
Growth and Antioxidant Responses of Lettuce (Lactuca sativa L.) to Arbuscular Mycorrhiza Inoculation and Seaweed Extract Foliar Application

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Morphological Traits
2.2. Chlorophyll Index Determination
2.3. Root Colonization
2.4. Total Protein Content
2.5. Total Carbohydrate Content
2.6. Total Antioxidant Activity (TAA)
2.7. Ascorbic Acid Content
2.8. Total Phenolics Content
2.9. Total Flavonoids Content
2.10. Statistical Analysis
3. Results
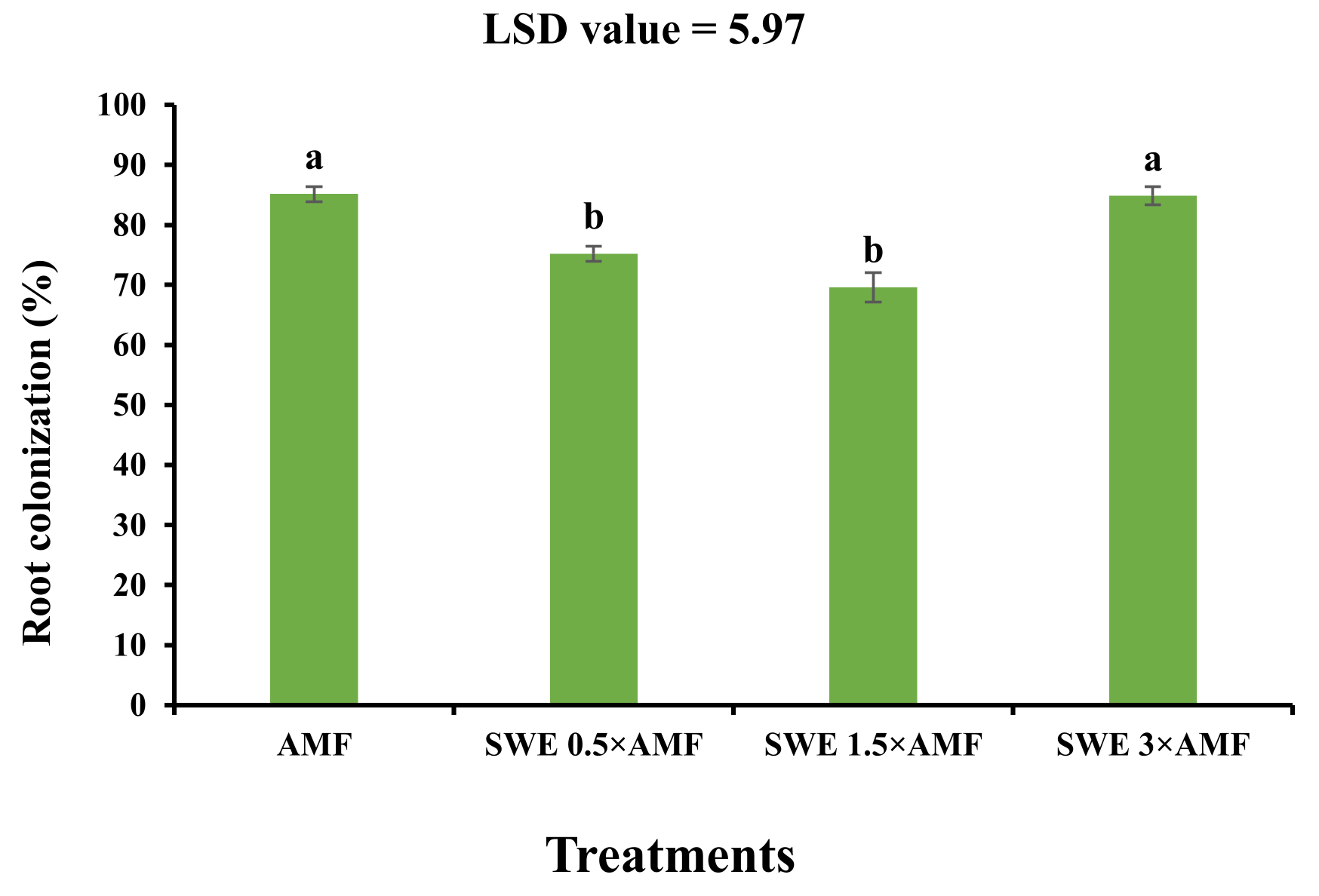
3.1. Root Colonization Percentage
3.2. Morphological Traits
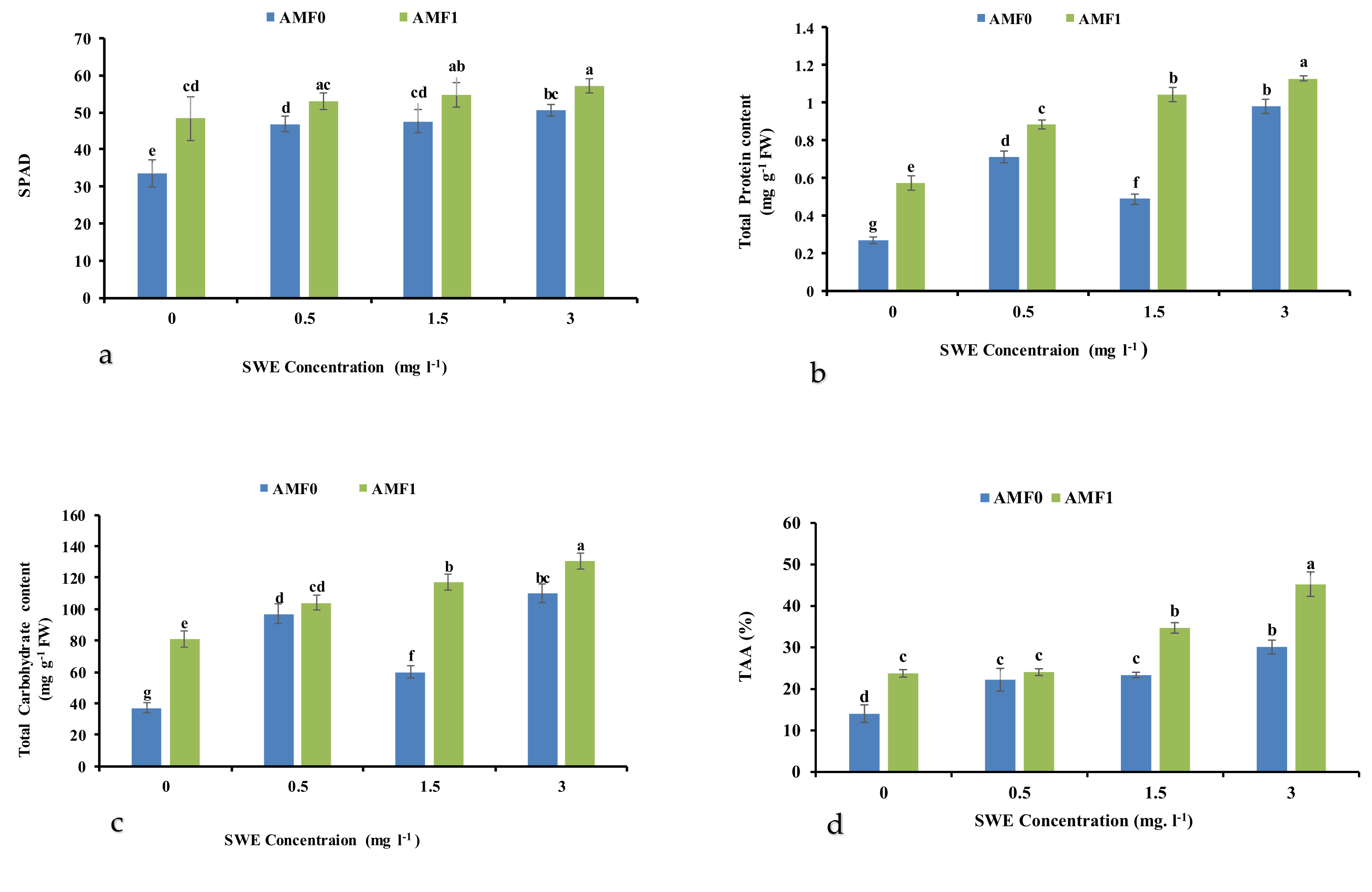
3.3. Chlorophyll Index (SPAD)
3.4. Total Protein Content
3.5. Total Carbohydrate Content
3.6. Total Antioxidant Activity (TAA)
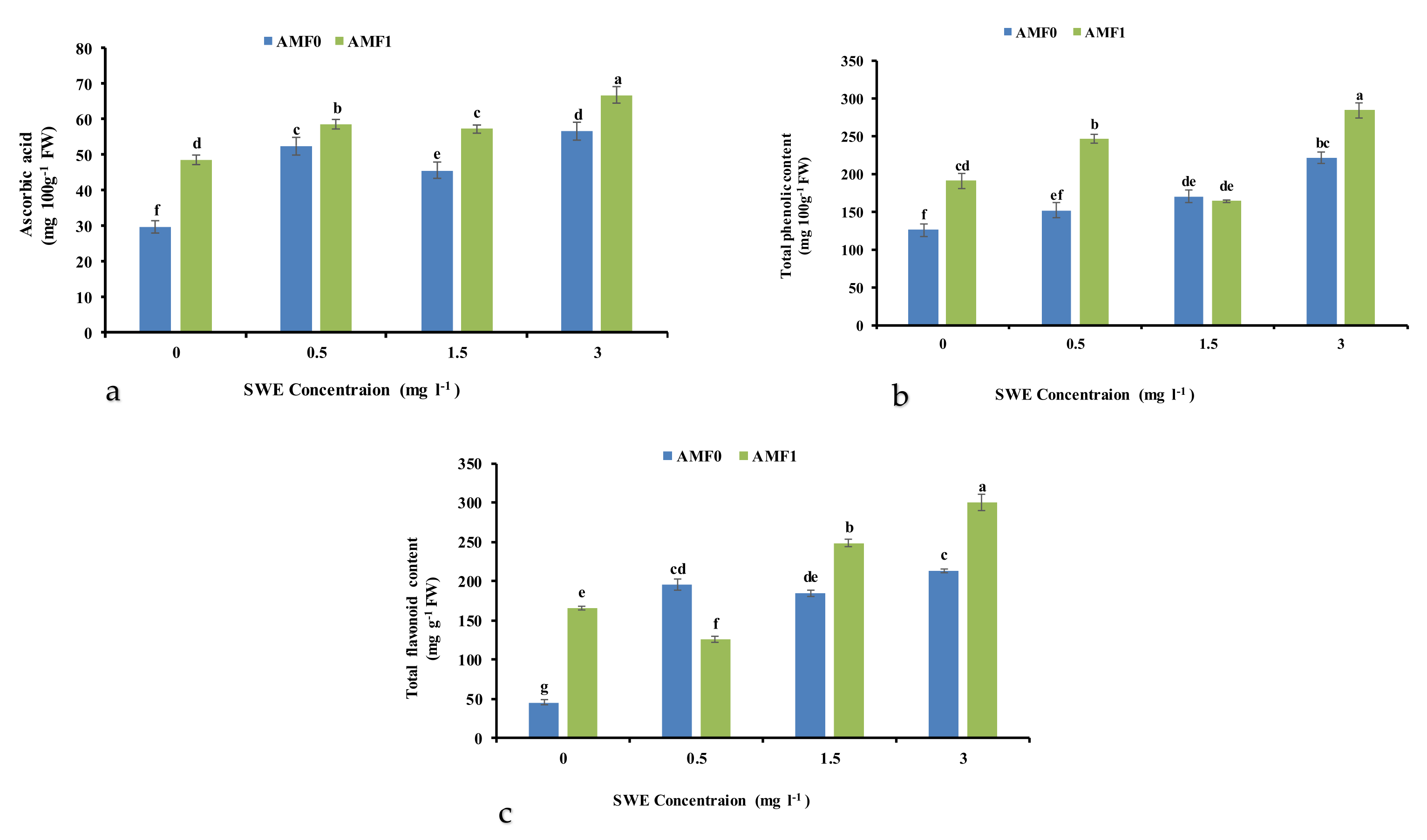
3.7. Ascorbic Acid Content
3.8. Total Phenolic and Flavonoid Content
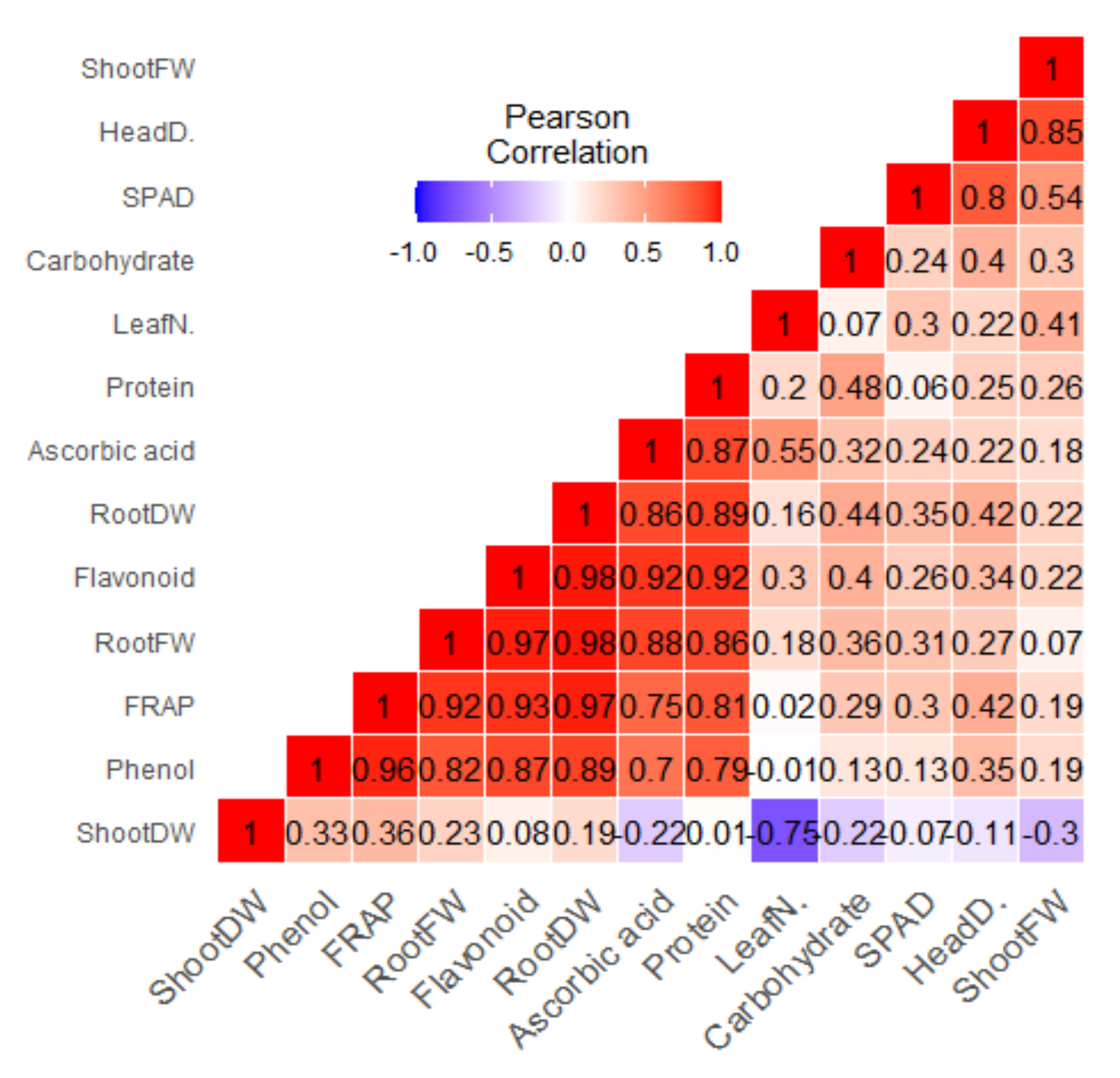
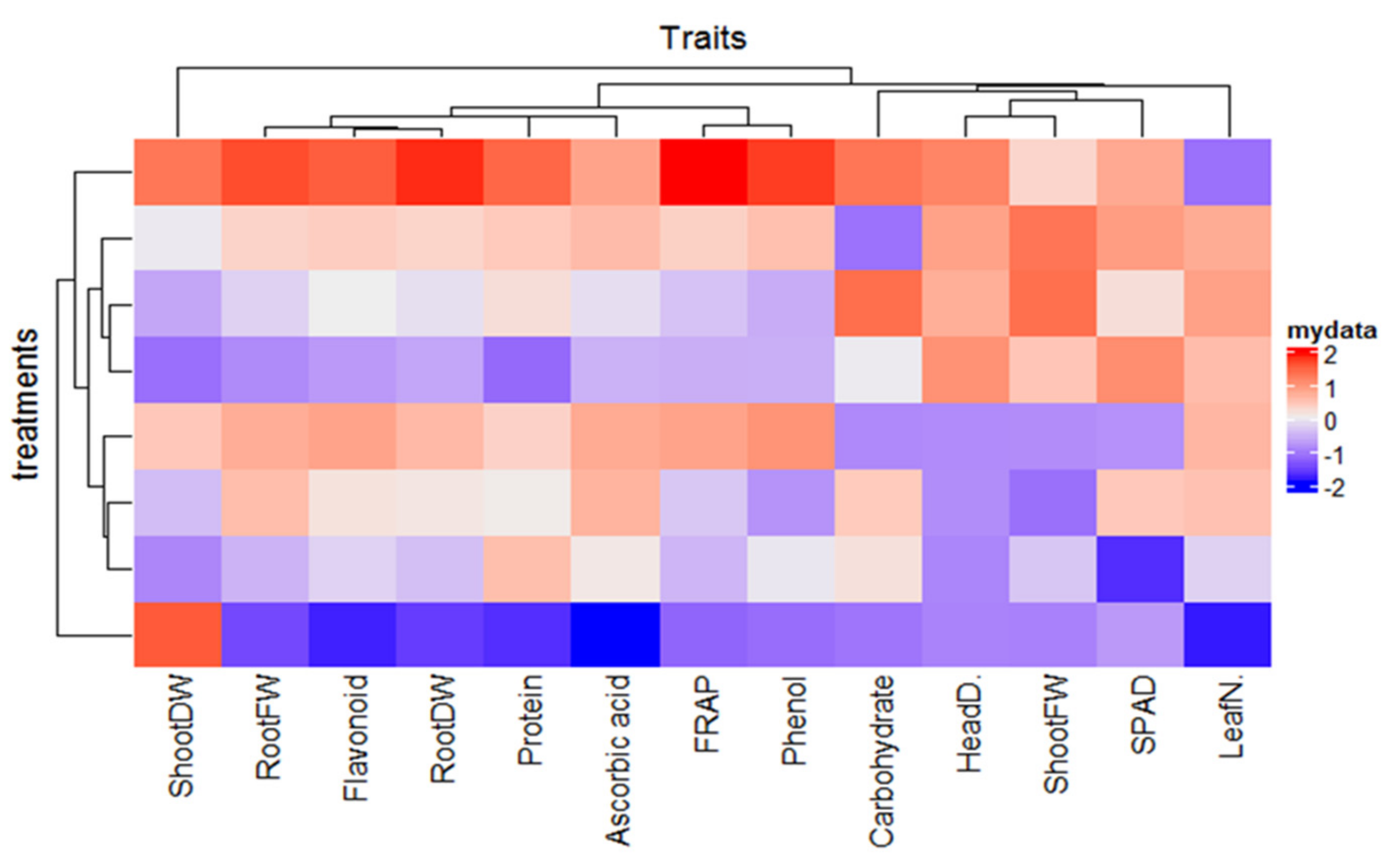
3.9. Correlation Matrix and Relative Expressions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, M.J.; Moon, Y.; Tou, J.C.; Mou, B.; Waterland, N.L. Nutritional value, bioactive compounds and health benefits of lettuce (Lactuca sativa L.). J. Food Compost. Anal. 2016, 49, 19–34. [Google Scholar] [CrossRef]
- FAOSTAT. UN Food and Agriculture Organization, Statistics Division. Production of Lettuce and Chicory by Countries. 2019. Available online: https://www.fao.org/faostat/en/#home (accessed on 4 March 2021).
- Agarwal, P.; Gupta, R.; Gill, I.K. Importance of biofertilizers in agriculture biotechnology. Ann. Biolog. Res. 2018, 9, 1–3. [Google Scholar]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in plant science: A global perspective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouphael, Y.; Colla, G. Biostimulants in agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef] [Green Version]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Battacharyya, D.; Babgohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed extracts as biostimulants in horticulture. Sci. Hortic. 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Türkmen, M.; Su, A. The effect of sea lettuce (Ulva lactuca) liquid fertilizer and zeolite combinations on the development of cucumber (Cucumis sativus). TURJAF. 2019, 7, 1021–1027. [Google Scholar] [CrossRef] [Green Version]
- Di Mola, I.; Cozzolino, E.; Ottaiano, L.; Giordano, M.; Rouphael, Y.; Colla, G.; Mori, M. Effect of vegetal-and seaweed extract-based biostimulants on agronomical and leaf quality traits of plastic tunnel-grown baby lettuce under four regimes of nitrogen fertilization. Agronomy 2019, 9, 571. [Google Scholar] [CrossRef] [Green Version]
- Chrysargyris, A.; Xylia, P.; Anastasiou, M.; Pantelides, I.; Tzortzakis, N. Effects of Ascophyllum nodosum seaweed extracts on lettuce growth, physiology and fresh-cut salad storage under potassium deficiency. J. Sci. Food Agric. 2018, 98, 5861–5872. [Google Scholar] [CrossRef]
- Jung, H.Y.; Kim, J.K. Complete reutilization of mixed mackerel and brown seaweed wastewater as a high-quality biofertiliser in open-flow lettuce hydroponics. J. Clean. Prod. 2020, 247, 119081. [Google Scholar] [CrossRef]
- Varma, A.; Prasad, R.; Tuteja, N. Mycorrhiza-Nutrient Uptake, Biocontrol, Ecorestoration; Springer: Berlin/Heidelberg, Germany, 2018; pp. 1–539. [Google Scholar]
- Santander, C.; Aroca, R.; Ruiz-Lozano, J.M.; Olave, J.; Cartes, P.; Borie, F.; Cornejo, P. arbuscular mycorrhiza effects on plant performance under osmotic stress. Mycorrhiza 2017, 27, 639–657. [Google Scholar] [CrossRef] [PubMed]
- Cobb, A.B. From Soil Ecology to Human Nutrition: Crop Symbiosis with Arbuscular Mycorrhizal Fungi in Agroecosystems. Ph.D. Thesis, Oklahoma State University, Stillwater, OK, USA, 2016; pp. 10–127. [Google Scholar]
- Bona, E.; Cantamessa, S.; Massa, N.; Manassero, P.; Marsano, F.; Copetta, A.; Lingua, G.; D’Agostino, G.; Gamalero, E.; Berta, G. Arbuscular mycorrhizal fungi and plant growth-promoting pseudomonads improve yield, quality and nutritional value of tomato: A field study. Mycorrhiza 2017, 27, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Saia, S.; Colla, G.; Raimondi, G.; Di Stasio, E.; Cardarelli, M.; Bonini, P.; Rouphael, Y. An endophytic fungi-based biostimulant modulated lettuce yield, physiological and functional quality responses to both moderate and severe water limitation. Sci. Hortic. 2019, 256, 108595. [Google Scholar] [CrossRef]
- Tarraf, W.; Ruta, C.; Tagarelli, A.; De Cillis, F.; De Mastro, G. Influence of arbuscular mycorrhizae on plant growth, essential oil production and phosphorus uptake of Salvia officinalis L. Ind. Crops Prod. 2017, 102, 144–153. [Google Scholar] [CrossRef]
- Kaur, S.; Suseela, V. Unraveling arbuscular mycorrhiza-induced changes in plant primary and secondary metabolome. Metabolites 2020, 10, 335. [Google Scholar] [CrossRef] [PubMed]
- Baslam, M.; Garmendia, I.; Goicoechea, N. Arbuscular mycorrhizal fungi (AMF) improved growth and nutritional quality of greenhouse-grown lettuce. J. Agric. Food Chem. 2011, 59, 5504–5515. [Google Scholar] [CrossRef] [PubMed]
- Baslam, M.; Goicoechea, N. Water deficit improved the capacity of arbuscular mycorrhizal fungi (AMF) for inducing the accumulation of antioxidant compounds in lettuce leaves. Mycorrhiza 2012, 22, 347–359. [Google Scholar] [CrossRef]
- Avio, L.; Sbrana, C.; Giovannetti, M.; Frassinetti, S. Arbuscular mycorrhizal fungi affect total phenolics content and antioxidant activity in leaves of oak leaf lettuce varieties. Sci. Hortic. 2017, 224, 265–271. [Google Scholar] [CrossRef]
- Phillips, J.M.; Hayman, D. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Koske, R.E.; Gemma, J.N. A modified procedure for staining roots to detect VA mycorrhizas. Mycol. Res. 1989, 92, 486–505. [Google Scholar] [CrossRef]
- McGonigle, T.; Miller, M.; Evans, D.; Fairchild, G.; Swan, J. A new method which gives an objective measure of colonization of roots by vesicular arbuscular mycorrhizal fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Giovannetti, M.; Mosse, B. An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, J.; Rayson, G.; Tian, B.; Lin, Y. Sensor array for carbohydrates and amino acids based on electrocatalytic modified electrodes. Anal. Chem. 1993, 65, 251–254. [Google Scholar] [CrossRef]
- Benzie, I.E.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of antioxidant power: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smirnoff, N. Ascorbic acid: Metabolism and functions of a multi-facetted molecule. Curr. Opin. Plant Biol. 2000, 3, 229–235. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. [14] Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Chang, C.C.; Yang, M.H.; Wen, H.M.; Chern, J.C. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug. Anal. 2002, 10, 178–182. [Google Scholar]
- Moradi, S.; Pasari, B.; Talebi, R. Study of the effects of mycorrhiza, fulvic acid, seaweed extract and urea on physiological traits and leaf yield of tobacco (Burley 21). Eur. J. Environ. Sci. 2019, 9, 33–40. [Google Scholar] [CrossRef]
- Kowalska, I.; Konieczny, A.; Gastol, M. Effect of mycorrhiza and the phosphorus content in a nutrient solution on the yield and nutritional status of lettuce grown on various substrates. J. Elem. 2015, 20, 631–642. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Summers, W.; Paszkowski, U. Mechanisms underlying establishment of arbuscular mycorrhizal symbioses. Annu. Rev. Phytopathol. 2018, 56, 135–160. [Google Scholar] [CrossRef]
- Dudaš, S.; Šola, I.; Sladonja, B.; Erhatić, R.; Ban, D.; Poljuha, D. The effect of biostimulant and fertilizer on “low input” lettuce production. Acta Bot. Croat. 2016, 75, 253–259. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; De Micco, V.; Arena, C.; Raimondi, G.; Colla, G.; De Pascale, S. Effect of Ecklonia maxima seaweed extract on yield, mineral composition, gas exchange, and leaf anatomy of zucchini squash grown under saline conditions. J. Appl. Phycol. 2017, 29, 459–470. [Google Scholar] [CrossRef]
- Kinany, S.E.; Achbani, E.; Faggroud, M.; Ouahmane, L.; Hilali, R.E.; Haggoud, A.; Bouamri, R. Effect of organic fertilizer and commercial arbuscular mycorrhizal fungi on the growth of micropropagated date palm cv. Feggouss. J. Saudi Soc. Agric. Sci. 2019, 18, 411–417. [Google Scholar] [CrossRef]
- Zhang, F.; Liu, M.; Li, Y.; Che, Y.; Xiao, Y. Effects of arbuscular mycorrhizal fungi, biochar and cadmium on the yield and element uptake of Medicago sativa. Sci. Total Environ. 2019, 655, 1150–1158. [Google Scholar] [CrossRef] [PubMed]
- Shukla, P.S.; Mantin, E.G.; Adil, M.; Bajpai, S.; Critchley, A.T.; Prithiviraj, B. Ascophyllum nodosum-based biostimulants: Sustainable applications in agriculture for the stimulation of plant growth, stress tolerance, and disease management. Front. Plant Sci. 2019, 10, 655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colla, G.; Cardarelli, M.; Bonini, P.; Rouphael, Y. Foliar applications of protein hydrolysate, plant and seaweed extracts increase yield but differentially modulate fruit quality of greenhouse tomato. HortScience 2017, 52, 1214–1220. [Google Scholar] [CrossRef]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural uses of plant biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef] [Green Version]
- Sosnowski, J.; Jankowski, K.; Wiśniewska-Kadżajan, B.; Malinowska, E.; Kolczarek, R.; Czeluściński, W.; Radzka, E. Shaping of chemical composition and digestibility of Medicago× varia T. Martyn under the influence of Ecklonia maxima seaweed extract. Fresenius. Environ. Bull. 2015, 24, 881–887. [Google Scholar]
- Bao, X.; Wang, Y.; Olsson, P.A. Arbuscular mycorrhiza under water—Carbon–phosphorus exchange between rice and arbuscular mycorrhizal fungi under different flooding regimes. Soil Biol. Biochem. 2019, 129, 169–177. [Google Scholar] [CrossRef]
- González-González, M.F.; Ocampo-Alvarez, H.; Santacruz-Ruvalcaba, F.; Sánchez-Hernández, C.V.; Casarrubias-Castillo, K.; Becerril-Espinosa, A.; Hernández-Herrera, R.M. Physiological, ecological, and biochemical implications in tomato plants of two plant biostimulants: Arbuscular mycorrhizal fungi and seaweed extract. Front. Plant Sci. 2020, 11, 999. [Google Scholar] [CrossRef] [PubMed]
- Paul, J.; Yuvaraj, P. Effect of seaweed liquid fertilizer of Colpomenia sinuosa (Mert. ex Roth) Derbes & Solier (Brown Seaweed) on Vigna radiata (L.) R. Wilczek, Koothankuzhi, Tirunelveli district, Tamil Nadu, India. Int. J. Pure App. Biosci. 2014, 2, 177–184. [Google Scholar]
- Patel, Z.; Bhalerao, P.P.; Gaikwad, S.S.; Sachin, A.J. Effect of foliar application of chemicals on growth and yield of garlic (Allium sativum L.) var. GG-4. Int. J. Chem. Stud. 2017, 5, 1035–1037. [Google Scholar]
- Subramanian, K.S.; Santhanakrishnan, P.; Balasubramanian, P. Responses of field grown tomato plants to arbuscular mycorrhizal fungal colonization under varying intensities of drought stress. Sci. Hortic. 2006, 107, 245–253. [Google Scholar] [CrossRef]
- Tiruvaimozhi, Y.V.; Varma, V.; Sankaran, M. Nitrogen fixation ability explains leaf chemistry and arbuscular mycorrhizal responses to fertilization. Plant Ecol. 2018, 219, 391–401. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Q.Y.; Tan, S.Y.; Zhuo, F.; Yang, D.J.; Ye, Z.H.; Jing, Y.X. Effect of Funneliformis mosseae on the growth, cadmium accumulation and antioxidant activities of Solanum nigrum. Appl. Soil Ecol. 2016, 98, 112–120. [Google Scholar] [CrossRef]
- Avio, L.; Maggini, R.; Ujvári, G.; Incrocci, L.; Giovannetti, M.; Turrini, A. Phenolics content and antioxidant activity in the leaves of two artichoke cultivars are differentially affected by six mycorrhizal symbionts. Sci. Hortic. 2020, 264, 109153. [Google Scholar] [CrossRef]
- Gholinezhad, E.; Darvishzadeh, R.; Moghaddam, S.S.; Popović-Djordjević, J. Effect of mycorrhizal inoculation in reducing water stress in sesame (Sesamum indicum L.): The assessment of agrobiochemical traits and enzymatic antioxidant activity. Agric. Water Manag. 2020, 238, 106234. [Google Scholar] [CrossRef]
- Fan, D.; Hodges, D.M.; Zhang, J.; Kirby, C.W.; Ji, X.; Locke, S.J.; Prithiviraj, B. Commercial extract of the brown seaweed Ascophyllum nodosum enhances phenolic antioxidant content of spinach (Spinacia oleracea L.) which protects Caenorhabditis elegans against oxidative and thermal stress. Food Chem. 2012, 124, 195–202. [Google Scholar] [CrossRef] [Green Version]
- De Sousa Oliveira, M.; da Silva Campos, M.A.; de Albuquerque, U.P.; da Silva, F.S.B. Arbuscular mycorrhizal fungi (AMF) affect biomolecules content in Myracrodruon urundeuva seedlings. Ind. Crops Prod. 2013, 50, 244–247. [Google Scholar] [CrossRef]
- Basavaraja, P.K.; Yogendra, N.D.; Zodape, S.T.; Prakash, R.; Ghosh, A. Effect of seaweed sap as foliar spray on growth and yield of hybrid maize. J. Plant Nutr. 2018, 41, 1851–1861. [Google Scholar] [CrossRef]
- Ashour, M.; Hassan, S.M.; Elshobary, M.E.; Ammar, G.A.; Gaber, A.; Alsanie, W.F.; El-Shenody, R. Impact of commercial seaweed liquid extract (TAM®) biostimulant and its bioactive molecules on Growth and Antioxidant Activities of Hot Pepper (Capsicum annuum). Plants 2021, 10, 1045. [Google Scholar] [CrossRef] [PubMed]
- de Assis, R.M.A.; Carneiro, J.J.; Medeiros, A.P.R.; de Carvalho, A.A.; da Cunha Honorato, A.; Carneiro, M.A.C.; Bertolucci, S.K.V.; Pinto, J.E.B.P. Arbuscular mycorrhizal fungi and organic manure enhance growth and accumulation of citral, total phenols, and flavonoids in Melissa officinalis L. Ind. Crops Prod. 2020, 158, 112981. [Google Scholar] [CrossRef]
- Hassan, S.M.; Ashour, M.; Soliman, A.A.; Hassanien, H.A.; Alsanie, W.F.; Gaber, A.; Elshobary, M.E. The potential of a new commercial seaweed extract in stimulating morpho-agronomic and bioactive properties of Eruca vesicaria (L.) Cav. Sustainability 2021, 13, 4485. [Google Scholar] [CrossRef]
- Mahmoud, S.H.; Salama, D.M.; El-Tanahy, A.M.; Abd El-Samad, E.H. Utilization of seaweed (Sargassum vulgare) extract to enhance growth, yield and nutritional quality of red radish plants. Ann. Agric. Sci. 2019, 64, 167–175. [Google Scholar] [CrossRef]
- Hao, Z.; Xie, W.; Chen, B. arbuscular mycorrhizal symbiosis affects plant immunity to viral infection and accumulation. Viruses 2019, 11, 534. [Google Scholar] [CrossRef] [Green Version]
- Berger, F.; Gutjahr, C. Factors affecting plant responsiveness to arbuscular mycorrhiza. Curr. Opin. Plant Biol. 2021, 59, 101994. [Google Scholar] [CrossRef]
- Talaat, N.B.; Shawky, B.T. Protective effects of arbuscular mycorrhizal fungi on wheat (Triticum aestivum L.) plants exposed to salinity. Environ. Exp. Bot. 2014, 98, 20–31. [Google Scholar] [CrossRef]
- Jannoura, R.; Joergensen, R.G.; Bruns, C. Organic fertilizer effects on growth, crop yield, and soil microbial biomass indices in sole and intercropped peas and oats under organic farming conditions. Eur. J. Agron. 2014, 52, 259–270. [Google Scholar] [CrossRef]
- Tarkka, M.T.; Frey-Klett, P. Mycorrhiza Helper Bacteria. In Mycorrhiza; Springer: Berlin/Heidelberg, Germany, 2008; pp. 113–132. [Google Scholar]
- Atayese, M.O. Field response of groundnut (Arachis hypogea L.) cultivars to mycorrhizal inoculation and phosphorus fertilizer in Abeokuta, South West Nigeria. Am.-Eurasian J. Agric. Tur. Environ. Sci. 2007, 2, 16–23. [Google Scholar]
- Boomsma, C.R.; Vyn, T.J. Maize drought tolerance: Potential improvements through arbuscular mycorrhizal symbiosis. Field Crops Res. 2008, 108, 14–31. [Google Scholar] [CrossRef]
- Urcoviche, R.C.; Gazim, Z.C.; Dragunski, D.C.; Barcellos, F.G.; Alberton, O. Plant growth and essential oil content of Mentha crispa inoculated with arbuscular mycorrhizal fungi under different levels of phosphorus. Ind. Crops Prod. 2015, 67, 103–107. [Google Scholar] [CrossRef]
- Kiran, S.; Kuşvuran, Ş.; Özkay, F.; Ellialtioğlu, Ş.Ş. Change in physiological and biochemical parameters under drought stress in salt-tolerant and salt-susceptible eggplant genotypes. Turk. J. Agric. For. 2019, 43, 593–602. [Google Scholar] [CrossRef]
- Lehmann, A.; Veresoglou, S.D.; Leifheit, E.F.; Rillig, M.C. Arbuscular mycorrhizal influence on zinc nutrition in crop plants—A meta-analysis. Soil Biol. Biochem. 2014, 69, 123–131. [Google Scholar] [CrossRef]
- Subaşi, İ. Seed fatty acid compositions and chemotaxonomy of wild Crambe (Brassicaceae) taxa in Turkey. Turk. J. Agric. For. 2020, 44, 662–670. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fertilization | Number of Leaves | Head Diameter (cm) | Head Fresh Weight (g) | Root Fresh Weight (g) | Head Dry Weight (g) | Root Dry Weight (g) |
|---|---|---|---|---|---|---|
| control | 25.5 ± 3.01d | 20.75 ± 3.17g | 72.5 ± 20.6f | 22.38 ± 2.74g | 1.42 ± 0.54e | 13.88 ± 1.45g |
| SWE1 | 28.5 ± 2.96c | 29.25 ± 2.39f | 134.2 ± 18.90e | 33.03 ± 1.26df | 2.02 ± 0.035bc | 20.9 ± 0.57f |
| SWE2 | 30.47 ± 2.75c | 34.87 ± 2.57e | 173 ± 27.1d | 32.25 ± 1.572f | 2.2 ± 0.035d | 25.88 ± 0.32d |
| SWE3 | 34.75 ± 1.62bc | 57.53 ± 3.54c | 220.5 ± 14.4c | 46.49 ± 2.275c | 3.02 ± 0.022bc | 30.01 ± 0.42c |
| AMF | 31.5 ± 1.83bc | 32.1 ± 2.39ef | 180.2 ± 22.5d | 35.6 ± 6.95ef | 2.57 ± 0.041cd | 23.4 ± 0.17e |
| AMF + SWE1 | 36 ± 2.61ab | 43.72 ± 1.95d | 199.1 ± 24.2de | 42.22 ± 2.351cd | 2.37 ± 0.056c | 28.38 ± 0.43c |
| AMF + SWE2 | 37 ± 2.21a | 66.48 ± 3.53b | 245.7 ± 18.6b | 52.11 ± 1.16b | 3.4 ± 0.127b | 32.86 ± 0.34b |
| AMF + SWE3 | 36.72 ± 3.83a | 76.02 ± 3.13a | 284.3 ± 17.01a | 63.83 ± 3.31a | 5.2 ± 0.437a | 39.7 ± 2.08a |
| LSD at 0.05% | 8.17 | 4.49 | 24.32 | 4.82 | 0.587 | 2.16 |
| Significance | ||||||
| AMF | ns | ** | ** | ** | ** | ** |
| SWE | ns | ** | ** | ** | ** | ** |
| AMF × SWE | ** | ** | * | * | ** | ** |
| S.O.V. | df | Total Protein Content (mg g−1 FW) | Carbohydrate Content (mg g−1 FW) | TAA (%) | Ascorbic acid Content (mg 100g−1 FW) | Total Phenolics Content (mg 100g−1 FW) | Total Flavonoids Content (mg g−1 FW) |
|---|---|---|---|---|---|---|---|
| AMF | 1 | 0.146 ** | 1708.3 ** | 392.91 ** | 262.43 ** | 7699.90 ** | 14,238.2 ** |
| SWE | 3 | 0.792 ** | 8346.9 ** | 663.45 ** | 961.32 ** | 19,875.95 ** | 48,877.1 ** |
| AMF × SWE | 3 | 0.007 ** | 103.2 * | 37.60 * | 125.95 ** | 3326.05 ** | 1733.1 ** |
| Error | 24 | 0.002 | 32.06 | 12.60 | 10.08 | 593.75 | 264.94 |
| C.V. | 5.51 | 6.15 | 13.05 | 5.92 | 12.55 | 8.82 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rasouli, F.; Amini, T.; Asadi, M.; Hassanpouraghdam, M.B.; Aazami, M.A.; Ercisli, S.; Skrovankova, S.; Mlcek, J. Growth and Antioxidant Responses of Lettuce (Lactuca sativa L.) to Arbuscular Mycorrhiza Inoculation and Seaweed Extract Foliar Application. Agronomy 2022, 12, 401. https://doi.org/10.3390/agronomy12020401
Rasouli F, Amini T, Asadi M, Hassanpouraghdam MB, Aazami MA, Ercisli S, Skrovankova S, Mlcek J. Growth and Antioxidant Responses of Lettuce (Lactuca sativa L.) to Arbuscular Mycorrhiza Inoculation and Seaweed Extract Foliar Application. Agronomy. 2022; 12(2):401. https://doi.org/10.3390/agronomy12020401
Chicago/Turabian StyleRasouli, Farzad, Trifa Amini, Mohammad Asadi, Mohammad Bagher Hassanpouraghdam, Mohammad Ali Aazami, Sezai Ercisli, Sona Skrovankova, and Jiri Mlcek. 2022. "Growth and Antioxidant Responses of Lettuce (Lactuca sativa L.) to Arbuscular Mycorrhiza Inoculation and Seaweed Extract Foliar Application" Agronomy 12, no. 2: 401. https://doi.org/10.3390/agronomy12020401