
The Use of Thermal Time to Describe and Predict the Growth and Nutritive Value of Lolium perenne L. and Bromus valdivianus Phil


Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Experimental Design
2.2. Climatic Information
2.3. Yield Components
2.4. Nutritive Value
2.5. Statistical Analysis and Model Selection
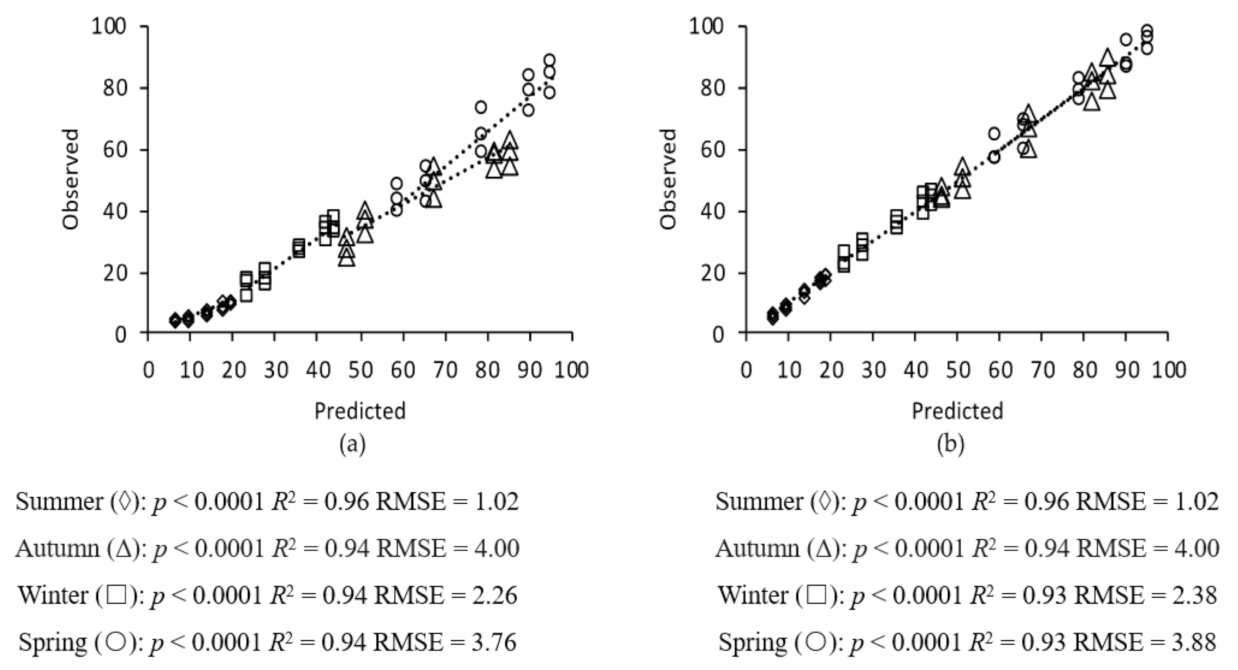
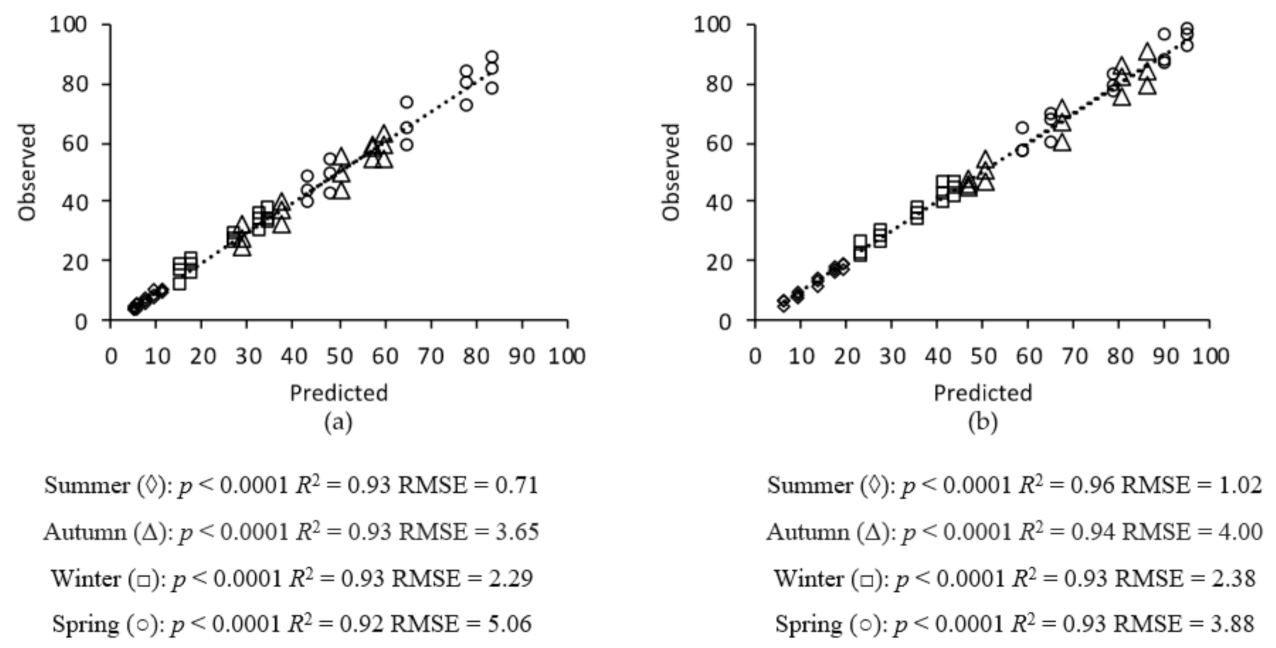
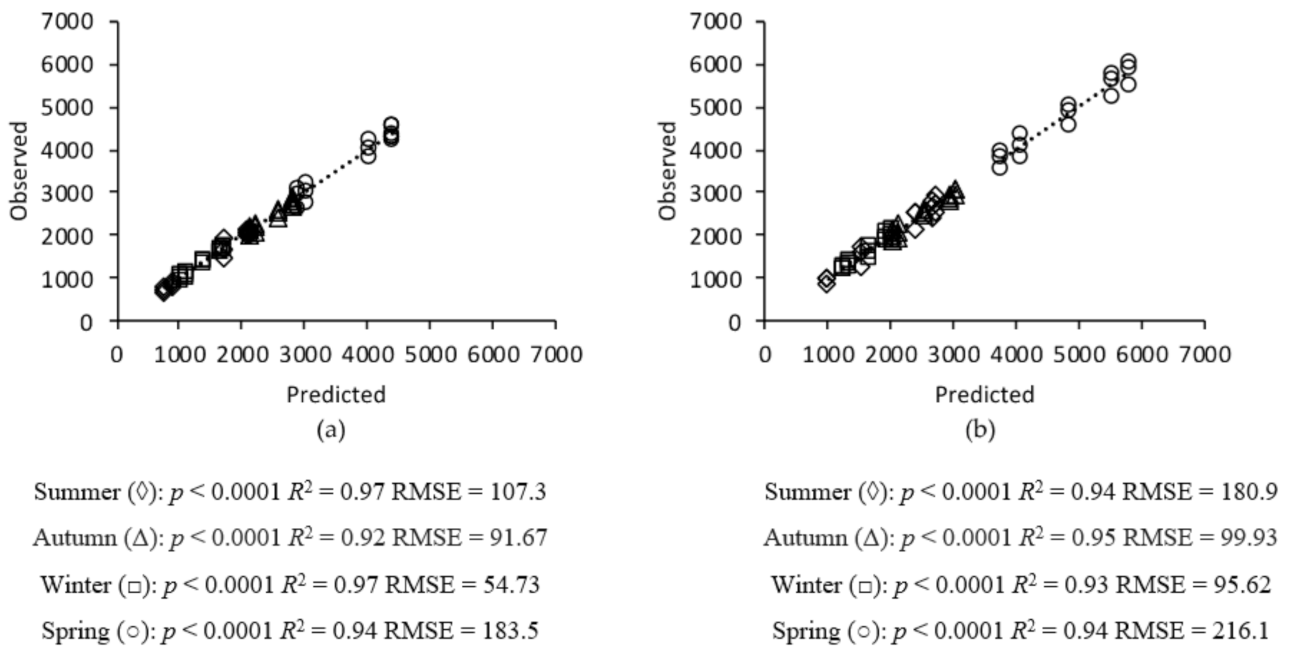
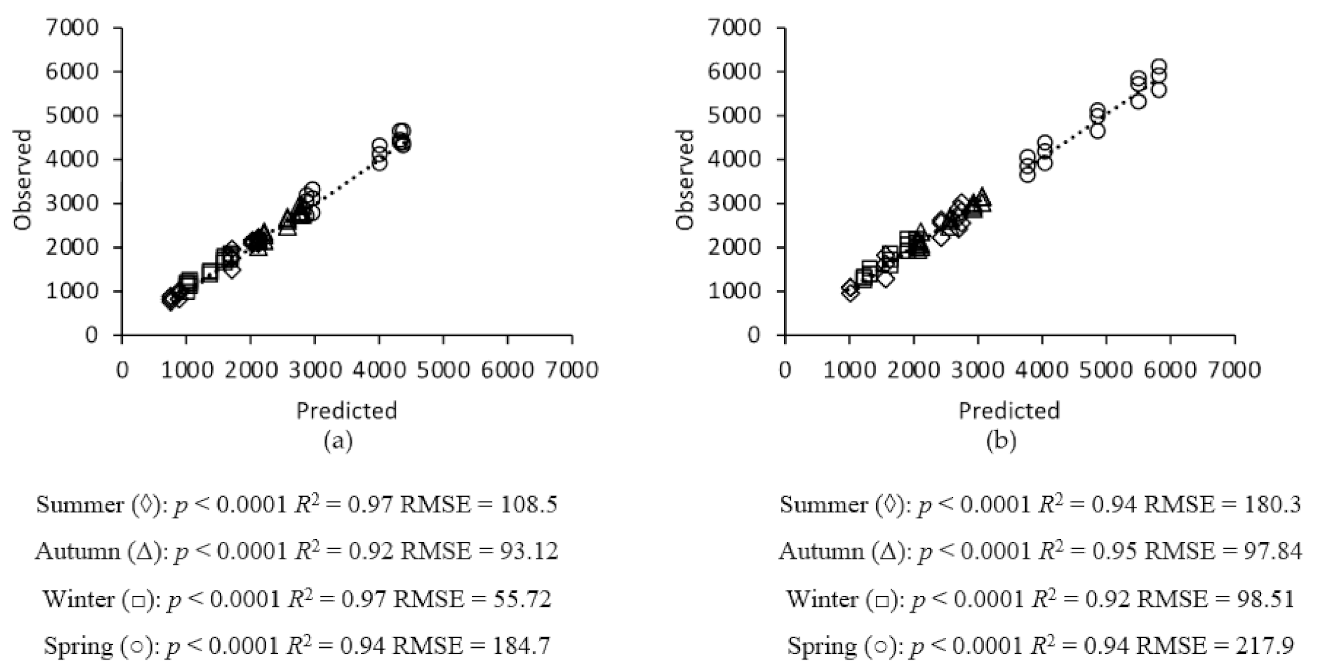
2.6. Model Validation
3. Results
3.1. Grass Growth
3.2. Nutritive Value
4. Discussion
4.1. Grass Growth
4.2. Nutritive Value
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McMaster, G.S.; Wilhelm, W. Growing degree-days: One equation, two interpretations. Agric. For. Meteorol. 1997, 87, 291–300. [Google Scholar] [CrossRef] [Green Version]
- Elnesr, M.; Alazba, A. An integral model to calculate the growing degree-days and heat units, a spreadsheet application. Comput. Electron. Agric. 2016, 124, 37–45. [Google Scholar] [CrossRef]
- Herz, K.; Dietz, S.; Haider, S.; Jandt, U.; Scheel, D.; Bruelheide, H. Predicting individual plant performance in grasslands. Ecol. Evol. 2017, 7, 8958–8965. [Google Scholar] [CrossRef]
- Babu, R.; Rao, K.L.N.; Ashokarani, Y.; Martiluther, M.; Prasad, P.R.K. Prediction of phenological stages of six maize (Zea mays L.) hybrids based on required growing degree days. Int. J. Chem. Stud. 2020, 8, 2564–2574. [Google Scholar]
- Ahanger, M.A.; Morad-Talab, N.; Abd-Allah, E.F.; Ahmad, P.; Hajiboland, R. Plant growth under drought stress: Significance of mineral nutrients. Water Stress Crop. Plants A Sustain. Approach 2016, 2, 649–668. [Google Scholar]
- McMaster, G.S.; Wilhelm, W.W. Phenological responses of wheat and barley to water and temperature: Improving simulation models. J. Agric. Sci. 2003, 141, 129–147. [Google Scholar] [CrossRef] [Green Version]
- Ostadian Bidgoly, R.; Balouchi, H.; Soltani, E.; Moradi, A. Effect of temperature and water potential on Carthamus tinctorius L. seed germination: Quantification of the cardinal temperatures and modeling using hydrothermal time. Ind. Crops. Prod. 2018, 113, 121–127. [Google Scholar] [CrossRef]
- Kiniry, J.R.; Kim, S.; Williams, A.S.; Lock, T.R.; Kallenbach, R.L. Simulating bimodal tall fescue growth with a degree-day-based process-oriented plant model. Grass Forage Sci. 2018, 73, 432–439. [Google Scholar] [CrossRef]
- Hunt, R.C.; Shipley, B.R.; Askew, A.P. A Modern Tool for Classical Plant Growth Analysis. Ann. Bot. 2002, 90, 485–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archontoulis, S.V.; Miguez, F.E. Nonlinear Regression Models and Applications in Agricultural Research. Agron. J. 2015, 107. [Google Scholar] [CrossRef] [Green Version]
- Paine, C.E.T.; Marthews, T.R.; Vogt, D.R.; Purves, D.; Rees, M.; Hector, A.; Turnbull, L.A. How to fit nonlinear plant growth models and calculate growth rates: An update for ecologists. Methods Ecol. Evol. 2012, 3, 245–256. [Google Scholar] [CrossRef]
- Hoffmann, W.A.; Poorter, H. Avoiding Bias in Calculations of Relative Growth Rate. Ann. Bot. 2002, 90, 37–42. [Google Scholar] [CrossRef] [Green Version]
- Thornley, J.H.; France, J. Mathematical Models in Agriculture: Quantitative Methods for the Plant, Animal and Ecological Sciences, 2nd ed.; CABI: Wallingford, UK, 2007; pp. 136–208. [Google Scholar]
- Zhao, J.; Zhao, Y.; Song, Z.; Liu, H.; Liu, Y.; Yang, R. Genetic analysis of the main growth traits using random regression models in Japanese flounder (Paralichthys olivaceus). Aquac. Res. 2018, 49, 1504–1511. [Google Scholar] [CrossRef]
- Choler, P. Growth response of temperate mountain grasslands to inter-annual variations in snow cover duration. Biogeosciences 2015, 12, 3885–3897. [Google Scholar] [CrossRef] [Green Version]
- McClung, C.R.; Davis, S.J. Ambient thermometers in plants: From physiological outputs towards mechanisms of thermal sensing. Curr. Biol. 2010, 20, R1086–R1092. [Google Scholar] [CrossRef] [Green Version]
- Calvache, I.; Balocchi, O.; Alonso, M.; Keim, J.P.; López, I.F. Thermal Time as a Parameter to Determine Optimal Defoliation Frequency of Perennial Ryegrass (Lolium perenne L.) and Pasture Brome (Bromus valdivianus Phil.). Agronomy 2020, 10, 620. [Google Scholar] [CrossRef]
- Monteith, J.L. Climate and the efficiency of crop production in Britain. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1977, 281, 277–294. [Google Scholar]
- Thompson, R.; Clark, R.M. Spatio-temporal modelling and assessment of within-species phenological variability using thermal time methods. Int. J. Biometeorol. 2006, 50, 312–322. [Google Scholar] [CrossRef]
- Biligetu, B.; Coulman, B. Responses of Three Bromegrass (Bromus) Species to Defoliation under Different Growth Conditions. Int. J. Agron. 2010, 2010, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Zhu, Y.; Cao, W.; Dai, T.; Jiang, D. Predicting the protein content of grain in winter wheat with meteorological and genotypic factors. Plant. Prod. Sci. 2006, 9, 323–333. [Google Scholar] [CrossRef]
- Leys, C.; Ley, C.; Klein, O.; Bernard, P.; Licata, L. Detecting outliers: Do not use standard deviation around the mean, use absolute deviation around the median. J. Exp. Soc. Psychol. 2013, 49, 764–766. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, E.A.; Naugler, C.; Far, B.H. Emerging Business Intelligence Framework for a Clinical Laboratory through Big Data Analytics. Emerging Trends in Computational Biology, Bioinformatics, and Systems Biology: Algorithms and Software Tools; Elsevier/Morgan Kaufmann: New York, NY, USA, 2015; Chapter 32; pp. 577–602. [Google Scholar]
- Coleman, S.W.; Henry, D.A. Nutritive value of herbage. In Sheep Nutrition; Freer, M., Dove, H., Eds.; CSIRO Publishing: Collingwood, UK, 2002; pp. 1–26. [Google Scholar]
- Chuine, I.; Cour, P.; Rousseau, D. Selecting models to predict the timing of flowering of temperate trees: Implications for tree phenology modelling. Plant Cell Environ. 1999, 22, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.; Kawamura, K.; Guo, W.; Xuan, T.D.; Lim, J.; Yuba, N.; Kurokawa, Y.; Obitsu, T.; Lv, R.; Tsumiyama, Y.; et al. A simple visible and near-infrared (V-NIR) camera system for monitoring the leaf area index and growth stage of Italian ryegrass. Comput. Electron. Agric. 2018, 144, 314–323. [Google Scholar] [CrossRef]
- Karimi, S.; Sadraddini, A.A.; Nazemi, A.H.; Xu, T.; Fard, A.F. Generalizability of gene expression programming and random forest methodologies in estimating cropland and grassland leaf area index. Comput. Electron. Agric. 2018, 144, 232–240. [Google Scholar] [CrossRef]
- Li, Q.; Xu, L.; Pan, X.; Zhang, L.; Li, C.; Yang, N.; Qi, J. Modeling phenological responses of Inner Mongolia grassland species to regional climate change. Environ. Res. Lett. 2016, 11, 015002. [Google Scholar] [CrossRef]
- Yan, W.; Wallace, D.H.; Ross, J. A Model of Photoperiod × Temperature Interaction Effects on Plant Development. Crit. Rev. Plant. Sci. 2011, 15, 63–96. [Google Scholar]
- Orloff, S.B.; Brummer, E.C.; Shrestha, A.; Putnam, D.H. Cool-Season Perennial Grasses Differ in Tolerance to Partial-Season Irrigation Deficits. Agron. J. 2016, 108, 692–700. [Google Scholar] [CrossRef]
- Bartholomew, P.W. Effect of Varying Temperature Regime on Phyllochron in Four Warm-Season Pasture Grasses. Agric. Sci. 2014, 5, 1000–1006. [Google Scholar] [CrossRef]
- Ruelland, E.; Zachowski, A. How plants sense temperature. Environ. Exp. Bot. 2010, 69, 225–232. [Google Scholar] [CrossRef]
- Rapacz, M.; Ergon, A.; Hoglind, M.; Jorgensen, M.; Jurczyk, B.; Ostrem, L.; Rognli, O.A.; Tronsmo, A.M. Overwintering of herbaceous plants in a changing climate. Still more questions than answers. Plant. Sci. 2014, 225, 34–44. [Google Scholar] [CrossRef]
- Lanier, W.E.; Bailey, L.L.; Muths, E. Integrating biology, field logistics, and simulations to optimize parameter estimation for imperiled species. Ecol. Model. 2016, 335, 16–23. [Google Scholar] [CrossRef]
- Pulina, A.; Lai, R.; Salis, L.; Seddaiu, G.; Roggero, P.P.; Bellocchi, G. Modelling pasture production and soil temperature, water and carbon fluxes in Mediterranean grassland systems with the Pasture Simulation model. Grass Forage Sci. 2018, 73, 272–283. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, H.; Okamoto, K.; Fukuhara, M. Estimation of aboveground grassland phytomass with a growth model using Landsat TM and climate data. Int. J. Remote Sens. 2010, 20, 2283–2294. [Google Scholar] [CrossRef]
- Chen, N.; Zhang, Y.; Zhu, J.; Li, J.; Liu, Y.; Zu, J.; Wang, L. Nonlinear responses of productivity and diversity of alpine meadow communities to degradation. Chin. J. Plant. Ecol. 2018, 42, 50–65. [Google Scholar]
- Bai, Y.; Wu, J.; Clark, C.M.; Pan, Q.; Zhang, L.; Chen, S.; Wang, Q.; Han, X.; Wisley, B. Grazing alters ecosystem functioning and C:N:P stoichiometry of grasslands along a regional precipitation gradient. J. Appl. Ecol. 2012, 49, 1204–1215. [Google Scholar] [CrossRef]
- Belesky, D.; Ruckle, J.; Abaye, A. Seasonal distribution of herbage mass and nutritive value of Prairiegrass (Bromus catharticus Vahl). Grass Forage Sci. 2007, 62, 301–311. [Google Scholar] [CrossRef]
- Davidson, A.; Da Silva, D.; Quintana, B.; DeJong, T.M. The phyllochron of Prunus persica shoots is relatively constant under controlled growth conditions but seasonally increases in the field in ways unrelated to patterns of temperature or radiation. Sci. Hortic. 2015, 184, 106–113. [Google Scholar] [CrossRef]
- Kumudini, S.; Andrade, F.H.; Boote, K.J.; Brown, G.A.; Dzotsi, K.A.; Edmeades, G.O.; Gocken, T.; Goodwin, M.; Halter, A.L.; Hammer, G.L.; et al. Predicting Maize Phenology: Intercomparison of Functions for Developmental Response to Temperature. Agron. J. 2014, 106, 2087–2097. [Google Scholar] [CrossRef] [Green Version]
- Robins, J.G. Evaluation of warm-season grasses nutritive value as alternatives to cool-season grasses under limited irrigation. Grassl. Sci. 2016, 62, 144–150. [Google Scholar] [CrossRef]
- Bilge, A.H.; Ozdemir, Y. Determining the Critical Point of a Sigmoidal Curve via its Fourier Transform. J. Phys. Conf. Ser. 2016, 738, 012062. [Google Scholar] [CrossRef] [Green Version]
- Pontes, L.S.; Carrere, P.; Andueza, D.; Louault, F.; Soussana, J. Seasonal productivity and nutritive value of temperate grasses found in semi-natural pastures in Europe: Responses to cutting frequency and N supply. Grass Forage Sci. 2007, 62, 485–496. [Google Scholar] [CrossRef]
- Ren, H.; Han, G.; Lan, Z.; Wan, H.; Schönbach, P.; Gierus, M.; Taube, F. Grazing effects on herbage nutritive values depend on precipitation and growing season in Inner Mongolian grassland. J. Plant. Ecol. 2016, 9, 712–723. [Google Scholar] [CrossRef] [Green Version]
- Mládek, J.; Hejcman, M.; Hejduk, S.; Duchoslav, M.; Pavlů, V. Community Seasonal Development Enables Late Defoliation Without Loss of Forage Quality in Semi-natural Grasslands. Folia Geobot. 2010, 46, 17–34. [Google Scholar] [CrossRef]
- Sanderson, M.A.; Stout, R.; Brink, G. Productivity, Botanical Composition, and Nutritive Value of Commercial Pasture Mixtures. Agron. J. 2016, 108, 450–456. [Google Scholar] [CrossRef]
- Bertilsson, J.; Akerlind, M.; Eriksson, T. The effects of high-sugar ryegrass/red clover silage diets on intake, production, digestibility, and N utilization in dairy cows, as measured in vivo and predicted by the NorFor model. J. Dairy Sci. 2017, 100, 7990–8003. [Google Scholar] [CrossRef] [PubMed]
- Gauch, H.G.; Hwang, J.T.; Fick, G.W. Model evaluation by comparison of model-based predictions and measured values. Agron. J. 2003, 95, 1442–1446. [Google Scholar] [CrossRef] [Green Version]
- Ruelle, E.; Hennessy, D.; Delaby, L. Development of the Moorepark St Gilles grass growth model (MoSt GG model): A predictive model for grass growth for pasture based systems. Eur. J. Agron. 2018, 99, 80–91. [Google Scholar] [CrossRef]
- Bruinenberg, M.; Valk, H.; Korevaar, H.; Struik, P. Factors affecting digestibility of temperate forages from seminatural grasslands: A review. Grass Forage Sci. 2002, 57, 292–301. [Google Scholar] [CrossRef]
- Lee, J.M.; Clark, A.J.; Roche, J.R. Climate-change effects and adaptation options for temperate pasture-based dairy farming systems: A review. Grass Forage Sci. 2013, 68, 485–503. [Google Scholar] [CrossRef]
- Tasset, E.; Boulanger, T.; Diquélou, S.; Laîné, P.; Lemauviel-Lavenant, S. Plant trait to fodder quality relationships at both species and community levels in wet grasslands. Ecol. Indic. 2019, 97, 389–397. [Google Scholar] [CrossRef]
- Cao, R.; Francisco-Fernández, M.; Anand, A.; Bastida, F.; González-Andújar, J.L. Modeling Bromus diandrus seedling emergence using nonparametric estimation. J. Agric. Biol. Environ. Stat. 2013, 18, 64–86. [Google Scholar] [CrossRef] [Green Version]
- Insua, J.R.; Agnusdei, M.G.; Berone, G.D.; Basso, B.; Machado, C.F. Modeling the Nutritive Value of Defoliated Tall Fescue Pastures Based on Leaf Morphogenesis. Agron. J. 2019, 111, 714–724. [Google Scholar] [CrossRef]
- Loaiza, P.A.; Balocchi, O.; Bertrand, A. Carbohydrate and crude protein fractions in perennial ryegrass as affected by defoliation frequency and nitrogen application rate. Grass Forage Sci. 2017, 72, 556–567. [Google Scholar] [CrossRef]
- Smart, A.J.; Schacht, W.H.; Volesky, J.D.; Moser, L.E. Seasonal Changes in Dry Matter Partitioning, Yield, and Crude Protein of Intermediate Wheatgrass and Smooth Bromegrass. Agron. J. 2006, 98, 986–991. [Google Scholar] [CrossRef] [Green Version]
- Jing, Q.; Jégo, G.; Bélanger, G.; Chantigny, M.H.; Rochette, P. Simulation of water and nitrogen balances in a perennial forage system using the STICS model. Field Crop. Res. 2017, 201, 10–18. [Google Scholar] [CrossRef]
- Persson, T.; Höglind, M.; Van Oijen, M.; Korhonen, P.; Palosuo, T.; Jégo, G.; Virkajärvi, P.; Bélanger, G.; Gustavsson, A.M. Simulation of timothy nutritive value: A comparison of three process-based models. Field Crop. Res. 2019, 231, 81–92. [Google Scholar] [CrossRef]
- Frary, A. Plant Physiology and Development. Rhodora 2015, 117, 397–399. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Season | Model | Leaf Blade Length (cm) | Accumulated Herbage Mass (kg DM ha−1) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| AIC | BIC | RMSE | R2 | AIC | BIC | RMSE | R2 | |||
| L. perenne | Summer | Logistic | 0.55 | 43.19 | 0.76 | 0.93 | 0.54 | 194.23 | 116.65 | 0.97 |
| Gompertz | 0.45 | 43.61 | 0.77 | 0.93 | 0.46 | 194.57 | 117.96 | 0.97 | ||
| Autumn | Logistic | 0.51 | 92.75 | 3.96 | 0.92 | 0.56 | 189.51 | 99.65 | 0.92 | |
| Gompertz | 0.49 | 92.84 | 3.97 | 0.92 | 0.44 | 189.98 | 101.23 | 0.92 | ||
| Winter | Logistic | 0.51 | 78.66 | 2.48 | 0.93 | 0.57 | 174.03 | 59.50 | 0.97 | |
| Gompertz | 0.49 | 78.76 | 2.48 | 0.93 | 0.43 | 174.57 | 60.57 | 0.97 | ||
| Spring | Logistic | 0.48 | 102.78 | 5.53 | 0.92 | 0.52 | 210.33 | 199.50 | 0.94 | |
| Gompertz | 0.52 | 102.61 | 5.50 | 0.92 | 0.48 | 210.52 | 200.77 | 0.94 | ||
| B. valdivianus | Summer | Logistic | 0.53 | 54.56 | 1.11 | 0.96 | 0.49 | 209.90 | 196.68 | 0.94 |
| Gompertz | 0.47 | 54.83 | 1.12 | 0.96 | 0.51 | 209.79 | 195.95 | 0.94 | ||
| Autumn | Logistic | 0.57 | 95.51 | 4.34 | 0.95 | 0.42 | 192.09 | 108.63 | 0.95 | |
| Gompertz | 0.43 | 96.06 | 4.42 | 0.94 | 0.58 | 191.46 | 106.36 | 0.95 | ||
| Winter | Logistic | 0.54 | 78.46 | 2.46 | 0.94 | 0.61 | 190.77 | 103.95 | 0.93 | |
| Gompertz | 0.46 | 78.76 | 2.48 | 0.93 | 0.39 | 191.67 | 107.09 | 0.92 | ||
| Spring | Logistic | 0.48 | 93.66 | 4.08 | 0.94 | 0.53 | 215.24 | 234.95 | 0.94 | |
| Gompertz | 0.52 | 93.51 | 4.06 | 0.94 | 0.47 | 215.48 | 236.88 | 0.94 | ||
| Species | Season | Model | Leaf Blade Length (cm) | Accumulated Herbage Mass (kg DM ha−1) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| a-GR | b-Pmax | c-AInf | d-ASup | a-GR | b-Pmax | c-AInf | d-ASup | |||
| L. perenne | Summer | Logistic | 0.006 ± 0.003 | 303.9 ± 28.1 | 4.25 ± 0.74 | 11.45 ± 1.25 | 0.015 ± 0.004 | 244.9 ± 9.7 | 764.7 ± 74.3 | 2106 ± 52.2 |
| Gompertz | 0.008 ± 0.004 | 288.1 ± 42.7 | 4.46 ± 0.55 | 12.67 ± 0.55 | 0.021 ± 0.004 | 222.3 ± 10.3 | 773.3 ± 67.8 | 2131 ± 62.7 | ||
| Autumn | Logistic | 0.007 ± 0.003 | 220.1 ± 26.2 | 25.56 ± 5.79 | 60.12 ± 2.81 | 0.01 ± 0.005 | 249.1 ± 17.1 | 2116.9 ± 70.9 | 2822 ± 51.9 | |
| Gompertz | 0.01 ± 0.005 | 201.8 ± 19.8 | 28.3 ± 3.00 | 60.83 ± 3.33 | 0.02 ± 0.005 | 225.1 ± 18.2 | 2126 ± 58.3 | 2842 ± 66.3 | ||
| Winter | Logistic | 0.009 ± 0.003 | 250.3 ± 17.2 | 14.86 ± 2.00 | 34.79 ± 1.5 | 0.009 ± 0.002 | 265.4 ± 11.9 | 1002 ± 47.3 | 1735 ± 44.1 | |
| Gompertz | 0.01 ± 0.004 | 227.5 ± 17.4 | 15.4 ± 1.45 | 35.53 ± 1.91 | 0.01 ± 0.003 | 242.8 ± 13.0 | 1023 ± 35.4 | 1776 ± 61.3 | ||
| Spring | Logistic | 0.008 ± 0.003 | 262.5 ± 21.2 | 41.97 ± 4.87 | 84.32 ± 4.35 | 0.01 ± 0.008 | 242.9 ± 16.0 | 2912 ± 120.5 | 4415 ± 84.4 | |
| Gompertz | 0.01 ± 0.004 | 240.6 ± 21.3 | 43.6 2 ± 3.26 | 86.44 ± 5.66 | 0.02 ± 0.008 | 220.3 ± 16.2 | 2916 ± 114.6 | 4429 ± 95.2 | ||
| B. valdivianus | Summer | Logistic | 0.006 ± 0.002 | 245.4 ± 21.1 | 4.84 ± 1.91 | 19.71 ± 1.30 | 0.01 ± 0.003 | 204.8 ± 15.4 | 884.03 ± 197 | 2736 ± 97.2 |
| Gompertz | 0.009 ± 0.003 | 224.6 ± 18.7 | 6.00 ± 1.08 | 20.44 ± 1.72 | 0.02 ± 0.005 | 188.1 ± 11.3 | 991.4 ± 117 | 2751 ± 107.2 | ||
| Autumn | Logistic | 0.009 ± 0.003 | 264.5 ± 15.2 | 45.38 ± 3.37 | 86.0 ± 3.10 | 0.009 ± 0.003 | 266.1 ± 13.7 | 2007 ± 79.7 | 3067 ± 75.3 | |
| Gompertz | 0.01 ± 0.004 | 241.5 ± 16.7 | 46.40 ± 2.58 | 88.27 ± 4.34 | 0.01 ± 0.004 | 245.5 ± 14.4 | 2039 ± 60.4 | 3118 ± 97 | ||
| Winter | Logistic | 0.007 ± 0.003 | 244.67 ± 21.7 | 22.2 ± 2.9 | 44.85 ± 2.00 | 0.008 ± 0.003 | 267.0 ± 18.6 | 1221 ± 83 | 2056 ± 79.3 | |
| Gompertz | 0.01 ± 0.004 | 221.9 ± 20 | 23.4 ± 1.73 | 45.78 ± 2.61 | 0.01 ± 0.004 | 242.3 ± 21.6 | 1237 ± 65 | 2113 ± 120 | ||
| Spring | Logistic | 0.006 ± 0.002 | 256.54 ± 22.4 | 56.6 ± 5.15 | 97 ± 4.20 | 0.007 ± 0.003 | 264.2 ± 19.5 | 3699 ± 226.8 | 5891 ± 210.7 | |
| Gompertz | 0.01 ± 0.003 | 235 ± 20.8 | 59.1 ± 2.86 | 99 ± 5.33 | 0.01 ± 0.004 | 241.4 ± 20.3 | 3788 ± 151 | 6025 ± 294.3 | ||
| Variable | Species | Season | Model | R2 | RMSE | p < |t| |
|---|---|---|---|---|---|---|
| CP (g 100 g−1) | L. Perenne | Summer | Y = 18.55(1.34) + 0.032(0.01) × AGDD−9.532e − 5(2.06 × 10−5) × AGDD2 | 0.87 | 1.08 | 0.001 |
| Autumn | Y = 27.98(0.36) − 0.02(0.001) × AGDD | 0.96 | 0.60 | 0.001 | ||
| Winter | Y = 20.69(0.32) − 0.01(0.001) × AGDD | 0.95 | 0.53 | 0.001 | ||
| Spring | Y = 22.92(0.54) − 0.03(0.001) × AGDD | 0.96 | 0.90 | 0.001 | ||
| B. valdivianus | Summer | Y = 17.83(1.12) + 0.02(0.009) × AGDD − 7.6161 × 10−5 (1.73 × 10−5) × AGDD2 | 0.85 | 0.91 | 0.001 | |
| Autumn | Y = 30.471(0.48) − 0.01(0.001) × AGDD | 0.91 | 0.81 | 0.001 | ||
| Winter | Y = 23.88(0.32) − 0.02(0.001) × AGDD | 0.96 | 0.54 | 0.001 | ||
| Spring | Y = 23.29(0.60) − 0.03(0.002) × AGDD | 0.95 | 1.00 | 0.001 | ||
| ME (Mcal kg DM−1) | L. perenne | Summer | Y = 2.24(0.06) + 0.002(5.1 × 10−4) × AGDD − 6.05 × 10−6 (9.2 × 10−7) × AGDD2 | 0.81 | 0.05 | 0.001 |
| Autumn | Y = 2.68(0.01) + 0.0003(8.6 × 10−5) × AGDD − 9.70 × 10−7 (1.5 × 10−7) × AGDD2 | 0.91 | 0.01 | 0.001 | ||
| Winter | Y = 2.61(3.03) + 2.6 × 10−4(3.6 × 10−4) × AGDD − 1.46 × 10−6 (5.5 × 10−7) × AGDD2 | 0.86 | 0.03 | 0.021 | ||
| Spring | Y = 2.59(0.02) + 2.6 × 10−4(2.2 × 10−4) × AGDD − 9.40 × 10−7 (4.0 × 10−7) × AGDD2 | 0.75 | 0.02 | 0.001 | ||
| B. valdivianus | Summer | Y = 2.29(0.02) + 7.2 × 10−4(1.7 × 10−4)×AGDD − 1.96 × 10−6 (3.1 × 10−7) × AGDD2 | 0.91 | 0.02 | 0.001 | |
| Autumn | Y = 2.57(0.01) + 3.0 × 10−4(1.13 × 10−4)×AGDD − 7.64 × 10−7(2.1 × 10−7) × AGDD2 | 0.69 | 0.01 | 0.004 | ||
| Winter | Y = 2.50(0.02) + 7.3 × 10−5(2.2 × 10−4) × AGDD − 9.11 × 10−7(4.1 × 10−4) × AGDD2 | 0.87 | 0.02 | 0.049 | ||
| Spring | Y = 2.50(0.03) + 8.2 ×10−4 (2.5 × 10−4) × AGDD − 2.11 × 10−6 (4.6 × 10−7) × AGDD2 | 0.83 | 0.02 | 0.001 | ||
| NDF (g 100 g−1) | L. perenne | Summer | Y = 42.37(0.45) + 0.01(0.001) × AGDD | 0.78 | 0.75 | 0.001 |
| Autumn | Y = 39.56(0.60) + 0.03(0.002) × AGDD | 0.95 | 1.00 | 0.001 | ||
| Winter | Y = 43.02(0.28) + 0.01(9.6 ×1 0−4) × AGDD | 0.94 | 0.47 | 0.001 | ||
| Spring | Y = 35.08(1.29) + 0.06(0.004) × AGDD | 0.93 | 2.13 | 0.001 | ||
| B. valdivianus | Summer | Y = 50.85(0.42) + 0.01(0.001) × AGDD | 0.90 | 0.71 | 0.001 | |
| Autumn | Y = 49.36(0.49) + 0.02(0.001) × AGDD | 0.92 | 0.82 | 0.001 | ||
| Winter | Y = 50.10(0.37) + 0.01(0.001) × AGDD | 0.91 | 0.61 | 0.001 | ||
| Spring | Y = 51.78(0.34) + 0.02(0.001) × AGDD | 0.98 | 0.57 | 0.001 | ||
| ADF (g 100 g−1) | L. Perenne | Summer | Y = 19.62(0.55) + 0.02(0.001) × AGDD | 0.92 | 0.92 | 0.001 |
| Autumn | Y = 20.34(0.49) + 0.02(0.001) × AGDD | 0.93 | 0.85 | 0.001 | ||
| Winter | Y = 20.90(0.70) + 0.01(0.002) × AGDD | 0.93 | 0.59 | 0.001 | ||
| Spring | Y = 17.80(0.80) + 0.04(0.002) × AGDD | 0.94 | 1.34 | 0.001 | ||
| B. valdivianus | Summer | Y = 28.23(0.47) + 0.01(0.001) × AGDD | 0.87 | 0.78 | 0.001 | |
| Autumn | Y = 23.70(0.49) + 0.02(0.001) × AGDD | 0.92 | 0.81 | 0.001 | ||
| Winter | Y = 22.81(0.70) + 0.02(0.002) × AGDD | 0.89 | 1.16 | 0.001 | ||
| Spring | Y = 20.51(0.80) + 0.03(0.002) × AGDD | 0.94 | 1.33 | 0.001 | ||
| WSC (g 1000 g−1) | L. Perenne | Summer | Y = 72.85(3.41) + 0.17(0.02) × AGDD − 1.8 × 10−4 (5.2 × 10−5) × AGDD2 | 0.93 | 2.75 | 0.004 |
| Autumn | Y = 89.71(4.88) + 0.17(0.04) × AGDD − 1.8 × 10−4 (7.5 × 10−5) × AGDD2 | 0.87 | 3.94 | 0.030 | ||
| Winter | Y = 74.46(5.27) + 0.20(0.01) × AGDD | 0.90 | 8.71 | 0.001 | ||
| Spring | Y = 60.97(8.4) + 0.41(0.07) × AGDD − 4.7 × 10−4 (1.3e−04) × AGDD2 | 0.91 | 6.80 | 0.003 | ||
| B. valdivianus | Summer | Y = 67.71(3.43) + 0.16(0.02) × AGDD − 1.9 × 10−4 (5.2 × 10−5) × AGDD2 | 0.90 | 2.77 | 0.003 | |
| Autumn | Y = 71.40(3.16) + 0.15(0.03) × AGDD − 1.6 × 10−4 (5.56 × 10−5) × AGDD2 | 0.91 | 2.92 | 0.013 | ||
| Winter | Y = 76.43(2.91) + 0.09(0.009) × AGDD | 0.88 | 4.82 | 0.001 | ||
| Spring | Y = 80.56(3.87) + 0.14(0.03) × AGDD − 1.5 × 10−4 (5.9 × 10−5) × AGDD2 | 0.88 | 3.13 | 0.021 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calvache, I.; Balocchi, O.; Arias, R.; Alonso, M. The Use of Thermal Time to Describe and Predict the Growth and Nutritive Value of Lolium perenne L. and Bromus valdivianus Phil. Agronomy 2021, 11, 774. https://doi.org/10.3390/agronomy11040774
Calvache I, Balocchi O, Arias R, Alonso M. The Use of Thermal Time to Describe and Predict the Growth and Nutritive Value of Lolium perenne L. and Bromus valdivianus Phil. Agronomy. 2021; 11(4):774. https://doi.org/10.3390/agronomy11040774
Chicago/Turabian StyleCalvache, Iván, Oscar Balocchi, Rodrigo Arias, and Máximo Alonso. 2021. "The Use of Thermal Time to Describe and Predict the Growth and Nutritive Value of Lolium perenne L. and Bromus valdivianus Phil" Agronomy 11, no. 4: 774. https://doi.org/10.3390/agronomy11040774