
Profiling of Water-Use Efficiency in Switchgrass (Panicum virgatum L.) and the Relationship with Cadmium Accumulation
by
 ,
,
Zhaoyang Hu
1,2 ,
,
Zhigang Fang
1,3,
Bingyu Hu
1,
Xuehui Wen
1,
Laiqing Lou
1 and
Qingsheng Cai
1,* 1
College of Life Sciences, Nanjing Agricultural University, Nanjing 210095, China
2
College of Bioscience and Bioengineering, Jiangxi Agricultural University, Nanchang 330045, China
3
College of Life and Geographic Sciences, Kashi University, Kashi 844006, China
*
Author to whom correspondence should be addressed.
Agronomy 2022, 12(2), 507; https://doi.org/10.3390/agronomy12020507
Submission received: 6 January 2022
/
Revised: 10 February 2022
/
Accepted: 16 February 2022
/
Published: 17 February 2022
(This article belongs to the Special Issue Grass Improvement through Integrated Omics, Physiology and Breeding Approaches)
Abstract
:Planting bioenergy crops with high water-use efficiency (WUE) on heavy metal-polluted land is a good practice for biomass production and phytoremediation. Switchgrass (Panicum virgatum L.), a C4 perennial bioenergy grass, is native to the United States. The relationship between the WUE and Cd accumulation of switchgrass has seldom been studied. Here, the WUE and Cd accumulation characteristics of 14 high-biomass switchgrass cultivars were investigated under Cd stress by hydroponic culture. The main results showed that Cd inhibited the instantaneous WUE in switchgrass seedlings and that the inhibition rate was more significant in the upland types than in the lowland types of switchgrass. A positive correlation was found between relative WUE and Cd accumulation in roots and shoots. The relative expression level of stomatal control-related genes (ERECTA and EPF1) in lowland cultivars with high WUE was higher than in upland cultivars with low WUE, both in control and Cd treatment conditions. The results suggest that it would be possible to further select and cultivate switchgrass with high WUE and a high capacity for Cd accumulation for phytoremediation in Cd-contaminated land.
1. Introduction
Switchgrass (Panicum virgatum L.) is a high-biomass biofuel crop native to the United States [1]. Its agricultural value lies in its rapid growth with low inputs, tolerance of various adverse environmental conditions, and wide geographic range [2]. Field studies have demonstrated that switchgrass can produce 540% more renewable energy as a biomass crop than the energy it consumes and that it comes with environmental benefits [3,4]. Additionally, switchgrass has been used for phytoremediation in marginal land contaminated with heavy metals, such as cadmium (Cd) and lead (Pb) [5,6].
Cd is one of the most toxic heavy metals for all organisms [7]. Cd in soil is easily absorbed by roots and accumulated in the edible portions of plants, resulting in a severe threat to human health through the food chain [8,9]. It has been estimated that 7% of the land in China is contaminated with Cd according to a survey by China’s ministry of environmental protection [10]. Cd may also cause plant damage even at very low concentrations and its toxic effects may alter plant water physiology [11,12]. For instance, P. euphratica cells treated with Cd exhibited a clear contraction of the cytoplasm, indicating interference with the water balance [13]. Plasma membrane permeability has been shown to be affected by Cd, leading to a decrease in the water content of plants [14]. Additionally, Cd toxicity has been shown to decrease plants’ water-use efficiency (WUE). Previous reports have demonstrated that Cd strongly inhibited the synthesis of chlorophylls and their stable binding to proteins, thereby damaging the photosynthetic apparatus and, in particular, decreasing the concentration of light-harvesting complex II and photosystems I (PSI) and II (PSII) [15,16]. Water is one of the crucial factors limiting crop production in numerous regions worldwide [17,18]. Other studies have indicated that increasing WUE can reduce water stress and improve productivity under stress [19,20]. Thus, research on WUE aims to discern the most effective use of water in order to achieve the highest possible agricultural yield or income with the lowest possible water consumption.
Recent studies have suggested that utilizing Cd-contaminated marginal lands to develop switchgrass is an option for harvesting biomass [21,22]. This could not only avoid competing with food crops for arable fields but also remove Cd from the soil for phytoremediation. Wullschleger et al. [23] studied 25 upland and 14 lowland types of switchgrass and found that different types of switchgrass had different WUEs. What is the relationship between the Cd accumulation and WUE of switchgrass? Here, we hypothesized that the WUE of different switchgrass cultivars would be inhibited under Cd stress and that this was related to stomatal closure, stomatal formation, and water uptake. In this study, we aimed to assay the WUEs of 14 switchgrass cultivars in response to Cd stress. Two cultivars were then selected in order to explore the mechanism behind differences in WUE.
2. Materials and Methods
2.1. Materials and Plant Cultivation
In this study, 14 cultivars of switchgrass (listed in Table 1) were used. Seedlings were cultured as described by Liu et al. [21]. Briefly, the seeds were shocked with 50% sulfuric acid (analytical reagent) for 30 min at 180 rpm and then washed with distilled water five times. The seeds were then sterilized with 10% NaClO for 15 min and rinsed thoroughly with distilled water. After that, the seeds were preliminarily germinated in a culture dish with filter paper and after a week the seedlings were transferred to a plastic container with clean quartz sand for further germination. Three weeks later, uniform seedlings were transferred to black beakers (1 L volume) filled with a modified ¼ Hoagland solution for a week and subsequently ½ Hoagland solution. The seedlings were treated with Cd when they grew five fully expanded leaves.
2.2. Cadmium Treatments
The seedlings were treated with 10 μmol L−1 Cd (supplied as CdCl2·2H2O), based on a previous study [21]. Seedlings not treated with Cd served as the control. Each treatment was replicated three times. After two weeks, the seedlings were harvested. The trials were conducted in a growth chamber with a 12 h light/12 h dark photoperiod (light intensity 150 μmol m−2s−1) with light/dark temperatures of 30/25 °C, respectively, and 70% relative humidity. The nutrient solution (pH 5.8) was renewed every 3 days. The positions of the beakers were randomly changed once a day to minimize position effects.
2.3. Estimation of Leaf Gas-Exchange Parameters and Instantaneous WUE
The leaf gas-exchange parameters of 14 switchgrass cultivars, including net photosynthetic rate (Pn), transpiration rate (E), intercellular CO2 concentration (Ci), and stomatal conductance (Gs), were determined with a LiCor-6400 portable photosynthesis system (LiCor Inc., Lincoln, CA, USA) equipped with an LED light source [24]. The light intensity of the leaf chamber, CO2 concentration, and leaf temperature were maintained at 1200 μmol m−2s−1. The instantaneous water-use efficiency (iWUE) and relative water use efficiency (RWUE) were calculated as follows [25]:
WUECd and WUEControl represent the WUE under Cd and control conditions, respectively.
2.4. Estimation of Relative Water Content
The first fully expanded tender leaves of each seedling were sampled and the fresh weight (FW) was immediately measured. Subsequently, the leaves were soaked in deionized water at room temperature for 8 h and the turgid weight (TW) was then determined. Afterward, the leaves were oven-dried to a constant weight at 80 °C and the dry weight (DW) was measured. The relative water content (RWC) and RWC reducing rate (%) were calculated using the following formula [26]:
where RWCcontrol (%) indicates relative water content in the controls and RWCCd (%) indicates relative water content under Cd stress.
2.5. Determination of Biomass, Cd Concentration, and Cd Accumulation
The harvest plants were immersed in 20 mmol L−1 EDTA-Na2 solution for 15 min to remove root surface adhering ions, rinsed with ultrapure water, and separated into roots and shoots [27]. Then, the samples were completely oven-dried at 80 °C. The biomass of each sample was recorded. The dried samples (0.2 g) were digested with mixed acid (HNO3 + HClO4 (87:13, v/v)). The digested crystals were diluted with 10 mL 2.5% HNO3 solution. The Cd concentration of samples was determined by an inductively coupled plasma optical emission spectrometer (ICP-OES, Perkin Elmer, Waltham, MA, USA). The accumulation of Cd in shoot samples was calculated according to the following formula [28]:
2.6. Measurements of Long-Term WUE
Based on the above results, a cultivar (Kanlow) with high iWUE and a cultivar (Cave-in-Rock) with low iWUE were selected to determine long-term WUE (LWUE).
LWUE was calculated according to the water loss and increase in dry weight. Each time the solution was replaced, the water loss was determined. After the seedlings were sampled, all samples were dried at 80 °C for 24 h and the dry biomass of samples was recorded [29]:
where 1 g water = 1 mL.
2.7. Measurements for Abscisic Acid Content
Two cultivars (Kanlow and Cave-in-Rock) were used to determine the abscisic acid (ABA) content. ABA was extracted from the root and shoot using the method of Zhang et al. [30]. Fresh samples (0.2 g) were ground into powder with liquid nitrogen, then 2 mL 80% chromatographic methanol was added to the mortar (19% ultrapure water, 1% acetic acid), and they were ground to a homogenous mixture over ice. The homogenate was transferred to a 10 mL centrifuge tube and the mixture was extracted for 12 h at 4 °C. After centrifuging at 12,000× g for 15 min at 4 °C, the supernatant was collected and then mixed with 1 mL 80% chromatographic methanol to precipitate thoroughly (19% ultrapure water, 1% acetic acid). The mixture was centrifuged again at 12,000× g for 15 min at 4 °C and the supernatant was filtered through a 0.22 μm hydrophobic membrane. After the C18 SPE cartridge (Waters, Milford, MA, USA) was activated with 6 mL ultrapure water and 6 mL chromatographic methanol and balanced with 6 mL 80% methanol for HPLC (19% ultrapure water, 1% acetic acid), the filtrate was passed through the cartridge. Finally, the filtrate was extracted with 3 mL ethyl acetate then dried using nitrogen gas, dissolved in a 0.5 mL mobile phase (methanol: 1% acetic acid, 55:45), and stored at −20 °C.
Abscisic acid content was determined using the Agilent 1260 Infinity LC system (Agilent Technologies, Santa Clara, CA, USA) with an Agilent SB-C18 column (4.6 × 50 mm; 1.8 μm; USA). The temperature of the column heater (holding HPLC column) was 27 °C, and the wavelength was 254 nm. The mobile phase was a mixture of methanol and 1% acetic acid (55/45), and the solvent flow rate was 0.3 mL min−1. The sample size was 2 μL.
2.8. RNA Isolation and Quantitative PCR Analysis
The relative expression levels of PvERECTA and PvEPF1 in Kanlow and Cave-in-Rock were analyzed by qRT-PCR. Plant cultivation conditions were the same as those mentioned earlier in this report. Total RNA was extracted from 0.2 g of shoots with TRIzol (Invitrogen™ TRIzol™) and cDNA was generated with a reverse transcription kit (HiScript® II Q RT SuperMix for qPCR, Vazyme). The PvERECTA and PvEPF1 expression levels were quantified using a ChamQ SYBR qPCR Master Mix (Vazyme) with a 7500 PCR system (Applied Biosystems, Foster City, CA, USA). The following primers were used for the PCR amplification: PvERECTA, forward, 5′-GATGTGTGGCCACTGACGCT-3′, reverse, 5′-ACTGCAAGCAGCACGGTCAT-3′; PvEPF1, forward, 5′-CTGGTAAGTGTGCGGCTGCT-3′, reverse, 5′-TTGAGTGCGCTGGTTGTCGT-3′. The actin gene was used as an internal standard with primers, forward, 5′-GGATGGCTTTAAGCAGAATGA-3′, reverse, 5′-CAAAACGCCCAGGTCTGACT-3′. Quantification of gene expression was performed using the comparative 2−ΔΔCT method, and for each treatment performed three independent biological replicates and three technical replicates were conducted for each sample [31].
2.9. Statistical Analysis
Data were expressed as the mean values ± standard errors (SEs). Statistical analyses were performed using SPSS 20.0 and a two-way analysis of variance (ANOVA). Figures were prepared by Origin 2016. Duncan’s multiple range test was applied to assess the significant differences (p < 0.05) between the mean values.
3. Results
3.1. Effects of Cd on Instantaneous WUE (iWUE) of Switchgrass
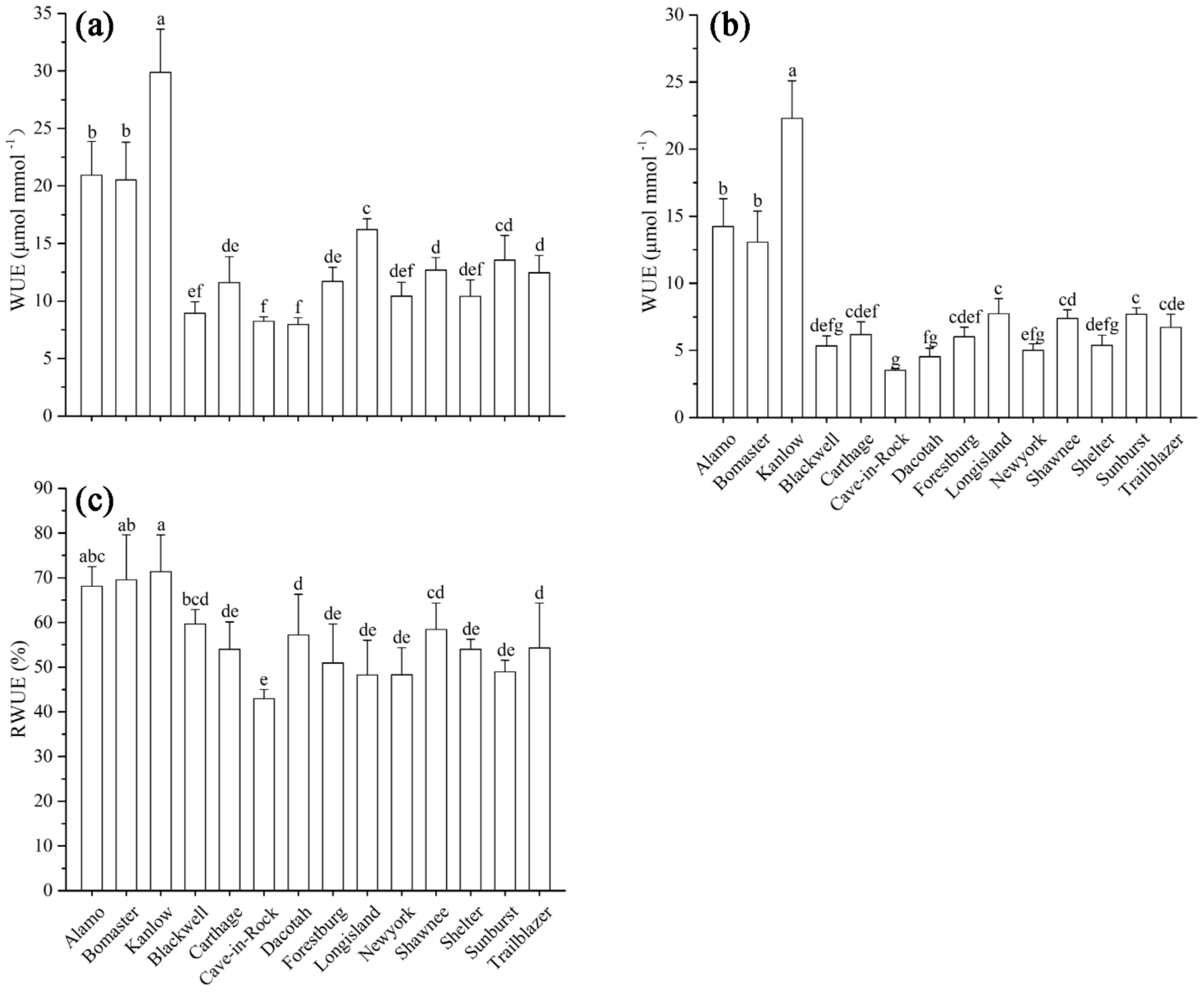
The iWUE was calculated based on Pn and E values (Figure S1). The iWUE values of different cultivars showed significant differences both in the control (Figure 1a) and Cd stress (Figure 1b) treatments. The iWUE values of the lowland-type cultivars (Alamo, Bomaster, and Kanlow) were higher than those of the upland-type cultivars under both control (Figure 1a) and Cd stress (Figure 1b) conditions. Additionally, the iWUE of Kanlow was highest, and significantly higher than those of the rest of cultivars under the control conditions (Figure 1a). Moreover, the iWUE of Kanlow was significantly higher than those of the rest of cultivars, and that of Cave-in-Rock was significantly lower than those of the remaining cultivars under Cd treatment (Figure 1b). As a result of the various iWUEs of different cultivars under control and Cd stress treatments, the RWUE of 14 cultivars of switchgrass was calculated according to the iWUE of different cultivars to further analyze the difference in response to Cd stress. The RWUE of the lowland-type switchgrass was higher than that of the upland types. The RWUE of Kanlow was the highest, while Cave-in-Rock had the lowest values (Figure 1c).
3.2. Effects of Cd on the RWC of Switchgrass
The results related to the RWC and RWC reducing rate of switchgrass under control and Cd treatment conditions are summarized in Table 2. Compared with the control, Cd stress affected the RWCs of switchgrass leaves. No significant difference occurred between the control and Cd groups among lowland types, while significant differences were exhibited among upland types. Similar trends were also observed in RWC reducing rates between lowland and upland types. The RWCs of different cultivars showed significant differences between the control and Cd stress treatments. Additionally, the RWC of Kanlow was highest, significantly higher than those of the rest of the cultivars under control and Cd stress condition. Moreover, the RWCs of Carthage and Sunburst were significantly lower than those of the remaining cultivars under control and Cd stress conditions, respectively. The RWC reducing rate of 14 cultivars of switchgrass was calculated according to the RWCs of different cultivars: the RWC reducing rate in lowland types was lower than that of upland types, and the maximum value was seen in Sunburst, then in Cave-in-Rock, while the RWC reducing rates of Kanlow were lowest (Table 2).
3.3. Cd Concentration, Cd Accumulations, and Their Relationship with RWUE
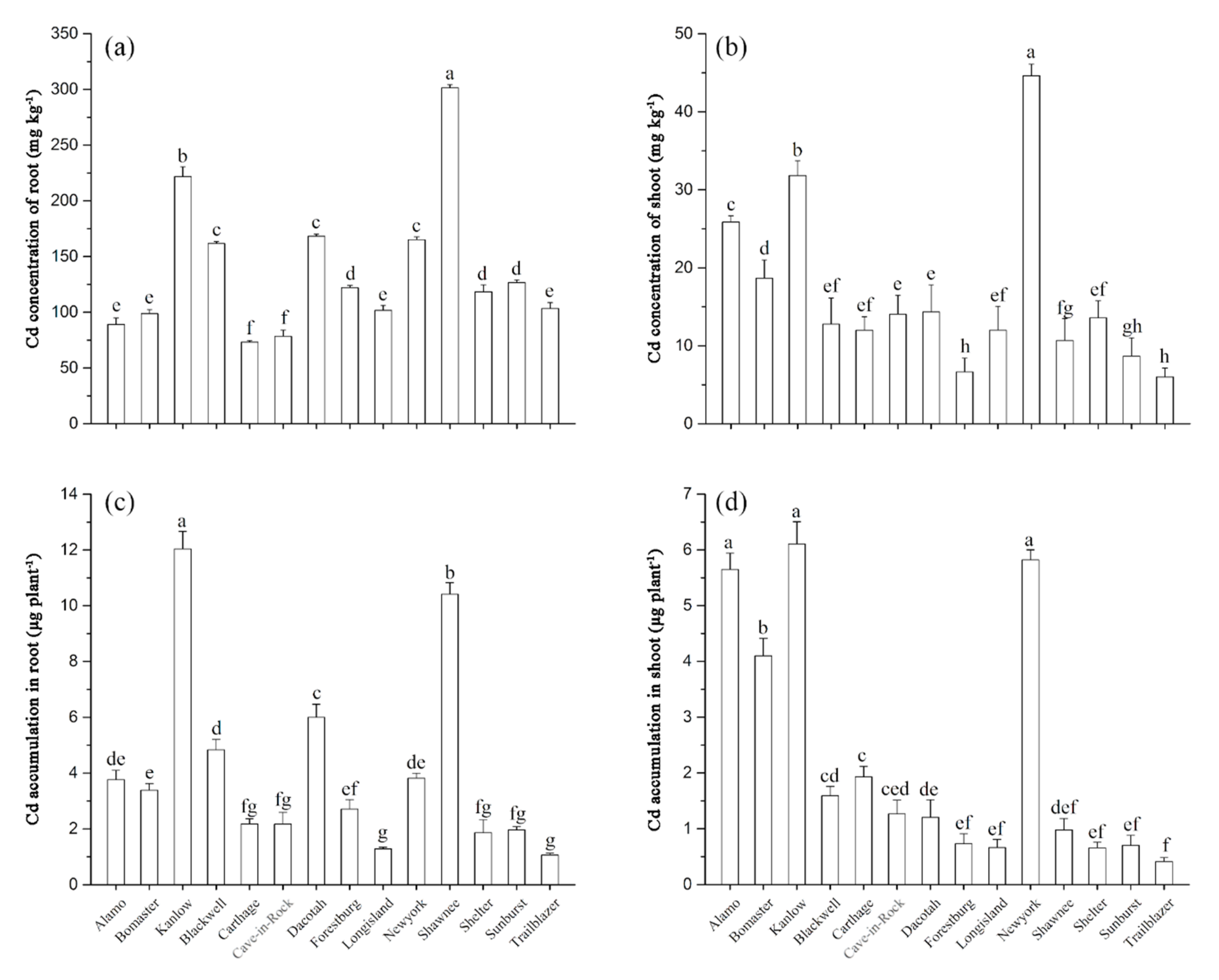
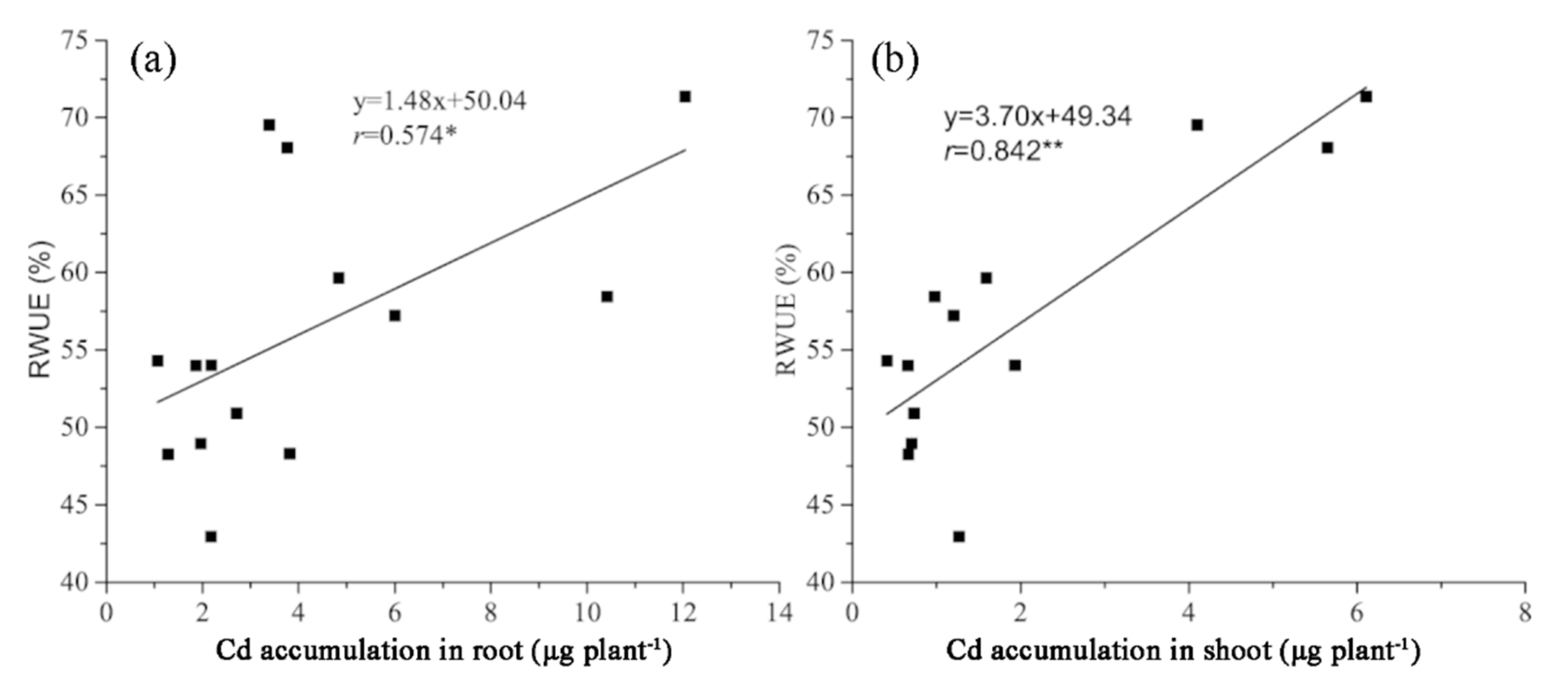
The Cd concentration in roots and shoots showed variation among cultivars (Figure 2a,b). The Cd concentration in the roots of Shawnee was significantly higher than in the other cultivars, followed by Kanlow (Figure 2a). The highest Cd concentration of shoots was found in New York, with the lowest being found in Trailblazer (Figure 2b). As for Cd accumulation calculated based on Cd concentration and biomass of plants (Figure S2), the accumulation of Cd in Kanlow was the highest in both roots and shoots (Figure 2c,d). Significant differences in Cd accumulation were observed among culti-vars (p < 0.05). A significant positive (p < 0.05) correlation was observed between the RWUE and Cd accumulations in roots (Figure 3a). Additionally, a very significant dif-ference (p < 0.01) between the RWUE and Cd accumulations in shoots was exhibited (Figure 3b).
3.4. Effects of Cd on Long-Term WUE of Switchgrass
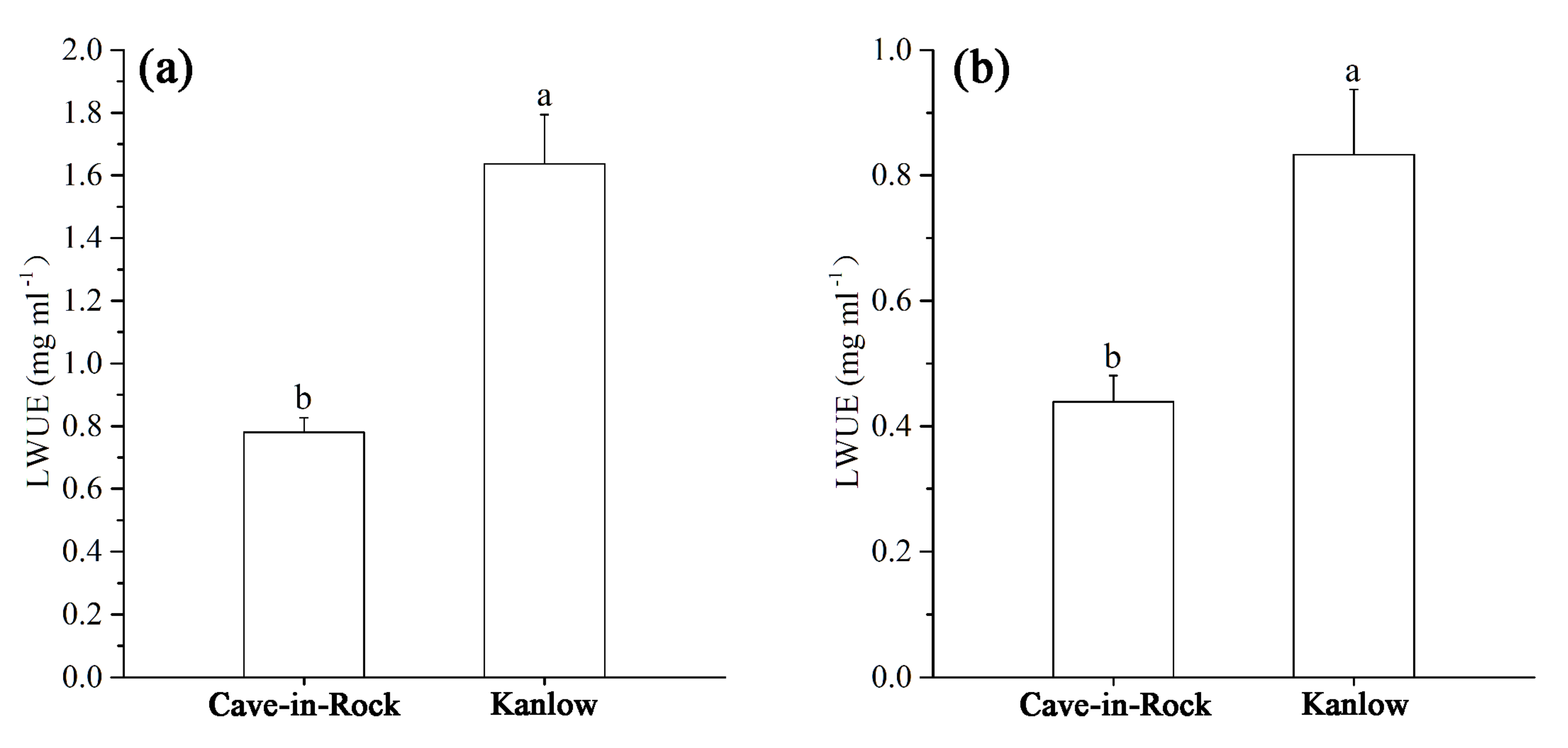
The LWUE of two cultivars with different iWUEs is shown in Figure 4. Cd stress inhibited the LWUE of both cultivars. The LWUE of Kanlow was significantly higher than that of Cave-in-Rock. As expected, Kanlow had the higher LWUE compared with Cave-in-Rock under Cd stress. Moreover, the LWUE of lowland-type switchgrass was higher than that of the upland types.
3.5. Effects of Cd on ABA Concentration of Switchgrass
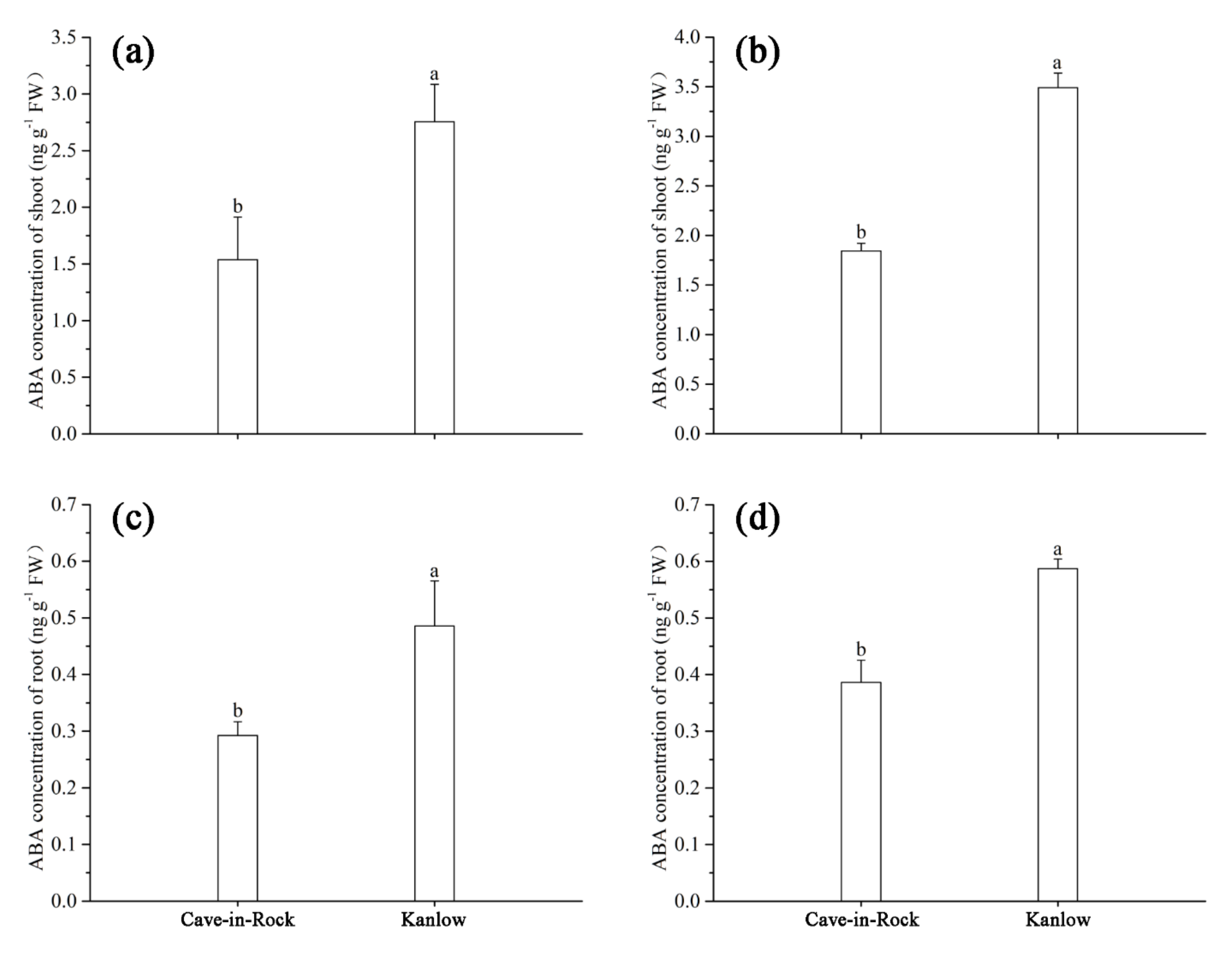
According to Figure 5, the ABA concentrations in Kanlow were significantly higher than those of Cave-in-Rock in both the roots and shoots of switchgrass under control and Cd stress conditions, and those of lowland-type Kanlow were significantly higher than those of upland-type Cave-in-Rock under both control and Cd exposure conditions (Figure 5).
3.6. Relative Expression of PvERECTA and PvEPF1
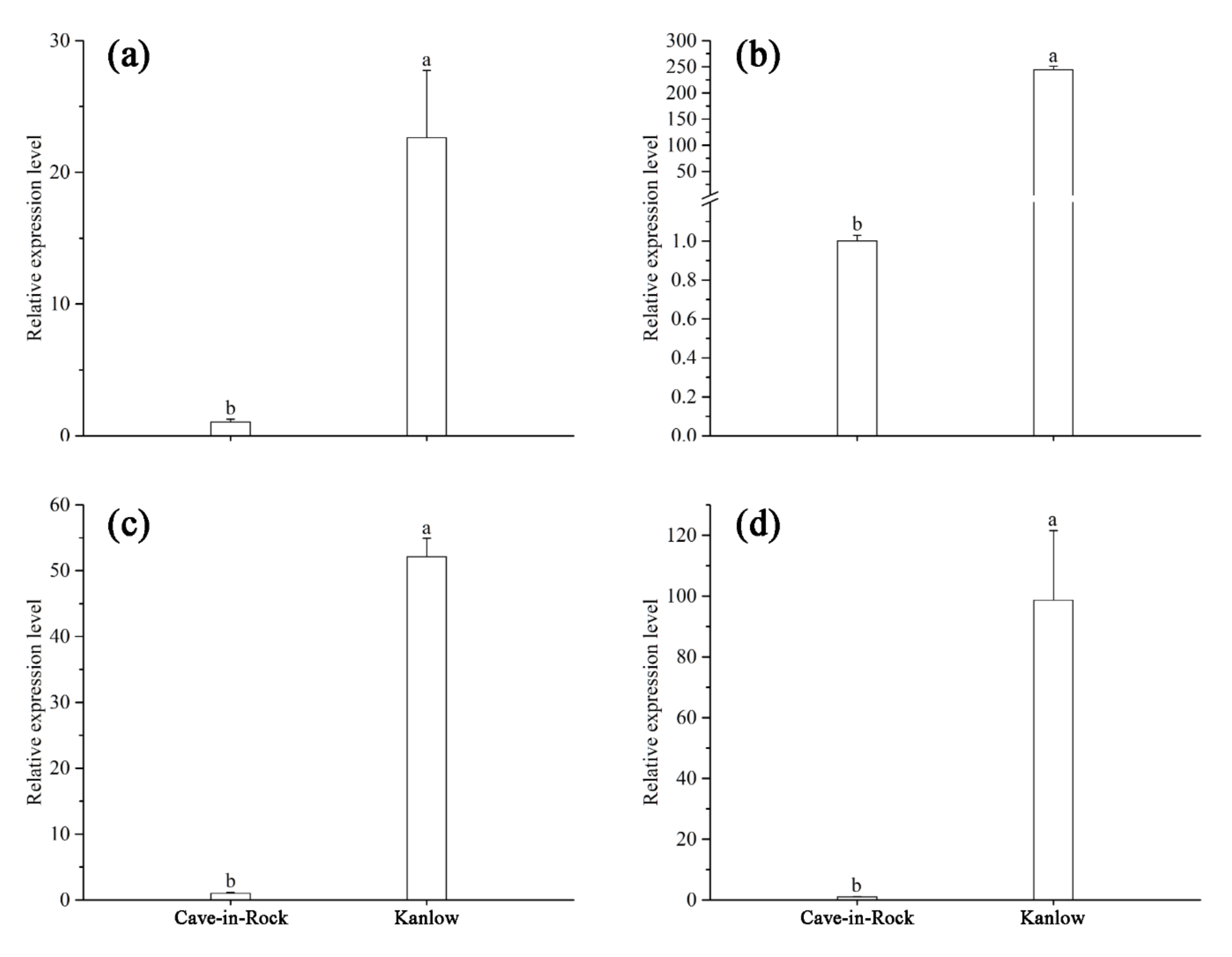
Quantitative RT-PCR was used to determine the relative expression levels of PvERECTA and PvEPF1 in the shoots of two cultivars. According to Figure 6, the relative expression levels of PvERECTA and PvEPF1 in Cave-in-Rock were significantly lower than those of PvERECTA and PvEPF1 in Kanlow, both in control and Cd stress conditions.
4. Discussion
The iWUE reflects the instantaneous carbon gain-to-water loss ratio [32]. In Silene paradoxa L. [33], Salicornia ramosissima [34], and Picris divaricata [35], several abiotic stresses, including heavy metal stress, have been shown to inhibit photosynthesis due to the limitation of leaf CO2 diffusion. In order to balance water losses and CO2 acquisition for growth, plants are prone to show a decrease in Gs under environmental stress [36,37]. In this study, the analysis of gas exchange parameters showed that Cd stress had an inhibitory effect on Gs. As the tradeoff between CO2 acquisition and water loss took place, the Pn, E, and Ci of different cultivars treated with Cd were also seriously influenced, which led to decreased iWUE. Thus, the water transport rate was decreased, as evidenced by the lowering of the water contents of the shoots [38]. In addition, the LWUE of two cultivars was measured and showed a similar trend to iWUE. The results further illustrated the reliability of iWUE calculated by Pn and E. To some extent, the decrease in RWC under Cd stress indicated diversity in the water-binding ability of different switchgrass cultivars. In our study, we found that the RWC of 14 switchgrass cultivars showed variation, irrespective of control conditions or Cd stress. The results partly agreed with Kiniry et al. [39]. The interaction between cultivar and Cd treatment in WUE and RWC did not show significant differences, meaning that the difference in WUE was related to the cultivars and that Cd influenced the WUE regardless of the cultivar. Moreover, it indicated that a cultivar with a high WUE and a high capacity for Cd accumulation has an inherent advantage. Additionally, a high iWUE was observed in lowland types, indicating that these were superior to upland types in terms of biomass gain relative to water consumption.
Stomata play a key role in modulating gas exchange between the atmosphere and the inter-cellular spaces of leaves [40]. In the Cd hyperaccumulator Picris divaricata, a decrease in Gs caused a reduction in stomatal density and interfered with guard cell function under Cd stress [35]. The increase in ABA content decreased or inhibited the stomata opening, thus reducing transpiration and water loss [41]. It was found that different plants exhibited different levels of ABA accumulation under water stress [42]. Similarly, the ABA content of switchgrass increased in response to Cd stress and in Kanlow was significantly higher than in Cave-in-Rock. It was proposed that ABA inhibits the stomatal opening, then reduces transpiration and water loss, causing the WUE to increase in switchgrass. It has also been shown that ABA is a stress hormone that is involved with various signal transduction pathways [43]. In Populus euphratica cells [44], mung bean seedlings [45], and oilseed rape [46], ABA has been shown to play a vital role in ameliorating Cd toxicity due to the increased ABA content in response to Cd stress. In this study, similar results were found in that the ABA content of three cultivars were significantly increased both in the roots and shoots under Cd stress. This was perhaps a comprehensive physiological response. The action of Cd on switchgrass incurred the increase of ABA content, which could alleviate Cd toxicity in the plant.
Stomata development has drawn the attention of many researchers. Shpak et al. [47] indicated that stomatal patterning was controlled by three ERECTA (ER)-family leucine-rich repeat receptor-like kinases (LRR-RLKs). ERECTA family genes, including ER, ERL1, and ERL2, regulate stomatal phenotypes [48]. ERECTA (ER) genes affect, but are not limited to affecting, cell–cell contact, epidermal cell expansion, and stomatal density [49,50,51]. ERECTA modulate stomatal density mainly through their role in epidermal pavement cell expansion. Epidermal patterning factors (EPFs) are a family of 11 related, small, secreted peptides. Several members of the EPF family regulate stomata formation in Arabidopsis leaves. With Arabidopsis epf mutants, plants lacking both EPF1 and EPF2 (epf1, epf2) exhibited higher stomatal density and lower iWUE [20]. The relative levels of expression of ERECTA and EPF1 homologous genes in switchgrass were detected in the shoots of different cultivars using quantitative RT-PCR. Regardless of control or Cd stress conditions, the relative expression levels of PvERECTA and PvEPF1 showed completely similar results in Kanlow and Cave-in-Rock. As a result, the density of stomata in Kanlow was reduced and the transpiration rate also decreased compared to Cave-in-Rock. The increase in relative expression levels of PvERECTA and PvEPF1 resulted in the reduction of stomatal density and an increased water absorption capacity. At the same time, Cd was more likely to be absorbed by the roots. As a result, the transpiration of switchgrass cultivars with high WUE was attenuated and photosynthesis in those cultivars was higher than in cultivars with low WUE. Therefore, plants with high WUE may accumulate more Cd in plant tissues than cultivars with low WUE.
5. Conclusions
The WUE of 14 switchgrass cultivars was illustrated in the study. Switchgrass cultivars with higher RWUE values showed higher accumulation of Cd. A change in stomata may be responsible for the diversity of WUE in switchgrass cultivars. Our results indicate that it would be possible to further select and cultivate switchgrass with a high WUE and a high capacity for Cd accumulation for developing switchgrass in Cd-contaminated land.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/agronomy12020507/s1, Figure S1: Effect of cadmium on photosynthesis of switchgrass; Figure S2: Biomass of roots and shoots of switchgrass under Cd stress.
Author Contributions
Conceptualization, Z.H., Z.F. and Q.C.; methodology, Z.H., Z.F. and B.H.; validation, Z.H., Z.F., B.H. and X.W.; formal analysis, L.L.; investigation, Q.C.; resources, Q.C.; data curation, Z.H., Z.F., B.H. and X.W.; writing—original draft preparation, Z.H. and B.H.; writing—review and editing, Z.H., Z.F. and Q.C.; supervision, Q.C.; project administration, L.L. and Q.C.; funding acquisition, Q.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Natural Science Foundation of China (NSFC), grant number 31372359.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Yuan, S.; Xu, B.; Zhang, J.; Xie, Z.; Cheng, Q.; Yang, Z.; Cai, Q.; Huang, B. Comprehensive analysis of CCCH-type zinc finger family genes facilitates functional gene discovery and reflects recent allopolyploidization event in tetraploid switchgrass. BMC Genom. 2015, 16, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balsamo, R.A.; Kelly, W.J.; Satrio, J.A.; Ruiz-Felix, M.N.; Fetterman, M.; Wynn, R.; Hagel, K. Utilization of grasses for potential biofuel production and phytoremediation of heavy metal contaminated soils. Int. J. Phytoremediat. 2015, 17, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.; Mielenz, J.R.; Xiao, X.; Ge, Y.; Hamilton, C.Y.; Rodriguez, M.; Chen, F.; Foston, M.; Ragauskas, A.; Bouton, J.; et al. Genetic manipulation of lignin reduces recalcitrance and improves ethanol production from switchgrass. Proc. Natl. Acad. Sci. USA 2011, 108, 3803–3808. [Google Scholar] [CrossRef] [Green Version]
- Schmer, M.R.; Vogel, K.P.; Mitchell, R.B.; Perrin, R.K. Net energy of cellulosic ethanol from switchgrass. Proc. Natl. Acad. Sci. USA 2008, 105, 464–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arora, K.; Sharma, S.; Monti, A. Bio-remediation of Pb and Cd polluted soils by switchgrass: A case study in India. Int. J. Phytoremediation 2015, 18, 704–709. [Google Scholar] [CrossRef]
- Wang, Q.; Gu, M.; Ma, X.; Zhang, H.; Wang, Y.; Cui, J.; Gao, W.; Gui, J. Model optimization of cadmium and accumulation in switchgrass (Panicum virgatum L.): Potential use for ecological phytoremediation in Cd-contaminated soils. Environ. Sci. Pollut. Res. 2015, 22, 16758. [Google Scholar] [CrossRef]
- Clemens, S.; Ma, J.F. Toxic heavy metal and metalloid accumulation in crop plants and foods. Annu. Rev. Plant Biol. 2016, 67, 489–512. [Google Scholar] [CrossRef] [Green Version]
- Duan, G.; Shao, G.; Tang, Z.; Chen, H.; Wang, B.; Tang, Z.; Yang, Y.; Liu, Y.; Zhao, F.J. Genotypic and environmental variations in grain cadmium and arsenic concentrations among a panel of high yielding rice cultivars. Rice 2017, 10, 9. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.J.; Ma, Y.; Zhu, Y.G.; Tang, Z.; McGrath, S.P. Soil contamination in China: Current status and mitigation strategies. Environ. Sci. Technol. 2015, 49, 750–759. [Google Scholar] [CrossRef]
- The Ministry of Environmental Protection. The Ministry of Land and Resources Report on the National Soil Contamination Survey. Available online: http://www.mee.gov.cn/gkml/sthjbgw/qt/201404/W020140417558995804588.pdf (accessed on 17 April 2014).
- He, S.; Yang, X.; He, Z.; Baligar, V.C. Morphological and physiological responses of plants to cadmium toxicity: A Review. Pedosphere 2017, 27, 421–438. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Adrees, M.; Ibrahim, M.; Tsang, D.C.W.; Zia-Ur-Rehman, M.; Zahir, Z.A.; Rinklebe, J.; Tack, F.M.G.; Ok, Y.S. A critical review on effects, tolerance mechanisms and management of cadmium in vegetables. Chemosphere 2017, 182, 90–105. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Wang, R.; Zhang, X.; Yu, Y.; Zhao, R.; Li, Z.; Chen, S. Hydrogen sulfide alleviates cadmium toxicity through regulations of cadmium transport across the plasma and vacuolar membranes in Populus euphratica cells. Plant Physiol. Bioch. 2013, 65, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, R.; Bertrand, A.; Reis, R.; Mourato, M.P.; Martins, L.L.; Gonzalez, A. Growth and physiological responses to cadmium stress of two populations of Dittrichia viscosa (L.) Greuter. J. Hazard. Mater. 2013, 244–245, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Kupper, H.; Parameswaran, A.; Leitenmaier, B.; Trtilek, M.; Setlik, I. Cadmium-induced inhibition of photosynthesis and long-term acclimation to cadmium stress in the hyperaccumulator Thlaspi caerulescens. New Phytol. 2007, 175, 655–674. [Google Scholar] [CrossRef]
- Benakova, M.; Ahmadi, H.; Ducaiova, Z.; Tylova, E.; Clemens, S.; Tuma, J. Effects of Cd and Zn on physiological and anatomical properties of hydroponically grown Brassica napus plants. Environ. Sci. Pollut. Res. 2017, 24, 20705–20716. [Google Scholar] [CrossRef]
- Ruggiero, A.; Punzo, P.; Landi, S.; Costa, A.; Oosten, M.; Grillo, S. Improving plant water use efficiency through molecular genetics. Horticulturae 2017, 3, 31. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, M.; Sallam, A.; Munir, M.; Ali-Dinar, H. Effects of deficit irrigation scheduling on water use, gas exchange, yield, and fruit quality of date palm. Agronomy 2021, 11, 2256. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [Green Version]
- Franks, P.J.; Doheny-Adams, T.; Britton-Harper, Z.J.; Gray, J.E. Increasing water-use efficiency directly through genetic manipulation of stomatal density. New Phytol. 2015, 207, 188–195. [Google Scholar] [CrossRef]
- Liu, C.; Lou, L.; Deng, J.; Li, D.; Yuan, S.; Cai, Q. Morph-physiological responses of two switchgrass (Panicum virgatum L.) cultivars to cadmium stress. Grassl. Sci. 2016, 62, 92–101. [Google Scholar] [CrossRef]
- Pandey, V.C.; Bajpai, O.; Singh, N. Energy crops in sustainable phytoremediation. Renew. Sustain. Energy Rev. 2016, 54, 58–73. [Google Scholar] [CrossRef]
- Wullschleger, S.D.; Davis, E.B.; Borsuk, M.E.; Gunderson, C.A.; Lynd, L. Biomass production in switchgrass across the United States: Database description and determinants of yield. Agron. J. 2010, 102, 1158–1168. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.H.; Wang, Y.; Wang, J.W.; Babla, M.; Zhao, C.; Garcia-Mata, C.; Sani, E.; Differ, C.; Mak, M.; Hills, A.; et al. Nitrate reductase mutation alters potassium nutrition as well as nitric oxide-mediated control of guard cell ion channels in Arabidopsis. New Phytol. 2016, 209, 1456–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, H.; Wu, L.; Filardo, F.; Yang, X.; Zhao, X.; Fu, D. Chemical and hydraulic signals regulate stomatal behavior and photosynthetic activity in maize during progressive drought. Acta Physiol. Plant 2017, 39, 125. [Google Scholar] [CrossRef]
- Shi, G.; Xia, S.; Ye, J.; Huang, Y.; Liu, C.; Zhang, Z. PEG-simulated drought stress decreases cadmium acccumulation in castor bean by altering root morphology. Environ. Exp. Bot. 2015, 111, 127–134. [Google Scholar] [CrossRef]
- Yoon, J.; Cao, X.; Zhou, Q.; Ma, L.Q. Accumulation of Pb, Cu, and Zn in native plants growing on a contaminated Florida site. Sci. Total Environ. 2006, 368, 456–464. [Google Scholar] [CrossRef]
- Fang, Z.; Lou, L.; Tai, Z.; Wang, Y.; Yang, L.; Hu, Z.; Cai, Q. Comparative study of Cd uptake and tolerance of two Italian ryegrass (Lolium multiflorum) cultivars. PeerJ 2017, 5, e3621. [Google Scholar] [CrossRef] [Green Version]
- Wituszynska, W.; Karpiński, S. Determination of Water Use Efficiency for Arabidopsis thaliana. Bio-Protocol 2014, 4, e1041. [Google Scholar] [CrossRef]
- Zhang, H.J.; Zhang, N.; Yang, R.C.; Wang, L.; Sun, Q.Q.; Li, D.B.; Cao, Y.Y.; Weeda, S.; Zhao, B.; Ren, S. Melatonin promotes seed germination under high salinity by regulating antioxidant systems, ABA and GA4 interaction in cucumber (Cucumis sativus L.). J. Pineal Res. 2014, 57, 269–279. [Google Scholar] [CrossRef]
- Song, G.; Yuan, S.; Wen, X.; Xie, Z.; Lou, L.; Hu, B.; Cai, Q.; Xu, B. Transcriptome analysis of Cd-treated switchgrass root revealed novel transcripts and the importance of HSF/HSP network in switchgrass Cd tolerance. Plant Cell Rep. 2018, 37, 1485–1497. [Google Scholar] [CrossRef]
- Cao, X.; Jia, J.B.; Li, H.; Li, M.C.; Luo, J.; Liang, Z.S.; Liu, T.X.; Liu, W.G.; Peng, C.H.; Luo, Z.B. Photosynthesis, water use efficiency and stable carbon isotope composition are associated with anatomical properties of leaf and xylem in six poplar species. Plant Biol. 2012, 14, 612–620. [Google Scholar] [CrossRef] [PubMed]
- Bazihizina, N.; Colzi, I.; Giorni, E.; Mancuso, S.; Gonnelli, C. Photosynthesizing on metal excess: Copper differently induced changes in various photosynthetic parameters in copper tolerant and sensitive Silene paradoxa L. populations. Plant Sci. 2015, 232, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Perez-Romero, J.A.; Redondo-Gomez, S.; Mateos-Naranjo, E. Growth and photosynthetic limitation analysis of the Cd-accumulator Salicornia ramosissima under excessive cadmium concentrations and optimum salinity conditions. Plant Physiol. Biochem. 2016, 109, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Ying, R.R.; Jiang, D.; Zeng, X.W.; Morel, J.L.; Tang, Y.T.; Qiu, R.L. Impaired leaf CO2 diffusion mediates Cd-induced inhibition of photosynthesis in the Zn/Cd hyperaccumulator Picris divaricata. Plant Physiol. Biochem. 2013, 73, 70–76. [Google Scholar] [CrossRef]
- Flexas, J.; Baron, M.; Bota, J.; Ducruet, J.M.; Galle, A.; Galmes, J.; Jimenez, M.; Pou, A.; Ribas-Carbo, M.; Sajnani, C.; et al. Photosynthesis limitations during water stress acclimation and recovery in the drought-adapted Vitis hybrid Richter-110 (V. berlandieri x V. rupestris). J. Exp. Bot. 2009, 60, 2361–2377. [Google Scholar] [CrossRef] [Green Version]
- Sagardoy, R.; Vazquez, S.; Florez-Sarasa, I.D.; Albacete, A.; Ribas-Carbo, M.; Flexas, J.; Abadia, J.; Morales, F. Stomatal and mesophyll conductances to CO2 are the main limitations to photosynthesis in sugar beet (Beta vulgaris) plants grown with excess zinc. New Phytol. 2010, 187, 145–158. [Google Scholar] [CrossRef]
- Fahr, M.; Laplaze, L.; Bendaou, N.; Hocher, V.; Mzibri, M.E.; Bogusz, D.; Smouni, A. Effect of lead on root growth. Front. Plant Sci. 2013, 4, 175. [Google Scholar] [CrossRef] [Green Version]
- Kiniry, J.R.; Lynd, L.; Greene, N.; Johnson, M.V.V.; Casler, M.; Laser, M.S. Biofuels and water use: Comparison of maize and switchgrass and general perspectives. In New Research on Biofuels; Wright, J.H., Evans, D.A., Eds.; Nova Science Publishers: New York, NY, USA, 2008; pp. 17–30. [Google Scholar]
- Azoulay-Shemer, T.; Palomares, A.; Bagheri, A.; Israelsson-Nordstrom, M.; Engineer, C.B.; Bargmann, B.O.; Stephan, A.B.; Schroeder, J.I. Guard cell photosynthesis is critical for stomatal turgor production, yet does not directly mediate CO2-and ABA-induced stomatal closing. Plant J. 2015, 83, 567–581. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.H.; Hills, A.; Batz, U.; Amtmann, A.; Lew, V.L.; Blatt, M.R. Systems dynamic modeling of the stomatal guard cell predicts emergent behaviors in transport, signaling, and volume control. Plant Physiol. 2012, 159, 1235–1251. [Google Scholar] [CrossRef] [Green Version]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Molecular responses to dehydration and low temperature: Differences and cross-talk between two stress signaling pathways. Curr. Opin. Plant Biol. 2000, 3, 217–223. [Google Scholar] [CrossRef]
- Pompeu, G.B.; Vilhena, M.B.; Gratao, P.L.; Carvalho, R.F.; Rossi, M.L.; Martinelli, A.P.; Azevedo, R.A. Abscisic acid-deficient sit tomato mutant responses to cadmium-induced stress. Protoplasma 2017, 254, 771–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Wang, S.; Zhao, N.; Deng, S.; Zhao, C.; Li, N.; Sun, J.; Zhao, R.; Yi, H.; Shen, X.; et al. Exogenous Abscisic Acid Alleviates Cadmium Toxicity by Restricting Cd2+ Influx in Populus euphratica Cells. J. Plant Growth Regul. 2016, 35, 827–837. [Google Scholar] [CrossRef]
- Li, S.W.; Leng, Y.; Feng, L.; Zeng, X.Y. Involvement of abscisic acid in regulating antioxidative defense systems and IAA-oxidase activity and improving adventitious rooting in mung bean (Vigna radiata L.) Wilczek seedlings under cadmium stress. Environ. Sci. Pollut. Res. 2014, 21, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Filardo, F.; Hu, X.; Zhao, X.; Fu, D. Cadmium stress alters the redox reaction and hormone balance in oilseed rape (Brassica napus L.) leaves. Environ. Sci. Pollut. Res. 2016, 23, 3758–3769. [Google Scholar] [CrossRef]
- Shpak, E.D.; McAbee, J.M.; Pillitteri, L.J.; Torii, K.U. Stomatal patterning and differentiation by synergistic interactions of receptor kinases. Science 2005, 309, 290–293. [Google Scholar] [CrossRef]
- Wang, Y.; Xue, X.; Zhu, J.-K.; Dong, J. Demethylation of ERECTA receptor genes by IBM1 histone demethylase affects stomatal development. Development 2016, 143, 4452–4461. [Google Scholar] [CrossRef] [Green Version]
- Ho, C.M.; Paciorek, T.; Abrash, E.; Bergmann, D.C. Modulators of stomatal lineage signal transduction alter membrane contact sites and reveal specialization among ERECTA kinases. Dev. Cell 2016, 38, 345–357. [Google Scholar] [CrossRef] [Green Version]
- Zanten, M.; Snoek, L.B.; Proveniers, M.C.; Peeters, A.J. The many functions of ERECTA. Trends Plant Sci. 2009, 14, 214–218. [Google Scholar] [CrossRef]
- Torii, K.U.; Mitsukawa, N.; Oosumi, T.; Matsuura, Y.; Yokoyama, R.; Whittier, R.F.; Komeda, Y. The Arabidopsis ERECTA gene encodes a putative receptor protein kinase with extracellular leucine-rich repeats. Plant Cell 1996, 8, 735–746. [Google Scholar]
Figure 1.
The iWUE and RWUE of switchgrass: iWUE under control (a) and Cd stress (b) conditions; RWUE (c). Different letters above the bars were significantly different at p < 0.05 (Duncan’s multiple range test). Values are means ± SE (n = 6).
Figure 1.
The iWUE and RWUE of switchgrass: iWUE under control (a) and Cd stress (b) conditions; RWUE (c). Different letters above the bars were significantly different at p < 0.05 (Duncan’s multiple range test). Values are means ± SE (n = 6).

Figure 2.
Cd concentration in roots (a) and shoots (b) and accumulations in roots (c) and shoots (d) of switchgrass under Cd stress. Different letters above the bars indicate significant differences at p < 0.05 (Duncan’s multiple range test). Values are means ± SE (n = 3).
Figure 2.
Cd concentration in roots (a) and shoots (b) and accumulations in roots (c) and shoots (d) of switchgrass under Cd stress. Different letters above the bars indicate significant differences at p < 0.05 (Duncan’s multiple range test). Values are means ± SE (n = 3).

Figure 3.
Correlation between RWUE and Cd accumulations in roots (a) and shoots (b) of different switchgrass cultivars under Cd stress. * p < 0.05, ** p < 0.01 (Duncan’s multiple range test).
Figure 3.
Correlation between RWUE and Cd accumulations in roots (a) and shoots (b) of different switchgrass cultivars under Cd stress. * p < 0.05, ** p < 0.01 (Duncan’s multiple range test).

Figure 4.
Effect of cadmium on long-term WUE of switchgrass: LWUE under control (a) and Cd stress (b) conditions. Different letters above the bars indicate significant differences at p < 0.05 (Duncan’s multiple range test). Values are means ± SE (n = 3).
Figure 4.
Effect of cadmium on long-term WUE of switchgrass: LWUE under control (a) and Cd stress (b) conditions. Different letters above the bars indicate significant differences at p < 0.05 (Duncan’s multiple range test). Values are means ± SE (n = 3).

Figure 5.
ABA concentration of switchgrass in shoots (a) and roots (c) under control conditions and in shoots (b) and roots (d) under Cd stress conditions. Values are means ± SE (n = 3). Different letters above the bars were significantly different at p < 0.05 (Duncan’s multiple range test).
Figure 5.
ABA concentration of switchgrass in shoots (a) and roots (c) under control conditions and in shoots (b) and roots (d) under Cd stress conditions. Values are means ± SE (n = 3). Different letters above the bars were significantly different at p < 0.05 (Duncan’s multiple range test).

Figure 6.
qRT-PCR analysis of PvERECTA under control (a) and Cd stress (b) conditions, and PvEPF1 under control (c) and Cd stress (d) conditions expressed in the shoots of switchgrass. Values are means ± SE (n = 3). Different letters above the bars were significantly different at p < 0.05 (Duncan’s test).
Figure 6.
qRT-PCR analysis of PvERECTA under control (a) and Cd stress (b) conditions, and PvEPF1 under control (c) and Cd stress (d) conditions expressed in the shoots of switchgrass. Values are means ± SE (n = 3). Different letters above the bars were significantly different at p < 0.05 (Duncan’s test).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Switchgrass cultivars used in the experiment.
| No. | Cultivar | Ecotype | No. | Cultivar | Ecotype |
|---|---|---|---|---|---|
| 1 | Alamo | Lowland | 8 | Forestburg | Upland |
| 2 | Bomaster | Lowland | 9 | Long Island | Upland |
| 3 | Kanlow | Lowland | 10 | New York | Upland |
| 4 | Blackwell | Upland | 11 | Shawnee | Upland |
| 5 | Carthage | Upland | 12 | Shelter | Upland |
| 6 | Cave-in-Rock | Upland | 13 | Sunburst | Upland |
| 7 | Dacotah | Upland | 14 | Trailblazer | Upland |
Table 2.
Relative water content (RWC) and RWC reducing rate of switchgrass under Cd stress.
| Cultivar | RWCcontrol (%) | RWCCd (%) | RWC Reducing Rate (%) |
|---|---|---|---|
| Alamo | 90.06 ± 0.95 ab | 88.13 ± 0.33 ab | 1.91 ± 0.56 cd |
| Bomaster | 89.18 ± 0.62 abc | 87.40 ± 0.57 ab | 1.99 ± 0.47 cd |
| Kanlow | 90.23 ± 0.49 a | 88.88 ± 0.26 a | 1.71 ± 0.31 d |
| Blackwell | 87.11 ± 0.69 cd | 83.82 ± 0.70 cde | 3.76 ± 0.56 abc |
| Carthage | 84.63 ± 0.87 d | 82.58 ± 0.3 de | 2.39 ± 0.53 bcd |
| Cave-in-Rock | 90.7 ± 0.57 a | 86.97 ± 0.98 b | 4.10 ± 0.62 ab |
| Dacotah | 86.12 ± 1.07 d | 84.00 ± 0.58 cde | 2.85 ± 0.63 bcd |
| Forestburg | 86.38 ± 0.95 cd | 83.42 ± 0.35 cde | 3.41 ± 0.57 bcd |
| Long Island | 87.43 ± 1.25 bcd | 84.62 ± 0.20 c | 3.17 ± 0.70 bcd |
| New York | 87.23 ± 1.30 cd | 84.48 ± 0.33 cd | 3.51 ± 0.72 bcd |
| Shawnee | 90.54 ± 0.30 a | 88.77 ± 0.51 ab | 1.95 ± 0.33 cd |
| Shelter | 86.58 ± 0.81 cd | 83.08 ± 0.65 cde | 4.03 ± 0.59 ab |
| Sunburst | 87.08 ± 0.58 cd | 82.41 ± 0.92 e | 5.36 ± 0.62 a |
| Trailblazer | 87.32 ± 1.09 cd | 83.83 ± 0.82 cde | 3.97 ± 0.76 ab |
RWCcontrol (%) indicates relative water content in control; RWCCd (%) indicates relative water content under Cd stress. Values (mean ± SE, n = 3) followed by different letters in the same columns are significantly different according to Duncan’s multiple range test (p < 0.05).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hu, Z.; Fang, Z.; Hu, B.; Wen, X.; Lou, L.; Cai, Q. Profiling of Water-Use Efficiency in Switchgrass (Panicum virgatum L.) and the Relationship with Cadmium Accumulation. Agronomy 2022, 12, 507. https://doi.org/10.3390/agronomy12020507
AMA Style
Hu Z, Fang Z, Hu B, Wen X, Lou L, Cai Q. Profiling of Water-Use Efficiency in Switchgrass (Panicum virgatum L.) and the Relationship with Cadmium Accumulation. Agronomy. 2022; 12(2):507. https://doi.org/10.3390/agronomy12020507
Chicago/Turabian StyleHu, Zhaoyang, Zhigang Fang, Bingyu Hu, Xuehui Wen, Laiqing Lou, and Qingsheng Cai. 2022. "Profiling of Water-Use Efficiency in Switchgrass (Panicum virgatum L.) and the Relationship with Cadmium Accumulation" Agronomy 12, no. 2: 507. https://doi.org/10.3390/agronomy12020507
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.