Understanding In Vitro Embryo Development through Classical Germination Measurements: A Case Study of Dragon’s Blood (Croton lechleri Müll Arg.)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fruit Collection and Processing
2.2. Seed Disinfestation and Embryo Development In Vitro Protocols
2.3. Embryo Development Measurements
2.4. Experimental Design and Statistical Analysis
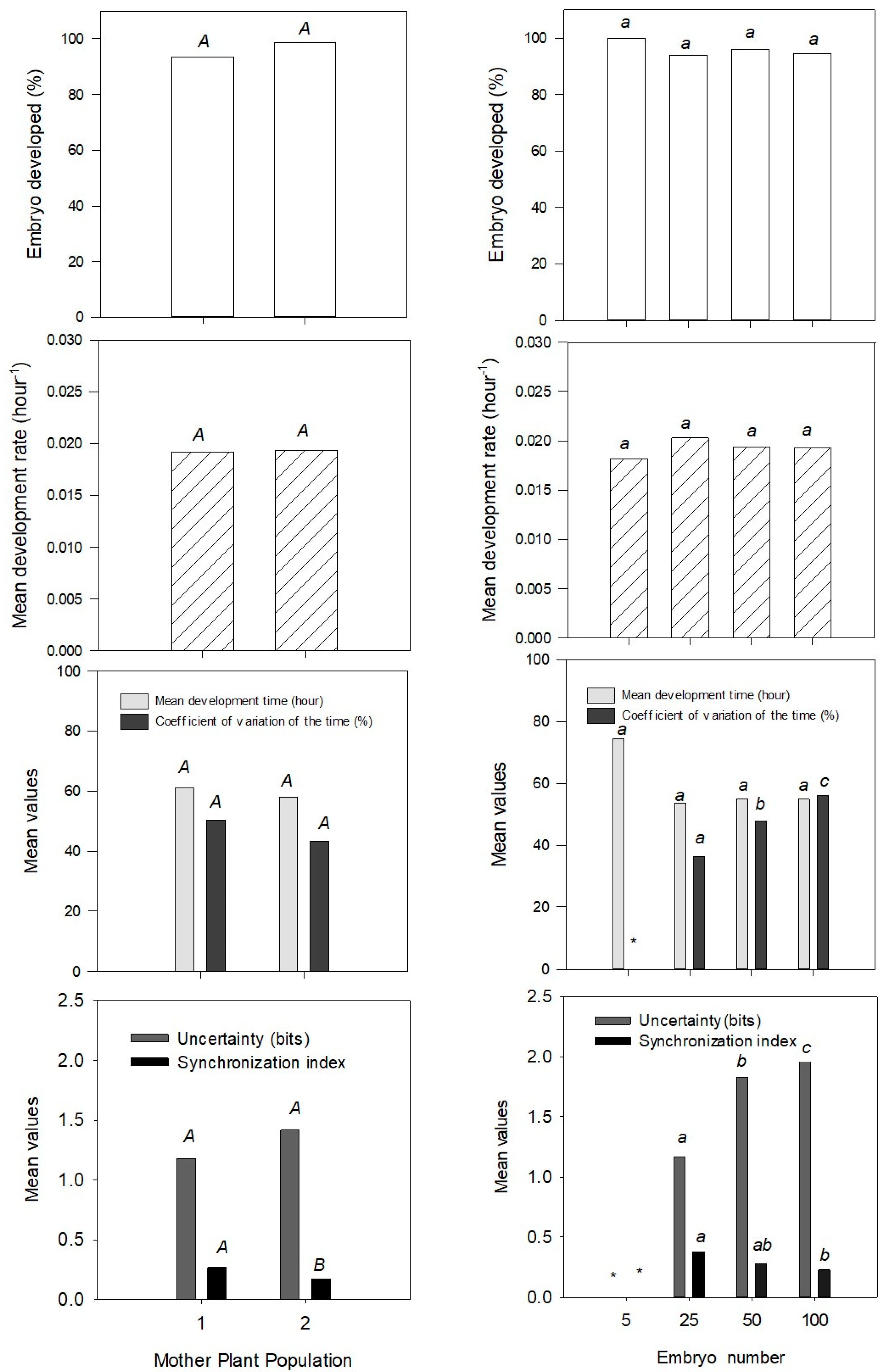
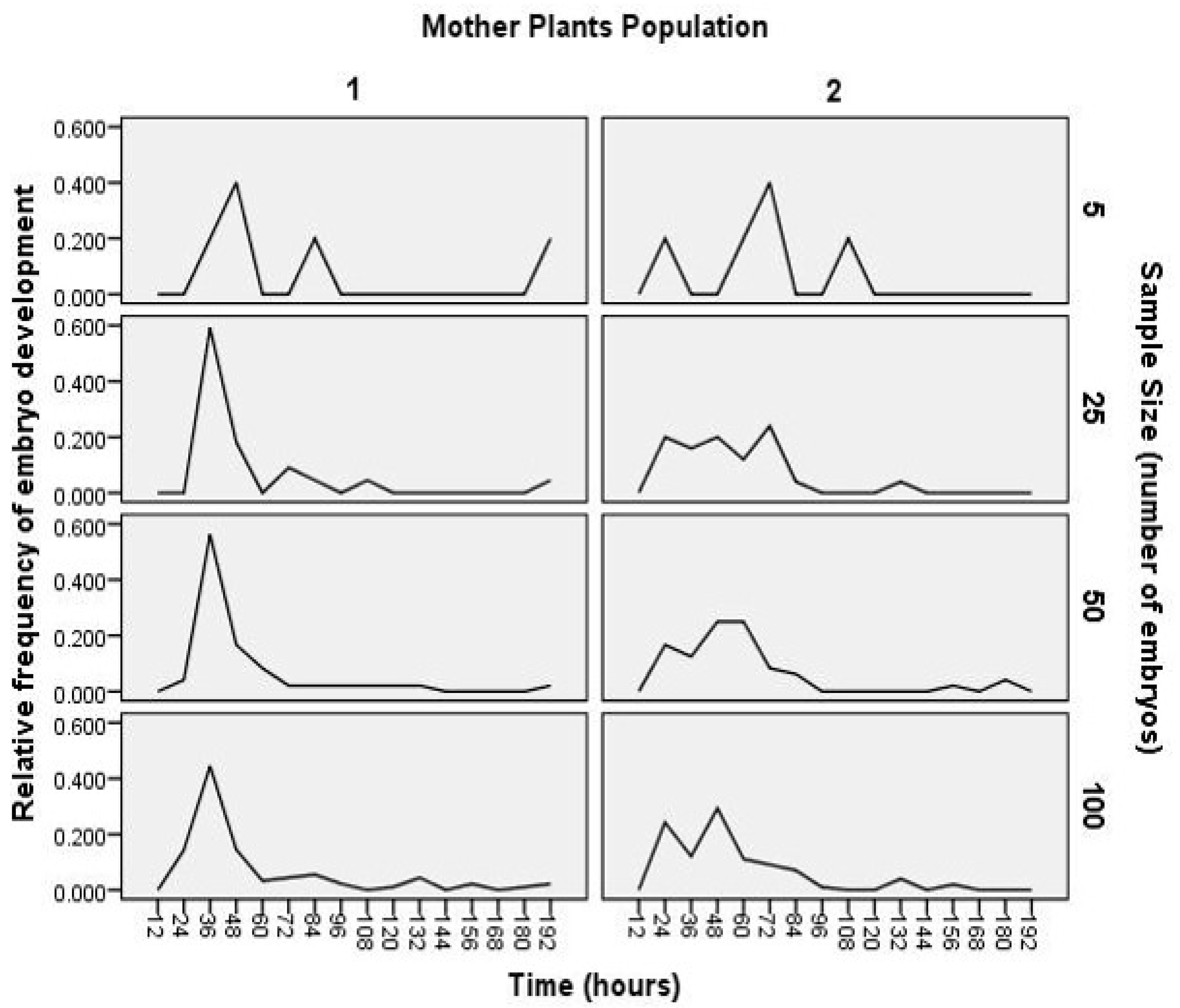
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thorpe, T. History of Plant Tissue Culture. Methods Mol. Biol. 2012, 877, 9–27. [Google Scholar] [PubMed]
- Thorpe, T.A. History of Plant Tissue Culture. Mol. Biotechnol. 2007, 37, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Gamborg, O.L. Plant Tissue Culture. Biotechnology. Milestones. Vitr. Cell. Dev. Biol. Plant 2002, 38, 84–92. [Google Scholar] [CrossRef]
- Davies, K.M.; Deroles, S.C. Prospects for the Use of Plant Cell Cultures in Food Biotechnology. Curr. Opin. Biotechnol. 2014, 26, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Vasil, I.K. A History of Plant Biotechnology: From the Cell Theory of Schleiden and Schwann to Biotech Crops. Plant Cell Rep. 2008, 27, 1423–1440. [Google Scholar] [CrossRef]
- Shahzad, A.; Sharma, S.; Parveen, S.; Saeed, T.; Shaheen, A.; Akhtar, R.; Yadav, V.; Upadhyay, A.; Ahmad, Z. Historical Perspective and Basic Principles of Plant Tissue Culture. In Plant Biotechnology: Principles and Applications; Springer: Singapore, 2017; pp. 1–36. [Google Scholar]
- Ranal, M.A.; De Santana, D.G. How and Why to Measure the Germination Process? Braz. J. Bot. 2006, 29, 1–11. [Google Scholar] [CrossRef]
- Nobel, P.S. Physicochemical and Environmental Plant Physiology; Academic Press: Amsterdam, The Netherlands, 2009. [Google Scholar]
- Hunt, R. Basic Growth Analysis; Springer: Dordrecht, The Netherlands, 1990; ISBN 978-0-04-445373-4. [Google Scholar]
- Roderick, M.L. On the Measurement of Growth with Applications to the Modelling and Analysis of Plant Growth. Funct. Ecol. 2000, 14, 244–251. [Google Scholar] [CrossRef]
- Pérez-Harguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. New Handbook for Standardised Measurement of Plant Functional Traits Worldwide. Aust. J. Bot. 2013, 61, 167. [Google Scholar] [CrossRef]
- Smith, R.H. Plant Tissue Culture: Techniques and Experiments; Academic Press: Cambridge, MA, USA, 1992; ISBN 9780126503401. [Google Scholar]
- Penfield, S. Seed Dormancy and Germination. Curr. Biol. 2017, 27, R874–R878. [Google Scholar] [CrossRef]
- Santana, D.G.; Ranal, M. Análise da Germinação: Um Enfoque Estatístico, 1st ed.; UNB: Brasília, Brasil, 2004. [Google Scholar]
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.M.; Nonogaki, H. Seeds; Springer: New York, NY, USA, 2013; ISBN 978-1-4614-4692-7. [Google Scholar]
- Ribeiro-Oliveira, J.P.; Ranal, M.A. Sample Size in Studies on the Germination Process. Botany 2016, 94, 103–115. [Google Scholar] [CrossRef]
- de Morais, A.R.; de Araújo, A.G.; Pasqual, M.; Peixoto, A.P.B. Estimação Do Tamanho de Parcela Para Experimento Com Cultura de Tecidos Em Videira. Semin. Cienc. Agrar. 2014, 35, 113. [Google Scholar] [CrossRef]
- Peixoto, A.P.B.; Faria, G.A.; Morais, A.R. de Modelos de Regressão Com Platô Na Estimativa Do Tamanho de Parcelas Em Experimento de Conservação in Vitro de Maracujazeiro. Ciên. Rural. 2011, 41, 1907–1913. [Google Scholar] [CrossRef]
- Minardi, B.D.; Voytena, A.P.L.; Randi, A.M.; Zaffari, G.R. Cultivo in Vitro de Embriões Zigóticos de Butia Eriospatha (Mart. Ex Drude) Becc. Insul. Rev. Botân. 2011, 40, 70–81. [Google Scholar]
- Jesus, M.A.S.; Pasqual, M.; Dutra, L.F.; Chagas, E.A. Cultivo in Vitro de Embriões Zigóticos de Jatropha. Ceres 2003, 50, 183–189. [Google Scholar]
- Costa, N.M.S.; Aloufa, M.A.I. Desenvolvimento in Vitro de Embriões Zigóticos de Tamareira. Rev. Ciên. Agronô. 2007, 38, 276–279. [Google Scholar]
- Ribeiro-Oliveira, J.P.; Ranal, M.A.; Garcia De Santana, D.; Pereira, L.A. Sufficient Sample Size to Study Seed Germination. Aust. J. Bot. 2016, 64, 295–301. [Google Scholar] [CrossRef]
- Campos, M.S. Avaliação da Biocompatibilidade da Seiva do Croton Lechleri (Sangue de Dragão) em Tecido Subcutâneo de Ratos; Universidade Estadual Paulista: Araçatuba, Brazil, 2009. [Google Scholar]
- Smith, B.A. A New Species of Croton (Euphorbiaceae) from Ecuador. Novon 2006, 16, 273–274. [Google Scholar] [CrossRef]
- Caruzo, M.B.R.; Cordeiro, I. Sinopse Da Tribo Crotoneae Dumort. (Euphorbiaceae s.s.) No Estado de São Paulo, Brasil. Hoehnea 2007, 34, 571–585. [Google Scholar] [CrossRef]
- Medonça, M.N.S.; Almeida, M.; Firmino, J.L.; Ramos, L.B.; Santos, A.M. Biometria Dos Frutos e Sementes de Sangue de Grado Croton Lechleri Muell. Arg Euphorbiaceae. In Proceedings of the Sociedade Brasileira para o Progresso da Ciência 66; Sociedade Brasileira para o Progresso da Ciência: Rio Branco, Brazil, 2014. [Google Scholar]
- Azevedo, K.; Alechandre, A.; Lima, A.; Leite, A.; Melo, T.; Costa, J.; Pereira, M.A.; Campos, C.A.; Lima, A. Guia Para a Extração de Sangue de Grado (Croton Lechleri Müll. Arg.): Recomendações Técnicas Para a Extração de Látex de Sangue de Grado (Sangue de Dragão). 2013. Available online: https://ipam.org.br/wp-content/uploads/2008/04/guia_para_a_extrac%CC%A7a%CC%83o_de_sangue_de_grado_.pdf (accessed on 7 June 2022).
- Fayad, W.; Fryknäs, M.; Brnjic, S.; Olofsson, M.H.; Larsson, R.; Linder, S. Identification of a Novel Topoisomerase Inhibitor Effective in Cells Overexpressing Drug Efflux Transporters. PLoS ONE 2009, 4, e7238. [Google Scholar] [CrossRef]
- De Marino, S.; Gala, F.; Zollo, F.; Vitalini, S.; Fico, G.; Visioli, F.; Iorizzi, M. Identification of Minor Secondary Metabolites from the Latex of Croton Lechleri (Muell-Arg) and Evaluation of Their Antioxidant Activity. Molecules 2008, 13, 1219–1229. [Google Scholar] [CrossRef]
- Montopoli, M.; Bertin, R.; Chen, Z.; Bolcato, J.; Caparrotta, L.; Froldi, G. Croton Lechleri Sap and Isolated Alkaloid Taspine Exhibit Inhibition against Human Melanoma SK23 and Colon Cancer HT29 Cell Lines. J. Ethnopharmacol. 2012, 144, 747–753. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, T.G.; Pereira, A.M.S.; Coppede, J.S.; França, S.C.; Ming, L.C.; Bertoni, B.W. Genetic Diversity Analysis of Croton Antisyphiliticus Mart. Using AFLP Molecular Markers. Genet. Mol. Res. 2016, 15, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro-Oliveira, J.P.; Ranal, M.A. Brazilian Forest Seeds: A Precarious Beginning, a Heady Present and the Future, Will It Be Promising?|Sementes Florestais Brasileiras: Início Precário, Presente Inebriante e o Futuro, Promissor? Cienc. Florest. 2014, 24, 771–784. [Google Scholar] [CrossRef]
- Quoirin, M.; Lepoivre, P. Etude de Milieux Adaptes Aux Cultures in Vitro de Prunus. Acta Hortic. 1977, 78, 437–442. [Google Scholar] [CrossRef]
- Machado, M.P.; Biasi, L.A.; Ritter, M.; Ribas, L.L.F.; Koehler, H.S.; Zanette, F. Meios de Cultura Na Micropropagação Do Porta-Enxerto de Videira “VR043-43” (Vitis Vinifera × Vitis Rotundifolia). Ciên. Rural. 2007, 37, 277–280. [Google Scholar] [CrossRef]
- Ribeiro-Oliveira, J.P.; Silveira, L.E.D.; Pinto, L.V.A.; Silva, E.A.A.; Hilhorst, H.W.M. Clues on an Intraspecific Communication System in Seed-Seedling Transition. Physiol. Plant 2021, 172, 1609–1618. [Google Scholar] [CrossRef]
- Neter, J.; Kutner, M.H.; Nachtsheim, C.J.; Wasserman, W. Applied Linear Statistical Models; Irwin: Chicago, IL, USA, 1985; Volume 4. [Google Scholar]
- Roach, D.A.; Wulff, R.D. Maternal Effects in Plants. Annu. Rev. Ecol. Syst. 1987, 18, 209–235. [Google Scholar] [CrossRef]
- Fenner, M. The Effects of the Parent Environment on Seed Germinability. Seed Sci. Res. 1991, 1, 75–84. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. How Much Influence Does the Paternal Parent Have on Seed Germination? Seed Sci. Res. 2019, 29, 1–11. [Google Scholar] [CrossRef]
- Labouriau, L.G.A. Germinação de Sementes; Secretaria Geral da Organização dos Estados Americanos: Caracas, Venezuela, 1983. [Google Scholar]
- Labouriau, L.G.A. On the Physiology of Seed Germination in Vicia Graminea I. Acad. Bras. Cienc. 1970, 42, 235–262. [Google Scholar]
- Labouriau, L.G.; Valadares, M.B. On the Physiology of Seed of Calotropis Procera. An. da Acad. Bras. de Ciên. 1976, 42, 235–264. [Google Scholar]
- Primack, R.B. Variation in the Phenology of Natural Populations of Montane Shrubs in New Zealand. J. Ecol. 1980, 68, 849. [Google Scholar] [CrossRef]
- Ranal, M.A.; de Santana, D.G.; Ferreira, W.R.; Mendes-Rodrigues, C. Calculating Germination Measurements and Organizing Spreadsheets. Rev. Bras. de Botân. 2009, 32, 849–855. [Google Scholar] [CrossRef]
- Ribeiro-Oliveira, J.P.; de Santana, D.G.; Pereira, V.J.; dos Santos, C.M. Data Transformation: An Underestimated Tool by Inappropriate Use. Acta Sci. Agron. 2018, 40, 1–11. [Google Scholar] [CrossRef]
- Penfield, S.; MacGregor, D.R. Effects of Environmental Variation during Seed Production on Seed Dormancy and Germination. J. Exp. Bot. 2017, 68, 819–825. [Google Scholar] [CrossRef] [PubMed]
- Mcnair, J.N.; Sunkara, A.; Frobish, D. How to Analyse Seed Germination Data Using Statistical Time-to-Event Analysis: Non-Parametric and Semi-Parametric Methods. Seed Sci. Res. 2012, 22, 77–95. [Google Scholar] [CrossRef]
- Waterworth, W.M.; Footitt, S.; Bray, C.M.; Finch-Savage, W.E.; West, C.E. DNA Damage Checkpoint Kinase ATM Regulates Germination and Maintains Genome Stability in Seeds. Proc. Natl. Acad. Sci. USA 2016, 113, 9647–9652. [Google Scholar] [CrossRef]
- Ajala-Luccas, D.; Ribeiro-Oliveira, J.P.; Duarte Silveira, L.E.; da Silva, E.A.A. An Integrative Insight on Dormancy Alleviation in Diaspores of Urochloa Humidicola (Rendle) Morrone & Zuloaga, a Tropical Grass with Great Economic and Ecological Impact. Plant Biol. 2018, 20, 252–262. [Google Scholar] [CrossRef]
- Silveira, L.E.D.; Ribeiro-Oliveira, J.P.; Ajala-Luccas, D.; Bravo, J.P.; da Silva, E.A.A. A Molecular Framework for the Embryo Growth in Germinating Seeds of Solanum Lycocarpum A. St.-Hil., a Nurse Plant Species. Ann. Appl. Biol. 2019, 175, 136–145. [Google Scholar] [CrossRef]
- de Souza, B.L.; Ribeiro-Oliveira, J.P.; Bravo, J.P.; Dias, G.F.; da Silva, E.A.A. What Happens When the Rain Is Back? A Hypothetical Model on How Germination and Post-Germination Occur in a Species from Transient Seed Banks. PLoS ONE 2020, 15, e0229215. [Google Scholar] [CrossRef]
- Ferreira, W.R.; Ranal, M.A.; de Santana, D.G.; Nogueira, A.P.O. Germination and Emergence Measurements Could Group Individuals and Species? Braz. J. Bot. 2015, 38, 457–468. [Google Scholar] [CrossRef]
- ISTA. International Rules for Seed Testing 2019. 2019. Available online: https://www.seedhealth.org/files/2019/02/ISTARules2019SHmethods7-019a-opt.pdf (accessed on 7 June 2022).
- Peet, R.K. Relative Diversity Indices. Ecology 1975, 56, 496–498. [Google Scholar] [CrossRef]
- Meier, V.D.; Lessman, K.J. Estimation of Optimum Field Plot Shape and Size for Testing Yield in Crambe Abyssinica Hochst. Crop Sci. 1971, 11, 648–650. [Google Scholar] [CrossRef]
- Ribeiro-Oliveira, J.P.; Ranal, M.A. Sample Size and Water Dynamics on Germinating Diaspores: The First Step for Physiological and Molecular Studies on the Germination Process. Plant Biosyst. 2018, 152, 840–847. [Google Scholar] [CrossRef]
- de Souza, R.A.; Hungria, M.; Franchini, J.C.; Chueire, L.M.d.O.; Barcellos, F.G.; Campo, R.J. Avaliação Qualitativa e Quantitativa Da Microbiota Do Solo e Da Fixação Biológica Do Nitrogênio Pela Soja. Pesqui. Agropecu. Bras. 2008, 43, 71–82. [Google Scholar] [CrossRef]
- Ribeiro-Oliveira, J.P.; Ranal, M.A.; de Santana, D.G. A Amplitude Amostral Interfere Nas Medidas de Germinação de Bowdichia Virgilioides Kunth? Cienc. Florest. 2013, 23, 623–634. [Google Scholar] [CrossRef]
- Pinto, L.V.A.; Da Silva, E.A.A.; Davide, A.C.; De Jesus, V.A.M.; Toorop, P.E.; Hilhorst, H.W.M. Mechanism and Control of Solanum Lycocarpum Seed Germination. Ann. Bot. 2007, 100, 1175–1187. [Google Scholar] [CrossRef]
- Dorneles, M.C.; Ranal, M.A.; de Santana, D.G. Germinação de Sementes e Emergência de Plântulas de Anadenanthera Colubrina (Vell.) Brenan Var. Cebil (Griseb.) Altschut, Fabaceae, Estabelecida Em Fragmentos Florestais Do Cerrado, MG. Ciên. Florest. 2013, 23, 291–304. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Statistics | Characters | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (Day) | CVt (%) | U (Bit) | Z | ED (%) | (Day−1) | ||||||||||
| ANOVA Assumptions | W | 0.924 | 0.951 | 0.983 | 0.951 | 0.962 | 0.927 | ||||||||
| (P) | (0.010) | (0.084) | (0.787) | (0.080) | (0.193) | (0.012) | |||||||||
| 1F | 2.688 | 2.059 | 3.200 | 1.635 | 4.721 | 2.323 | |||||||||
| (P) | (0.26) | (0.78) | (0.011) | (0.161) | (0.001) | (0.049) | |||||||||
| 2F | 3.666 | 3.592 | 0.648 | 0.565 | 20.848 | 1.790 | |||||||||
| (P) | (0.066) | (0.069) | (0.428) | (0.459) | (0.000) | (0.192) | |||||||||
| Model | Source of Variation | DF | MS | 3F (P) | MS | 3F (P) | MS | MS | 3F (P) | DF | MS | 3F (P) | MS | 3F (P) | |
| Factorial ANOVA | Sample Size | 2 | 0.022 | 0.740 (0.5372) | 0.619 | 28.427 (0.000) | 3.257 | 419.583 (0.0000) | 0.943 | 19.882 (0.0000) | 3 | 0.150 | 1.261 (0.3068) | 0.0002 | 0.273 (0.8442) |
| Mother Plant Population | 1 | 0.002 | 0.051 (0.8222) | 0.046 | 2.117 (0.1568) | 0.022 | 2.846 (0.1027) | 0.240 | 5064 (0.0325) | 1 | 0.486 | 4.085 (0.0529) | 0.00004 | 0.066 (0.7997) | |
| Sample Size × Mother Plant Population | 2 | 0.005 | 0.152 (0.9276) | 0.032 | 1.1492 (0.2383) | 0.010 | 1.300 (0.2940) | 0.072 | 1.521 (0.2307) | 3 | 0.171 | 1.440 (0.2520) | 0.0001 | 0.142 (0.9340) | |
| Block | 4 | 0.048 | - | 0.039 | - | 0.011 | - | 0.038 | - | 4 | 0.003 | - | 0.008 | - | |
| Error | 20 | 0.030 | - | 0.022 | - | 0.008 | - | 0.047 | - | 28 | 0.019 | 0.006 | |||
| CV (%) | 7.79 | 41.14 | 10.40 | 49.28 | - | - | 1.89 | 18.16 | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Albuquerque, J.P.; Ribeiro-Oliveira, J.P.; de Oliveira-Júnior, J.B.; da Silva Costa, F.H.; Amaral da Silva, E.A. Understanding In Vitro Embryo Development through Classical Germination Measurements: A Case Study of Dragon’s Blood (Croton lechleri Müll Arg.). Agriculture 2023, 13, 1618. https://doi.org/10.3390/agriculture13081618
de Albuquerque JP, Ribeiro-Oliveira JP, de Oliveira-Júnior JB, da Silva Costa FH, Amaral da Silva EA. Understanding In Vitro Embryo Development through Classical Germination Measurements: A Case Study of Dragon’s Blood (Croton lechleri Müll Arg.). Agriculture. 2023; 13(8):1618. https://doi.org/10.3390/agriculture13081618
Chicago/Turabian Stylede Albuquerque, Janai Pereira, João Paulo Ribeiro-Oliveira, João Bosco de Oliveira-Júnior, Frederico Henrique da Silva Costa, and Edvaldo Aparecido Amaral da Silva. 2023. "Understanding In Vitro Embryo Development through Classical Germination Measurements: A Case Study of Dragon’s Blood (Croton lechleri Müll Arg.)" Agriculture 13, no. 8: 1618. https://doi.org/10.3390/agriculture13081618