
Silicon Application Induced Alleviation of Aluminum Toxicity in Xaraés Palisadegrass


 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design and Treatments
2.3. Xaraés Palisadegrass Growth and Harvest
2.4. Evaluations
2.4.1. Number of Leaves and Tillers, Shoot Dry Mass, and Fibers Content
2.4.2. Silicon and Al Concentrations in Leaf and Root Tissue and Nutrient Concentration in Diagnostic Leaves (Nutritional Status)
2.4.3. Silicon and Al Roots to Shoots Translocation Factor
2.5. Statistical Analysis
2.6. Principal Components Analysis
3. Results
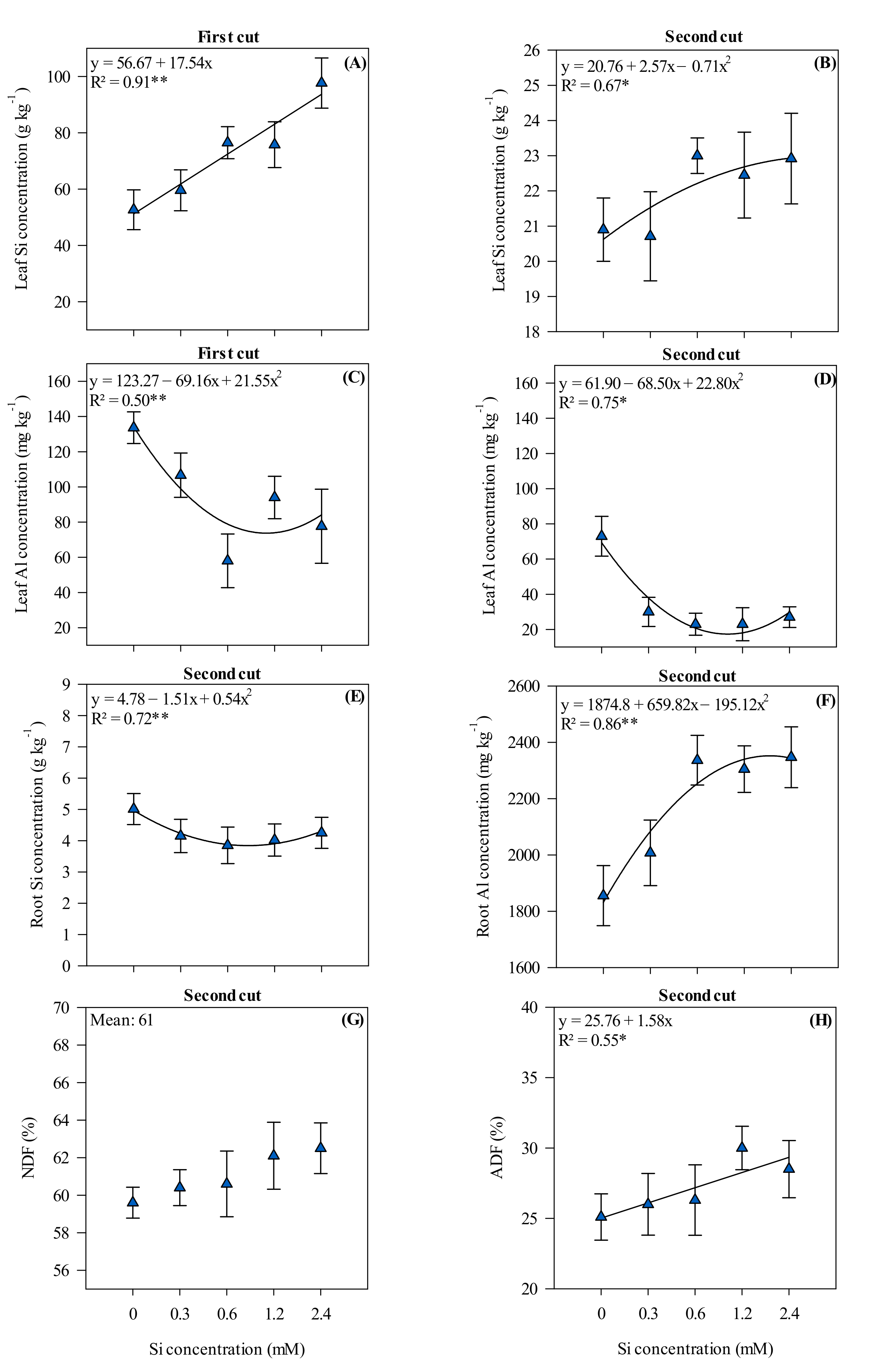
3.1. Silicon and Aluminum Content in Leaves and Roots, Neutral, and Acid Detergent Fiber Content in Xaraés Palisadegrass
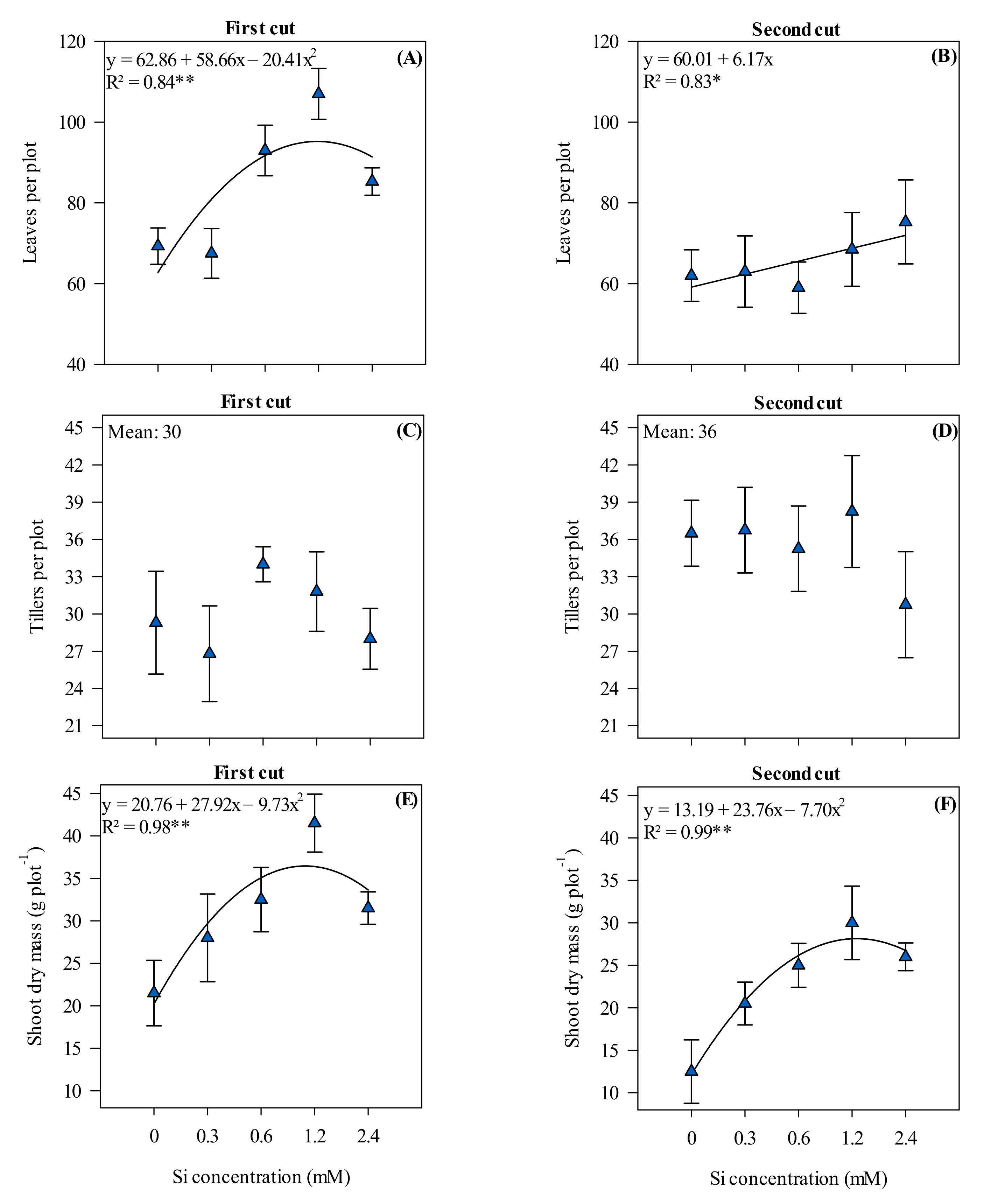
3.2. Number of Leaves and Tillers Per Plot and Shoot Dry Mass of Xaraés Palisadegrass
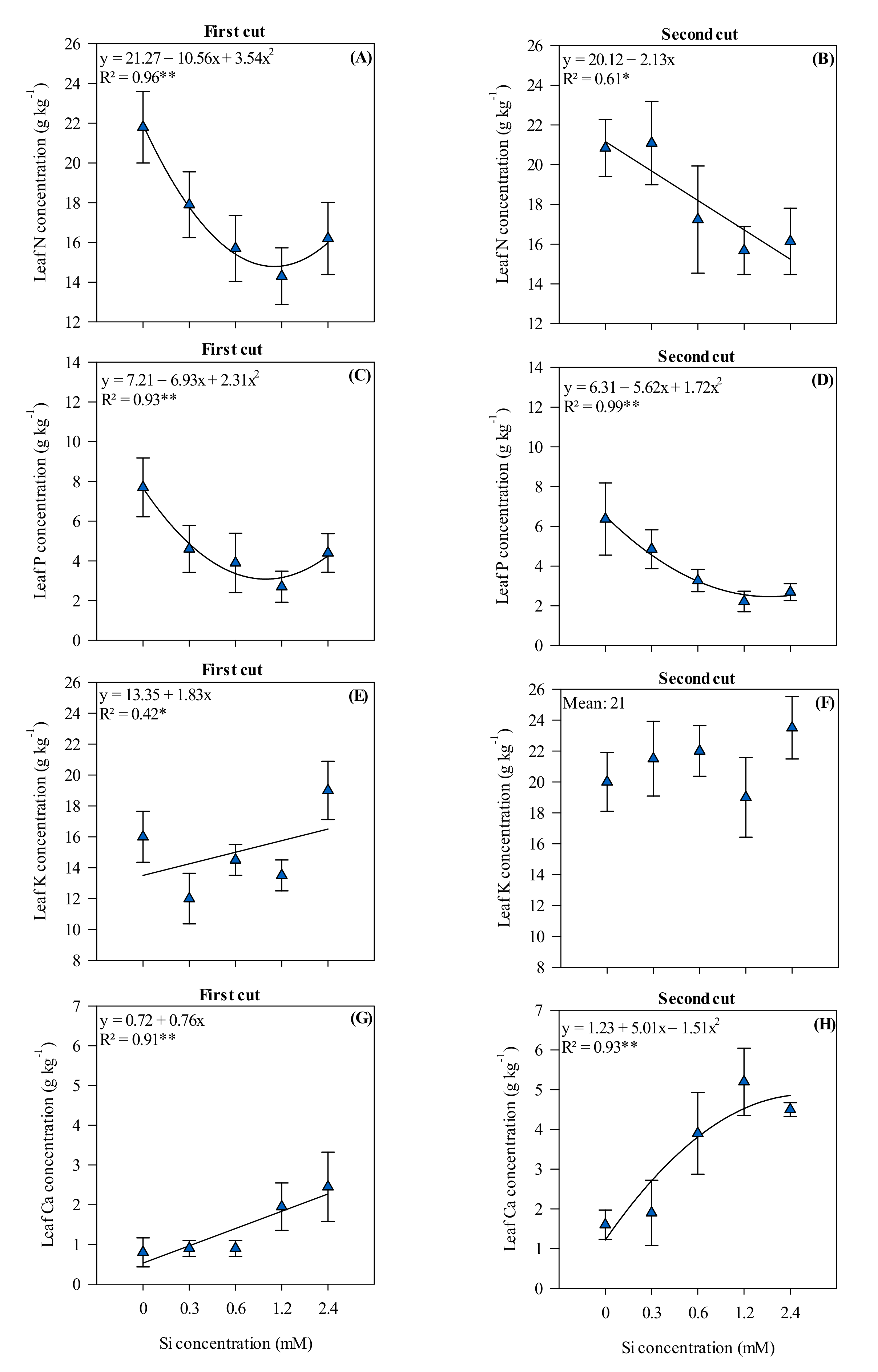
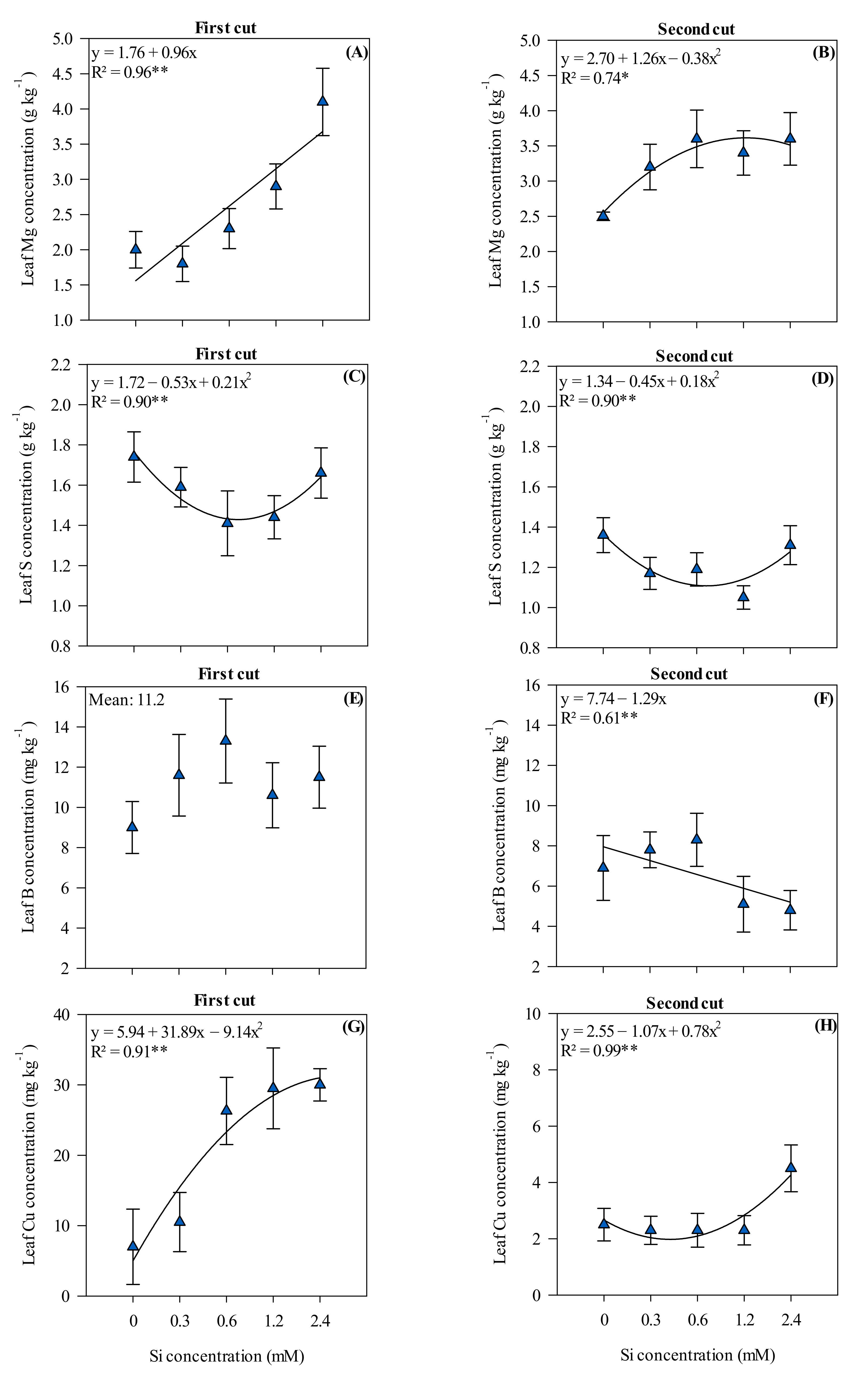
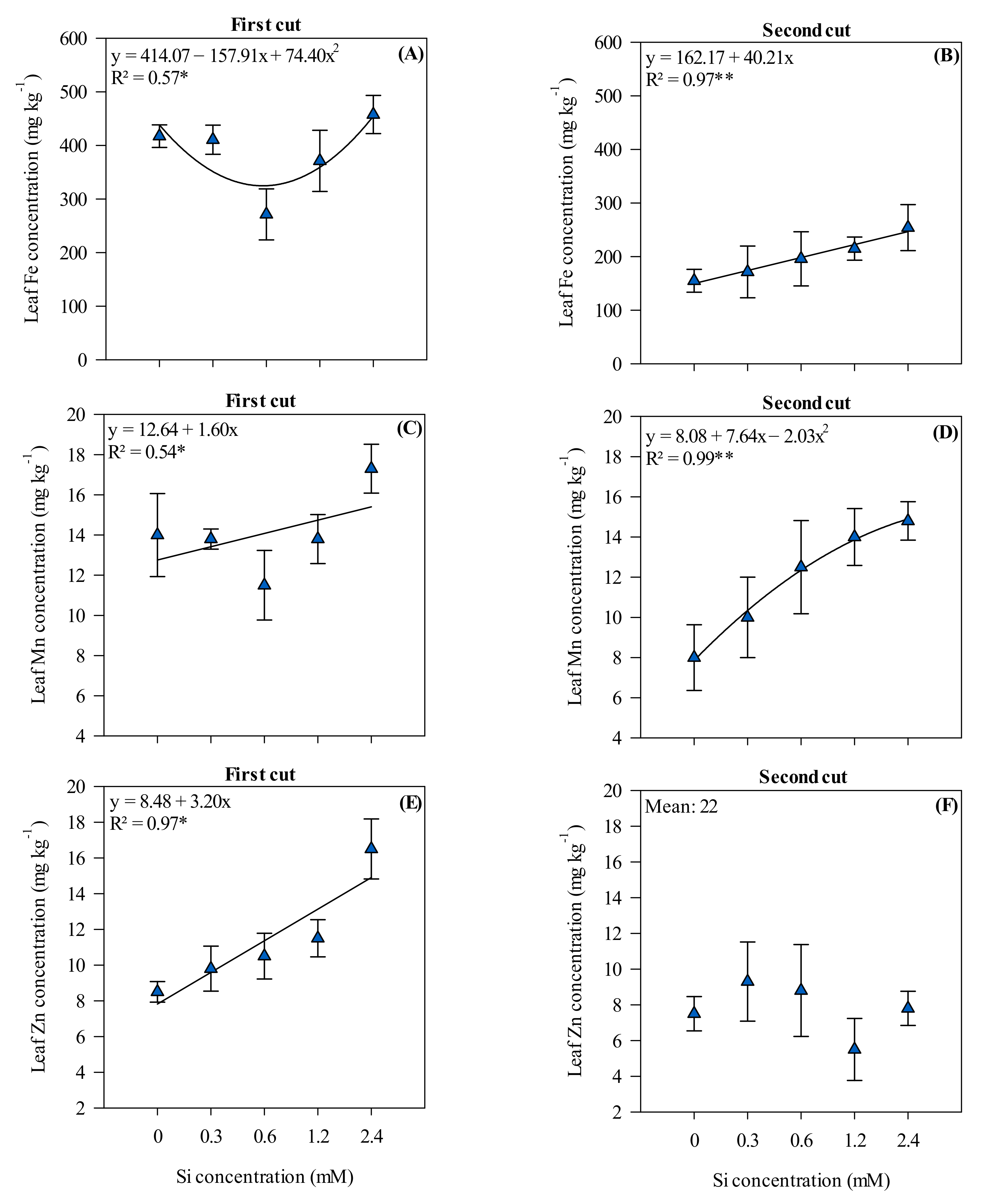
3.3. Leaf Nutrient Concentrations in Xaraés Palisadegrass
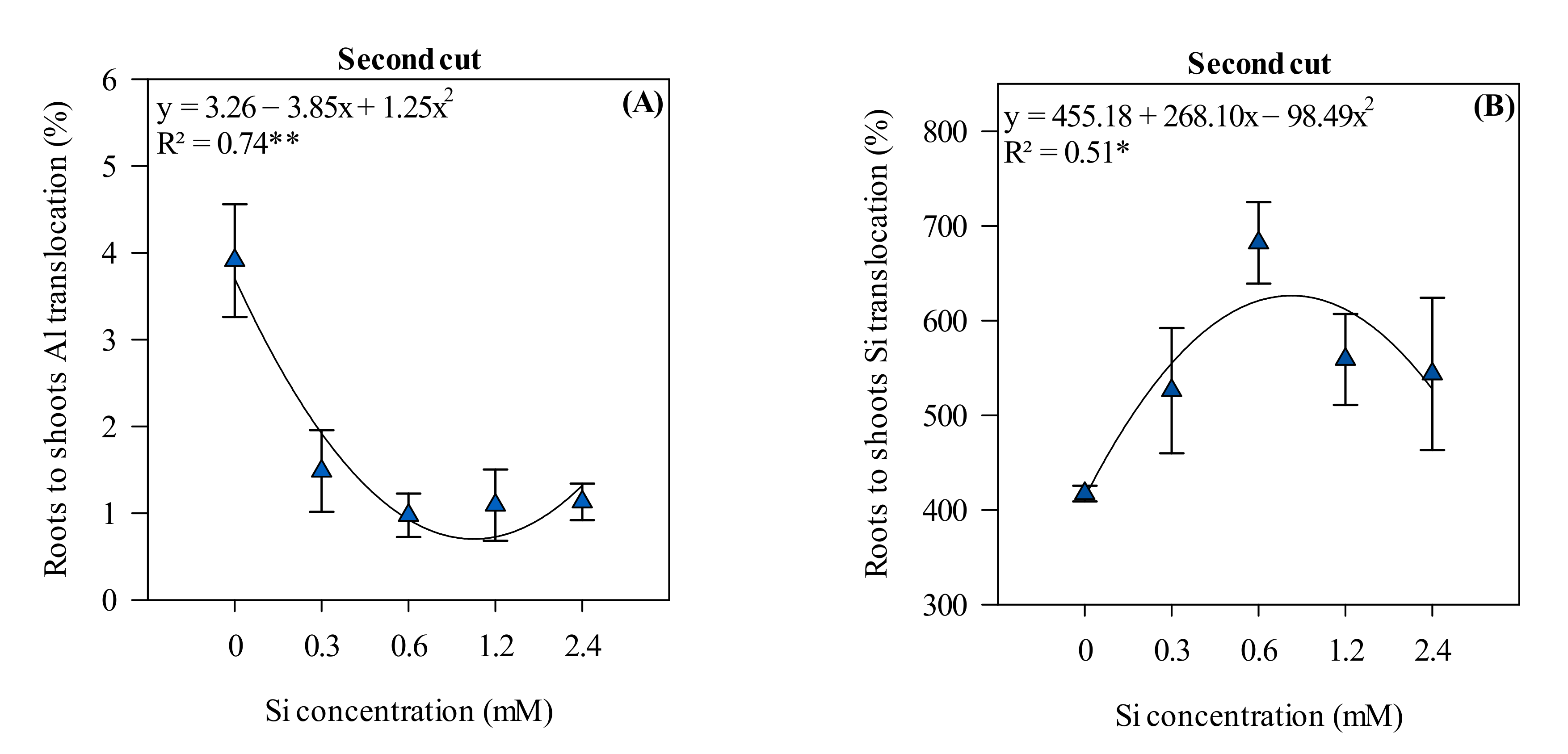
3.4. Roots to Shoots Al and Si Translocation
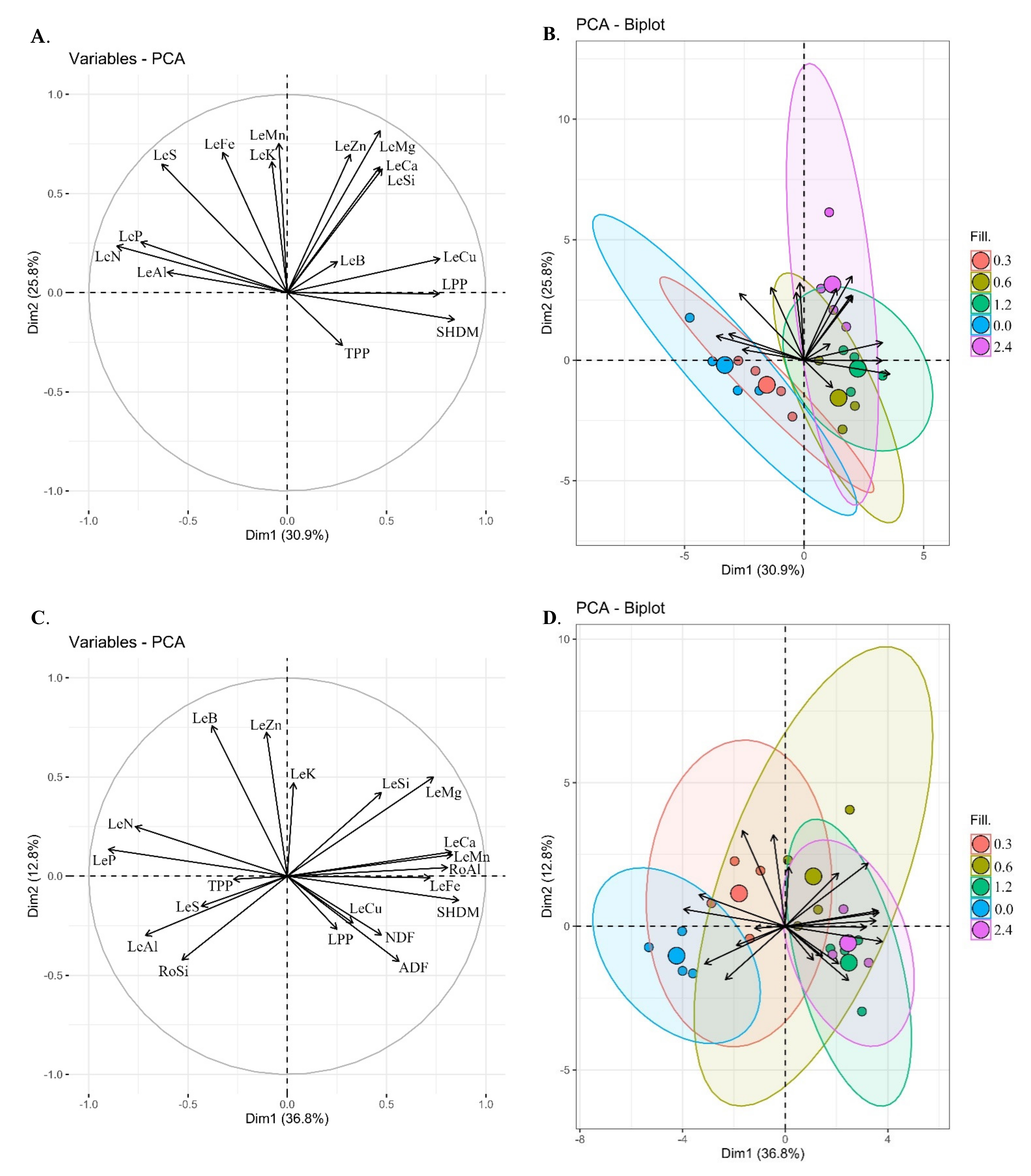
3.5. Principal Component Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Galindo, F.S.; Buzetti, S.; Teixeira Filho, M.C.M.; Dupas, E. Rates and sources of nitrogen fertilizer application on yield and quality of Panicum maximum cv. Mombasa. Idesia 2019, 37, 67–73. [Google Scholar] [CrossRef]
- Fernandes, J.C.; Buzetti, S.; Dupas, E.; Teixeira Filho, M.C.M.; Andreotti, M. Sources and rates of nitrogen fertilizer used in Mombasa guineagrass in the Brazilian Cerrado region. Afr. J. Agric. Res. 2015, 10, 2076–2082. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, P.P.A.; Corte, R.; Silva, S.; Rodriguez, P.; Sakamoto, L.; Pedroso, A.; Tullio, R.; Berndt, A. The effect of grazing system intensification on the growth and meat quality of beef cattle in the Brazilian Atlantic Forest biome. Meat Sci. 2018, 139, 157–161. [Google Scholar] [CrossRef]
- Costa, C.M.; da Costa, A.B.G.; de Farias Theodoro, G.; dos Santos, G.; Difante, A.L.C.G.; Santana, J.C.S.; Camargo, F.C.; de Almeida, E.M. The 4R management for nitrogen fertilization in tropical forage: A review. Aust. J. Crop Sci. 2020, 14, 1834–1837. [Google Scholar] [CrossRef]
- Galindo, F.S.; Beloni, T.; Buzetti, S.; Filho, M.C.M.T.; Dupas, E.; Ludkiewicz, M.G.Z. Technical and economic viability and nutritional quality of Mombasa guinea grass silage production. Acta Sci. Agron. 2018, 40, 36395. [Google Scholar] [CrossRef]
- Pontigo, S.; Godoy, K.; Jiménez, H.; Gutiérrez-Moraga, A.; Mora, M.L.; Cartes, P. Silicon-mediated alleviation of aluminum toxicity by modulation of Al/Si uptake and antioxidant performance in ryegrass plants. Front. Plant Sci. 2017, 8, 642. [Google Scholar] [CrossRef] [Green Version]
- De Sousa, A.; Saleh, A.M.; Habeeb, T.H.; Hassan, Y.M.; Zrieq, R.; Wadaan, M.A.; Hozzein, W.N.; Selim, S.; Matos, M.; AbdElgawad, H. Silicon dioxide nanoparticles ameliorate the phytotoxic hazards of aluminum in maize grown on acidic soil. Sci. Total Environ. 2019, 693, 133636. [Google Scholar] [CrossRef] [PubMed]
- Worthington, M.; Perez, J.G.; Mussurova, S.; Silva-Cordoba, A.; Castiblanco, V.; Arango, J.A.C.; Jones, C.; Fernandez-Fuentes, N.; Skot, L.; Dyer, S.; et al. A new genome allows the identification of genes associated with natural variation in aluminium tolerance in Brachiaria grasses. J. Exp. Bot. 2021, 72, 302–319. [Google Scholar] [CrossRef] [PubMed]
- De Jesus, L.R.; Batista, B.L.; da Silva Lobato, A.K. Silicon reduces aluminum accumulation and mitigates toxic effects in cowpea plants. Acta Physiol. Plant. 2017, 39, 138. [Google Scholar] [CrossRef]
- De Freitas, L.B.; Fernandes, D.M.; Maia, S.C.M.; Fernandes, A.M. Effects of silicon on aluminum toxicity in upland rice plants. Plant Soil 2017, 420, 263–275. [Google Scholar] [CrossRef]
- Singh, S.; Tripathi, D.K.; Singh, S.; Sharma, S.; Dubey, N.; Chauhan, D.; Vaculik, M. Toxicity of aluminium on various levels of plant cells and organism: A review. Environ. Exp. Bot. 2017, 137, 177–193. [Google Scholar] [CrossRef]
- Horst, W.J.; Wang, Y.; Eticha, D. The role of the root apoplast in aluminium-induced inhibition of root elongation and in aluminium resistance of plants: A review. Ann. Bot. 2010, 106, 185–197. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Kobayashi, Y.; Matsumoto, H. Lipid peroxidation is an early symptom triggered by aluminum, but not the primary cause of elongation inhibition in pea roots. Plant Physiol. 2001, 125, 199–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, N.; Gaurav, S.S.; Kumar, A. Molecular basis of aluminium toxicity in plants: A review. Am. J. Plant Sci. 2013, 4, 21–37. [Google Scholar] [CrossRef] [Green Version]
- Goodwin, S.B.; Sutter, T.R. Microarray analysis of Arabidopsis genome response to aluminum stress. Biol. Plant. 2009, 53, 85–99. [Google Scholar] [CrossRef]
- Vega, I.; Nikolic, M.; Pontigo, S.; Godoy, K.; Mora, M.L.; Cartes, P. Silicon improves the production of high antioxidant or structural phenolic compounds in barley cultivars under aluminum stress. Agronomy 2019, 9, 388. [Google Scholar] [CrossRef] [Green Version]
- Luyckx, M.; Hausman, J.-F.; Lutts, S.; Guerriero, G. Silicon and plants: Current knowledge and technological perspectives. Front. Plant Sci. 2017, 8, 411. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Gao, L.; Dong, S.; Sun, Y.; Shen, Q.; Guo, S. Role of silicon on plant–pathogen interactions. Front. Plant Sci. 2017, 8, 701. [Google Scholar] [CrossRef] [Green Version]
- Haynes, R.J. What effect does liming have on silicon availability in agricultural soils? Geoderma 2019, 337, 375–383. [Google Scholar] [CrossRef]
- Galindo, F.S.; Buzetti, S.; Rodrigues, W.L.; Boleta, E.; Silva, V.M.; Tavanti, R.F.R.; Fernandes, G.C.; Biagini, A.L.C.; Rosa, P.A.L.; Filho, M.C.M.T. Inoculation of Azospirillum brasilense associated with silicon as a liming source to improve nitrogen fertilization in wheat crops. Sci. Rep. 2020, 10, 1–18. [Google Scholar] [CrossRef]
- Galindo, F.S.; Pagliari, P.H.; Buzetti, S.; Rodrigues, W.L.; Santini, J.M.K.; Boleta, E.; Rosa, P.A.L.; Nogueira, T.A.R.; Lazarini, E.; Filho, M.C.M.T. Can silicon applied to correct soil acidity in combination with Azospirillum brasilense inoculation improve nitrogen use efficiency in maize? PLoS ONE 2020, 15, e0230954. [Google Scholar] [CrossRef]
- Cuong, T.X.; Ullah, H.; Datta, A.; Hanh, T.C. Effects of silicon-based fertilizer on growth, yield and nutrient uptake of rice in tropical zone of Vietnam. Rice Sci. 2017, 24, 283–290. [Google Scholar] [CrossRef]
- Keeping, M.G.; Miles, N.; Rutherford, R.S. Liming an acid soil treated with diverse silicon sources: Effects on silicon uptake by sugarcane (Saccharum spp. hybrids). J. Plant Nutr. 2017, 40, 1417–1436. [Google Scholar] [CrossRef]
- Schaller, J.; Turner, B.; Weissflog, A.; Pino, D.; Bielnicka, A.W.; Engelbrecht, B.M.J. Silicon in tropical forests: Large variation across soils and leaves suggests ecological significance. Biogeochemistry 2018, 140, 161–174. [Google Scholar] [CrossRef]
- Caubet, M.; Cornu, S.; Saby, N.P.A.; Meunier, J.-D. Agriculture increases the bioavailability of silicon, a beneficial element for crop, in temperate soils. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Pedreira, C.; Silva, V.; Pedreira, B.; Sollenberger, L.E. Herbage accumulation and organic reserves of palisadegrass in response to grazing management based on canopy targets. Crop Sci. 2017, 57, 2283–2293. [Google Scholar] [CrossRef]
- Vandevenne, F.; Struyf, E.; Clymans, W.; Meire, P. Agricultural silica harvest: Have humans created a new loop in the global silica cycle? Front. Ecol. Environ. 2012, 10, 243–248. [Google Scholar] [CrossRef]
- Schaller, J.; Puppe, D.; Kaczorek, D.; Ellerbrock, R.; Sommer, M. Silicon cycling in soils revisited. Plants 2021, 10, 295. [Google Scholar] [CrossRef]
- Guntzer, F.; Keller, C.; Meunier, J.-D. Benefits of plant silicon for crops: A review. Agron. Sustain. Dev. 2012, 32, 201–213. [Google Scholar] [CrossRef] [Green Version]
- Bakhat, H.F.; Bibi, N.; Zia, Z.; Abbas, S.; Hammad, H.M.; Fahad, S.; Ashraf, M.R.; Shah, G.M.; Rabbani, F.; Saeed, S. Silicon mitigates biotic stresses in crop plants: A review. Crop Prot. 2018, 104, 21–34. [Google Scholar] [CrossRef]
- Liang, Y.; Nikolic, M.; Belanger, R.R.; Gong, H.; Song, A. Silicon biogeochemistry and bioavailability in soil. In Silicon in Agriculture; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2015; pp. 45–68. [Google Scholar]
- Linden, C.V.; Delvaux, B. The weathering stage of tropical soils affects the soil-plant cycle of silicon, but depending on land use. Geoderma 2019, 351, 209–220. [Google Scholar] [CrossRef]
- Mohanty, S.; Nayak, A.K.; Swain, C.K.; Dhal, B.; Kumar, A.; Tripathi, R.; Shahid, M.; Lal, B.; Gautam, P.; Dash, G.K.; et al. Silicon enhances yield and nitrogen use efficiency of tropical low land rice. Agron. J. 2019, 112, 758–771. [Google Scholar] [CrossRef]
- Kochian, L.V.; Piñeros, M.A.; Liu, J.; Magalhaes, J.V. Plant adaptation to acid soils: The molecular basis for crop aluminum resistance. Annu. Rev. Plant Biol. 2015, 66, 571–598. [Google Scholar] [CrossRef] [PubMed]
- Rao, I.M.; Miles, J.W.; Beebe, S.E.; Horst, W.J. Root adaptations to soils with low fertility and aluminium toxicity. Ann. Bot. 2016, 118, 593–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arroyave, C.; Barceló, J.; Poschenrieder, C.; Tolrà, R. Aluminium-induced changes in root epidermal cell patterning, a distinctive feature of hyperresistance to Al in Brachiaria decumbens. J. Inorg. Biochem. 2011, 105, 1477–1483. [Google Scholar] [CrossRef]
- Wenzl, P.; Patiño, G.M.; Chaves, A.L.; Mayer, J.E.; Rao, I.M. The high level of aluminum resistance in signalgrass is not associated with known mechanisms of external aluminum detoxification in root apices. Plant Physiol. 2001, 125, 1473–1484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Głazowska, S.; Baldwin, L.; Mravec, J.; Bukh, C.; Hansen, T.; Jensen, M.M.; Fangel, J.U.; Willats, W.G.T.; Glasius, M.; Felby, C.; et al. The impact of silicon on cell wall composition and enzymatic saccharification of Brachypodium distachyon. Biotechnol. Biofuels 2018, 11, 171. [Google Scholar] [CrossRef]
- Hussain, S.; Shuxian, L.; Mumtaz, M.; Shafiq, I.; Iqbal, N.; Brestic, M.; Shoaib, M.; Sisi, Q.; Li, W.; Mei, X.; et al. Foliar application of silicon improves stem strength under low light stress by regulating lignin biosynthesis genes in soybean (Glycine max (L.) Merr.). J. Hazard. Mater. 2021, 401, 123256. [Google Scholar] [CrossRef]
- Teixeira, G.C.M.; Prado, R.D.M.; Rocha, A.M.S.; dos Santos, L.C.N.; Sarah, M.M.D.S.; Gratão, P.L.; Fernandes, C. Silicon in pre-sprouted sugarcane seedlings mitigates the effects of water deficit after transplanting. J. Soil Sci. Plant Nutr. 2020, 20, 849–859. [Google Scholar] [CrossRef]
- Arroyave, C.; Tolrà, R.; Thuy, T.; Barceló, J.; Poschenrieder, C. Differential aluminum resistance in Brachiaria species. Environ. Exp. Bot. 2013, 89, 11–18. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil; College of Agriculture: Berkeley, CA, USA, 1950. [Google Scholar]
- Van Soest, P.J. Nutritional Ecology of the Ruminant, 2nd ed.; Cornell University Press: Ithaca, NY, USA, 1994. [Google Scholar] [CrossRef]
- Korndörfer, G.H.; Pereira, H.S.; Nolla, A. Silicon Analysis: Soil, Plant and Fertilizer; Boletim Técnico: Uberlândia, Brazil, 2004. (In Portuguese) [Google Scholar]
- Coskun, D.; Britto, D.T.; Huynh, W.Q.; Kronzucker, H.J. The role of silicon in higher plants under salinity and drought stress. Front. Plant Sci. 2016, 7, 1072. [Google Scholar] [CrossRef] [Green Version]
- Malavolta, E.; Vitti, G.C.; Oliveira, S.A. Evaluation of the Nutritional Status of Plants: Principles and Applications, 2nd ed.; Potafos: Piracicaba, Brazil, 1997; p. 319. (In Portugese) [Google Scholar]
- Abichequer, A.D.; Bohnen, H. Eficiência de absorção, translocação e utilização de fósforo por variedades de trigo. Rev. Bras. Ciência Solo 1998, 22, 21–26. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 15 September 2020).
- Lawley, D.N.; Maxwell, A.E. Factor analysis as a statistical method. J. R. Stat. Soc. Ser. D Stat. 1962, 12, 209. [Google Scholar] [CrossRef]
- Singh, V.P.; Tripathi, D.K.; Kumar, D.; Chauhan, D.K.; Singh, D.P. Influence of exogenous silicon addition on aluminium tolerance in rice seedlings. Biol. Trace Elem. Res. 2011, 144, 1260–1274. [Google Scholar] [CrossRef]
- Shen, X.; Xiao, X.; Dong, Z.; Chen, Y. Silicon effects on antioxidative enzymes and lipid peroxidation in leaves and roots of peanut under aluminum stress. Acta Physiol. Plant. 2014, 36, 3063–3069. [Google Scholar] [CrossRef]
- Jaiswal, S.K.; Naamala, J.; Dakora, F.D. Nature and mechanisms of aluminium toxicity, tolerance and amelioration in symbiotic legumes and rhizobia. Biol. Fertil. Soils 2018, 54, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Bhat, J.A.; Shivaraj, S.M.; Singh, P.; Navadagi, D.B.; Tripathi, D.K.; Dash, P.K.; Solanke, A.U.; Sonah, H.; Deshmukh, R. Role of silicon in mitigation of heavy metal stresses in crop plants. Plants 2019, 8, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodson, M.J.; Evans, D.E. Aluminium–silicon interactions in higher plants: An update. J. Exp. Bot. 2020, 71, 6719–6729. [Google Scholar] [CrossRef] [PubMed]
- Kopittke, P.M.; Gianoncelli, A.; Kourousias, G.; Green, K.; McKenna, B.A. Alleviation of al toxicity by Si is associated with the formation of Al–Si complexes in root tissues of sorghum. Front. Plant Sci. 2017, 8, 2189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cárcamo-Fincheira, P.; Reyes-Díaz, M.; Omena-García, R.P.; Vargas, J.R.; Alvear, M.; Florez-Sarasa, I.; Rosado-Saouza, L.; Rengel, Z.; Fernie, A.R.; Nunes-Nesi, A.; et al. Metabolomic analyses of highbush blueberry (Vaccinium corymbosum L.) cultivars revealed mechanisms of resistance to aluminum toxicity. Environ. Exp. Bot. 2021, 183, 104338. [Google Scholar] [CrossRef]
- Haynes, R.J. Significance and role of Si in crop production. In Advances in Agronomy; Elsevier BV: Amsterdam, The Netherlands, 2017; Volume 146, pp. 83–166. [Google Scholar]
- Ma, J.F.; Tamai, K.; Ichii, M.; Wu, G.F. A rice mutant defective in Si uptake. Plant Physiol. 2002, 130, 2111–2117. [Google Scholar] [CrossRef] [Green Version]
- Hodson, M.; White, P.; Mead, A.; Broadley, M.R. Phylogenetic variation in the silicon composition of plants. Ann. Bot. 2005, 96, 1027–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reboredo, F.; Lidon, F.C.; Pessoa, F.; Duarte, M.P.; Silva, M.J. The uptake of macronutrients by an active silicon accumulator plant growing in two different substrata. Emir. J. Food Agric. 2013, 25, 986–993. [Google Scholar] [CrossRef] [Green Version]
- Werner, J.C.; Paulino, V.T.; Cantarella, H. Forages. In Liming and Fertilization Recommendations for the State of São Paulo; van Raij, B., van Cantarella, H., Quaggio, J.A., Furlani, A.M.C., Eds.; Instituto Agronômico de Campinas: Campinas, Brazil, 1997; p. 285. (In Portuguese) [Google Scholar]
- Neu, S.; Schaller, J.; Dudel, E.G. Silicon availability modifies nutrient use efficiency and content, C:N:P stoichiometry, and productivity of winter wheat (Triticum aestivum L.). Sci. Rep. 2017, 7, 40829. [Google Scholar] [CrossRef]
- Xu, D.; Gao, X.; Gao, T.; Mou, J.; Li, J.; Bu, H.; Zhang, R.; Li, Q. Interactive effects of nitrogen and silicon addition on growth of five common plant species and structure of plant community in alpine meadow. Catena 2018, 169, 80–89. [Google Scholar] [CrossRef]
- Hurtado, A.C.; Chiconato, D.A.; Prado, R.D.M.; Junior, G.D.S.S.; Felisberto, G. Silicon attenuates sodium toxicity by improving nutritional efficiency in sorghum and sunflower plants. Plant Physiol. Biochem. 2019, 142, 224–233. [Google Scholar] [CrossRef]
- Zhu, Y.; Gong, H. Beneficial effects of silicon on salt and drought tolerance in plants. Agron. Sustain. Dev. 2014, 34, 455–472. [Google Scholar] [CrossRef] [Green Version]
- Adrees, M.; Ali, S.; Rizwan, M.; Rehman, M.Z.U.; Ibrahim, M.; Abbas, F.; Farid, M.; Qayyum, M.F.; Irshad, M.K. Mechanisms of silicon-mediated alleviation of heavy metal toxicity in plants: A review. Ecotoxicol. Environ. Saf. 2015, 119, 186–197. [Google Scholar] [CrossRef]
- Crusciol, C.A.; Artigiani, A.C.; Arf, O.; Filho, A.C.C.; Soratto, R.; Nascente, A.S.; Alvarez, R.C.F. Soil fertility, plant nutrition, and grain yield of upland rice affected by surface application of lime, silicate, and phosphogypsum in a tropical no-till system. Catena 2016, 137, 87–99. [Google Scholar] [CrossRef] [Green Version]
- Crusciol, C.A.C.; de Arruda, D.P.; Fernandes, A.M.; Antonangelo, J.; Alleoni, L.R.F.; do Nascimento, C.A.C.; Rossato, O.B.; McCray, J.M. Methods and extractants to evaluate silicon availability for sugarcane. Sci. Rep. 2018, 8, 916. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Fang, X.; Zhang, R.; Gao, T.; Bu, H.; Du, G. Influences of nitrogen, phosphorus and silicon addition on plant productivity and species richness in an alpine meadow. AoB Plants 2015, 7, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Camargo, M.S.; Korndörfer, G.H.; Wyler, P. Silicate fertilization of sugarcane cultivated in tropical soils. Field Crops Res. 2014, 167, 64–75. [Google Scholar] [CrossRef]
- Camargo, M.S.; Korndörfer, G.H.; Foltran, D.E. Silicon absorption and stalk borer incidence by sugarcane varieties in two ratoons. Biosci. J. 2014, 30, 1304–1313. (In Portuguese) [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Si (mM) | 0 | 0.3 | 0.6 | 1.2 | 2.4 |
|---|---|---|---|---|---|
| Stock solution | Volume (mL L−1) | ||||
| NaSiO3 (0.5 M) | - | 0.6 | 1.2 | 2.4 | 4.8 |
| KH2PO4 (1 M) | 1 | 1 | 1 | 1 | 1 |
| KCl (1 M) | 5 | 5 | 5 | 5 | 5 |
| Ca(NO3)2 (1 M) | 5 | 5 | 5 | 5 | 5 |
| MgSO4 (1 M) | 2 | 2 | 2 | 2 | 2 |
| NH4Cl (1 M) | 5 | 5 | 5 | 5 | 5 |
| Micro—Fe * | 1 | 1 | 1 | 1 | 1 |
| Fe-EDTA ** | 1 | 1 | 1 | 1 | 1 |
| AlCl3.6H2O (0.3 M) | 3.3 | 3.3 | 3.3 | 3.3 | 3.3 |
| First Cut | ||||
| Parameters | PC1 | PC2 | PC3 | PC4 |
| Shoot dry mass | 0.38 | −0.07 | −0.26 | 0.16 |
| Leaves per plot | 0.34 | −0.01 | 0.27 | 0.13 |
| Tillers per plot | 0.12 | −0.13 | 0.51 | 0.12 |
| Leaf Al concentration | −0.31 | 0.05 | −0.20 | 0.41 |
| Leaf Si concentration | 0.21 | 0.31 | 0.10 | −0.26 |
| Leaf N concentration | −0.39 | 0.12 | 0.20 | −0.01 |
| Leaf P concentration | −0.34 | 0.13 | 0.34 | −0.15 |
| Leaf K concentration | −0.03 | 0.32 | 0.44 | 0.06 |
| Leaf Ca concentration | 0.21 | 0.31 | 0.14 | 0.17 |
| Leaf Mg concentration | 0.21 | 0.40 | 0.06 | 0.06 |
| Leaf S concentration | −0.28 | 0.32 | −0.04 | −0.05 |
| Leaf B concentration | 0.11 | 0.08 | −0.16 | −0.75 |
| Leaf Cu concentration | 0.35 | 0.08 | −0.01 | 0.15 |
| Leaf Fe concentration | −0.15 | 0.35 | −0.28 | 0.08 |
| Leaf Mn concentration | −0.02 | 0.37 | −0.25 | 0.22 |
| Leaf Zn concentration | 0.14 | 0.34 | −0.09 | −0.11 |
| Variance (%) | 30.9 | 25.8 | 12.95 | 7.53 |
| Cumulative variance (%) | 30.9 | 56.8 | 69.7 | 77.2 |
| Eigenvalues | 4.95 | 4.14 | 2.07 | 1.20 |
| Second cut | ||||
| Parameters | PC1 | PC2 | PC3 | PC4 |
| Shoot dry mass | 0.32 | −0.08 | −0.11 | −0.13 |
| Leaves per plot | 0.09 | −0.17 | 0.43 | −0.30 |
| Tillers per plot | −0.10 | −0.01 | −0.01 | −0.46 |
| Leaf Al concentration | −0.30 | −0.19 | 0.08 | 0.26 |
| Leaf Si concentration | 0.17 | 0.30 | −0.10 | −0.12 |
| Leaf N concentration | −0.31 | 0.16 | 0.09 | −0.29 |
| Leaf P concentration | −0.33 | 0.08 | 0.12 | −0.04 |
| Leaf K concentration | 0.01 | 0.30 | 0.41 | −0.33 |
| Leaf Ca concentration | 0.30 | 0.08 | 0.07 | 0.12 |
| Leaf Mg concentration | 0.27 | 0.31 | 0.16 | 0.11 |
| Leaf S concentration | −0.16 | −0.09 | 0.45 | 0.08 |
| Leaf B concentration | −0.14 | 0.47 | −0.17 | 0.14 |
| Leaf Cu concentration | 0.12 | −0.15 | 0.29 | 0.01 |
| Leaf Fe concentration | 0.26 | −0.01 | 0.33 | 0.21 |
| Leaf Mn concentration | 0.31 | 0.07 | 0.23 | 0.09 |
| Leaf Zn concentration | −0.04 | 0.45 | 0.11 | 0.35 |
| Root Si concentration | −0.20 | −0.26 | 0.19 | 0.40 |
| Root Al concentration | 0.31 | 0.03 | −0.09 | −0.02 |
| Neutral detergent fiber | 0.17 | −0.19 | −0.13 | −0.04 |
| Acid detergent fiber | 0.21 | −0.27 | −0.13 | 0.02 |
| Variance (%) | 36.9 | 12.82 | 10.81 | 9.16 |
| Cumulative variance (%) | 36.9 | 49.7 | 60.5 | 69.6 |
| Eigenvalues | 7.37 | 2.56 | 2.16 | 1.83 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baggio, G.; Dupas, E.; Galindo, F.S.; Megda, M.M.; Pereira, N.C.M.; Luchetta, M.O.; Tritapepe, C.A.; da Silva, M.R.; Jalal, A.; Teixeira Filho, M.C.M. Silicon Application Induced Alleviation of Aluminum Toxicity in Xaraés Palisadegrass. Agronomy 2021, 11, 1938. https://doi.org/10.3390/agronomy11101938
Baggio G, Dupas E, Galindo FS, Megda MM, Pereira NCM, Luchetta MO, Tritapepe CA, da Silva MR, Jalal A, Teixeira Filho MCM. Silicon Application Induced Alleviation of Aluminum Toxicity in Xaraés Palisadegrass. Agronomy. 2021; 11(10):1938. https://doi.org/10.3390/agronomy11101938
Chicago/Turabian StyleBaggio, Guilherme, Elisângela Dupas, Fernando Shintate Galindo, Marcio Mahmoud Megda, Nathália Cristina Marchiori Pereira, Monique Oliveira Luchetta, Caio Augusto Tritapepe, Marcelo Rinaldi da Silva, Arshad Jalal, and Marcelo Carvalho Minhoto Teixeira Filho. 2021. "Silicon Application Induced Alleviation of Aluminum Toxicity in Xaraés Palisadegrass" Agronomy 11, no. 10: 1938. https://doi.org/10.3390/agronomy11101938