectural changes can be triggered by complex signaling mechanisms such as brassinosteroid signaling, auxin transporter, and auxin signaling that are involved in the regulation of root architecture under low N conditions [19,20,27,28]. Root morphogenesis is regulated by cell division and expansion [21]. Cell wall biosynthesis is involved in the processes of cell division, elongation, and differentiation [14], and affects cell size and shape. Cell wall structure and cell wall metabolism are strongly influenced by various abiotic stresses, such as drought, flooding, extreme temperature, salinization, and heavy metals [29]. A previous study found that the amount of several cell-wall-related proteins in rapeseed roots increased under N deficiency, which participates in the adaptive elongation of roots [26]. Nitrogen also has a significant effect on cell wall composition [30]. It has been reported that ammonium could reduce the content of pectin and hemicellulose in cell walls of rice root tips [31]. Cell elongation also promotes cell wall components (such as cellulose, hemicellulose, pectin, etc.) synthesis to maintain cell wall thickness [22]. Xin et al. identified proteins related to cellulose, hemicellulose, and pectin under different N conditions and proposed that the response of rice root morphology to N availability could be achieved through coordination of relevant proteins involved in cell structure, cell wall modification, and cell division [22]. Our results also showed that low N stress changed the cell wall composition of rapeseed roots, accompanied by a significant increase in two different types of pectin and cellulose content, but had no remarkable effect on hemicellulose and lignin content (Figure 3C,E–G). A study on sorghum seedlings also confirmed that N deficiency affected root cell wall components by upregulated expression of cell-wall-related genes, including cellulose synthase-like proteins and pectate lyase [32]. The analysis of pectinase, cellulase, and pectin monosaccharide group, and the expression of related genes, indicated that the increase of pectin and cellulose was mainly attributed to inhibited degradation of pectin and cellulose in root cell walls induced by N deficiency. Pectin demethylation was also promoted by low N treatment (Figure 3D), and the increased PME activity, upregulated PME genes and downregulated PMEI genes, also proved the results (Figure 4D and Table 2). As the main component of plant cell walls, pectin has many functions in plants, such as promoting cell adhesion, providing structural support, affecting the formation of secondary cell walls, etc. [33,34]. It has been reported that PME activity appeared to create an acidic environment within the cell walls, causing cell wall loosening [35]. Cellulose is part of cell walls, and it performs physiological functions in plants and also plays a skeleton supporting role in plant cell walls and has excellent tensile strength [36]. It is widely known that root morphogenesis is controlled by cell division and expansion, and these are related to the loosening and hardening of cell walls. In this study, low N treatment induced promoting pectin demethylation, more pectin and cellulose in root cell walls, which were conducive to promote cell wall loosening and provide more components for cell wall synthesis, thus promoting root cell division and elongation, ultimately promoting root elongation.
4. Materials and Methods
4.1. Experimental Design
Zhongshuang “11” rapeseed variety was chosen as the study material and cultured in a nutrient solution in an illuminated room. The temperature of the illumination room was set at 22 °C–24 °C, the light cycle was 14 h (light)/10 h (dark), and the light intensity was 300–320 μmol/m2/s. The humidity was maintained at 60–75%. The macro and microelements were provided according to Hoagland and Arnon’s nutrient solution formula. In the experiment, two different N treatments were set: low N (LN: 0.3 mmol/L NO3−) and normal N (CK: 6.0 mmol/L NO3−), and six biological replicates were set for each treatment. The germinated and consistently growing rapeseed plants were chosen and transplanted into the shading culture plates. After being cultured in the normal nutrient solution for 5 days, the seedlings were transferred into the nutrient solution with different N concentrations for 10 days.
4.2. Determination Indicators and Methods
Growth parameter: during the treatment period, the root and leaf phenotypes of rapeseed were observed and recorded, and the root length was measured after being harvested.
Measurement of photosynthetic pigment content: according to the previously described method [37], the photosynthetic pigment of fresh leaves was extracted by ethanol (95%). The absorbance of chlorophyll a, b, and carotenoid was gained by UV–VIS spectrophotometry (TU-1810, PGENERAL, Beijing, China) at wavelengths of 470, 649, and 665 nm, respectively. The chlorophyll a, chlorophyll b, and carotenoid contents were calculated according to their corresponding formulae: chlorophyll a (mg/L) = 13.95A665−6.88A649, chlorophyll b (mg/L) = 24.96A649−7.32A665, carotenoid (mg/L) = .
Transmission electron microscope (TEM) imaging: the TEM slices of roots were prepared using the method of Kong et al. [38]. Ultrathin sections were examined with a TEM (Hitachi 500 electron microscope) at an acceleration voltage of 60 kV, and representative plant images were chosen for each N treatment.
Dry matter accumulation: after harvesting, the roots and leaves of rapeseed seedlings were sampled and then the enzymes in plants were first inactivated at 105 °C for 30 min, and later dried at 75 °C for 3 days, and finally weighed to measure the dry weight.
Determination of total N content: about 0.1 g of the sample was weighed after being crushed and evenly mixed, and then placed into a triangular flask, and 5 mL of concentrated sulfuric acid was added for digestion on an electric heating plate. When the samples were clear, the hydrogen peroxide was added and then transferred to a volumetric flask. After dry filtration and dilution, the AA3 flow analyzer (autoanalyzer 3, seal, Germany) was used to determine the total N content.
Extraction of root cell walls was conducted based on the methods of Weigel and Jager [39] and Rathore [40]: after the fresh root was finely ground in a precooled quartz mortar with a homogenate solution (pH = 7.2) of sucrose, Tris-HCl, and dithiothreitol, the mixed solution was centrifuged at 2000× g at 4 °C for 10 min, and the precipitate was considered as cell wall material.
Separation of root cell wall components: the cell wall material was further separated into ion-bound pectin (ISP), covalently bound pectin (CSP), cellulose, and hemicellulose, as previously described by Redgwell and Selvendran [41] with slight modification. Firstly, the ground cell wall powder was suspended in 0.05 M sodium acetate buffer (pH 6.5) containing 0.05 M CDTA and stirred for 12 h at 24 °C in a horizontal shaker, and the supernatant was defined as CDTA-soluble pectin (CSP). Then, the CDTA-insoluble pellet was resuspended in 0.05 M Na2CO3 and incubated for 12 h at 24 °C. After centrifugation at 5000× g for 20 min, the supernatant was designated as ion-soluble pectin (ISP). Finally, the remaining residue was added to 20 mL of 4 M KOH solution, extracted by shaking at 24 °C for 3 h, and centrifuged. The supernatant was KOH-soluble component, which was mainly hemicellulose, and the last residue was mainly cellulose.
Determination of cell wall components: the contents of ISP, CSP, cellulose, and hemicellulose were respectively determined using corresponding kits (Keming Biotechnology Co., Ltd., Suzhou, China).
Determination of cell wall lignin content: lignin content in root samples was determined using the acetyl bromide method [42,43]. Briefly, 10 mg of dry root samples were incubated in 0.6 mL of a 25% acetyl bromide solution (w/w, AcBr in glacial acetic acid) in 2 mL Eppendorf tubes at 80 °C for 3 h. After cooling to 25 °C and centrifuging at 12,000× g for 5 min, 0.1 mL of the above mixture was transferred into a new 2 mL Eppendorf tube. Then, 0.4 mL of 2 M NaOH and 0.07 mL of 0.5 M hydroxylamine hydrochloride were added and the mixture was thoroughly vortexed. Subsequently, 100 μL of the mixture was transferred to a 96-well microplate followed by 200 μL of glacial acetic acid. Finally, the percentage of acetyl bromide soluble lignin was measured by UV–Vis spectrophotometry (TU-1810, PGENERAL, Beijing, China) at a wavelength of 280 nm. Lignin concentration was expressed as A280 g−1 DW.
Determinations of pectin methylation degree: by the method of Anthon and Barrett [44], with some modifications, the methylation degrees of ISP and CSP were measured. After the pectin extraction with NaOH (1.5 M) was treated at 25 °C for 30 min, the mixed solution of H2SO4 (0.75 M), Tris-HCl (0.2 M, pH 7.5), MBTH (3 mg/mL), and alcohol oxidase (0.01 units/mL) was added. Then, the reaction was terminated by a solution containing ammonium ferric sulfate (5 mg/mL) and sulfamic acid (5 mg/mL). Finally, after incubation at 25 °C for 20 min, the absorbance was measured at 620 nm using a spectrophotometer (TU-1810, PGENERAL, Beijing, China). Pectin demethylation degree = 100 – pectin methylation degree.
The activity of pectinase and cellulose analysis: the activities of the PME, pectate lyase, polygalacturonase, β-galactosidase, and cellulase were measured with the available commercial PME-2-G, PL-1-G, PG-1-G, β-GALB-1-Y, and CL-2-Y kits, respectively, following the manufacturer’s guidelines (Suzhou Comin Biotechnology Co., Ltd. Suzhou, China).
High-throughput sequencing data analysis of the transcriptome: the mapped reads of each sample were assembled using StringTie (http://ccb.Jhu.edu/software/stringtie/, accessed on 1 February 2022). Then, the transcriptomes from the samples were merged to reconstruct a comprehensive transcriptome using perl scripts. After the final transcriptome was generated, StringTie and edgeR (http://bioconductor.org/packages/release/bioc/html/edgeR.html, accessed on 1 February 2022) were used to estimate the expression levels of all the transcripts. StringTie was used to examine the expression levels of the mRNAs by calculating the FPKM (fragments per kilobase of transcript sequence per million base pairs sequenced). The differentially expressed genes (DEGs) related to cell wall remodeling were defined as those with p-value < 0.05.
4.3. Statistical Analysis
The data were statistically analyzed by SAS 9.1.3 (SAS Institute, Cary, NC, USA). ANOVA analysis based on the Student’s t-test method was used to determine the significance of differences between LN and HN treatments. The * and ** represent the statistical significances at the levels of p < 0.05 and p < 0.01, respectively.
5. Conclusions
Compared with normal N treatment, low N treatment significantly promoted rapeseed root elongation and improved root dry mass accumulation, root/shoot ratio, and N physiological efficiency, while reducing photosynthetic pigment content and dry matter accumulation in leaves. Analysis of the main chemical components of root cell walls showed that low N increased the demethylation degree of ISP and CSP, the content of CSP, and cellulose in root cell walls, which were closely related to promoted PME activity, and inhibited pectin degradation enzymes, cellulase. The low-N-increased demethylation degree of pectin and content of pectin and cellulose in cell walls was conducive to cell wall loosening and cell wall synthesis during cell division and elongation, ultimately promoting root adaptive elongation.
Author Contributions
H.T. and X.W. designed the experiments; H.T. performed the experiments and conducted the determination and analysis of laboratory indexes; X.W. and H.S. performed critical data interpretation; X.W. wrote the first draft of the manuscript; Z.Z. reviewed and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was partially supported by the Natural Science Foundation of Hunan Provincial (2020JJ5242; 2021JJ0004), the National Natural Science Foundation of China (32072664), and the National Oilseed Rape Production Technology System of China.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Coruzzi, G.; Bush, D.R. Nitrogen and Carbon Nutrient and Metabolite Signaling in Plants. Plant Physiol. 2001, 125, 61–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malagoli, P.; Laine, P.; Rossato, L.; Ourry, A. Dynamics of nitrogen uptake and mobilization in field-grown winter oilseed rape (Brassica napus) from stem extension to harvest. ii. An 15N-labelling-based simulation model of n partitioning between vegetative and reproductive tissues. Ann. Bot. 2005, 7, 1187–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roger, S.B.; Kindred, D.R. Analysing nitrogen responses of cereals to prioritize routes to the improvement of nitrogen use efficiency. J. Exp. Bot. 2009, 7, 1939–1951. [Google Scholar]
- Gao, K.; Chen, F.J.; Yuan, L.X.; Zhang, F.S.; Mi, G.H. A comprehensive analysis of root morphological changes and nitrogen allocation in maize in response to low nitrogen stress. Plant Cell Environ. 2014, 38, 740–750. [Google Scholar] [CrossRef]
- Li, X.X.; Zeng, R.S.; Liao, H. Improving crop nutrient efficiency through root architecture modifications. J. Integr. Plant Biol. 2015, 58, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Chun, L.; Mi, G.H.; Li, J.S.; Chen, F.J.; Zhang, F.S. Genetic Analysis of Maize Root Characteristics in Response to Low Nitrogen Stress. Plant Soil 2005, 276, 369–382. [Google Scholar] [CrossRef]
- Eghball, B.; Maranville, J.W. Root Development and Nitrogen Influx of Corn Genotypes Grown under Combined Drought and Nitrogen Stresses. Agron. J. 1993, 85, 147–152. [Google Scholar] [CrossRef]
- Feil, B.; Thiraporn, R.; Geisler, G.; Stamp, P. Root traits of maize seedlings-indicators of nitrogen efficiency. Plant Soil 1990, 123, 155–159. [Google Scholar] [CrossRef]
- Vamerali, T.; Saccomani, M.; Bona, S.; Mosca, G.; Guarise, M.; Ganis, A. A comparison of root characteristics in relation to nutrient and water stress in two maize hybrids. Plant Soil 2003, 255, 157–167. [Google Scholar] [CrossRef]
- Mi, G.H.; Chen, F.J.; Wu, Q.P.; Lai, N.W.; Yuan, L.X.; Zhang, F.S. Ideotype root architecture for efficient nitrogen acquisition by maize in intensive cropping systems. Sci. China Life Sci. 2010, 53, 1369–1373. [Google Scholar] [CrossRef]
- Ju, C.; Buresh, R.J.; Wang, Z.; Zhang, H.; Liu, L.; Yang, J.; Zhang, J. Root and shoot traits for rice varieties with higher grain yield and higher nitrogen use efficiency at lower nitrogen rates application. Field Crops Res. 2015, 175, 47–55. [Google Scholar] [CrossRef]
- Scheres, B.; Benfey, P.; Dolan, L. Root development. In The Arabidopsis Book; Somerville, C.R., Meyerowitz, E.M., Eds.; America Society of Plant Biologists: Rockville, MD, USA, 2002. [Google Scholar]
- Giehl, R.F.H.; Gruber, B.D.; Von Wirén, N. It’s time to make changes: Modulation of root system architecture by nutrient signals. J. Exp. Bot. 2014, 65, 769–778. [Google Scholar] [CrossRef]
- Xia, M.; Chen, A.L.; Yu, F. The effect of the node on the development of ground tissue cell walls during the rapid elongation growth of phyllostachys edulis ‘Pachyloen’ culm. Acta Agri. Univ. Jiangxi 2018, 40, 68–74. [Google Scholar]
- Tenhaken, R. Cell wall remodeling under abiotic stress. Front. Plant Sci. 2015, 5, 771. [Google Scholar] [CrossRef] [Green Version]
- Franciosini, A.; Rymen, B.; Shibata, M.; Favero, D.S.; Sugimoto, K. Molecular networks orchestrating plant cell growth. Curr. Opin. Plant Biol. 2017, 35, 98–104. [Google Scholar] [CrossRef]
- Simone, L.; Sergio, E. Nitrate uptake affects cell wall synthesis and modeling. Front. Plant Sci. 2017, 8, 1376. [Google Scholar]
- Fukuda, H. Plant Cell Wall Patterning and Cell Shape; Wiley: Hoboken, NJ, USA, 2014. [Google Scholar]
- Mounier, E.; Pervent, M.; Ljung, K.; Gojon, A.; Nacry, P. Auxin-mediated nitrate signaling by NRT1.1 participates in the adaptive response of Arabidopsis root architecture to the spatial heterogeneity of nitrate availability. Plant Cell. Environ. 2015, 37, 162–174. [Google Scholar] [CrossRef]
- Vidal, E.A.; Viviana, A.; Cheng, L.; Geraint, P.; Green, P.J.; Coruzzi, G.M.; Gutiérrez, R.A. Nitrate-responsive miR393/AFB3 regulatory module controls root system architecture in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2010, 107, 4477–4482. [Google Scholar] [CrossRef] [Green Version]
- Strader, L.C.; Chen, G.L.; Bartel, B. Ethylene directs auxin to control root cell expansion. Plant J. 2010, 64, 874–884. [Google Scholar] [CrossRef] [Green Version]
- Xin, W.; Zhang, L.N.; Gao, J.P.; Zhang, W.Z.; Yi, J.; Zhen, X.X.; Bi, C.Y.; He, D.W.; Liu, S.M.; Zhao, X.Y. Adaptation Mechanism of Roots to Low and High Nitrogen Revealed by Proteomic Analysis. Rice 2021, 14, 5. [Google Scholar] [CrossRef]
- Liu, Q.X.; Ren, T.; Liao, S.P. Effect of different nitrogen application on seedling growth and allocation of carbon and nitrogen in oilseed rape (Brassica napus L.). Chin. J. Oil Crop Sci. 2019, 41, 92–100. [Google Scholar]
- Wang, Y.; Mi, G.H.; Chen, F.J. Response of root morphology to nitrate supply and its contribution to nitrogen accumulation in maize. J. Plant Nutr. 2004, 27, 2189–2202. [Google Scholar] [CrossRef]
- Rasmussen, I.S.; Dresbll, D.B.; Thorup-Kristensen, K. Winter wheat cultivars and nitrogen (N) fertilization-effects on root growth, n uptake efficiency and n use efficiency. Eur. J. Agron. 2015, 68, 38–49. [Google Scholar] [CrossRef]
- Qin, L.; Walk, T.C.; Han, P.; Chen, L.; Zhang, S.; Li, Y.; Hu, X.; Xie, L.; Yang, Y.; Liu, J.; et al. Adaption of Roots to Nitrogen Deficiency Revealed by 3D Quantification and Proteomic Analysis. Plant Physiol. 2019, 179, 329–347. [Google Scholar] [CrossRef] [Green Version]
- Jia, Z.T.; Giehl, R.F.H.; von Wirén, N. The Root Foraging Response under Low Nitrogen Depends on DWARF1-Mediated Brassinosteroid Biosynthesis. Plant Physiol. 2020, 183, 998–1010. [Google Scholar] [CrossRef]
- Ötvös, K.; Marconi, M.; Vega, A.; O’Brien, J.; Johnson, A.; Abualia, R.; Antonielli, L.; Montesinos, J.C.; Zhang, Y.; Tan, S.; et al. Modulation of plant root growth by nitrogen source-defined regulation of polar auxin transport. EMBO J. 2021, 40, e106862. [Google Scholar] [CrossRef]
- Le Gall, H.; Philippe, F.; Domon, J.-M.; Gillet, F.; Pelloux, J.; Rayon, C. Cell Wall Metabolism in Response to Abiotic Stress. Plants 2015, 4, 0112. [Google Scholar] [CrossRef]
- Allison, G.G.; Morris, C.; Lister, S.J.; Barraclough, T.; Yates, N.; Shield, I.; Donnison, I.S. Effect of nitrogen fertiliser application on cell wall composition in switchgrass and reed canary grass. Biomass Bioenergy 2012, 40, 19–26. [Google Scholar] [CrossRef]
- Wang, W. Study on the mechanism of nitrogen forms affecting aluminum toxicity and aluminum affecting manganese toxicity in rice [D]. Univ. Chin. Acad. Sci. 2014. [Google Scholar]
- Rivai, R.R.; Miyamoto, T.; Awano, T.; Takada, R.; Tobimatsu, Y.; Umezawa, T.; Kobayashi, M. Nitrogen deficiency results in changes to cell wall composition of sorghum seedlings. Sci. Rep. 2021, 11, 23309. [Google Scholar] [CrossRef] [PubMed]
- Atmodjo, M.A.; Hao, Z.; Mohnen, D. Evolving Views of Pectin Biosynthesis. Annu. Rev. Plant Biol. 2013, 64, 747–779. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.J.; Shen, X.P.; Cao, J.S. The metabolism and function of homogalacturonan in plant cell wall. Chin. J. Cell Biol. 2014, 36, 93–98. [Google Scholar]
- Kumar, N.; Srivastava, G.; Chand, D. Cell wall hydrolases: Expansion or senescence in roses (Rosa hybrida L.). Hort. Environ. Biotechnol. 2008, 49, 376–380. [Google Scholar]
- Jin, K.X.; Jiang, Z.H.; Liu, X.G.; Yang, S.M.; Tian, G.L.; Ma, J.F. Research advance in cellulose fibril aggregates structure of plant cell wall. Mater. Rev. 2019, 33, 2997–3002. [Google Scholar]
- Morales, F.; Abadía, A.; Belkhodja, R.; Abadia, J. Iron deficiency-induced changes in the photosynthetic pigment composition of field-grown pear (Pyrus communis L.) leaves. Plant Cell Environ. 1994, 17, 1153–1160. [Google Scholar] [CrossRef]
- Kong, Y.; Xu, X.Y.; Zhu, L. Cyanbactericidal effect of Streptomyces sp. HJC-D1 on microcystis auruginosa. PLoS ONE 2013, 8, e57654. [Google Scholar]
- Weigel, J.J.; Jager, H.J. Subcellular distribution and chemical form of cadmium in bean plant. Plant Physiol. 1980, 65, 480–482. [Google Scholar] [CrossRef] [Green Version]
- Rathore, V.S.; Bajaj, Y.P.S.; Wittwer, S.H. Subcellular Localization of Zinc and Calcium in Bean (Phaseolus vulgaris L.) Tissues. Plant Physiol. 1972, 49, 207–211. [Google Scholar] [CrossRef] [Green Version]
- Redgwell, R.J.; Selvendran, R.R. Structural features of cell-wall polysaccharides of onion Allium cepa. Carbohydr. Res. 1986, 157, 183–199. [Google Scholar] [CrossRef]
- Papa, G.; Varanasi, P.; Sun, L.; Cheng, G.; Stavila, V.; Holmes, B.; Simmons, B.A.; Adani, F.; Singh, S. Exploring the effect of different plant lignin content and composition on ionic liquid pretreatment efficiency and enzymatic saccharification of Eucalyptus globulus L. mutants. Bioresour. Technol. 2012, 117, 352–359. [Google Scholar] [CrossRef]
- Pauly, M.; Keegstra, K. Cell-wall carbohydrates and their modification as a resource for biofuels. Plant J. 2008, 54, 559–568. [Google Scholar] [CrossRef]
- Anthon, G.E.; Barrett, D.M. Comparison of Three Colorimetric Reagents in the Determination of Methanol with Alcohol Oxidase. Application to the Assay of Pectin Methylesterase. J. Agric. Food Chem. 2004, 52, 3749–3753. [Google Scholar] [CrossRef]
Figure 1.
The growth of rapeseed under different N treatment. (A) The growth of the whole seedlings; (B) leaf size and anthocyanin accumulation in leaves; (C) the root morphology reflected by root scanner.
Figure 1.
The growth of rapeseed under different N treatment. (A) The growth of the whole seedlings; (B) leaf size and anthocyanin accumulation in leaves; (C) the root morphology reflected by root scanner.

Figure 2.
The root length (A) and photosynthetic pigment content (B) in rapeseed under different N treatment. The * and ** represent the statistical significances at the levels of p < 0.05 and p < 0.01, respectively.
Figure 2.
The root length (A) and photosynthetic pigment content (B) in rapeseed under different N treatment. The * and ** represent the statistical significances at the levels of p < 0.05 and p < 0.01, respectively.

Figure 3.
The responses of cell wall structure and compositions to different N treatment. (A) Cell wall extraction ratio; (B) TEM images of roots; (C,D) content and demethylation degree of ionic soluble pectin (ISP) and chelated soluble pectin (CSP); (E–G) concentrations of cellulose, hemicellulose, and lignin in root cell walls. The * and ** represent the statistical significances at the levels of p < 0.05 and p < 0.01, respectively.
Figure 3.
The responses of cell wall structure and compositions to different N treatment. (A) Cell wall extraction ratio; (B) TEM images of roots; (C,D) content and demethylation degree of ionic soluble pectin (ISP) and chelated soluble pectin (CSP); (E–G) concentrations of cellulose, hemicellulose, and lignin in root cell walls. The * and ** represent the statistical significances at the levels of p < 0.05 and p < 0.01, respectively.

Figure 4.
The enzyme activity related to pectin and cellulose in rapeseed roots under different N treatment. (A–E) Activities of β-galactosidase, pectate lyase, polygalacturonase, pectin methylesterase, and cellulase in rapeseed roots. The * represents the statistical significance at the levels of p < 0.05.
Figure 4.
The enzyme activity related to pectin and cellulose in rapeseed roots under different N treatment. (A–E) Activities of β-galactosidase, pectate lyase, polygalacturonase, pectin methylesterase, and cellulase in rapeseed roots. The * represents the statistical significance at the levels of p < 0.05.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The difference of dry mass, N content, root/shoot ratio, and N physiological efficiency between rapeseed in low and normal N levels.
Table 1.
The difference of dry mass, N content, root/shoot ratio, and N physiological efficiency between rapeseed in low and normal N levels.
| Treatment | Organ | Dry Mass (g) | Total N Content (mg/g) | N Accumulation (mg/Plant) | Root/Shoot Ratio (%) | N Physiological Efficiency (%) |
|---|---|---|---|---|---|---|
| LN | Leaf | 0.200 ± 0.02 | 22.19 ± 0.73 | 4.19 ± 0.28 | 19.66 * ± 0.92 | 47.83 ** ± 2.49 |
| Root | 0.044 * ± 0.01 | 15.62 ± 0.14 | 0.59 ± 0.02 | |||
| CK | Leaf | 0.295 * ± 0.01 | 42.18 ** ± 1.30 | 10.25 ** ± 0.73 | 13.43 ± 0.85 | 24.36 ± 1.09 |
| Root | 0.037 ± 0.01 | 39.35 * ± 1.08 | 1.05 * ± 0.09 |
Note: The * and ** represent the statistical significances at the levels of p < 0.05 and p < 0.01, respectively.
Table 2.
The transcription level of genes related to pectinase and cellulose in root of rapeseed under different N treatment.
Table 2.
The transcription level of genes related to pectinase and cellulose in root of rapeseed under different N treatment.
| Gene ID | Transcription Level | Fold Change | log2(Fold Change) | Regulation | Significant | Description | |
|---|---|---|---|---|---|---|---|
| LN | CK | ||||||
| BnaAnng22760D | 20.49 | 44.66 | 0.46 | −1.12 | down | yes | Pectate lyase |
| BnaA02g13340D | 5.09 | 17.88 | 0.28 | −1.81 | down | yes | Pectate lyase |
| BnaA07g16550D | 3.50 | 5.30 | 0.66 | −0.60 | down | no | Pectate lyase |
| BnaA09g55050D | 5.71 | 6.13 | 0.93 | −0.10 | down | no | Pectate lyase |
| BnaC08g25770D | 3.55 | 4.66 | 0.76 | −0.39 | down | no | Pectate lyase |
| BnaCnng18640D | 0.13 | 0.87 | 0.15 | −2.76 | down | yes | polygalacturonase |
| BnaA01g10390D | 0.27 | 1.54 | 0.18 | −2.49 | down | yes | polygalacturonase |
| BnaC01g18170D | 13.52 | 6.933 | 1.95 | 0.96 | up | no | β-galactosidase 12 |
| BnaA01g15330D | 12.76 | 7.23 | 1.77 | 0.82 | up | no | β-galactosidase 12 |
| BnaC02g12030D | 3.14 | 24.36 | 0.13 | −2.96 | down | yes | β-galactosidase 12 |
| BnaA03g10550D | 6.25 | 24.71 | 0.25 | −1.98 | down | yes | β-galactosidase 12 |
| BnaAnng30540D | 4.34 | 26.93 | 0.16 | −2.63 | down | yes | β-galactosidase 12 |
| BnaC03g13250D | 7.16 | 19.73 | 0.36 | −1.46 | down | yes | β-galactosidase 12 |
| BnaA02g08480D | 8.66 | 31.33 | 0.28 | −1.86 | down | yes | β-galactosidase 12 |
| BnaA06g00690D | 46.57 | 10.92 | 4.27 | 2.09 | up | yes | PME1 |
| BnaC06g41950D | 15.24 | 4.66 | 3.27 | 1.71 | up | yes | PME1 |
| BnaA05g14120D | 10.78 | 10.97 | 0.98 | −0.02 | down | no | PME1 |
| BnaC06g41940D | 9.21 | 9.70 | 0.95 | −0.07 | down | no | PME2 |
| BnaC01g37010D | 74.38 | 20.67 | 3.60 | 1.85 | up | yes | PME3 |
| BnaA01g29470D | 39.47 | 12.30 | 3.21 | 1.68 | up | yes | PME3 |
| BnaC05g39340D | 79.58 | 64.15 | 1.24 | 0.31 | up | no | PME3 |
| BnaA05g25150D | 96.59 | 90.11 | 1.07 | 0.10 | up | no | PME3 |
| BnaA01g37100D | 0.26 | 0.23 | 1.12 | 0.16 | up | no | PME3 |
| BnaA06g31060D | 8.48 | 6.88 | 1.23 | 0.30 | up | no | PME31 |
| BnaC07g25560D | 4.54 | 4.06 | 1.12 | 0.16 | up | no | PME31 |
| BnaA08g11860D | 1.82 | 1.74 | 1.05 | 0.06 | up | no | PME44 |
| BnaC03g66720D | 4.63 | 4.54 | 1.02 | 0.03 | up | no | PME44 |
| BnaC04g23290D | 1.26 | 1.58 | 0.79 | −0.33 | down | no | PME61 |
| BnaA09g55410D | 0.55 | 0.34 | 1.60 | 0.68 | up | no | PME61 |
| BnaA02g21340D | 6.40 | 26.41 | 0.24 | −2.05 | down | yes | PMEI1 |
| BnaC02g46740D | 23.70 | 57.45 | 0.41 | −1.28 | down | yes | PMEI1 |
| BnaA09g51160D | 0.04 | 0.11 | 0.33 | −1.60 | down | yes | cellulase 2 |
| BnaC06g43770D | 6.55 | 23.25 | 0.28 | −1.83 | down | yes | cellulase 3 |
| BnaA07g23740D | 1.16 | 4.24 | 0.27 | −1.88 | down | yes | cellulase |
| BnaC05g09040D | 0.17 | 0.36 | 0.48 | −1.07 | down | yes | cellulase |
| BnaC07g23300D | 5.17 | 16.97 | 0.30 | −1.72 | down | yes | cellulase |
| BnaA07g28820D | 15.26 | 10.54 | 1.45 | 0.53 | up | no | cellulase |
| BnaC06g24550D | 5.79 | 5.97 | 0.97 | −0.04 | down | no | cellulase |
| BnaA10g05530D | 50.15 | 51.63 | 0.97 | −0.04 | down | no | cellulase |
| BnaC09g26420D | 64.25 | 60.88 | 1.06 | 0.08 | up | no | cellulase |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tian, H.; Song, H.; Wu, X.; Zhang, Z. Responses of Cell Wall Components to Low Nitrogen in Rapeseed Roots. Agronomy 2022, 12, 1044. https://doi.org/10.3390/agronomy12051044
AMA Style
Tian H, Song H, Wu X, Zhang Z. Responses of Cell Wall Components to Low Nitrogen in Rapeseed Roots. Agronomy. 2022; 12(5):1044. https://doi.org/10.3390/agronomy12051044
Chicago/Turabian StyleTian, Hui, Haixing Song, Xiuwen Wu, and Zhenhua Zhang. 2022. "Responses of Cell Wall Components to Low Nitrogen in Rapeseed Roots" Agronomy 12, no. 5: 1044. https://doi.org/10.3390/agronomy12051044
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.