
Molecular Characterization of Three Chemosensory Proteins from Carposina sasakii

Research Institute of Pomology, Chinese Academy of Agricultural Sciences, 98 South Street, Xinghai, Huludao 125100, China
*
Author to whom correspondence should be addressed.
Agriculture 2023, 13(11), 2066; https://doi.org/10.3390/agriculture13112066
Submission received: 5 September 2023
/
Revised: 24 October 2023
/
Accepted: 25 October 2023
/
Published: 27 October 2023
(This article belongs to the Special Issue Advances in Integrated Pest Management Strategies)
Abstract
:The peach fruit moth, Carposina sasakii, is part of the Carposinidae, and is harmful to the families Rosaceae and Rhamnaceae. C. sasakii lays eggs on the hairy surface of the fruit’s stalk cavity and calyx end. After hatching, the moth can bore into the fruits and feed on the flesh inside. Chemosensory proteins (CSPs) are a class of low-molecular-weight soluble carrier proteins that are highly evolutionarily conserved. To enhance our understanding of the recognition of host plant volatiles by CSPs of C. sasakii, the expression patterns and binding characteristics of CsasCSP7, CsasCSP9 and CsasCSP11 in C. sasakii were investigated. In our study, the results of real-time quantitative polymerase chain reaction (qPCR) assays demonstrate that CsasCSP7 and CsasCSP9 transcripts were abundantly expressed in the antennae of males, and CsasCSP11 was highly expressed in the wings of females. Fluorescence competitive binding assays with 38 candidate ligands showed that CsasCSP7 could bind to benzaldehyde and dodecanal, whereas CsasCSP9 bound to butyl octanoate, decanal and (-)-beta-pinene. CsasCSP11 could also bind to1-hexanol, beta-ocimene and 6-methyl-5-hepten-2-one. Our results suggest that CsasCSP7, CsasCSP9 and CsasCSP11 may play a crucial role in locating the host plant of C. sasakii.
1. Introduction
The peach fruit moth, Carposina sasakii, is one of the Carposinidae, and is widely distributed in North Korea, South Korea, Japan, the Russian Far East and China. It is harmful to apple, jujube, pear and other fruit trees from the families Rosaceae and Rhamnaceae [1]. Female adult C. sasakii prefers to lay eggs on the surface of the fruit, especially in the calyx pits of apples. In the case of peach fruits, which are covered with fine hairs, the insect lays eggs all over the fruit surface [2]. After hatching, C. sasakii can bore into the fruits and feed on the flesh inside, making it difficult to reduce larvae numbers using chemicals and causing losses in fruit production [3]. Preventing oviposition could be one of the most effective methods of controlling C. sasakii. The adult females release a sex pheromone composed of two odorants, (Z)-7-eicosen-11-one and (Z)-7-nonadecen-11-one, which attracts adult males for mating, and the pheromone has been used to trap adult males for controlling and monitoring the C. sasakii population. However, it is not sufficiently effective for mating disruption [4]. Overall, more efficacious management processes are urgently required.
Insects communicate with their environment through olfaction when detecting food and habitats, as well as when finding mates and escaping predators. To successfully perform such behaviors, insects must respond to chemical stimulation at the right moment. Insects’ olfactory systems are modified depending on age, feeding state, circadian rhythm, and mating status. Disrupting the chemical communication system as a method to disrupt mating is a possible novel, ecologically friendly, and environmentally friendly approach for pest control [5].
The sex pheromone of Bombyx mori was identified first, and from then on, research into pest olfactory systems has developed rapidly. Pest behavior regulation techniques based on the olfactory system are internationally recognized as a green plant protection technology. Compared to other techniques, olfactory behavior regulation technology has five characteristics: safety, selectivity, efficiency, effectiveness and compatibility [6]. There are two important types of compounds used as part of the technology: one relates to the volatile organic compounds released by plants, and the other is the sex pheromones released by insects. Both of them have several advantages. First, pheromones are natural compounds that are generally used in very small quantities (billionth of a unit). Second, pheromones break down relatively quickly in the environment (they do not leave residues). Third, they are highly specific—they do not act on non-target organisms. Fourth, they allow the rapid detection of insects in the field, aiding agricultural decision-making [7].
Plant volatile organic compounds (VOCs) are major vehicles of information transfer between organisms, and they mediate many ecological interactions. Several herbivorous insects utilize plant chemical cues to identify hosts for feeding. Given the multifunctional nature of induced VOC emissions, it is safe to assume that, with changes in the selective environment, the major function of an information-mediating trait, such as VOC emission, will shift [8]. Several insects employ plant volatile organic compounds as olfactory cues when seeking a host plant, enabling them to discriminate between host plants and nonhost genotypes. Herbivorous insects can search for their hosts by plant volatiles, and the natural enemies of herbivores can find their prey by plant volatiles. Fruit-feeding insects are, logically, attracted to fruit volatiles. For these insects, fruits are not only a nutritional resource but also ideal mating sites. Blends of fruit volatiles indicate mating opportunities and may stimulate calling (pheromone release) and mating behaviors [9]. From an evolutionary perspective, there is a strong selection pressure on insects to develop sophisticated means for detecting and locating food resources upon which survival and reproduction depend [10].
In most species of moths, the sex pheromone is a blend of several components comprising fatty acid derivatives, usually alcohols, aldehydes, or acetates connected by a C10-C18 straight chain [11], and is produced and secreted by the female pheromone glands (PGs) located between the eighth and ninth abdominal segments. Sex pheromones induce sexual behavior in insects. A male insect may smell the sex pheromone released by the female and then attempt to copulate with her [12]. To date, sex pheromones, especially female-produced sex attractants, have been the most widely used semiochemicals in attract-and-kill techniques. They are selective, often attracting only the target species [13].
The perception of these compounds depends on the olfactory receptor neurones (ORNs) in sensillae, mostly found on the insects’ antennae, which can recognize individual molecular structures [14]. After odor compounds enter the sensilla via the numerous pores on its surface, these molecules encounter an aqueous barrier, the sensillum lymph [11]. In the lymph, there are many protein families that determine the operation of the odor compounds. Chemosensory proteins (CSPs) are one of them.
CSPs were first reported in Drosophila melanogaster and were identified as olfactory segment D (OS-D) at the time [15]. Antennal protein 10 was also found in Drosophila melanogaster. Subsequently, proteins with close similarities to OS-D were also found on the antennae of several stick insects (identity 30–45%) [16,17,18]. A protein with a 47% structural similarity to OS-D, named CLP-1, was found on the lips of Cactoblastis cactorum, and this protein is expressed in females and not in males [19]. Similar proteins have also been found in Anopheles gambiae, referred to as sensory appendage proteins (SAPs) [20,21]. CSPs were officially named in 1999, and seven soluble proteins with structural similarities to OS-D were discovered in the antennae of Schistocerca gregaria [22].
CSP is highly conserved in evolution, with 40–50% identical residues between species with more orthologous phylogenetic development compared to odorant-binding proteins (OBPs), which are only conserved at 10–15% [23]. The cause may be related to the disulfide bond. In OBP, where three disulfide bonds bind to each other to keep the protein stable, the effect of residue substitution is very limited, while in CSP, two disulfide bonds are connected to adjacent cysteines, and conservative residue sequences are folded to form hydrophobic pockets and maintain protein stability [24].
We have evaluated the relative expression levels of CSPs from different tissues in C. sasakii, and the results showed that CsasCSP7 and CsasCSP9 are highly expressed in the antennae of the male and female moth, while CsasCSP11 is extremely highly expressed in the wings. In order to better understand the function of CsasCSPs in the olfactory communication system, we analyzed the characteristics of the CsasCSPs sequence, constructed the recombinant expression vector of CsasCSPs in a prokaryotic expression system, and tested the binding affinities of CsasCSPs to volatile molecules through fluorescence competitive binding assays. Our study enriches our understanding of the functions of CSPs from C. sasakii and complex olfactory mechanisms in insects.
2. Materials and Methods
2.1. Insect Rearing and Sample Collection
The larvae of C. sasakii were collected from the apple orchard of the Institute of Pomology, Chinese Academy of Agricultural Sciences (CAAS), Liaoning province (latitude 40.61° N, longitude 120.73° E), China. The pests were maintained under constant conditions of 25 ± 1 °C, 70 ± 5% relative humidity, and a 15:9 (L:D) h photoperiod. They were fed a 10% honey solution. C sasakii commenced eclosion after 4:00 p.m., and the virgin moths mated after 9:00 p.m. on the same day. Therefore, we infer that C. sasakii reached sexual maturity on the day of the eclosion [25]. We separately dissected 400, 100, 100, 100, 100, and 100 1-day-old adults to get different tissues of antennae (200 males and 200 females), heads without antennae (20 males and 20 females, respectively), thoraxes and abdomens (10 males and 10 females, respectively), legs, and wings (50 males and 50 females, respectively). The samples were placed in RNase-free centrifuge tubes under an optical microscope. Three biological replicates were prepared. All samples were immediately immersed in liquid nitrogen and stored at −80 °C.
2.2. Total RNA Extraction and cDNA Synthesis
The total RNA of different tissue samples was extracted using a Trizol Reagent handbook (TAKARA BIO Inc., Shiga, Japan). The integrity and concentration of RNA were assessed via 1.5% agarose gel electrophoresis and an ultra-microspectrophotometer, and the first-strand cDNA was synthesized with 1 μg of total RNA using a PrimeScript RT reagent Kit with a gDNA eraser (TAKARA BIO Inc., Shiga, Japan) according to the manufacturer’s instructions. This was then subjected to PCR and q-PCR.
2.3. cDNA Cloning and Sequence Analysis
PCR assays were performed in a mixture of 40 μL containing 4 μL of antennae cDNA (100 ng), 20 μL of Primer fix Kit, 2 μL of each primer (10 μM), and 12 μL of RNase-Free Water. The PCR amplification procedure was as follows: predenaturation at 95 °C for 3 min; 35 cycles of 95 °C for 30 s; 55 °C for 30 s; 72 °C for 45 s; and final extension at 72 °C for 10 min. The PCR product was visualized using 1% agarose gel electrophoresis; the recovered target gene was then ligated into the pMD19-T vector (TAKARA BIO Inc., Shiga, Japan) and then transformed into DH5α competent cells (TAKARA BIO Inc., Shiga, Japan) for splicing sequencing (BGI Genomics, Beijing, China).
The open reading frames (ORFs) for these sequences were found using the ORF Finder (http://www.ncbi.nlm.nih.gov/gorf/gorf.html, accessed on 9 March 2022). The signal peptides of CsasCSPs were predicted using the SignalP 5.0 Server (http://www.cbs.dtu.dk/services/SignalP/, accessed on 9 March 2022) [26]. The molecular weight was computed using ProtParam (http://web.expasy.org/protparam/, accessed on 1 May 2022). The isoelectric point was also calculated using the same software. The C. sasakii CSPs were chosen for phylogenetic analysis along with CSPs from Lepidoptera, Diptera, Hemiptera and Coleoptera, which were downloaded from NCBI (http://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 1 October 2021). DNAMAN (Lynnon Biosoft, San Ramon, CA, USA) was used for multiple sequence alignment. A phylogenetic tree was constructed in MEGA7.0 using the neighbor-joining method with 1000 bootstrap replicates and clarified with iTOL (https://itol.embl.de/, accessed on 10 October 2021).
2.4. Expression Levels of Three CSPs mRNA from C. sasakii
The RT-qPCR assay was carried out using the CFX96 Connect Real-Time Detection System produced by BioRad in the USA using β-actin and elongation factor 1α (EF-1α) as the endogenous gene, the cDNA obtained as above as the template, and the female head (without antennae) as a control. In the experiment, RT-qPCR primers were designed using Beacon Design 8.0 (Table 1), and the amplification efficiencies were 90–110%. Three biological replicates were performed, and two technical replicates were also performed. The specific operation is detailed below.
Reaction system: Each RT-qPCR reaction was conducted in a 20 μL reaction mixture containing 10 μL of SYBR Master Mix, 1 μL of sample cDNA, 0.5 μL of each primer (10 µmol/L), and 8 μL of sterilized ddH2O. Reaction procedure: denaturation at 95 °C for 3 min; 40 cycles of 95 °C for 10 s, 55 °C for 30 s, and 72 °C for 30 s.
After the program, the Ct values of the endogenous and target genes were obtained. According to the endogenous gene, the relative expression of the target gene was calculated from the 2−ΔΔCt. A significant difference analysis of the target genes between different tissues was undertaken using a one-way analysis of variance (ANOVA), and then Tukey’s honestly significant difference (p < 0.05) method was employed for multiple comparisons. These were implemented through the SPSS Statistics 19.0 software.
2.5. Expression and Purification of Recombinant CsasCSPs Proteins
The target gene was amplified through adult antennae cDNA, and primers were designed using Primer Premier 5 (Table 1). The PCR product was ligated to the pMD-19T vector, then to the pET-28a vector (EcoRⅠ, XhoⅠ). It was then transformed into BL21 (DE3) competent cells [3]. After positive verification had been performed, the bacterial solution was cultured in 500 mL of liquid Luria–Bertani (LB) medium containing 50 μg/mL of kanamycin at 37 °C. When the OD600 reached 0.6–0.8, the recombinant protein was induced at 37 °C for 10 h using isopropyl β-d-1-thiogalactopyranoside (IPTG) at a final concentration of 1 mM. The cells were harvested via centrifugation at 12,000 rpm for 10 min, suspended in lysis buffer (80 mM Tris-HCl, 200 mM NaCl, 1 mM EDTA, 4% glycerol, pH 7.2 and 0.5 mM PMSF), sonicated in ice for 3 s, over five passes, and then centrifuged again. The 15% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) assay showed that these proteins persisted in the inclusion body. A refolding inclusion body assessment was conducted via the redox method. The proteins were concentrated in the 10 kDa and 30 kDa ultrafiltration cube. Before the 15% SDS-PAGE analysis, the solutions containing the proteins were dialyzed using 500 mL of Tris-HCl buffer at a final concentration of 20 mM three times at 4 °C.
2.6. Fluorescence Competitive Binding Assay
Thirty-six common volatile components and two sex pheromones from C. sasakii were chosen as ligands for use in the fluorescence competitive binding assays (Table 2). Fluorescence binding assays were performed using a multimode reader. The fluorescent emission spectra of the probe, 1-Aminoanthraquinone (1-AMA, Sigma–Aldrich, St. Louis, MO, USA), were recorded in the range of 400 to 750 nm with an excitation wavelength of 260 nm. To measure the affinity of 1-AMA to each protein, 2 μM of protein in 50 mM Tris–HCl (pH 7.2) was titrated with 1 mM 1-AMA to final concentrations ranging from 2 to 80 μM. Dissociation constants for the protein/1-AMA complex were inferred from Scatchard plots of the binding data. Ligands with concentrations ranging from 1 to 30 µM were added to the mixtures of protein and probe at the same final concentration of 2 mM. The dissociation constants of the competitors were calculated from their corresponding IC50 values (concentrations of ligands halving the initial fluorescence value of 1-AMA), using the equation Ki = [IC50]/ (1 + [1-AMA]/K1-AMA), where [1-AMA] is the free concentration of 1-AMA, [IC50] is the ligand concentration displacing 50% of the fluorescent reporter, and K1-AMA is the dissociation constant of the complex protein/1-AMA. All values were obtained from three independent measurements. The protein with 100% activity was assumed, inferred via the stoichiometry assessment of a 1:1 ratio of protein/ligand, and the data were analyzed as above.
3. Results
3.1. Sequence Analysis of CsasCSPs
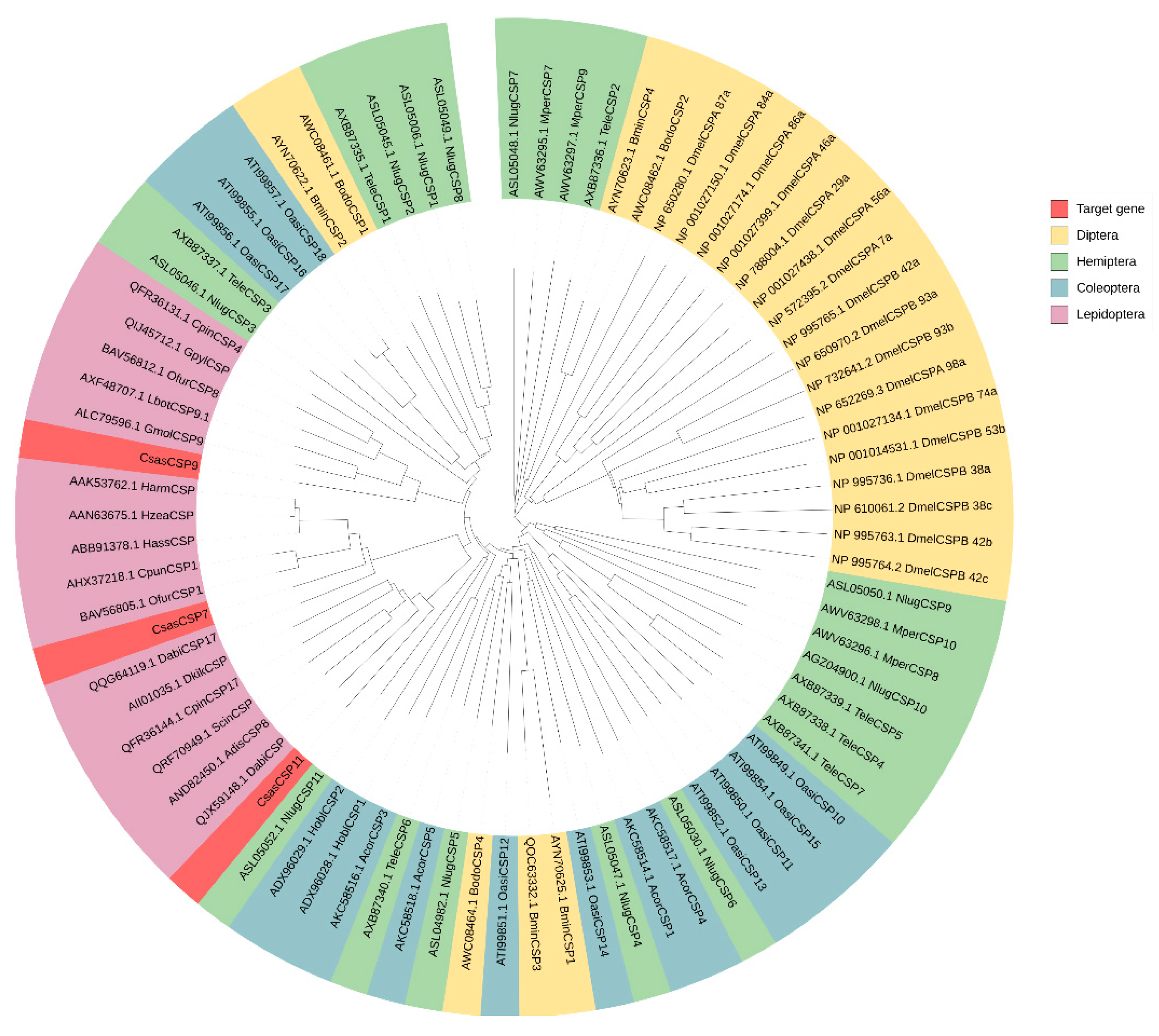
The length of the open reading frame (ORF) of CsasCSP7, CsasCSP9 and CsasCSP11 was 387-bp, 432-bp and 402-bp, respectively. This encoded 128 amino acids, 143 amino acids and 133 amino acids, respectively. At the N-terminus of the polypeptide chain, CsasCSP7 was predicted to contain a signal peptide consisting of 19 amino acid residues. CsasCSP9 was predicted to contain a signal peptide consisting of 15 amino acid residues. CsasCSP11 was predicted to contain a signal peptide consisting of 24 amino acid residues. As the signal peptide was cleaved off, the MW of the mature protein CsasCSP7 was 13.25 kDa with an isoelectric point of 5.36. The predicted MW of the CsasCSP9 protein was 14.54 kDa, and the isoelectric point was 4.93. The predicted MV of the CsasCSP11 protein was 13.15kDa, and the isoelectric point was 8.53. The results of multiple alignments show that CsasCSP7, CsasCSP9 and CsasCSP11 presented a typical four-cysteine signature, forming two pairs of disulfide bonds (Figure 1). To analyze the phylogenetic relationships of CsasCSPs with other insects, including Lepidoptera, Diptera, Hemiptera and Coleoptera, a phylogenetic tree was constructed. The results clearly show that CsasCSPs were located in the Lepidoptera group. Compared to other orders, Lepidoptera were explicitly clustered together (Figure 2).
3.2. Tissue Expression Patterns of CsasCSPs
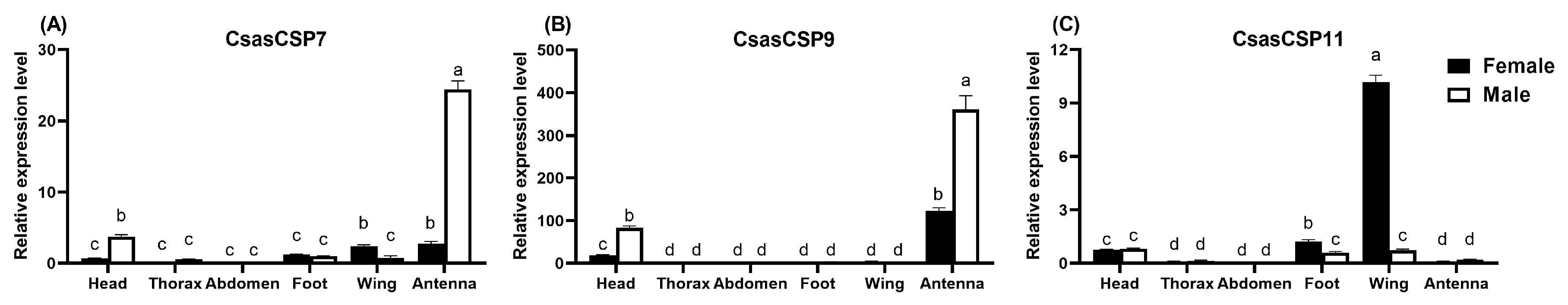
To better understand the function of CsasCSPs, their expression patterns in different tissues were measured via RT-qPCR. The expression analysis revealed similar expression patterns of CsasCSP9 in both males and females. CsasCSP9 was predominantly expressed in the antennae of both sexes, with the lowest expression observed in the other tissues. CsasCSP7 and CsasCSP11 showed broad expression profiles in the whole body of male and female adults, but CsasCSP7 was predominantly expressed in the male antennae, and CsasCSP11 was expressed at a significantly higher level in female wings (Figure 3).
3.3. Bacterial Expression and Purification of CsasCSPs
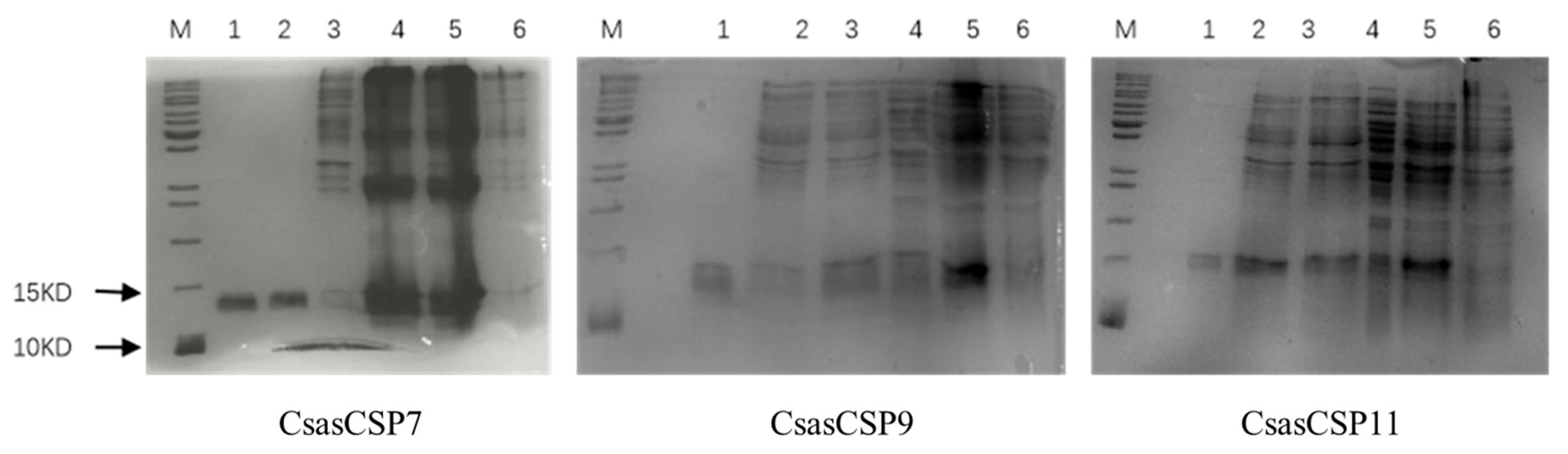
The pET-28a (+)/CsasCSPs proteins were assessed for expression in vitro (Escherichia coli BL21 (DE3) cells), and the target proteins were found more abundantly in the insoluble fraction containing inclusion bodies compared to the supernatant when induced with IPTG via 15% SDS-PAGE. The 15% SDS-PAGE analysis revealed that the final purified proteins were present in single bands of approximately 13.25 kDa, 14.54 kDa and 13.15 kDa, as is consistent with the MW of the recombinant proteins (Figure 4).
3.4. Binding Properties of Recombinant CsasCSPs
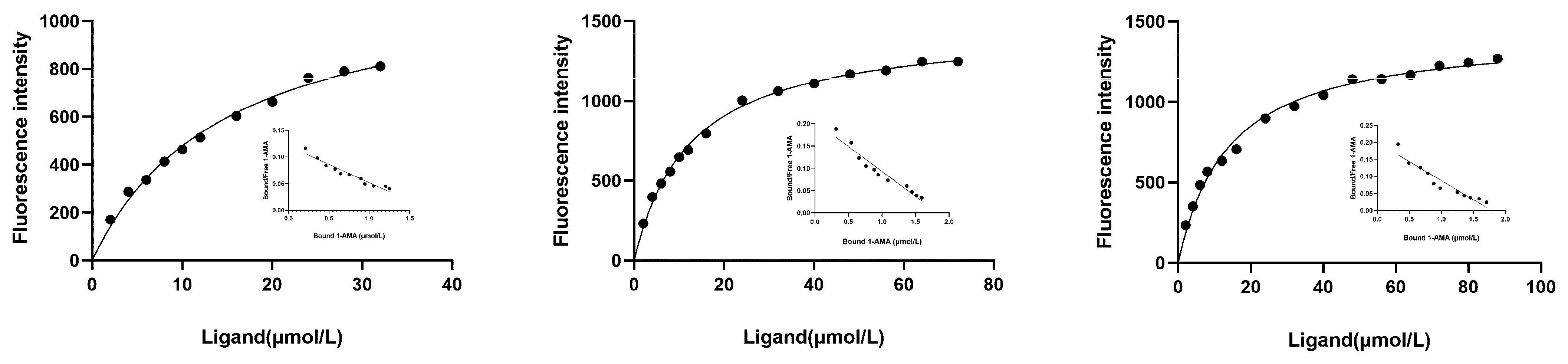
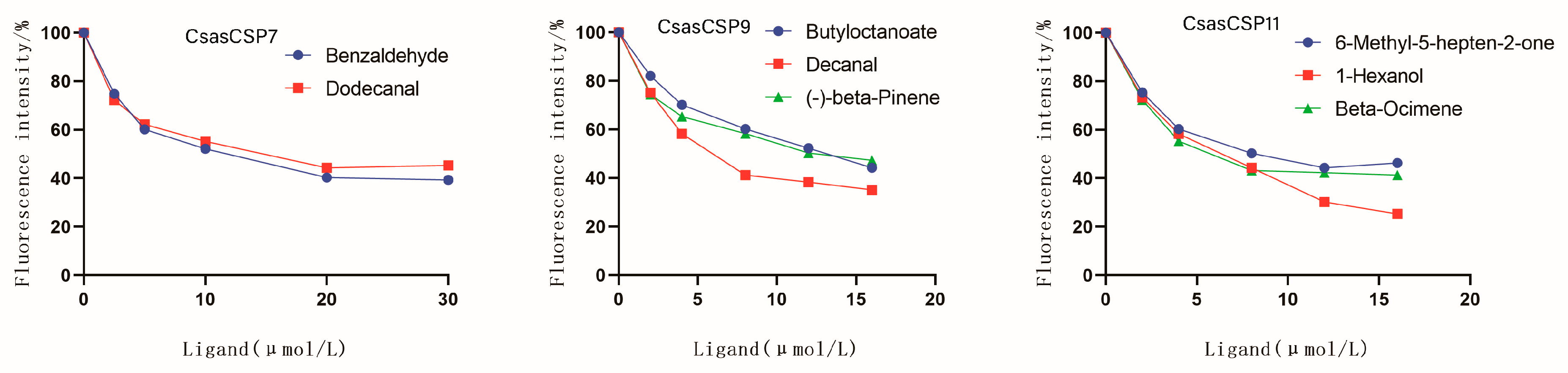
For the further characterization of CsasCSPs ligand-binding affinity to host volatiles, 1-AMA was used as a competitive fluorescent reporter. When 1-AMA was added dropwise to the protein solution, the maximum emission peak at 420 nm was shifted to approximately 560 nm. CsasCSP7, CsasCSP9 and CsasCSP11 bind to the fluorescent probe 1-AMA rather weakly, with dissociation constants of 20.10, 12.39 and 13.83 μM, respectively, yielded by the Scatchard equation (Figure 5). Therefore, the Ki (the ratio of the dissociation constant of the competitor to the partition coefficient of the competitor in the aqueous phase) and IC50 (the concentration of the competitor when the fluorescence intensity of the complex protein/1-AMA declined to 50% of the initial fluorescence) values of the ligands with CsasCSPs are presented in Table 3, Table 4 and Table 5. Compounds that we tested that reduced the fluorescence intensity of the complex systems to 50% or lower were regarded as binding ligands. This group of chemicals includes two sex pheromone components and 36 plant volatiles. Two volatiles, benzaldehyde (Ki = 7.25 ± 0.23 µM) and dodecanal (Ki = 13.61 ± 0.54 µM), were tested for use as potential ligands for CsasCSP7. CsasCSP9 showed high capacities for binding to three chemical volatiles, namely, butyl octanoate (Ki = 1.47 ± 0.13 µM), decanal (Ki = 1.65 ± 0.31 µM) and (-)-beta-pinene (Ki = 14.26 ± 0.62 µM). Three of the ligands tested, 1-hexanol (Ki = 8.13 ± 0.78 µM), beta-ocimene (Ki = 1.83 ± 0.66 µM) and 6-methyl-5-hepten-2-one (Ki = 0.71 ± 0.07 µM), exhibited relatively strong binding affinities for CsasCSP11. Besides this, the CsasCSPs showed no affinity for the components of sex pheromones (Figure 6).
4. Discussion
In this work, we cloned and sequenced the cDNA of CsasCSP7, CasaCSP9 and CasaCSP11, and analyzed the signal peptides, isoelectric points, protein molecular weights and protein properties of the CsasCSP7, CasaCSP9, and CasaCSP11 proteins. They all showed four conserved cysteines and two disulfide bonds, as is typical of the CSP family.
CSP is distributed in various parts of an insect’s body, such as the antennae [27,28], legs [29], abdomen [30], midguts [31], sensillum lymph [32] and wings [33]. There have also been reports showing that CSP is expressed differently at different times [34]. Before now, many studies have shown that CSPs have many different functions.
CSPMbraA and CSPMbraB, expressed in the antennae and pheromone glands, were thought to be involved in the transport of hydrophobic molecules through different aqueous media, such as the sensillar lymph and the pheromonal gland cytosol, in Mamestra brassicae [35,36]. LmigCSP91 was found to be expressed in the male organs of Locusta migratoria, but after mating, it was also found in female organs [37]. This was also the case for Apis mellifera AmelCSP3 [38]. It can be speculated that these proteins are key to the transportation of pheromones.
CSP is also thought to be closely related to the reproduction, regeneration and development of insects. Protein p10 was found in the regenerated legs of Periplaneta americana larvae at levels 30 times higher than in normal legs. Fluorescent immunology showed that p10 was present only in a newly formed epidermis [39,40]. Apis mellifera AmelCSP5 was specifically expressed only in the gonads of female adults. When the gene encoding AmelCSP5 was silenced, eggs did not fully develop and did not hatch. This protein has been shown to play a vital role in the development of eggs [41,42]. Solenopsis invicta SinvCSP9 and AmelCSP5 belong to the same evolutionary branch. Gene silencing not only affected the synthesis of fatty acids, but also the development and molting of the stratum corneum [43]. The HamCSP4 of Helicoverpa armigera was expressed in large quantities in the mouthparts, and has been verified as a surfactant, suggesting that it may be involved in dissolving the terpenoids in nectar, given the affinity of HarmCSP4 with terpenoids [44]. LmigCSP IIII (Lmig EST6) in Locusta migratoria exhibited a strong binding capacity with the non-host plant compound α-amyl cinnamaldehyde (AMCAL). AMCAL has been shown to be an effective food repellent for use against locusts. Locusts were less sensitive to this compound after LmicsP IIII had been knocked out [45]. Bemisia tabaci BtabCSP11 was highly expressed in the abdomen. After it was silenced, the fertility of female whiteflies decreased significantly, indicating that it may be involved in their reproduction [46].
In our study, the results show that CasaCSP7 and CasaCSP9 were highly expressed in antennae, and CasaCSP11 was highly expressed in the wings. Mythimna separata MsepCSP5 was mainly expressed in the antennae compared to other tissues, and combined with some rice volatiles, suggesting its potential major role in chemodetection [47]. Monochamus alternatus MaltCSP5 was also reported to be expressed in the antennae of both male and female adults, and the protein showed a high binding activity to most pine volatiles [48]. Apolygus lucorum AlucCSP2 and AlucCSP3 were specifically expressed in female wings. AlucCSP4 was expressed relatively highly in female wings but was also expressed in other tissues. Interestingly, the capacity of the three proteins to bind to cotton secondary metabolites is better than their capacity to bind to plant volatiles [33]. Therefore, it is hypothesized that not only the antennae-predominant CasaCSP7 and CasaCSP9, but also the wing-predominant CasaCSP11, may both bind to host volatiles during olfactory recognition.
Ligand-binding experiments have demonstrated that CsasCSPs undergo highly selective binding to volatile compounds, including benzaldehyde, dodecanal, butyl octanoate, decanal, (-)-beta-pinene, 1-hexanol, beta-ocimene and 6-methyl-5-hepten-2-one. In previous studies, benzaldehyde, decanal, 6-methyl-5-hepten-2-one and β-pinene were identified as promoting aggregation behavior in insects [49,50,51,52].
Benzaldehyde is not only very common, but also a predominant component of the floral scents of many species [53]. In single sensillum recording experiments on both male and female Sitona humeralis antennae, olfactory sensory neurons (OSN) were stimulated successfully using benzaldehyde in a clear dose-dependent manner. Field experiments revealed that traps baited with benzaldehyde caught significantly more S. humeralis than unbaited traps, indicating an attractant effect. There were no apparent differences between the sexes [54]. Meanwhile, behavioral experiments were conducted on Spodoptera littoralis, and the results showed that the larvae were attracted to benzaldehyde at 100 and 10 μg/μL [55]. Rhopalosiphum padi males responded positively to benzaldehyde. The release of benzaldehyde with a conspecific sex pheromone increased the catch rates of both species of aphid [56]. Trapping tests undertaken in the field also showed that benzaldehyde has a lure effect on Conotrachelus nenuphar [57]. In 2015, an article confirmed that benzaldehyde has insecticidal activity against Galleria mellonella (100% insect mortality) [58]. All this suggests that benzaldehyde can be used in the development of new insecticides.
A strong binding affinity between CsasCSP7 and dodecanal was also shown. It has been shown that MsepCSP5 is abundantly expressed in the antennae of adult females and selectively recognizes dodecanal [47]. The dodecanal identified in the headspace of acidified chicken feces elicited electroantennogram responses from the antennae of Culex quinquefasciatus females [59]. The dodecanal of 50 and 100 ppm exhibited oviposition-deterring activities against gravid Aedes aegypti females. [60]. Olfactometer experiments with synthetic chemical compounds revealed the significant attraction of Bactrocera dorsalis female flies to dodecanal [61]. In addition, dodecanal was considered a sex pheromone because it was identified on extracts of filter paper contaminated by young Cephalonomia tarsalis females. It also had an arresting effect on males, but not on females [62]. CsasCSP7 showed sensitivities to benzaldehyde and dodecanal, suggesting that CsasCSP7 plays an important role in the identification of host plants in the olfactory communication system.
CsasCSP9 showed strong capacities for binding to three volatiles, namely, butyl octanoate, decanal and (-)-beta-pinene. The electroantennographic results demonstrate that decanal elicited significant antennal responses in Quadrastichus mendeli. Bioassays confirmed that Q. mendeli was repelled by decanal [63]. On the other hand, wind tunnel experiments showed that decanal attracted male Lutzomyia longipalpis [64]. A pure compound of decanal and 0.03 ppm decanal were also reported as attractants for Heterorhabditis bacteriophora and Steinernema kraussei, respectively [65]. Compared with normal Sitobion avenae SaveOBP9, in experiments with SaveOBP9 knockdown, wheat aphids showed significantly decreased attractive responses toward decanal [66]. Similarly, after silencing LmigOBP1 in Locusta migratoria, the nymphs showed significantly decreased electroantennography (EAG) responses to decanal [67]. In GC-EAD, the antennae of gravid female Ostrinia nubilalis responded to decanal, which is an established maize volatile [68]. Interestingly, Morawo et al. found that decanal elicited 0.82 mV EAG response in Microplitis croceipes, which was the highest response value. However, decanal accounts for only 1% of cotton-fed host volatiles [69].
β-pinene is a ubiquitous plant terpenoid and elicits host recognition behaviors in female Conogethes punctiferalis [70]. Under four-arm olfactometer bioassays, the Porphyrophora sophorae larvae showed a clear preference for β-pinene, and β-pinene proved to be the major chemical cue used by P. sphorae neonates when searching for the roots of their host plant [71]. In a series of novel hydronopylformamides derivatives synthesized from a naturally occurring compound, (-)-β-pinene, four exhibited repellency against Blattella germanica at a concentration of 20 mg/mL [72]. CsasCSP9 had a binding affinity to butyl octanoate, decanal and (-)-beta-pinene, suggesting that CsasCSP9 can transport host volatiles in the olfactory communication system.
The compound 6-methyl-5-hepten-2-one is released by animals, is also a major component identified from flower scents and may play an important role outside animal-host seeking [73]. In Triatoma dimidiate, both females and males were attracted to 6-methyl-5-hepten-2-one [74]. Diaziella yangi and Lipothymus sp enter Ficus curtipes in response to the body odors of obligate wasps and one of the main compounds emitted by figs—6-methyl-5-hepten-2-one [75]. 6-methyl-5-hepten-2-one, which has been identified in zebra skin, was one of the three ketones (acetophenone and geranylacetone) that reduced the catch levels of Glossina pallidipes in field trials [76]. The mixture of 6-methyl-5-hepten-2-one and geranylacetone in a 1:1 ratio increased the repellency of DEET to Anopheles gambiae, Culex quinquefasciatus and Aedes aegypti when present at low concentrations. In olfactometer trials, 6-methyl-5-hepten-2-one interfered with the attraction of mosquitoes to a host [77]. A mixture of two compounds, 6-methyl-5-hepten-2-one and geranylacetone, showed significant repellency towards Culicoides impunctatus in the field [78]. The electrophysiological testing of 6-methyl-5-hepten-2-one using five fly species (Musca autumnalis, Haematobia irritans, Hydrotaea irritans, Stomoxys calcitrans and Wohlfahrtia magnifica) showed that it was physiologically active towards the flies tested. At certain concentrations, 6-methyl-5-hepten-2-one increased upwind flight in flies and reduced fly loads [79]. As such, 6-methyl-5-hepten-2-one may have the potential to be developed into novel repellents. Meanwhile, volatile characterization with gas chromatography–mass spectrometry identified the contents of Gelis agilis emissions as 6-methyl-5-hepten-2-one, a known insect defense semiochemical that acts as an alarm pheromone in ants [80].
β-ocimene is a key plant volatile with multiple relevant functions in plants, depending on the organ and the time of emission. AmelCSP4 has the best affinity with β-ocimene. In AmelCSP4, Tyr98 and Asp67 are involved in β-ocimene binding [81]. Tea plants infested with Ectropis obliqua larvae triggered neighboring plants to release β-ocimene, which repels E. obliqua adults, especially mated females. The levels of β-ocimene released by infested tea plants increased rapidly [82]. Pest-infested plants (Silene latifolia) emitted higher amounts of β-ocimene [83], as did Morus alba [84]. Compared with healthy plants, the treatment of Chinese cabbage with β-ocimene inhibited the growth and reproduction of Myzus persicae, and we saw that winged aphids preferred to stay on the healthy plants. Aphidius gifuensis, however, was shown to prefer cabbage treated with β-ocimene [85]. Similar results have been reported elsewhere [86]. In male Hyphantria cunea, when β-ocimene was added to a sex pheromone, the efficiency of trapping male moths increased [84]. Subtractive bioassays performed in a dual-choice olfactometer showed that a 3-component terpenoid plant-derived blend comprising (E)-linalool oxide, β-pinene and β-ocimene was more attractive to female Anopheles gambiae [87].
In a study on Adelphocoris lineolatus, 1-hexanol was identified as the sex pheromone antagonist [88]. However, further research found that AlinOBP10 had a higher binding affinity for 1-hexanol. In Y-tube olfactometer trials, 1-hexanol was repellent against female adults [89]. In a feeding test of Ips typographus, high concentrations of 1-hexanol solution were shown to cause food rejection (AFI = 1.00), and males were found to be more sensitive than females [90]. In Grapholita molesta, recombinant GmolCSP8 (rGmolCSP8) also showed the strongest binding affinity to 1-hexanol. In our research, CsasCSP11 showed sensitivities to these three compounds, providing evidence that CsasCSP11 may be involved in the detection of plant volatiles.
5. Conclusions
Two of the three genes selected were highly expressed in male antennae. Unfortunately, the two genes showed nonsignificant binding affinities for sex pheromones of C. sasakii, but were able to bind to plant volatiles. Different compounds will produce different results in different insects. The results for some of these compounds are consistent with our experimental results, while others are in opposition, and these compounds have attractive/repellent effects on insects. In conclusion, our results show that CsasCSP7, CsasCSP9 and CsasCSP11 may have a dual role in host-seeking. The characterization and function of CsasCSP7, CsasCSP9 and CsasCSP11 from C. sasakii contributes to our understanding of the underlying mechanisms of olfactory communication in insects.
Author Contributions
Conceptualization, L.S. and G.Q.; methodology, L.S. and G.Q.; software, L.L. and Q.Y.; validation, W.Y. and L.S.; formal analysis, L.L.; investigation, L.S. and G.Q.; resources, L.S. and G.Q.; data curation, L.S. and L.L.; writing—original draft preparation, L.L.; writing—review and editing, L.L. and L.S.; visualization, L.L., H.Z. and Q.Y.; supervision, L.S.; project administration, L.S. and G.Q.; funding acquisition, L.S. and G.Q. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Applied Basic Research Project of Liaoning Province (2022JH2/101300192), the Agricultural Science and Technology Innovation Program (CAAS-ASTIP-2021-RIP-05) and the National Natural Science Funds (31601643).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- Wu, Y.P.; Zhao, J.L.; Su, T.J.; He, Q.S.; Xie, J.L.; Zhu, C.D. The complete mitochondrial genome of Carposina sasakii (Lepidoptera: Carposinidae). Mitochondrial DNA 2016, 27, 1432–1434. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.J.; Song, W.; Yue, L.; Guo, S.K.; Chen, J.C.; Gong, Y.J.; Hoffmann, A.A.; Wei, S.J. Chromosome-level genome of the peach fruit moth Carposina sasakii (Lepidoptera: Carposinidae) provides a resource for evolutionary studies on moths. Mol. Ecol. Resour. 2021, 21, 834–848. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.Q.; Qiu, G.S.; Li, Y.Y.; Zhang, H.J.; Yan, W.T.; Yue, Q.; Sun, L. N Molecular characterization and functional analysis of pheromone binding proteins and general odorant binding proteins from Carposina sasakii matsumura (Lepidoptera: Carposinidae). Pest. Manag. Sci. 2019, 75, 234–245. [Google Scholar] [CrossRef] [PubMed]
- Han, K.S.; Jin, K.J.; Choi, K.H.; Sun, W.L.; Boo, K.S. Sex pheromone composition and male trapping of the peach fruit moth, Carposina sasakii (matsumura) (Lepidoptera: Carposinidae) in korea. J. Asia-Pac. Entomol. 2000, 3, 83–88. [Google Scholar] [CrossRef]
- Gadenne, C.; Barrozo, R.B.; Anton, S. Plasticity in insect olfaction: To smell or not to smell? Annu. Rev. Entomol. 2016, 61, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Liu, Y.; Wang, B.; Wang, G.R. Research status, opportunities and challenges of pest olfactory behavior regulation technology. Chin. Sci. Found. 2020, 4, 441–446. (In Chinese) [Google Scholar]
- Brezolin, A.N.; Martinazzo, J.; Muenchen, D.K.; De Cezaro, A.M.; Rigo, A.A.; Steffens, C.; Steffens, J.; Blassioli-Moraes, M.C.; Borges, M. Tools for detecting insect semiochemicals: A review. Anal. Bioanal. Chem. 2018, 410, 4091–4108. [Google Scholar] [CrossRef]
- Kalske, A.; Shiojiri, K.; Uesugi, A.; Sakata, Y.; Morrell, K.; Kessler, A. 2019. Insect herbivory selects for volatile-mediated plant-plant communication. Curr. Biol. 2005, 29, 3128–3133. [Google Scholar] [CrossRef]
- Xu, H.; Turlings, T.C.J. Plant volatiles as mate-finding cues for insects. Trends Plant Sci. 2018, 23, 100–111. [Google Scholar] [CrossRef]
- Bruce, T.J.; Pickett, J.A. Perception of plant volatile blends by herbivorous insects--finding the right mix. Phytochemistry 2011, 72, 1605–1611. [Google Scholar] [CrossRef]
- Koutroumpa, F.A.; Jacquin-Joly, E. Sex in the night: Fatty acid-derived sex pheromones and corresponding membrane pheromone receptors in insects. Biochimie 2014, 107, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Regnier, F.E.; Law, J.H. Insect pheromones. J. Lipid Res. 1968, 9, 541–551. [Google Scholar] [CrossRef]
- Gregg, P.C.; Del Socorro, A.P.; Landolt, P.J. Advances in attract-and-kill for agricultural pests: Beyond pheromones. Annu. Rev. Entomol. 2018, 63, 453–470. [Google Scholar] [CrossRef] [PubMed]
- Ruther, J.; Reinecke, A.; Tolasch, T.; Hilker, M. Make love not war: A common arthropod defence compound as sex pheromone in the forest cockchafer Melolontha hippocastani. Oecologia 2001, 128, 44–47. [Google Scholar] [CrossRef] [PubMed]
- McKenna, M.P.; Hekmat-Scafe, D.S.; Gaines, P.; Carlson, J.R. Putative drosophila pheromone-binding proteins expressed in a subregion of the olfactory system. J. Biol. Chem. 1994, 269, 16340–16347. [Google Scholar] [CrossRef]
- Pikielny, C.W.; Hasan, G.; Rouyer, F.; Rosbash, M. Members of a family of drosophila putative odorant-binding proteins are expressed in different subsets of olfactory hairs. Neuron 1994, 12, 35–49. [Google Scholar] [CrossRef]
- Mameli, M.; Tuccini, A.; Mazza, M.; Petacchi, R.; Pelosi, P. Soluble proteins in chemosensory organs of phasmids. Insect Biochem. Mol. Biol. 1996, 26, 875–882. [Google Scholar] [CrossRef]
- Tuccini, A.; Maida, R.; Rovero, P.; Mazza, M.; Pelosi, P. Putative odorant-binding protein in antennae and legs of Carausius morosus (insecta, phasmatodea). Insect Biochem. Mol. Biol. 1996, 26, 19–24. [Google Scholar] [CrossRef]
- Maleszka, R.; Stange, G. Molecular cloning, by a novel approach, of a cdna encoding a putative olfactory protein in the labial palps of the moth Cactoblastis cactorum. Gene 1997, 202, 39–43. [Google Scholar] [CrossRef]
- Biessmann, H.; Nguyen, Q.K.; Le, D.; Walter, M.F. Microarray-based survey of a subset of putative olfactory genes in the mosquito Anopheles gambiae. Insect Mol. Biol. 2005, 14, 575–589. [Google Scholar] [CrossRef]
- Biessmann, H.; Walter, M.F.; Dimitratos, S.; Woods, D. Isolation of cdna clones encoding putative odourant binding proteins from the antennae of the malaria-transmitting mosquito, Anopheles gambiae. Insect Mol. Biol. 2002, 11, 123–132. [Google Scholar] [CrossRef]
- Angeli, S.; Ceron, F.; Scaloni, A.; Monti, M.; Monteforti, G.; Minnocci, A.; Petacchi, R.; Pelosi, P. Purification, structural characterization, cloning and immunocytochemical localization of chemoreception proteins from Schistocerca gregaria. Eur. J. Biochem. 1999, 262, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, P.; Zhou, J.J.; Ban, L.P.; Calvello, M. Soluble proteins in insect chemical communication. Cell. Mol. Life Sci. 2006, 63, 1658–1676. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, P.; Iovinella, I.; Zhu, J.; Wang, G.; Dani, F.R. Beyond chemoreception: Diverse tasks of soluble olfactory proteins in insects. Biol. Rev. Camb. Philos. Soc. 2018, 93, 184–200. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.Y.; Zhang, N.X.; Shu, Z.Q.; Huang, K.X. Observation of mating and egg-laying habits of adult worms (carposina niponensis wal.). Insect Knowl. 1964, 6, 271–273. (In Chinese) [Google Scholar]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; Von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Younas, A.; Waris, M.I.; Shaaban, M.; Ul Qamar, M.T.; Wang, M.Q. Appraisal of MsepCSP14 for chemosensory functions in Mythimna separata. Insect Sci. 2022, 29, 162–176. [Google Scholar] [CrossRef]
- Zhang, Z.K.; Lei, Z.R. Identification, expression profiling and fluorescence-based binding assays of a chemosensory protein gene from the western flower thrips, Frankliniella occidentalis. PLoS ONE 2015, 10, e0117726. [Google Scholar] [CrossRef]
- Zhang, C.; Tang, B.; Zhou, T.; Yu, X.; Hu, M.; Dai, W. Involvement of chemosensory protein bodocsp1 in perception of host plant volatiles in Bradysia odoriphaga. J. Agric. Food Chem. 2021, 69, 10797–10806. [Google Scholar] [CrossRef]
- Li, C.; Sun, K.; Li, D.; Liu, D. Functional characterization of chemosensory protein AmalCSP5 from apple buprestid beetle, Agrilus mali (Coleoptera: Buprestidae). J. Econ. Entomol. 2021, 114, 348–359. [Google Scholar] [CrossRef]
- Yi, X.; Qi, J.; Zhou, X.; Hu, M.Y.; Zhong, G.H. Differential expression of chemosensory-protein genes in midguts in response to diet of Spodoptera litura. Sci. Rep. 2017, 7, 296. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Wang, S.N.; Li, K.M.; Liu, J.T.; Zheng, Y.; Shan, S.; Yang, Y.Q.; Li, R.J.; Zhang, Y.J.; Guo, Y.Y. Identification of odorant binding proteins and chemosensory proteins in Microplitis mediator as well as functional characterization of chemosensory protein 3. PLoS ONE 2017, 12, e0180775. [Google Scholar] [CrossRef] [PubMed]
- Hua, J.F.; Zhang, S.; Cui, J.J.; Wang, D.J.; Wang, C.Y.; Luo, J.Y.; Lv, L.M.; Ma, Y. Functional characterizations of one odorant binding protein and three chemosensory proteins from Apolygus lucorum (meyer-dur) (Hemiptera: Miridae) legs. J. Insect Physiol. 2013, 59, 690–696. [Google Scholar] [CrossRef] [PubMed]
- Waris, M.I.; Younas, A.; Ameen, A.; Rasool, F.; Wang, M.Q. Expression profiles and biochemical analysis of chemosensory protein 3 from Nilaparvata lugens (Hemiptera: Delphacidae). J. Chem. Ecol. 2020, 46, 363–377. [Google Scholar] [CrossRef]
- Jacquin-Joly, E.; Vogt, R.G.; François, M.C.; Nagnan-Le Meillour, P. Functional and expression pattern analysis of chemosensory proteins expressed in antennae and pheromonal gland of Mamestra brassicae. Chem. Senses 2001, 26, 833–844. [Google Scholar] [CrossRef]
- Nagnan-Le Meillour, P.; Cain, A.H.; Jacquin-Joly, E.; François, M.C.; Ramachandran, S.; Maida, R.; Steinbrecht, R.A. Chemosensory proteins from the proboscis of Mamestra brassicae. Chem. Senses 2000, 25, 541–553. [Google Scholar] [CrossRef]
- Zhou, X.H.; Ban, L.P.; Iovinella, I.; Zhao, L.J.; Gao, Q.; Felicioli, A.; Sagona, S.; Pieraccini, G.; Pelosi, P.; Zhang, L.; et al. Diversity, abundance, and sex-specific expression of chemosensory proteins in the reproductive organs of the locust Locusta migratoria manilensis. Biol. Chem. 2013, 394, 43–54. [Google Scholar] [CrossRef]
- Baer, B.; Zareie, R.; Paynter, E.; Poland, V.; Millar, A.H. Seminal fluid proteins differ in abundance between genetic lineages of honeybees. J. Proteom. 2012, 75, 5646–5653. [Google Scholar] [CrossRef]
- Kitabayashi, A.N.; Arai, T.; Kubo, T.; Natori, S. Molecular cloning of cdna for p10, a novel protein that increases in the regenerating legs of periplaneta americana (American cockroach). Insect Biochem. Mol. Biol. 1998, 28, 785–790. [Google Scholar] [CrossRef]
- Nomura, A.; Kawasaki, K.; Kubo, T.; Natori, S. Purification and localization of p10, a novel protein that increases in nymphal regenerating legs of periplaneta americana (American cockroach). Int. J. Dev. Biol. 1992, 36, 391–398. [Google Scholar]
- Forêt, S.; Wanner, K.W.; Maleszka, R. Chemosensory proteins in the honey bee: Insights from the annotated genome, comparative analyses and expressional profiling. Insect Biochem. Mol. Biol. 2007, 37, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Maleszka, J.; Forêt, S.; Saint, R.; Maleszka, R. Rnai-induced phenotypes suggest a novel role for a chemosensory protein csp5 in the development of embryonic integument in the honeybee (Apis mellifera). Dev. Genes. Evol. 2007, 217, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Lu, Y.; Zeng, L.; Liang, G.; He, X. Si-csp9 regulates the integument and moulting process of larvae in the red imported fire ant, Solenopsis invicta. Sci. Rep. 2015, 5, 9245. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.L.; Guo, H.; Huang, L.Q.; Pelosi, P.; Wang, C.Z. Unique function of a chemosensory protein in the proboscis of two helicoverpa species. J. Exp. Biol. 2014, 217, 1821–1826. [Google Scholar] [CrossRef]
- Jiang, X.; Xu, H.; Zheng, N.; Yin, X.; Zhang, L. A chemosensory protein detects antifeedant in locust (Locusta migratoria). Insects 2020, 12, 1. [Google Scholar] [CrossRef]
- Zeng, Y.; Merchant, A.; Wu, Q.; Wang, S.; Kong, L.; Zhou, X.; Xie, W.; Zhang, Y. A chemosensory protein btabcsp11 mediates reproduction in Bemisia tabaci. Front. Physiol. 2020, 11, 709. [Google Scholar] [CrossRef]
- Younas, A.; Waris, M.I.; Chang, X.Q.; Shaaban, M.; Abdelnabby, H.; Ul Qamar, M.T.; Wang, M.Q. A chemosensory protein MsepCSP5 involved in chemoreception of oriental armyworm Mythimna separata. Int. J. Biol. Sci. 2018, 14, 1935–1949. [Google Scholar] [CrossRef]
- Ali, S.; Ahmed, M.Z.; Li, N.; Ali, S.A.I.; Wang, M.Q. Functional characteristics of chemosensory proteins in the sawyer beetle Monochamus alternatus hope. Bull. Entomol. Res. 2019, 109, 34–42. [Google Scholar] [CrossRef]
- Chénier, J.V.; Philogène, B.J. Field responses of certain forest coleoptera to conifer monoterpenes and ethanol. J. Chem. Ecol. 1989, 15, 1729–1745. [Google Scholar] [CrossRef]
- Hassaballa, I.B.; Torto, B.; Sole, C.L.; Tchouassi, D.P. Exploring the influence of different habitats and their volatile chemistry in modulating sand fly population structure in a leishmaniasis endemic foci, kenya. PLOS Neglected Trop. Dis. 2021, 15, e0009062. [Google Scholar] [CrossRef]
- Jumean, Z.; Gries, R.; Unruh, T.; Rowland, E.; Gries, G. Identification of the larval aggregation pheromone of codling moth, Cydia pomonella. J. Chem. Ecol. 2005, 31, 911–924. [Google Scholar] [CrossRef]
- Siljander, E.; Gries, R.; Khaskin, G.; Gries, G. Identification of the airborne aggregation pheromone of the common bed bug, cimex lectularius. J. Chem. Ecol. 2008, 34, 708–718. [Google Scholar] [CrossRef] [PubMed]
- Farré-Armengol, G.; Fernández-Martínez, M.; Filella, I.; Junker, R.R.; Peñuelas, J. Deciphering the biotic and climatic factors that influence floral scents: A systematic review of floral volatile emissions. Front. Plant Sci. 2020, 11, 1154. [Google Scholar] [CrossRef] [PubMed]
- Lohonyai, Z.; Vuts, J.; Kárpáti, Z.; Koczor, S.; Domingue, M.J.; Fail, J.; Birkett, M.A.; Tóth, M.; Imrei, Z. Benzaldehyde: An alfalfa-related compound for the spring attraction of the pest weevil Sitona humeralis (Coleoptera: Curculionidae). Pest. Manag. Sci. 2019, 75, 3153–3159. [Google Scholar] [CrossRef] [PubMed]
- De Fouchier, A.; Sun, X.; Caballero-Vidal, G.; Travaillard, S.; Jacquin-Joly, E.; Montagné, N. Behavioral effect of plant volatiles binding to Spodoptera littoralis larval odorant receptors. Front. Behav. Neurosci. 2018, 12, 264. [Google Scholar] [CrossRef] [PubMed]
- Pope, T.W.; Campbell, C.A.; Hardie, J.; Pickett, J.A.; Wadhams, L.J. Interactions between host-plant volatiles and the sex pheromones of the bird cherry-oat aphid, Rhopalosiphum padi and the damson-hop aphid. Phorodon humuli. J. Chem. Ecol. 2007, 33, 157–165. [Google Scholar] [CrossRef]
- Piñero, J.C.; Prokopy, R.J. Field evaluation of plant odor and pheromonal combinations for attracting plum curculios. J. Chem. Ecol. 2003, 29, 2735–2748. [Google Scholar] [CrossRef]
- Ullah, I.; Khan, A.L.; Ali, L.; Khan, A.R.; Waqas, M.; Hussain, J.; Lee, I.J.; Shin, J.H. Benzaldehyde as an insecticidal, antimicrobial, and antioxidant compound produced by photorhabdus temperata M1021. J. Microbiol. 2015, 53, 127–133. [Google Scholar] [CrossRef]
- Cooperband, M.F.; McElfresh, J.S.; Millar, J.G.; Cardé, R.T. Attraction of female Culex quinquefasciatus say (Diptera: Culicidae) to odors from chicken feces. J. Insect Physiol. 2008, 54, 1184–1192. [Google Scholar] [CrossRef]
- Bezerra-Silva, P.C.; Dutra, K.A.; Santos, G.K.; Silva, R.C.; Iulek, J.; Milet-Pinheiro, P.; Navarro, D.M. Evaluation of the activity of the essential oil from an ornamental flower against Aedes aegypti: Electrophysiology, molecular dynamics and behavioral assays. PLoS ONE 2016, 11, e0150008. [Google Scholar] [CrossRef]
- Collatz, J.; Tolasch, T.; Steidle, J.L. Mate finding in the parasitic wasp Cephalonomia tarsalis (ashmead): More than one way to a female’s heart. J. Chem. Ecol. 2009, 35, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Jayanthi, K.; Saravan, P.D.; Kumar, P.; Vyas, M. Odour cues from fruit arils of Artocarpus heterophyllus attract both sexes of oriental fruit flies. J. Chem. Ecol. 2021, 47, 552–563. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.Y.; Liu, Z.J.; Wang, X.Y.; Zhang, Z.L.; Lu, W.; Zheng, X.L. Electroantennographic and olfactory responses of Quadrastichus mendeli to eucalyptus volatiles induced by the gall-forming insect Leptocybe invasa. Pest. Manag. Sci. 2022, 78, 2405–2416. [Google Scholar] [CrossRef] [PubMed]
- Magalhães-Junior, J.T.; Oliva-Filho, A.A.; Novais, H.O.; Mesquita, P.R.R.; Rodrigues, M.; Pinto, M.C.; Barrouin-Melo, S.M. Attraction of the sandfly lutzomyia longipalpis to possible biomarker compounds from dogs infected with Leishmania infantum. Med. Vet. Entomol. 2019, 33, 322–325. [Google Scholar] [CrossRef]
- Laznik, Ž.; Trdan, S. Attraction behaviors of entomopathogenic nematodes (steinernematidae and heterorhabditidae) to synthetic volatiles emitted by insect damaged potato tubers. J. Chem. Ecol. 2016, 42, 314–322. [Google Scholar] [CrossRef]
- Li, J.; Zhang, L.; Wang, X. An odorant-binding protein involved in perception of host plant odorants in locust Locusta migratoria. Arch. Insect Biochem. Physiol. 2016, 91, 221–229. [Google Scholar] [CrossRef]
- Ullah, R.M.K.; Quershi, S.R.; Adeel, M.M.; Abdelnabby, H.; Waris, M.I.; Duan, S.G.; Wang, M.Q. An odorant binding protein (saveobp9) involved in chemoreception of the wheat aphid Sitobion avenae. Int. J. Mol. Sci. 2020, 21, 8331. [Google Scholar] [CrossRef]
- Molnár, B.P.; Tóth, Z.; Fejes-Tóth, A.; Dekker, T.; Kárpáti, Z. Electrophysiologically-active maize volatiles attract gravid female european corn borer, Ostrinia nubilalis. J. Chem. Ecol. 2015, 41, 997–1005. [Google Scholar] [CrossRef]
- Morawo, T.; Burrows, M.; Fadamiro, H. Electroantennogram response of the parasitoid, Microplitis croceipes to host-related odors: The discrepancy between relative abundance and level of antennal responses to volatile compound. F1000Res 2016, 5, 2725. [Google Scholar] [CrossRef]
- Du, Y.; Zhang, J.; Yan, Z.; Ma, Y.; Yang, M.; Zhang, M.; Zhang, Z.; Qin, L.; Cao, Q. Host preference and performance of the yellow peach moth (Conogethes punctiferalis) on chestnut cultivars. PLoS ONE 2016, 11, e0157609. [Google Scholar] [CrossRef]
- Liu, X.F.; Chen, H.H.; Li, J.K.; Zhang, R.; Turlings, T.C.; Chen, L. Volatiles released by chinese liquorice roots mediate host location behaviour by neonate Porphyrophora sophorae (Hemiptera: Margarodidae). Pest. Manag. Sci. 2016, 72, 1959–1964. [Google Scholar] [CrossRef] [PubMed]
- Liao, S.; Liu, Y.; Si, H.; Xiao, Z.; Fan, G.; Chen, S.; Wang, P.; Wang, Z. Hydronopylformamides: Modification of the naturally occurring compound (-)-β-pinene to produce insect repellent candidates against Blattella germanica. Molecules 2017, 22, 1004. [Google Scholar] [CrossRef] [PubMed]
- Dekel, A.; Yakir, E.; Bohbot, J.D. The sulcatone receptor of the strict nectar-feeding mosquito toxorhynchites amboinensis. Insect Biochem. Mol. Biol. 2019, 111, 103174. [Google Scholar] [CrossRef] [PubMed]
- May-Concha, I.; Rojas, J.C.; Cruz-López, L.; Millar, J.G.; Ramsey, J.M. Volatile compounds emitted by Triatoma dimidiata, a vector of chagas disease: Chemical analysis and behavioural evaluation. Med. Vet. Entomol. 2013, 27, 165–174. [Google Scholar] [CrossRef]
- Gu, D.; Yang, D.R. Utilisation of chemical signals by inquiline wasps in entering their host figs. J. Insect Physiol. 2013, 59, 1065–1068. [Google Scholar] [CrossRef]
- Olaide, O.Y.; Tchouassi, D.P.; Yusuf, A.A.; Pirk, C.W.W.; Masiga, D.K.; Saini, R.K.; Torto, B. Zebra skin odor repels the savannah tsetse fly, Glossina pallidipes (Diptera: Glossinidae). PLOS Neglected Trop. Dis. 2019, 13, e0007460. [Google Scholar] [CrossRef]
- Logan, J.G.; Stanczyk, N.M.; Hassanali, A.; Kemei, J.; Santana, A.E.; Ribeiro, K.A.; Pickett, J.A.; Mordue Luntz, A.J. Arm-in-cage testing of natural human-derived mosquito repellents. Malar. J. 2010, 9, 239. [Google Scholar] [CrossRef]
- Logan, J.G.; Seal, N.J.; Cook, J.I.; Stanczyk, N.M.; Birkett, M.A.; Clark, S.J.; Gezan, S.A.; Wadhams, L.J.; Pickett, J.A.; Mordue, A.J. Identification of human-derived volatile chemicals that interfere with attraction of the scottish biting midge and their potential use as repellents. J. Med. Entomol. 2009, 46, 208–219. [Google Scholar] [CrossRef]
- Birkett, M.A.; Agelopoulos, N.; Jensen, K.M.; Jespersen, J.B.; Pickett, J.A.; Prijs, H.J.; Thomas, G.; Trapman, J.J.; Wadhams, L.J.; Woodcock, C.M. The role of volatile semiochemicals in mediating host location and selection by nuisance and disease-transmitting cattle flies. Med. Vet. Entomol. 2004, 18, 313–322. [Google Scholar] [CrossRef]
- Malcicka, M.; Bezemer, T.M.; Visser, B.; Bloemberg, M.; Snart, C.J.; Hardy, I.C.; Harvey, J.A. Multi-trait mimicry of ants by a parasitoid wasp. Sci. Rep. 2015, 5, 8043. [Google Scholar] [CrossRef]
- Wu, F.; Feng, Y.; Han, B.; Hu, H.; Feng, M.; Meng, L.; Ma, C.; Yu, L.; Li, J. Mechanistic insight into binding interaction between chemosensory protein 4 and volatile larval pheromones in honeybees (Apis mellifera). Int. J. Biol. Macromol. 2019, 141, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Jing, T.; Qian, X.; Du, W.; Gao, T.; Li, D.; Guo, D.; He, F.; Yu, G.; Li, S.; Schwab, W.; et al. Herbivore-induced volatiles influence moth preference by increasing the β-ocimene emission of neighbouring tea plants. Plant Cell Environ. 2021, 44, 3667–3680. [Google Scholar] [CrossRef] [PubMed]
- Cozzolino, S.; Fineschi, S.; Litto, M.; Scopece, G.; Trunschke, J.; Schiestl, F.P. Herbivory increases fruit set in silene latifolia: A consequence of induced pollinator-attracting floral volatiles. J. Chem. Ecol. 2015, 41, 622–630. [Google Scholar] [CrossRef]
- Tang, R.; Zhang, J.P.; Zhang, Z.N. Electrophysiological and behavioral responses of male fall webworm moths (Hyphantria cunea) to herbivory-induced mulberry (Morus alba) leaf volatiles. PLoS ONE 2012, 7, e49256. [Google Scholar] [CrossRef] [PubMed]
- Kang, Z.W.; Liu, F.H.; Zhang, Z.F.; Tian, H.G.; Liu, T.X. Volatile β-ocimene can regulate developmental performance of peach aphid Myzus persicae through activation of defense responses in chinese cabbage brassica pekinensis. Front. Plant Sci. 2018, 9, 708. [Google Scholar] [CrossRef]
- Cascone, P.; Iodice, L.; Maffei, M.E.; Bossi, S.; Arimura, G.; Guerrieri, E. Tobacco overexpressing β-ocimene induces direct and indirect responses against aphids in receiver tomato plants. J. Plant Physiol. 2015, 173, 28–32. [Google Scholar] [CrossRef]
- Jacob, J.W.; Tchouassi, D.P.; Lagat, Z.O.; Mathenge, E.M.; Mweresa, C.K.; Torto, B. Independent and interactive effect of plant- and mammalian- based odors on the response of the malaria vector, Anopheles gambiae. Acta Trop. 2018, 185, 98–106. [Google Scholar] [CrossRef]
- Koczor, S.; Vuts, J.; Caulfield, J.C.; Withall, D.M.; Sarria, A.; Pickett, J.A.; Birkett, M.A.; Csonka, É.B.; Tóth, M. Sex pheromone of the alfalfa plant bug, Adelphocoris lineolatus: Pheromone composition and antagonistic effect of 1-hexanol (Hemiptera: Miridae). J. Chem. Ecol. 2021, 47, 525–533. [Google Scholar] [CrossRef]
- Sun, L.; Gu, S.H.; Xiao, H.J.; Zhou, J.J.; Guo, Y.Y.; Liu, Z.W.; Zhang, Y.J. The preferential binding of a sensory organ specific odorant binding protein of the alfalfa plant bug Adelphocoris lineolatus alinobp10 to biologically active host plant volatiles. J. Chem. Ecol. 2013, 39, 1221–1231. [Google Scholar] [CrossRef]
- Faccoli, M.; Blazenec, M.; Schlyter, F. Feeding response to host and nonhost compounds by males and females of the spruce bark beetle Ips typographus in a tunneling microassay. J. Chem. Ecol. 2005, 31, 745–759. [Google Scholar] [CrossRef]
Figure 1.
Multiple sequence comparison between CasaCSPs and Lepidoptera. Lepidoptera included Athetis dissimilis (AND82450.1), Conogethes pinicolalis (QFR36131.1, QFR36144.1), Conogethes punctiferalis (AHX37218.1), Dendrolimus kikuchii (AII01035.1), Dioryctria abietella (QQG64119.1, QJX59148.1), Glyphodes pyloalis (QIJ45712.1), Grapholita molesta (ALC79596.1), Helicoverpa armigera (AAK53762.1), Helicoverpa assulta (ABB91378.1), Helicoverpa zea (AAN63675.1), Lobesia botrana (AXF48707.1), Ostrinia furnacalis (BAV56805.1, BAV56812.1) and Semiothisa cinerearia (QRF70949.1). The red stars explain four highly conserved cysteine residues of the CSP family.
Figure 1.
Multiple sequence comparison between CasaCSPs and Lepidoptera. Lepidoptera included Athetis dissimilis (AND82450.1), Conogethes pinicolalis (QFR36131.1, QFR36144.1), Conogethes punctiferalis (AHX37218.1), Dendrolimus kikuchii (AII01035.1), Dioryctria abietella (QQG64119.1, QJX59148.1), Glyphodes pyloalis (QIJ45712.1), Grapholita molesta (ALC79596.1), Helicoverpa armigera (AAK53762.1), Helicoverpa assulta (ABB91378.1), Helicoverpa zea (AAN63675.1), Lobesia botrana (AXF48707.1), Ostrinia furnacalis (BAV56805.1, BAV56812.1) and Semiothisa cinerearia (QRF70949.1). The red stars explain four highly conserved cysteine residues of the CSP family.

Figure 2.
Phylogenetic tree of CsasCSPs and other organisms. Diptera included Bactrocera minax and Drosophila melanogaster; Hemiptera included Myzus persicae, Nilaparvata lugens and Tropidothorax elegans; Coleoptera included Holotrichia oblita, Anomala corpulenta and Oedaleus asiaticus; Lepidoptera included Athetis dissimilis, Conogethes pinicolalis, Conogethes punctiferalis, Dendrolimus kikuchii, Dioryctria abietella, Glyphodes pyloalis, Grapholita molesta, Helicoverpa armigera, Helicoverpa assulta, Helicoverpa zea, Lobesia botrana, Ostrinia furnacalis and Semiothisa cinerearia.
Figure 2.
Phylogenetic tree of CsasCSPs and other organisms. Diptera included Bactrocera minax and Drosophila melanogaster; Hemiptera included Myzus persicae, Nilaparvata lugens and Tropidothorax elegans; Coleoptera included Holotrichia oblita, Anomala corpulenta and Oedaleus asiaticus; Lepidoptera included Athetis dissimilis, Conogethes pinicolalis, Conogethes punctiferalis, Dendrolimus kikuchii, Dioryctria abietella, Glyphodes pyloalis, Grapholita molesta, Helicoverpa armigera, Helicoverpa assulta, Helicoverpa zea, Lobesia botrana, Ostrinia furnacalis and Semiothisa cinerearia.

Figure 3.
Expression levels of the CsasCSP7, CsasCSP9 and CsasCSP11 genes between male and female C. sasakii. (A). CsasCSP7; (B) CsasCSP9; (C) CsasCSP11. The letters on the top of each bar means the statistic differences (p < 0.05).
Figure 3.
Expression levels of the CsasCSP7, CsasCSP9 and CsasCSP11 genes between male and female C. sasakii. (A). CsasCSP7; (B) CsasCSP9; (C) CsasCSP11. The letters on the top of each bar means the statistic differences (p < 0.05).

Figure 4.
SDS-PAGE analysis of expressed CsasCSP7, CsasCSP9 and CsasCSP11 proteins. 1: The purified protein; 2: The refolding protein; 3: Supernatant of the induced; 4: Inclusion body of the induced; 5: Expressed product induced with IPTG; 6: Expressed product without induction with IPTG; M: Standard protein marker.
Figure 4.
SDS-PAGE analysis of expressed CsasCSP7, CsasCSP9 and CsasCSP11 proteins. 1: The purified protein; 2: The refolding protein; 3: Supernatant of the induced; 4: Inclusion body of the induced; 5: Expressed product induced with IPTG; 6: Expressed product without induction with IPTG; M: Standard protein marker.

Figure 5.
Binding curves of 1-AMA to CsasCSPs.

Figure 6.
Competitive binding curves of CsasCSPs with compounds.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Primers used in this study.
| Primer Name | Primer Sequence (5′-3′) | Length (bp) | Purpose |
|---|---|---|---|
| CsasCSP7-F | AGGTTATTGAGCATCTGATTAAG | 95 | Fluorescence quantification |
| CsasCSP7-R | TTCATACTTCTTTCTCCACTTG | ||
| CsasCSP9-F | GTTATGGAGTACATCATAGATC | 102 | |
| CsasCSP9-R | TTTCTTCTCTTCATACTTACTC | ||
| CsasCSP11-F | CAAGTAGTCCGATACATTAGG | 125 | |
| CsasCSP11-R | TAATCATCAGAAGCGAAGAAT | ||
| CsasCSP7-F | CGGGATCCATGGAAGAAAAGTATTCGGACAAATA | 342 | Prokaryotic expression |
| CsasCSP7-R | TGGAATTCCTATTTTTCAGGTATTTCAATACCCCT | ||
| CsasCSP9-F | CGGGATCCATGCGCCCCGAAGAGCACT | 432 | |
| CsasCSP9-R | TGGAATTCTTATGGCCTTGACGGTGCG | ||
| CsasCSP11-F | CGGGATCCATGGATGAGGAGCAGTATACAGATAGAT | 333 | |
| CsasCSP11-R | TGGAATTCTTAATCATCAGAAGCGAAGAATG |
Note: The restriction sites are underlined.
Table 2.
Plant volatiles and sex compounds for this experiment.
| Code | Prospective Ligand | CAS | Molecular Weight /g·mol−1 | Purity/% |
|---|---|---|---|---|
| L1 | 2-Methylbutyl acetate | 624-41-9 | 130.18 | 99 |
| L2 | Butyl butyrate | 109-21-7 | 144.21 | >99 |
| L3 | Butyl heptanoate | 5454-28-4 | 186.29 | >99 |
| L4 | Butyl octanoate | 589-75-3 | 200.32 | >99 |
| L5 | Ethyl butyrate | 105-54-4 | 116.16 | 99 |
| L6 | Ethyl heptanoate | 106-30-9 | 158.24 | >98 |
| L7 | Ethyl hexanoate | 123-66-0 | 144.21 | 99 |
| L8 | Hexyl hexanoate | 6378-65-0 | 200.32 | 98 |
| L9 | Isoamyl acetate | 123-92-2 | 130.19 | 99 |
| L10 | Methyl jasmonate | 39,924-52-2 | 224.3 | 95 |
| L11 | Propyl octanoate | 624-13-5 | 186.29 | 98 |
| L12 | Tert-butyl acetate | 540-88-5 | 116.15 | 99 |
| L13 | (Z)-3-Hexenyl acetate | 3681-71-8 | 142.2 | 98 |
| L14 | Benzaldehyde | 100-52-7 | 106.12 | >99 |
| L15 | Decanal | 112-31-2 | 156.27 | 97 |
| L16 | Dodecanal | 112-54-9 | 184.32 | >95 |
| L17 | (E)-Hex-2-enal | 6728-26-3 | 98.14 | 98 |
| L18 | Hexanal | 66-25-1 | 100.16 | 97 |
| L19 | Honanal | 124-19-6 | 142.24 | 96 |
| L20 | Octanal | 124-13-0 | 128.215 | 97 |
| L21 | 1-Hexanol | 111-27-3 | 102.18 | >98 |
| L22 | 2-Ethylhexanol | 104-76-7 | 130.22 | 99 |
| L23 | 3-Methyl-1-butanol | 123-51-3 | 88.15 | 99 |
| L24 | (E)-2-Hexen-1-ol | 928-95-0 | 100.16 | 97 |
| L25 | (Z)-Hex-3-en-1-ol | 928-96-1 | 100.16 | ≥98 |
| L26 | Decane | 124-18-5 | 142.29 | >99 |
| L27 | Hexadecane | 544-76-3 | 226.45 | >98 |
| L28 | Octadecane | 593-45-3 | 254.49 | 98 |
| L29 | Pentadecane | 629-62-9 | 212.41 | 98 |
| L30 | Tetradecane | 629-59-4 | 198.39 | ≥99 |
| L31 | alpha-Farnesene | 502-61-4 | 204.25 | >99 |
| L32 | Beta-Ocimene | 13,877-91-3 | 136.23 | >90 |
| L33 | (-)-beta-Pinene | 18,172-67-3 | 136.24 | >94 |
| L34 | Myrcene | 123-35-3 | 136.236 | >80 |
| L35 | 6-Methyl-5-hepten-2-one | 110-93-0 | 126.2 | >98 |
| L36 | Benzonitrile | 100-47-0 | 103.12 | >99 |
| L37 | Z-7-Eicosene-11-one | 63,408-44-6 | 294.5 | >99 |
| L38 | Z-7-Nonadecen-11-one | 63,408-45-7 | 280.5 | >99 |
Table 3.
Binding affinities of CsasCSP7 to all tested ligands.
| Ligand | Ki | IC50 | Ligand | Ki | IC50 | Ligand | Ki | IC50 |
|---|---|---|---|---|---|---|---|---|
| 2-Methylbutyl acetate | - | >50 | Benzaldehyde | 7.25 ± 0.23 | 8.00 ± 0.59 | Hexadecane | - | >50 |
| Butyl butyrate | - | >50 | Decanal | - | >50 | Octadecane | - | >50 |
| Butyl heptanoate | - | >50 | Dodecanal | 13.61 ± 0.54 | 14.97 ± 0.60 | Pentadecane | - | >50 |
| Butyl octanoate | - | >50 | (E)-Hex-2-enal | - | >50 | Tetradecane | - | >50 |
| Ethyl butyrate | - | >50 | Hexanal | - | >50 | alpha-Farnesene | - | >50 |
| Ethyl heptanoate | - | >50 | Honanal | - | >50 | Beta-Ocimene | - | >50 |
| Ethyl hexanoate | - | >50 | Octanal | - | >50 | (-)-beta-Pinene | - | >50 |
| Hexyl hexanoate | - | >50 | 1-Hexanol | - | >50 | Myrcene | - | >50 |
| Isoamyl acetate | - | >50 | 2-Ethylhexanol | - | >50 | 6-Methyl-5-hepten-2-one | - | >50 |
| Methyl jasmonate | - | >50 | 3-Methyl-1-butanol | - | >50 | Benzonitrile | - | >50 |
| Propyl octanoate | - | >50 | (E)-2-Hexen-1-ol | - | >50 | Z-7-Eicosene-11-one | - | >50 |
| Tert-butyl acetate | - | >50 | (Z)-Hex-3-en-1-ol | - | >50 | Z-7-Nonadecen-11-one | - | >50 |
| (Z)-3-Hexenyl acetate | - | >50 | Decane | - | >50 |
IC50 labeled “>50” indicates that binding affinities could not be calculated with the tested ligand concentrations, so Ki of the ligands is represented as “-”.
Table 4.
Binding affinities of CsasCSP9 to all tested ligands.
| Ligand | Ki | IC50 | Ligand | Ki | IC50 | Ligand | Ki | IC50 |
|---|---|---|---|---|---|---|---|---|
| 2-Methylbutyl acetate | - | >50 | Benzaldehyde | - | >50 | Hexadecane | - | >50 |
| Butyl butyrate | - | >50 | Decanal | 1.65 ± 0.31 | 1.92 ± 0.34 | Octadecane | - | >50 |
| Butyl heptanoate | - | >50 | Dodecanal | - | >50 | Pentadecane | - | >50 |
| Butyl octanoate | 1.47 ± 0.13 | 1.71 ± 0.16 | (E)-Hex-2-enal | - | >50 | Tetradecane | - | >50 |
| Ethyl butyrate | - | >50 | Hexanal | - | >50 | alpha-Farnesene | - | >50 |
| Ethyl heptanoate | - | >50 | Honanal | - | >50 | Beta-Ocimene | - | >50 |
| Ethyl hexanoate | - | >50 | Octanal | - | >50 | (-)-beta-Pinene | 14.26 ± 0.62 | 16.56 ± 0.24 |
| Hexyl hexanoate | - | >50 | 1-Hexanol | - | >50 | Myrcene | - | >50 |
| Isoamyl acetate | - | >50 | 2-Ethylhexanol | - | >50 | 6-Methyl-5-hepten-2-one | - | >50 |
| Methyl jasmonate | - | >50 | 3-Methyl-1-butanol | - | >50 | Benzonitrile | - | >50 |
| Propyl octanoate | - | >50 | (E)-2-Hexen-1-ol | - | >50 | Z-7-Eicosene-11-one | - | >50 |
| Tert-butyl acetate | - | >50 | (Z)-Hex-3-en-1-ol | - | >50 | Z-7-Nonadecen-11-one | - | >50 |
| (Z)-3-Hexenyl acetate | - | >50 | Decane | - | >50 |
IC50 labeled “>50” indicates that binding affinities could not be calculated with the tested ligand concentrations, so Ki of the ligands is represented as “-”.
Table 5.
Binding affinities of CsasCSP11 to all tested ligands.
| Ligand | Ki | IC50 | Ligand | Ki | IC50 | Ligand | Ki | IC50 |
|---|---|---|---|---|---|---|---|---|
| 2-Methylbutyl acetate | - | >50 | Benzaldehyde | - | >50 | Hexadecane | - | >50 |
| Butyl butyrate | - | >50 | Decanal | - | >50 | Octadecane | - | >50 |
| Butyl heptanoate | - | >50 | Dodecanal | - | >50 | Pentadecane | - | >50 |
| Butyl octanoate | - | >50 | (E)-Hex-2-enal | - | >50 | Tetradecane | - | >50 |
| Ethyl butyrate | - | >50 | Hexanal | - | >50 | alpha-Farnesene | - | >50 |
| Ethyl heptanoate | - | >50 | Honanal | - | >50 | Beta-Ocimene | 1.83 ± 0.66 | 2.09 ± 0.76 |
| Ethyl hexanoate | - | >50 | Octanal | - | >50 | (-)-beta-Pinene | - | >50 |
| Hexyl hexanoate | - | >50 | 1-Hexanol | 8.13 ± 0.78 | 9.31 ± 0.39 | Myrcene | - | >50 |
| Isoamyl acetate | - | >50 | 2-Ethylhexanol | - | >50 | 6-Methyl-5-hepten-2-one | 0.71 ± 0.07 | 0.81 ± 0.08 |
| Methyl jasmonate | - | >50 | 3-Methyl-1-butanol | - | >50 | Benzonitrile | - | >50 |
| Propyl octanoate | - | >50 | (E)-2-Hexen-1-ol | - | >50 | Z-7-Eicosene-11-one | - | >50 |
| Tert-butyl acetate | - | >50 | (Z)-Hex-3-en-1-ol | - | >50 | Z-7-Nonadecen-11-one | - | >50 |
| (Z)-3-Hexenyl acetate | - | >50 | Decane | - | >50 |
IC50 labeled “>50” indicates that binding affinities could not be calculated with the tested ligand concentrations, so Ki of the ligands is represented as “-”.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, L.; Qiu, G.; Zhang, H.; Yue, Q.; Yan, W.; Sun, L. Molecular Characterization of Three Chemosensory Proteins from Carposina sasakii. Agriculture 2023, 13, 2066. https://doi.org/10.3390/agriculture13112066
AMA Style
Liu L, Qiu G, Zhang H, Yue Q, Yan W, Sun L. Molecular Characterization of Three Chemosensory Proteins from Carposina sasakii. Agriculture. 2023; 13(11):2066. https://doi.org/10.3390/agriculture13112066
Chicago/Turabian StyleLiu, Liu, Guisheng Qiu, Huaijiang Zhang, Qiang Yue, Wentao Yan, and Lina Sun. 2023. "Molecular Characterization of Three Chemosensory Proteins from Carposina sasakii" Agriculture 13, no. 11: 2066. https://doi.org/10.3390/agriculture13112066
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.