Congruity of the Polymorphisms in the Expressed and Noncoding Parts of the Gli-B1 Locus in Common Wheat


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Alleles at the Gli-B1 Locus and PCR Sequence Length Polymorphism in the Set of Cultivars Studied
3. Results
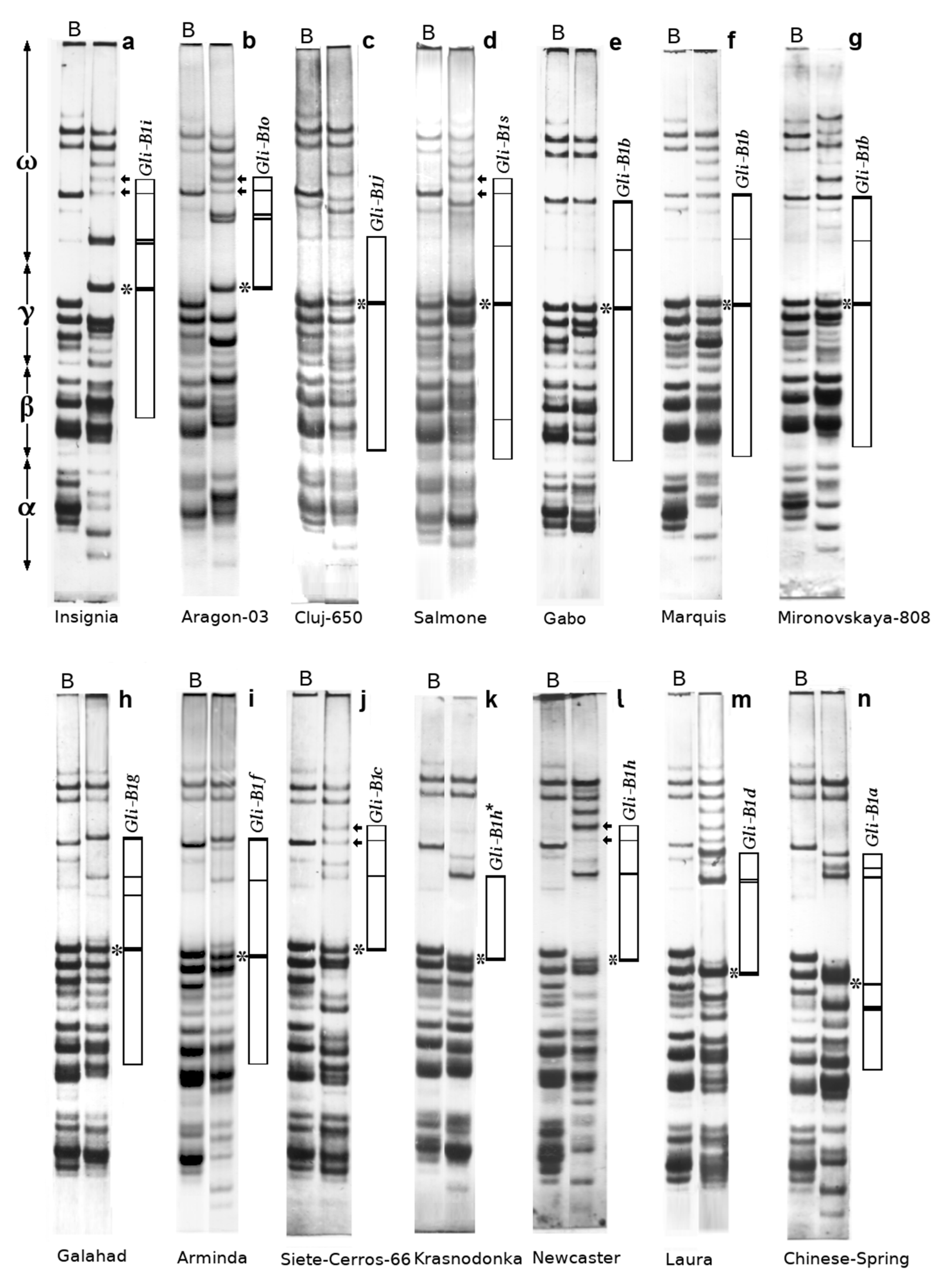
3.1. Allelic Variation at the Gli-B1 Locus and PCR Product Length Polymorphism among the Cultivars Studied
3.1.1. Allelic Variation
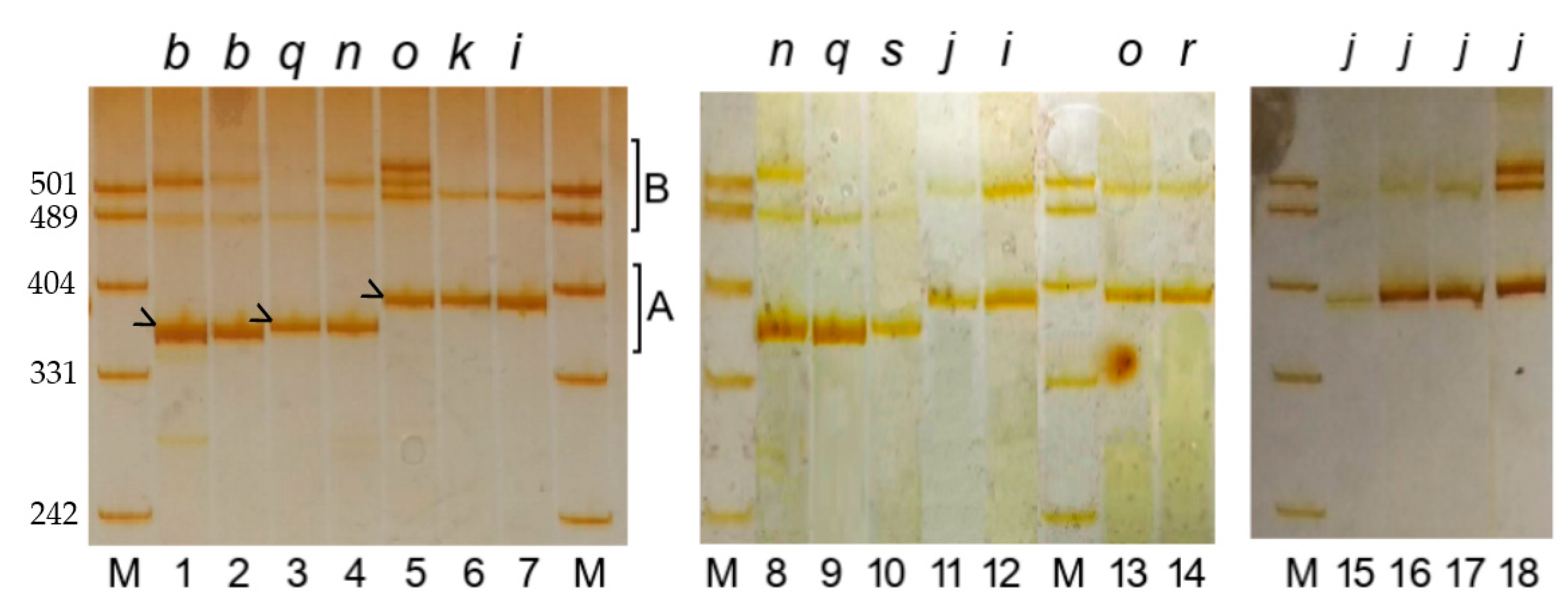
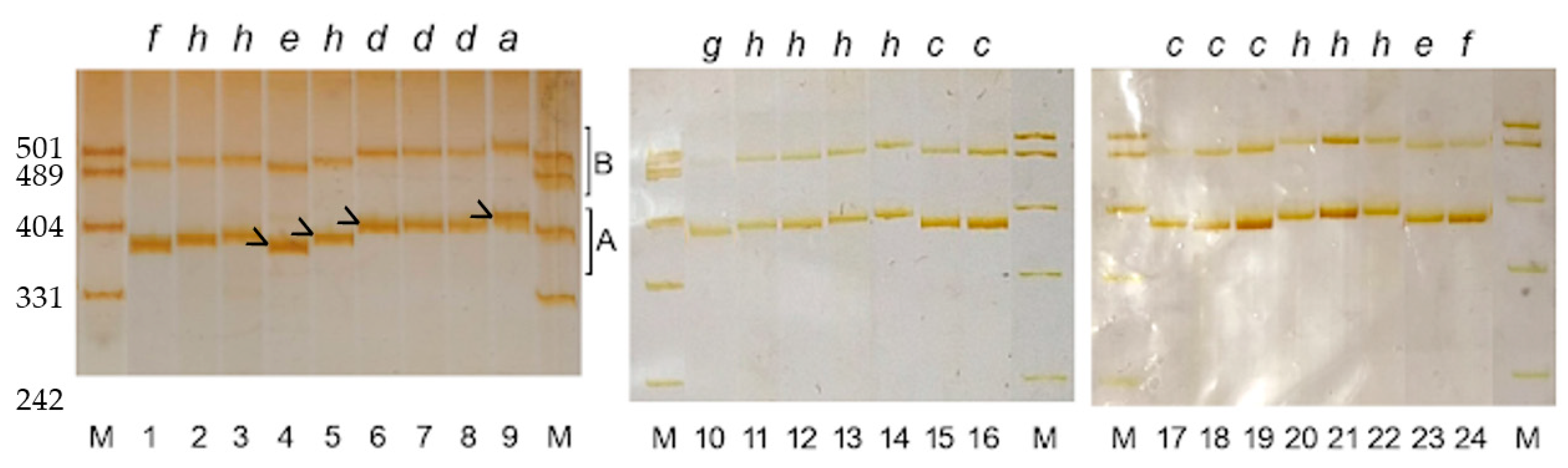
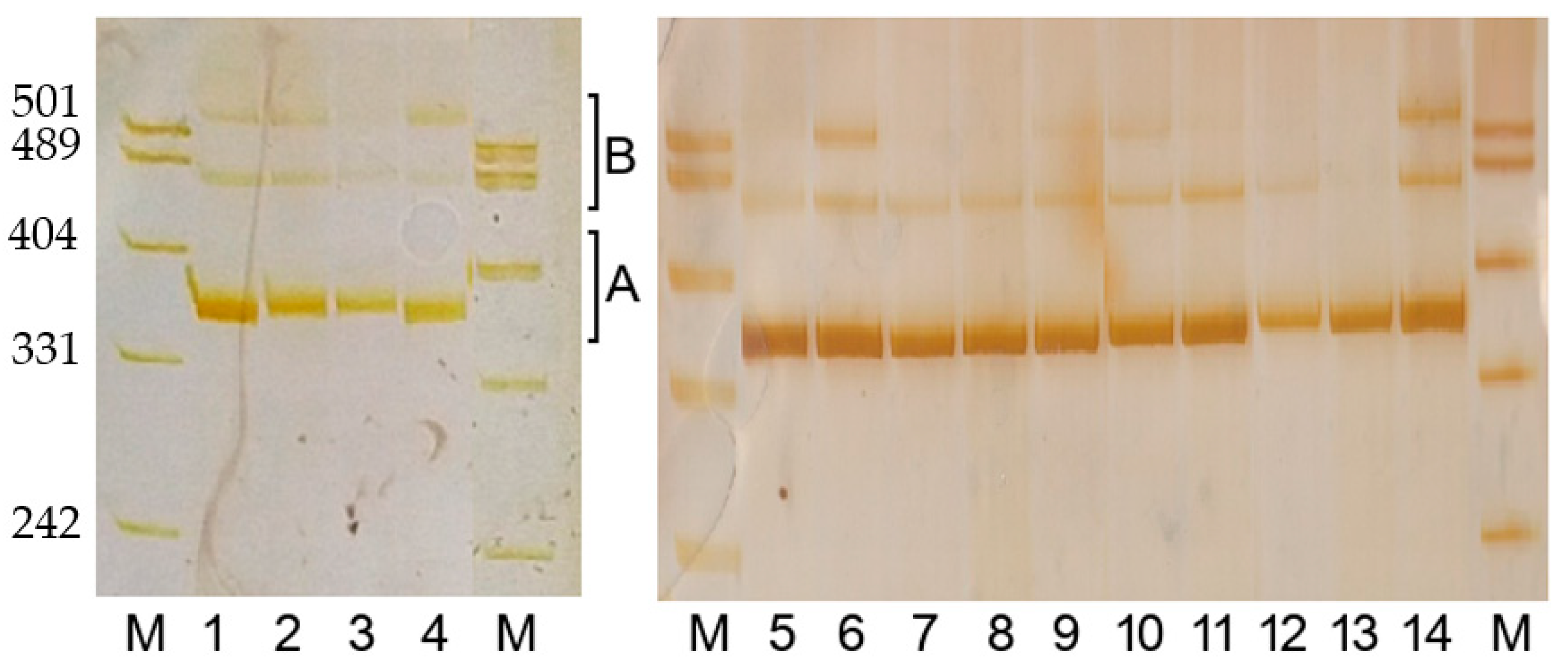
3.1.2. PCR Product Length Polymorphism
3.2. The Correspondence between PCR Product Polymorphism and Allelic Variation at the Gli-B1 Locus
3.3. The Probable Recombinational Nature of the Allele Gli-B1j
3.4. Variation in PCR Product Length among Cultivars Having Identical Alleles at the Gli-B1 Locus
3.5. Polymorphism of the PCR Products in “Zone B” of the Electrophoregram
3.6. The PCR Sequence Corresponds to Genotype, Not to the Label of the Grain Sample under Study
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Payne, P.I. Genetics of wheat storage proteins and the effect of allelic variation on bread-making quality. Annu. Rev. Plant Physiol. 1987, 38, 141–153. [Google Scholar] [CrossRef]
- Shewry, P.R.; Halford, N.G.; Lafiandra, D. Genetics of wheat gluten proteins. Adv. Genet. 2003, 49, 111–184. [Google Scholar] [PubMed]
- Gobaa, S.; Bancel, E.; Branlard, G.; Kleijer, G.; Stamp, P. Proteomic analysis of wheat recombinant inbred lines: Variations in prolamin and dough rheology. J. Cereal Sci. 2008, 47, 610–619. [Google Scholar] [CrossRef]
- Shewry, P.R.; Halford, N.G. Cereal seed storage proteins: Structures, properties and role in grain utilization. J. Exp. Bot. 2002, 53, 947–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barak, S.; Mudgil, D.; Khatkar, B.S. Biochemical and functional properties of wheat gliadins: A review. Crit. Rev. Food Sci. Nutr. 2015, 55, 357–368. [Google Scholar] [CrossRef]
- Sozinov, A.A.; Poperelya, F.A. Genetic classification of prolamins and its use for plant breeding. Ann. Technol. Agric. 1980, 29, 229–245. [Google Scholar]
- Metakovsky, E.; Melnik, V.A.; Rodriguez-Quijano, M.; Upelniek, V.P.; Carrillo, J.M. A catalog of glidin alleles: Polymorphism of 20th-century common wheat germplasm. Crop J. 2018, 6, 629–641. [Google Scholar] [CrossRef]
- Sandhu, D.; Champoux, J.A.; Bondareva, N.; Gill, K.S. Identification and physical localization of useful genes and markers to a major gene-rich region on wheat group 1S chromosomes. Genetics 2001, 157, 1735–1747. [Google Scholar]
- Saintenac, C.; Faure, S.; Remay, A.; Choulet, F.; Ravel, C.; Paux, E.; Balfourier, F.; Feuillet, C.; Sourdille, P. Variation in crossover rates across a 3-Mb contig of bread wheat (Triticum aestivum) reveals the presence of a meiotic recombination hotspot. Chromosoma 2011, 120, 185–198. [Google Scholar] [CrossRef]
- Jordan, K.W.; Wang, S.; He, F.; Chao, S.; Lun, Y.; Paux, E.; Sourdille, P.; Sherman, J.; Akhunova, A.; Blake, N.K.; et al. The genetic architecture of genome-wide recombination rate variation in allopolyploid wheat revealed by nested association mapping. Plant J. 2018, 95, 1039–1054. [Google Scholar] [CrossRef] [Green Version]
- Gill, K.S.; Gill, B.S.; Endo, T.R.; Taylor, T. Identification and high-density mapping of gene-rich regions in chromosome group 1 of wheat. Genetics 1996, 144, 1883–1891. [Google Scholar] [PubMed]
- Huang, X.Q.; Cloutier, S. Molecular characterization and genomic organization of low molecular weight glutenin subunit genes at the Glu-3 loci in hexaploid wheat (Triticum aestivum L.). Theor. Appl. Genet. 2008, 116, 953–966. [Google Scholar] [CrossRef] [PubMed]
- Qi, P.F.; Wei, Y.M.; Ouellet, T.; Chen, Q.; Tan, X.; Zheng, Y.L. The γ-gliadin multigene family in common wheat (Triticum aestivum) and its closely related species. BMC Genom. 2009, 10, 168–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Herpen, T.W.J.M.; Goryunova, S.V.; van der Schoot, J.; Mitreva, M.; Salentijn, E.; Vorst, O.; Schenk, M.F.; van Veelen, P.A.; Koning, F.; van Soest, L.J.M.; et al. Alpha-gliadin genes from the A, B, and D genomes of wheat contain different sets of celiac disease epitopes. BMC Genom. 2006, 7, 1. [Google Scholar] [CrossRef]
- von Büren, M. Polymorphisms in two homoeologous gamma-gliadin genes and the evolution of cultivated wheat. Genet. Res. Crop Evol. 2001, 48, 205–220. [Google Scholar] [CrossRef]
- Zhang, W.; Gianibelli, M.C.; Ma, W.; Rampling, L.R.; Gale, K.R. Identification of SNPs and development of allele-specific PCR markers for gamma-gliadin alleles in Triticum Aestivum. Theor. Appl. Genet. 2003, 107, 130–138. [Google Scholar] [CrossRef]
- Metakovsky, E.; Melnik, V.A.; Pascual, L.; Wrigley, C.W. Gliadin genotypes worldwide for spring wheats (Triticum aestivum L.). 2. Strong differentiation of polymorphism between countries and regions of origin. J. Cereal Sci. 2019, 87, 311–317. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Jackson, E.A.; Morel, M.H.; Sontag-Strohm, T.; Branlard, G.; Metakovsky, E.; Redaelli, R. Proposal for combining the classification systems of alleles of Gli-1 and Glu-3 loci in bread wheat (Triticum aestivum L.). J. Genet. Breed. 1996, 50, 321–336. [Google Scholar]
- Cao, W.; Scoles, G.; Hucl, P.; Chibbar, R.N. The use of RAPD analysis to classify Triticum accessions. Theor. Appl. Genet. 1999, 98, 602–607. [Google Scholar] [CrossRef]
- Metakovsky, E.; Melnik, V.A.; Pascual, L.; Romanov, G.A.; Wrigley, C.W. Types, frequences and value of intra-varietal genotypic non-uniformity in common wheat cultivars: Authentic biotypes and foreign seeds. J. Cereal Sci. 2019, 89. [Google Scholar] [CrossRef]
- Metakovsky, E.; Branlard, G.; Chernakov, V.M.; Upelniek, V.P.; Redaelli, R.; Pogna, N.E. Recombiation mapping of some chromosome 1A-, 1B-, 1D- and 6B-controlled gliadins and low molecular weight glutenin subunits in common wheat. Theor. Appl. Genet. 1997, 94, 788–795. [Google Scholar] [CrossRef]
- Gao, S.; Gu, Y.Q.; Wu, J.; Coleman-Derr, D.; Huo, N.; Crossman, C.; Jia, J.; Zuo, Q.; Ren, Z.; Anderson, O.D.; et al. Rapid evolution and complex structural organization in genomic regions harboring multiple prolamin genes in the polyploid wheat genome. Plant Mol. Biol. 2007, 65, 189–203. [Google Scholar] [CrossRef] [PubMed]
- Charles, M.; Belcram, H.; Just, J.; Huneau, C.; Violett, A.; Couloux, A.; Segurens, B.; Carter, M.; Huteau, V.; Coriton, O.; et al. Dynamics and differential proliferation of transposable elements during the evolution of the B and A genomes of wheat. Genetics 2008, 180, 1071–1086. [Google Scholar] [CrossRef] [Green Version]
- Wicker, T.; Mayer, K.F.X.; Gundlach, H.; Martis, M.; Steuernagel, B.; Scholz, U.; Šimková, H.; Kubaláková, M.; Choulet, F.; Taudien, S.; et al. Frequent gene movement and pseudogene evolution is common to the large and complex genomes of wheat, barley, and their relatives. Plant Cell 2011, 23, 1706–1718. [Google Scholar] [CrossRef] [Green Version]
- Pogna, N.E.; Metakovsky, E.; Redaelli, R.; Raineri, F.; Dachkevitch, T. Recombination mapping of Gli-5, a new gliadin-coding locus on chromosome 1A and 1B in common wheat. Theor. Appl. Genet. 1993, 87, 113–121. [Google Scholar] [CrossRef]
- Koval, S.F.; Metakovsky, E.; Kudryavtsev, A.M.; Sozinov, A.A. Non-accidental combinations of alleles at the Hg-1 and Rg-1 loci with definite alleles at the Gli-A1 and Gli-B1 loci in wheat. Selskokhoz. Biologia 1986, 2, 31–36. [Google Scholar]
- Metakovsky, E.; Pogna, N.E.; Biancardi, A.M.; Redaelli, R. Gliadin allele composition of common wheat cultivars grown in Italy. J. Genet. Breed. 1994, 48, 55–66. [Google Scholar]
- Metakovsky, E.; Gòmez, M.; Vàzquez, J.F.; Carrillo, J.M. High genetic diversity of Spanish common wheats as judged from gliadin alleles. Plant Breed. 2000, 119, 37–42. [Google Scholar] [CrossRef]
- Dvorak, J.; Luo, M.C.; Akhunov, E.D. NI Vavilov theory of centres of diversity in the light of current understanding of wheat diversity, domestication and evolution. Czech J. Genet. Plant Breed. 2011, 47, S20–S27. [Google Scholar] [CrossRef] [Green Version]
- Endo, T.R.; Gill, B.S. Somatic karyotype, heterochromatin distribution, and nature of chromosome differentiation in common wheat, Triticum aestivum L. em Thell. Chromosome 1984, 89, 361–369. [Google Scholar] [CrossRef]
- Friebe, B.; Gill, B.S. C-band polymorphism and structural rearrangements detected in common wheat (Triticum aestivum). Euphytica 1994, 78, 1–5. [Google Scholar]
- Dvořák, J.; Chen, K.C. Distribution of nonstructural variation between wheat cultivars along chromosome arm 6Bp: Evidence from the linkage map and physical map of the arm. Genetics 1984, 106, 325–333. [Google Scholar]
- Dubcovsky, J.; Luo, M.-C.; Dvořák, J. Differentiation between homoeologous chromosomes 1A of wheat and 1Am of Triticum monococcum and its recognition by the wheat Ph1 locus. Proc. Nat. Acad. Sci. USA 1995, 92, 6645–6649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dvořák, J.; McGuire, P.E. Nonstructural chromosome differentiation among wheat cultivars, with special reference to differentiation of chromosomes in related species. Genetics 1981, 97, 391–414. [Google Scholar] [PubMed]
- Metakovsky, E.; Chernakov, V.M.; Upelniek, V.P.; Redaelli, R.; Dardavet, M.; Branlard, G.; Pogna, N.E. Minor ω-gliadin-coding loci on chromosome 1A of common wheat: A revision. J. Genet. Breed. 1996, 50, 277–286. [Google Scholar]
- Dubcovsky, J.; Echaide, M.; Giancola, S.; Rousset, M.; Luo, M.-C.; Joppa, L.R.; Dvořák, J. Seed-storage-protein loci in RFLP maps of diploid, tetraploid and hexaploid wheat. Theor. Appl. Genet. 1997, 95, 1169–1180. [Google Scholar] [CrossRef] [Green Version]
- Dvořák, J.; Appels, R. Investigation of homologous crossing over and sister chromatid exchange in the wheat Nor-B2 locus coding for rRNA and Gli-B2 locus coding for gliadins. Genetics 1986, 113, 1037–1056. [Google Scholar]
- Choulet, F.; Alberti, A.; Theil, S.; Glover, N.; Barbe, V.; Daron, J.; Pingault, L.; Sourdille, P.; Couloux, A.; Paux, E.; et al. Structural and functional partitioning of bread wheat chromosome 3B. Science 2014, 345, 1249721. [Google Scholar] [CrossRef]
- Darrier, B.; Rimbert, H.; Balfourier, F.; Pingault, L.; Josselin, A.A.; Servin, B.; Navarro, J.; Choulet, F.; Paux, E.; Sourdille, P. High-resolution mapping of crossover events in the hexaploid wheat genome suggest a universal recombination mechanism. Genetics 2017, 206, 1373–1388. [Google Scholar] [CrossRef] [Green Version]
- Dvořák, J.; Yang, Z.L.; You, F.M.; Luo, M.C. Deletion polymorphism in wheat chromosome regions with contrasting recombination rates. Genetics 2004, 168, 1665–1675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thuillet, A.-C.; Bru, D.; David, J.; Roumet, P.; Santoni, S.; Sourdille, P.; Bataillon, T. Direct estimation of mutation rate for 10 microsatellite loci in durum wheat, Triticum turgidum (L.) Thell. ssp durum desf. Mol. Biol. Evol. 2002, 19, 122–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raquin, A.-L.; Depaulis, F.; Lambert, A.; Galic, N.; Brabant, P.; Goldringer, I. Experimental estimation of mutation rates in a wheat population with a gene genealogy approach. Genetics 2008, 179, 2195–2211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.Q.; Börner, A.; Röder, M.S.; Ganal, M.W. Assessing genetic diversity of wheat (Triticum aestivum L.) germplasm using microsatellite markers. Theor. Appl. Genet. 2002, 105, 699–707. [Google Scholar] [CrossRef]
- Wicker, T.; Stein, N.; Albar, L.; Feuillet, C.; Schlagenhauf, E.; Keller, B. Analysis of a contiguous 211 Kb sequence in diploid wheat (Triticum monococcum) reveals multiple mechanisms of genome evolution. Plant J. 2001, 26, 307–316. [Google Scholar] [CrossRef]
- Isidore, E.; Scherrer, B.; Chalhoub, B.; Feuillet, C.; Keller, B. Ancient haplotypes resulting from extensive molecular rearrangements in the wheat A genome have been maintained in species of three different ploidy levels. Genome Res. 2005, 15, 526–536. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar or Grain Sample 1 | Habit | Country | Year | Allele Gli-B1 | Primer | Length 2 | |
|---|---|---|---|---|---|---|---|
| 1 | Bezostaya-1 | w | Russia | 1959 | b | GliB1.1 | 369 |
| 2 | Gabo | s | Australia | 1942 | b | GliB1.1 | 369 |
| 3 | Marquis | s | Canada | 1907 | b | GliB1.1 | 369 |
| 4 | Mironovskaya-808 | w | Ukraine | 1963 | b | GliB1.1 | 369 |
| 5 | “Kavkaz” 3 | b | GliB1.1 | 369 | |||
| 6 | Federation | s | Australia | 1901 | i | GliB1.1 | 400 |
| 7 | Insignia | s | Australia | 1946 | i | GliB1.1 | 400 |
| 8 | Mentana | s | Italy | 1913 | k | GliB1.1 | 400 |
| 9 | Pane-247 | w | Spain | 1960 | k | GliB1.1 | 400 |
| 10 | Titien | w | France | 1985 | m | GliB1.1 | 400 |
| 11 | Aragon-03 | s, w | Spain | 1940 | o | GliB1.1 | 400 |
| 12 | Potam-70 | s | Mexico | 1970 | p | GliB1.1 | 400 |
| 13 | Gazul | s | Spain | 1992 | r | GliB1.1 | 400 |
| 14 | “Argelato” 3 | m | GliB1.1 | 400 | |||
| 15 | Cluj-650 | w | Rumania | 1954 | j | GliB1.1 | 400 |
| 16 | Intensivnaya | i | Kyrgyzstan | 1978 | n | GliB1.1 | 372 |
| 17 | Goelent | w | France | 1985 | q | GliB1.1 | 372 |
| 18 | Salmone | w | Italy | 1980 | s | GliB1.1 | 372 |
| 19 | Cartaya | s | Spain | l | GliB1.1 | - | |
| 20 | Chinese-Spring | s | China | - | a | GliB1.2 | 415 |
| 21 | Cajeme-71 | s | Mexico, USA | 1971 | d | GliB1.2 | 409 |
| 22 | Katepwa | s | Canada | 1981 | d | GliB1.2 | 409 |
| 23 | Laura | s | Canada | 1986 | d | GliB1.2 | 409 |
| 24 | Pavon-76 | s | Mexico | 1976 | d | GliB1.2 | 409 |
| 25 | Rinconada | s | Spain | 1981 | d | GliB1.2 | 409 |
| 26 | Roblin | s | Canada | 1986 | d | GliB1.2 | 409 |
| 27 | Suneca | s | Australia | 1981 | d | GliB1.2 | 409 |
| 28 | Yecora-S | s | Mexico | 1972 | d | GliB1.2 | 409 |
| 29 | “Inia-66” 3 | d | GliB1.2 | 409 | |||
| 30 | Diego | s | Spain | 1983 | c | GliB1.2 | 397 |
| 31 | Prinqual | s | France | 1978 | c | GliB1.2 | 397 |
| 32 | Siete-Cerros-66 | s | Mexico | 1966 | c | GliB1.2 | 397 |
| 33 | Escualo | w | Spain | 1981 | e | GliB1.2 | 397 |
| 34 | Glenlea | s | Canada | 1972 | e | GliB1.2 | 397 |
| 35 | Arminda | w | Netherlands | 1976 | f | GliB1.2 | 397 |
| 36 | Cappelle-Desprez | w | France | 1946 | f | GliB1.2 | 397 |
| 37 | Darius | w | France | 1974 | f | GliB1.2 | 397 |
| 38 | Recital | w | France | 1986 | f | GliB1.2 | 397 |
| 39 | Sideral | w | France | 1990 | f | GliB1.2 | 397 |
| 40 | “Libero” 3 | f | GliB1.2 | 397 | |||
| 41 | Argelato | s, w | Italy | 1964 | g | GliB1.2 | 397 |
| 42 | Galahad | w | UK | 1983 | g | GliB1.2 | 397 |
| 43 | Ardec | s | Belgium | 1979 | h | GliB1.2 | 402 |
| 44 | Caia | s | Portugal | - | h | GliB1.2 | 403 |
| 45 | Krasnodonka | w | Ukraine | 1969 | h | GliB1.2 | 401 |
| 46 | Newcaster | w | USA | 1946 | h | GliB1.2 | 403? 4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popovych, Y.; Chebotar, S.; Melnik, V.; Rodriguez-Quijano, M.; Pascual, L.; Rogers, W.J.; Metakovsky, E. Congruity of the Polymorphisms in the Expressed and Noncoding Parts of the Gli-B1 Locus in Common Wheat. Agronomy 2020, 10, 1510. https://doi.org/10.3390/agronomy10101510
Popovych Y, Chebotar S, Melnik V, Rodriguez-Quijano M, Pascual L, Rogers WJ, Metakovsky E. Congruity of the Polymorphisms in the Expressed and Noncoding Parts of the Gli-B1 Locus in Common Wheat. Agronomy. 2020; 10(10):1510. https://doi.org/10.3390/agronomy10101510
Chicago/Turabian StylePopovych, Yulia, Sabina Chebotar, Viktor Melnik, Marta Rodriguez-Quijano, Laura Pascual, William John Rogers, and Eugene Metakovsky. 2020. "Congruity of the Polymorphisms in the Expressed and Noncoding Parts of the Gli-B1 Locus in Common Wheat" Agronomy 10, no. 10: 1510. https://doi.org/10.3390/agronomy10101510