
Regulation of bta-miRNA29d-3p on Lipid Accumulation via GPAM in Bovine Mammary Epithelial Cells

1
Tianjin Key Laboratory of Animal Molecular Breeding and Biotechnology, Tianjin Engineering Research Center of Animal Healthy Farming, Institute of Animal Science and Veterinary, Tianjin Academy of Agricultural Sciences, Tianjin 300381, China
2
College of Life Science, Nankai University, Tianjin 300071, China
3
College of Animal Science and Technology, Henan University of Animal Husbandry and Economy, Zhengzhou 450046, China
4
College of Animal Science and Animal Medicine, Tianjin Agricultural University, Tianjin 300384, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Agriculture 2023, 13(2), 501; https://doi.org/10.3390/agriculture13020501
Submission received: 9 December 2022
/
Revised: 13 February 2023
/
Accepted: 16 February 2023
/
Published: 20 February 2023
(This article belongs to the Special Issue Breeding, Genetics and Safety Production of Dairy Cattle)
Abstract
:MicroRNAs (miRNAs) are small RNA molecules consisting of approximately 22 nucleotides that are engaged in the regulation of various bio-processes. There is growing evidence that miR-29 is a key regulator of hepatic lipid metabolism. Mimics and inhibitors of bta-miRNA29d-3p were transiently transfected in bovine mammary epithelial cells (BMECs) to reveal the regulation of bta-miRNA29d-3p on lipid accumulation in BMECs. Results showed that overexpression of bta-miRNA29d-3p significantly inhibited the expression of genes related to triglyceride (TAG) synthesis, namely DGAT1 and mitochondrial glycerol-3-phosphate acyltransferase (GPAM, p < 0.01) and down-regulated TAG levels in cells (p < 0.05). The expression of fatty acid synthesis and desaturation-related genes FASN, SCD1, and ACACA, and transcription factor SREBF1 also decreased. Interference of bta-miRNA29d-3p significantly increased the expression of GPAM, DGAT1, FASN, SCD1, ACACA, and SREBF1 (p < 0.01), and significantly upregulated the concentration of TAG in cells. Furthermore, a luciferase reporter assay confirmed that GPAM is a direct target of bta-miRNA29d-3p. In summary, bta-miRNA29d-3p modulates fatty acid metabolism and TAG synthesis by regulating genes related to lipid metabolism in BMECs and targeting GPAM. Thus, bta-miRNA29d-3p plays an important role in controlling mammary lipid synthesis in cows.
1. Introduction
Milk has gradually become an indispensable part of the daily diet of humans. Milk is high in fat, and it contains unsaturated fatty acids that are effective in preventing high blood cholesterol and cardiovascular disease in humans [1]. The composition and content of fatty acids are also strong correlates of the nutrition of milk [2]. Increasing lipid content and ameliorating lipid composition could help improve milk quality. One of the essential organs of lactating animals is the breast. Its development and lactation are regulated by cytokines, hormones, and some natural biological functional substances [3]. MicroRNA (miRNA) research in terms of breast development and secretion control in mammals (cows, mice, etc.) has also progressed to a certain extent.
MiRNAs are small, siRNA-like molecules that bind to targeted mRNAs, inhibiting their translation and accelerating their degradation at the post-transcriptional level [4]. MiRNA regulates the biometabolism of most lactating animal tissues, such as the liver, fat, and skeletal muscle in humans, dairy cows, and dairy goats, as well as in mice [5]. The contributions of miRNAs to milk gland evolution are mainly achieved by regulating the proliferation and differentiation of mammary epithelial cells, the differentiation of mammary stem cells, and the development of glandular vesicles and milk ducts [6]. The miR-29 family mainly includes miR-29a, miR-29b, miR-29c, miR-29d-3p, and miR-29e. Reports on the structure, function, and regulation of the miR-29 family are mainly focused on research related to humans and mice at present.
In ruminants, miR-29 was shown to target PTX3 in goat granulosa cells through activation of PI3K/AKT/mTOR and Erk1/2 signaling pathways to stimulate granulosa cell multiplication and suppress steroidogenesis and apoptosis [7]. However, studies in non-ruminants have shown that miR-29 not only participates in fatty acid oxidation but also leads to the synthesis of TAG and, thus, performs an instrumental role in controlling fatty acid metabolism. He et al. used microarray technology to study rat skeletal muscle and discovered that miR-29 expression was significantly higher in a type II diabetic group than in healthy rats [8]. In addition, reduced fatty acid content and significantly decreased plasma cholesterol and triglyceride (TAG) levels were observed in the livers of miR-29 knockdown mice [9]. These findings offer the possibility that bta-miRNA29d-3p regulates fatty acid composition and TAG content in dairy cows.
MiRNAs have become significant regulators of glycolipid metabolism among many tissues [5]. However, to date, studies on bta-miRNA29d-3p in the dairy mammary gland have not been reported. This study explored the effect of bta-miRNA29d-3p on the expression of genes related to lipid production and TAG content in mammary epithelial cells of dairy cows and revealed its role in regulating milk lipids.
2. Materials and Methods
2.1. Cell Culture and Transfection
The BMECs used in this experiment were provided by Mr. Chen Zhi from the College of Animal Science and Technology, Yangzhou University. The mammary gland tissue samples used for BMEC isolation were collected from three healthy Holstein cows (4 years old, second parity, non-pregnant) at peak lactation. The psiCHECK-2 vector (psiCHECK-2 plasmid) was presented by Professor Jun Luo from Northwest A&S University.
On the basis of the available literature [10], BMECs were cultured by 60 mm cell dishes in a 5% CO2 incubator at 37 °C. The medium contained 10% fetal bovine serum, 5 μg/mL insulin, 100 U/mL penicillin and streptomycin, 10 ng/mL epidermal growth factor, 1 μg/mL hydrocortisone, and 90% DMEM/F12. The medium was replaced once in 24 h.
When the cell confluence reached 80%, the cells were transfected with Lipofectamine® RNAiMAX Transfection Reagent, and the transfection complex was prepared by referring to the product’s instructions. The total volume of transfection complexes was 100 μL per well. miR-29d mimic (miR-29d-3p) and its negative control (Con miR) and miR-29d inhibitor (anti-miR-29d-3p) and its negative control (Con Inh) were transiently transfected into mammary epithelial cells at a final concentration of 50 nM, and the remaining volume was replenished with DMEM/F-12. The transfection complexes were mixed well and incubated at room temperature for 20 min. The cultured BMECs were digested into separate suspensions and added to a 24-well plate. Finally, the incubated transfection complexes were added drop-wise and incubated at 37 °C in a 5% CO2 incubator. After 48 h, intracellular miR-29d-3p mRNA levels were examined using real-time fluorescence quantification (RT-qPCR). Each treatment was performed in three biological replicates.
2.2. RNA Extraction and Real-Time Quantitative PCR
Cells were lysed to carry out RNA extraction after 48 h of transfection, and the RNA extraction method was carried out using the cultured cell/bacterial total RNA extraction kit from Tiangen Biochemical Technology (Beijing, China) Co., Ltd. (DP430). The extracted RNA was reverse transcribed with the PrimeScriptTM RT kit (Perfect Real time, Takara Bio Inc., Kusatsu, Japan), where the bta-miRNA29d-3p reverse transcription primers were designed using the stem–loop method. The bta-miRNA29d-3p reverse-transcribed cDNA was used as a template, and the 18s gene was used as an internal reference gene for RT-qPCR amplification of the gene by using SYBR Green (SYBR Premix Ex Taq II, Perfect Real Time, Takara Bio Inc., Kusatsu, Japan). Then, a 20 μL system was designed as follows: SYBR GreenMix, 10 μL; PCR Forward Primer, 0.8 μL; PCR Reverse Primer, 0.8 μL; DNA template, 2 μL; and sterilized water, 6.0 μL. The PCR reaction program was 40 cycles of 95 °C pre-denaturation for 30 s, 95 °C for 5 s, and 60 °C for 30 s.
The genes detected included fatty acid synthase (FASN), glycerol-3-phosphate acyltransferase (GPAM), peroxisome proliferator-activated receptor γ (PPARG1), diacylglycerol acyltransferase 1 (DGAT1), sterol regulatory element-binding factor 1 (SREBF1), liver X receptor alpha (LXRA), and acetyl-coenzyme A carboxylase alpha (ACACA). The internal controls were ubiquitously expressed transcript (UXT) and mitochondrial ribosomal protein L39 (MRPL39). The sequences of primers with qPCR are shown in Table 1.
2.3. Detection of Triglycerides in BMECs
The transfection operation was the same as above. Analyses were performed in triplicate. After the cells were transfected for 48 h and replaced with new medium, the cells were washed twice with PBS, and 1 mL of lysis buffer was added to each well. An appropriate amount of lysate was reserved for BCA protein quantitative determination. TAG content was detected and calculated in accordance with the instructions of the TAG test kit from Beijing Applygen. A TAG standard curve was constructed to specify the triglyceride content of the samples, after which the TAG content was corrected for total cellular protein concentration per mg.
2.4. Construction of psiCHECK-2-GPAM 3′UTR Recombinant Plasmid
To generate a reporter gene for luciferase assays, we amplified bovine GPAM 3′-UTR fragments by PCR and cloned them into the XhoI/NotI digest site of the dual-luciferase reporter gene vector psiCHECK-2 to construct a dual-luciferase reporter gene vector containing the wild-type bovine GPAM 3′UTR sequence. Meanwhile, a dual-luciferase expression vector containing a mutated GPAM 3′-UTR sequence was constructed by overlap PCR. The primer sequences are given in Table 2. All constructs were confirmed by sequencing.
2.5. Dual Luciferase Assays
After mammary epithelial cells were transfected for 48 h, they were lysed and a dual-luciferase assay was performed by referring to the Promega dual reporter gene assay kit. The cells were washed twice using PBS. In each well of the 24-well opaque assay plate, 65 μL of cell lysate was added. The firefly luciferase activity value (F value) was first obtained, and then the firefly luciferase activity was terminated, the Renilla luciferase activity was activated, and the Renilla luciferase activity (R value) was read. Relative luciferase activity = R value/F value, which is the relative transcriptional activity.
2.6. Statistical Analysis
All data were analyzed in SAS 9.4 (Cary, NC, USA) using reusable two-factor analysis of variance and expressed as mean ± standard error of the mean (SEM). Each experiment was repeated at least three times. Relative quantitative analysis was performed using the 2−ΔΔCt method, and the data were analyzed for significance. p < 0.05 indicated a significant difference (*, p < 0.05; **, p < 0.01).
3. Results
3.1. Detection of Overexpression and Interference Efficiency of bta-miRNA29d-3p in BMECs
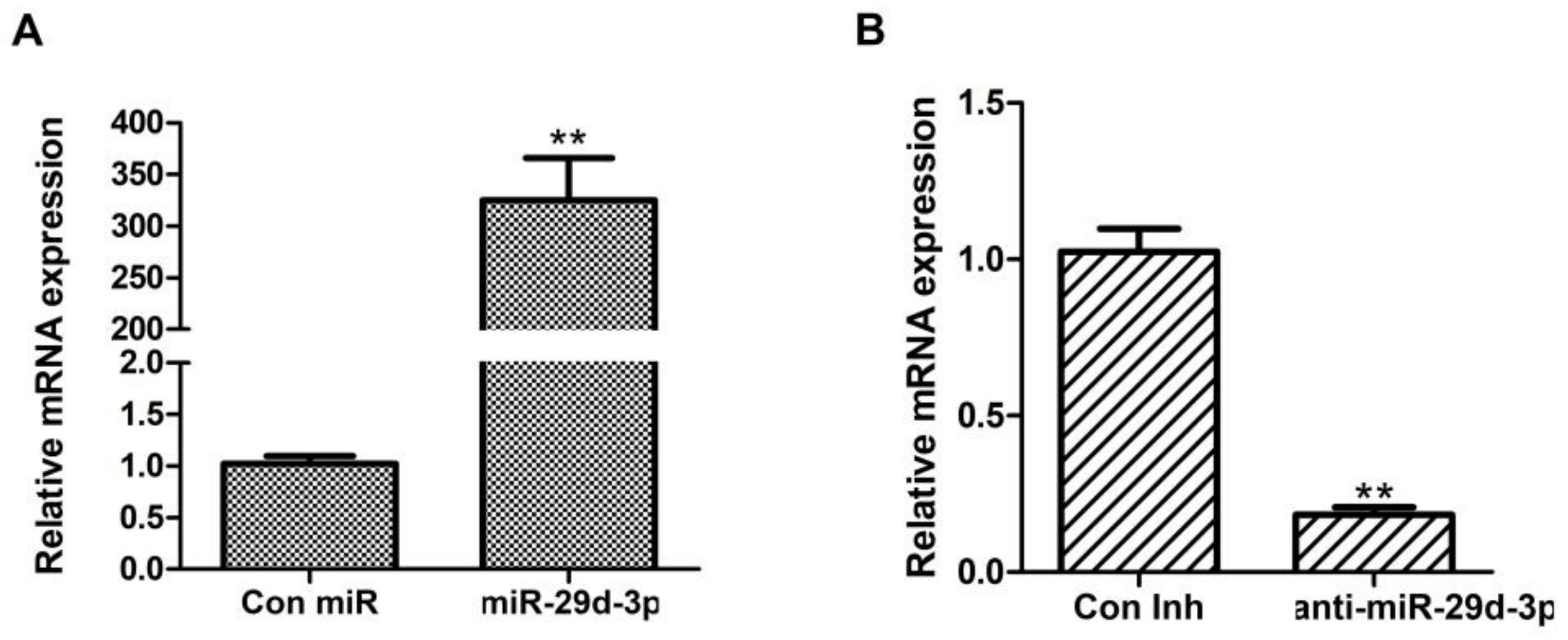
According to Figure 1, the expression of bta-miRNA29d-3p increased by approximately 300-fold compared with that of Con miR (p < 0.01, Figure 1A), and the expression of anti-miR-29d-3p significantly decreased by approximately 80% compared with that of Con Inh (p < 0.01, Figure 1B). This result indicated that miR-29d-3p and anti-miR-29d-3p were transfected with good efficiency.
3.2. Overexpression of bta-miRNA29d-3p Affects the Expression of Genes Related to Lipid Metabolism in BMECs
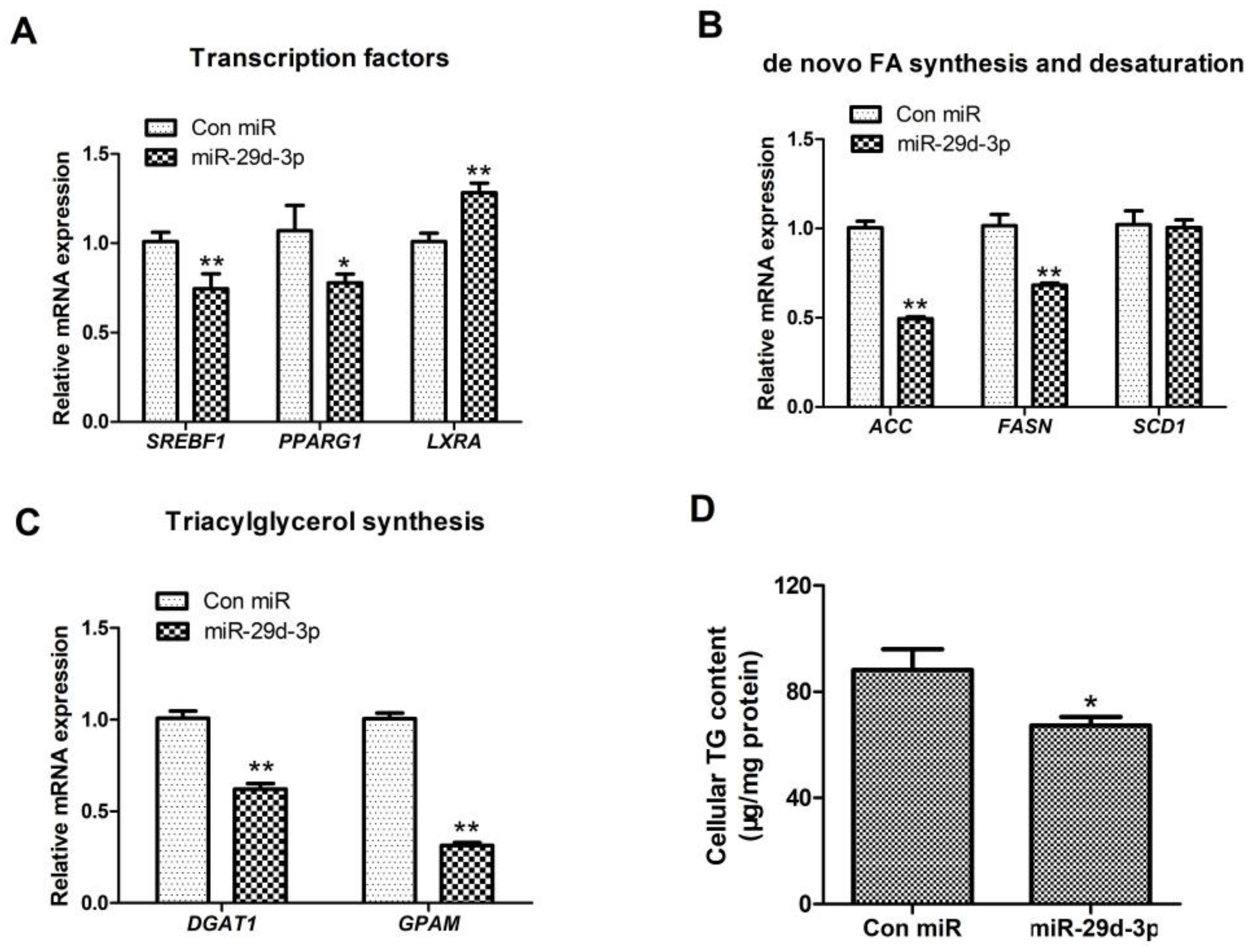
On the basis of Figure 2, after bta-miRNA29d-3p was overexpressed and the expression of SREBF1 was clearly decreased, whereas that of LXRA was markedly upregulated (p < 0.01, Figure 2A). The synthesis and desaturation of fatty acid-related gene (ACACA, FASN, and SCD1) expression were remarkably reduced (p < 0.01, Figure 2B), and that of TAG synthesis-related genes DGAT1 and GPAM was significantly down-regulated (p < 0.01, Figure 2C).
3.3. Interference of bta-miRNA29d-3p Promotes the Expression of Genes Related to Lipid Metabolism in BMECs
3.4. Effect of bta-miRNA29d-3p on TAG Content in BMECs
3.5. Prediction of bta-miRNA29d-3p Target Genes
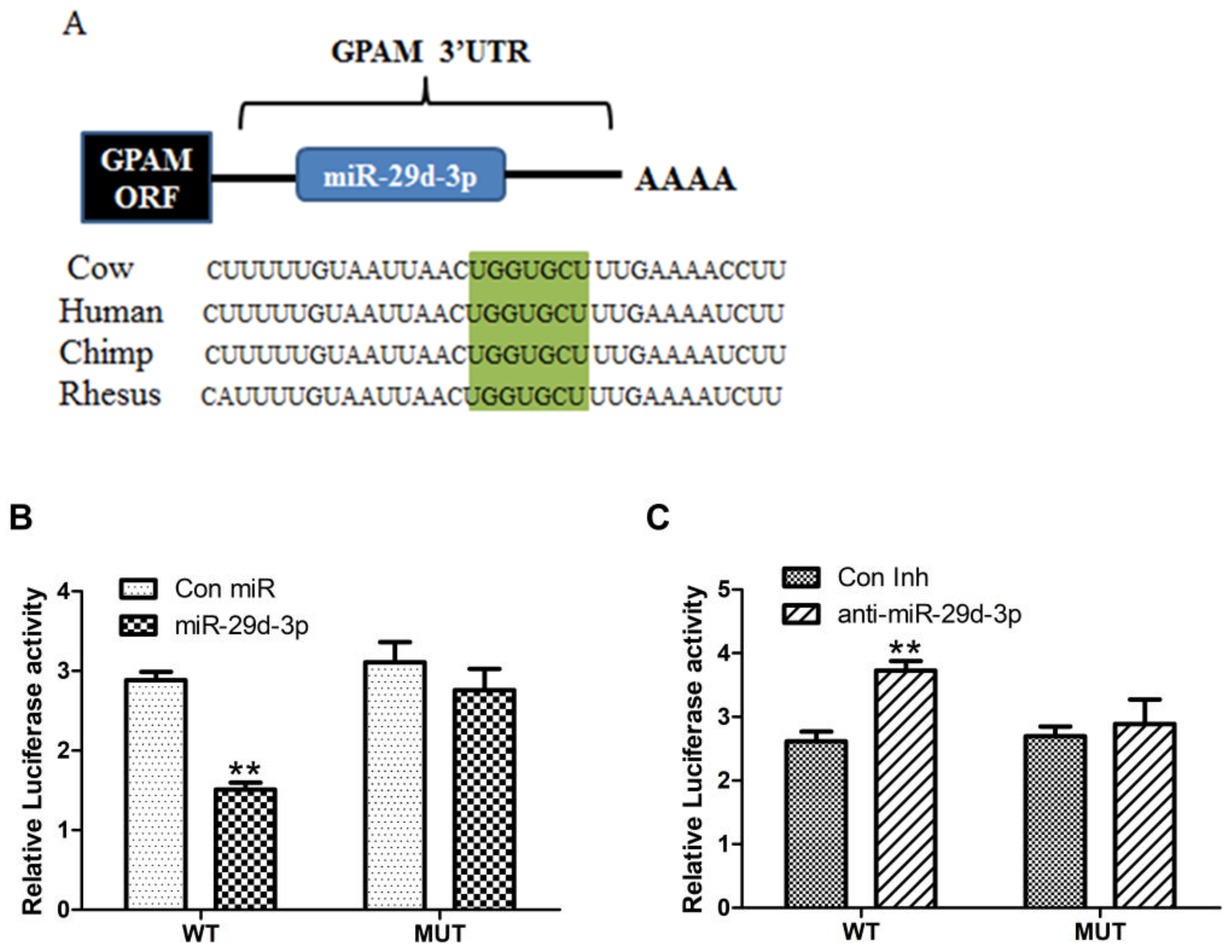
As shown in Figure 4, the target gene associated with fatty acid metabolism was GPAM, and bta-miRNA29d-3p was more conserved in human and chimpanzee sequences (Figure 4A). When the GPAM 3′-UTR was wild-type, bta-miRNA29d-3p overexpression significantly decreased the activity of the dual-luciferase reporter gene (p < 0.01, Figure 4B), whereas interference with the bta-miRNA29d-3p gene significantly increased the activity of the dual-luciferase reporter (p < 0.01, Figure 4C). When the GPAM 3′-UTR was mutant, bta-miRNA29d-3p showed an absence of effect towards the activity of dual-luciferase reporter gene expression. Therefore, bta-miRNA29d-3p acts directly on the cow GPAM 3′-UTR target site.
4. Discussion
The miR-29 family plays a core role in the etiology and pathogenesis of osteoarthritis (OA), osteoporosis, cardiac and renal diseases, and immune disorders [17]. Furthermore, miR-29 participates in the modulation of multiple cellular functions and in lipid metabolism [7,18,19]. In this study, combined with previous research of miR-29’s effects on lactation in ruminants, the hypothesis was that bta-miRNA29d-3p has an effect on lipid metabolism in BMECs. In vitro experiments confirmed that pharmacological inhibition of miR-29 significantly down-regulated hepatic cholesterol and adipose synthesis de novo [9]. Moreover, miR-29 was found to be an important regulator of lipid oxidation by overexpression or deletion in human or primary human skeletal muscle cells [5]. Consequently, we investigated the regulatory mechanism of bta-miRNA29d-3p on milk lipids to provide a basis for further improvement of lactation performance of cows and their progeny.
Transcription factors (TF) are an important class of proteins involved in the regulation of gene transcription. They are key regulators of various signaling pathways. In this study, overexpression of bta-miRNA29d-3p dramatically down-regulated the expression of SREBF1, a member of a family of TFs that binds to the ER and acts centrally involved in fatty acid, phospholipid, and cholesterol synthesis [20]. Overexpression of nSREBP-1a or -1c was suggested to significantly upregulate the expression of its target genes ACC, FASN, and stearoyl coenzyme A desaturase 1 (SCD1) and remarkably enhanced the pri-miR-29 and mature miR-29 expression in glioblastoma (GBM) cells [21]. Overall, miR-29 suppressed SREBP-1 and the lipid synthesis pathway in GBM cells as a negatively regulated factor. The results of this research are in agreement with the findings reported in the literature above. Accordingly, bta-miRNA29d-3p may control lipid metabolism-related gene expression through SREBF1, which consequently impacts fatty acid synthesis.
Interestingly, we found extremely significant upregulation of PPARG1 expression by interfering with bta-miRNA29d-3p. However, it has been demonstrated that inhibition of miR-29s can cause hypermethylation of overall DNA and augments the levels of methylation in the promoters of lactation-associated genes, including PPARG [22]. The peroxisome proliferator-activated receptor (PPAR) family is an important member of the nuclear hormone receptor superfamily, and their physiological functions are mainly associated with fatty acid metabolism, glucose metabolism, and cell proliferation and differentiation [16]. There is evidence that PPARA controls not only fatty acid oxidation-related genes, but also other genes concerned with its synthesis, including ACACA, FASN, and SCD1 [16]. While PPARγ is expressed at a high level in adipose tissue, it is a key coordinator of the transcriptional cascade response for adipocyte differentiation [23]. There are two homotypes of PPARG, PPARG1 and PPARG2, which are splice variants of the PPARG gene via alternative promoters [24]. The results of the in vitro culture of mammary epithelial cell lines showed that PPARG-specific agonist ROSI treatment of mammary epithelial cells was able to upregulate fatty acid metabolism-related genes, such as ACACA, FASN, DGAT1, and SREBF1, and clarified that PPARG directly regulates the expression of some genes in mammary tissue and affects the fatty acid metabolic network [25,26,27]. To summarize the above reports, we presume that bta-miRNA29d-3p may have a vital effect in the control of lipid metabolism in BMECs by affecting the expression of PPARG.
Changes in DGAT1 expression were also found in the current study. DGAT1 is found to be expressed at a high level in adipose tissue, the liver, and the small intestine, all of which are more active in TAG synthesis [28]. For dairy cows, the non-conservative substitution of alanine for lysine in the DGAT1 gene could have a significant effect on milk composition and yield. Evidence showed that mice with DGAT1 gene deletion have reduced absorption of triacylglycerols [29]. Through interference with bta-miRNA29d-3p expression, DGAT1 mRNA expression was significantly increased, along with an increase in intracellular TAG content. A combination of the above series of studies on lipid metabolism by bta-miRNA29d-3p showed that the synergistic effect of bta-miRNA29d-3p and DGAT1 affects lipid synthesis in BMECs.
Screening and identifying miRNAs and their target genes are the key steps in studying the functional mechanism of miRNAs. The bioinformatics approach could provide theoretical guidance for the identification of miRNA target genes, and it has an essential role in studying the functional mechanisms of miRNAs. In this study, GPAM was identified as a target gene related to lipid metabolism by online study of the bta-miRNA29d-3p gene. GPAM is the limiting enzyme in the first step of TAG synthesis, and intracellular TAG levels are regulated by GPAM enzyme content. Knockdown of GPAM significantly reduced the expression of genes associated with TAG synthesis and lipid metabolism in bovine embryonic fibroblasts [30]. Furthermore, GPAM plays a dynamic role in TAG synthesis. For example, a 10-fold increase in GPAM activity in 3T3-L1 adipocyte differentiation and a 5-fold increase in GPAM activity during TAG synthesis were observed in neonatal liver hepatocytes [31]. The present findings on TAG synthesis have mainly focused on both PPAR and SREBP regarding the transcriptional regulation of the gene, but there are also reports confirming that GPAM may have a greater regulatory role on TAG synthesis [31,32]. To summarize, GPAM, as a target gene of bta-miRNA29d-3p, performs a key role in TAG regulation in cells, and its mutation can be used as an effective marker for selective breeding of dairy cows.
Taken together, the bta-miRNA29d-3p gene has a significant function in the control of lipid accumulation in dairy cows, and its regulatory mechanism is linked to the expression of lipid synthesis-associated genes in BMECs. However, the specific regulatory mechanism of the bta-miRNA29d-3p gene on lipid synthesis in mammary epithelial cells of dairy cows needs to be studied in depth. Additional studies on relevant non-coding RNAs in the mammary lipid metabolism pathway in dairy cows could help to better utilize functional miRNAs to regulate the fatty acid composition of milk in the future, which has important application value for dairy development.
5. Conclusions
In this study, overexpression of bta-miRNA29d-3p down-regulated the expression of DGAT1 and GPAM, which are related to TAG synthesis (p < 0.01), and that of FASN, ELOVL4, ACACA, and SREBF1, which are related to FA synthesis and prolongation, significantly down-regulated intracellular TAG levels (p < 0.05). Interfering with the expression of the bta-miRNA29d-3p gene showed the opposite result. The downstream target gene of bta-miRNA29d-3p was predicted to be GPAM. The results provide a theoretical basis for the regulatory mechanism of the bta-miRNA29d-3p gene in the lipid synthesis of BMECs.
Author Contributions
Conceptualization, D.Y.; funding acquisition, D.Y.; supervision, Y.M.; formal analysis, M.Z. and L.C.; validation, S.Z.; writing—original draft, X.Z.; writing—review and editing, J.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was jointly supported by the Innovative Research and Experimental Projects for Young Researchers of Tianjin Academy of Agricultural Sciences (No. 2021010 to D.Y.); the National Natural Science Foundation of China (No.31702095 to D.Y.) and Natural Science Foundation of Tianjin (No.18JCYBJC30200 to D.Y.); the Breeding innovation research project (2022ZYCX010 to Y.M.); the Tianjin Science and Technology project (22ZXZYSN00020 to Y.M.); and the Tianjin “131” Innovative Talents Team (No.20180338 to Y.M.).
Institutional Review Board Statement
All animal collection and protocols were authorized by the Animal Care and Use Committee of the Tianjin Institute of Animal Husbandry and Veterinary Medicine (2022015). All procedures were performed with minimal pain.
Data Availability Statement
The data are available online at https://doi.org/10.6084/m9.figshare.21393204.v1, accessed on 25 October 2022.
Acknowledgments
We thank Yi Ma for supporting the experiments, Dawei Yao for technical guidance, Zhi Chen for providing bovine mammary epithelial cells, and Jun Luo for providing the plasmids.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Curadi, M.C.; Leotta, R.; Contarini, G.; Orlandi, M. Milk fatty acids from different horse breeds compared with cow, goat and human milk. Maced. J. Anim. Sci. 2012, 2, 79–82. [Google Scholar] [CrossRef]
- Hanuš, O.; Samková, E.; Křížová, L.; Hasoňová, L.; Kala, R. Role of Fatty Acids in Milk Fat and the Influence of Selected Factors on Their Variability—A Review. Molecules 2018, 23, 1636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennighausen, L.; Robinson, G.W. Information networks in the mammary gland. Nat. Rev. Mol. Cell Biol. 2005, 6, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Sassi, Y.; Avramopoulos, P.; Ramanujam, D.P.; Grüter, L.; Werfel, S.; Giosele, S.; Brunner, A.-D.; Esfandyari, D.; Papadopoulou, A.-S.; De Strooper, B.; et al. Cardiac myocyte miR-29 promotes pathological remodeling of the heart by activating Wnt signaling. Nat. Commun. 2017, 8, 1614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massart, J.; Sjögren, R.J.O.; Lundell, L.S.; Mudry, J.M.; Franck, N.; O’Gorman, D.J.; Egan, B.; Zierath, J.R.; Krook, A. Altered miR-29 Expression in Type 2 Diabetes Influences Glucose and Lipid Metabolism in Skeletal Muscle. Diabetes 2017, 66, 1807–1818. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Zhang, Q.; Zhang, M.; Yang, J.C. Research Progress on Regulation Effect of microRNA on Mammary Gland Devel-opment and Lactation. China Anim. Husb. Vet. Med. 2015, 42, 663–667. [Google Scholar] [CrossRef]
- Wang, P.; Liu, S.; Zhu, C.; Duan, Q.; Jiang, Y.; Gao, K.; Bu, Q.; Cao, B.; An, X. MiR-29 regulates the function of goat granulosa cell by targeting PTX3 via the PI3K/AKT/mTOR and Erk1/2 signaling pathways. J. Steroid Biochem. Mol. Biol. 2020, 202, 105722. [Google Scholar] [CrossRef]
- He, A.; Zhu, L.; Gupta, N.; Chang, Y.; Fang, F. Overexpression of Micro Ribonucleic Acid 29. highly Up-Regulated in Diabetic Rats, Leads to Insulin Resistance in 3T3-L1 Adipocytes. Mol. Endocrinol. 2007, 21, 2785–2794. [Google Scholar] [CrossRef]
- Kurtz, C.L.; Fannin, E.E.; Toth, C.L.; Pearson, D.S.; Vickers, K.C.; Sethupathy, P. Inhibition of miR-29 has a significant lipid-lowering benefit through suppression of lipogenic programs in liver. Sci. Rep. 2015, 5, 12911. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Cao, X.; Lu, Q.; Zhou, J.; Wang, Y.; Wu, Y.; Mao, Y.; Xu, H.; Yang, Z. circ01592 regulates unsaturated fatty acid metabolism through adsorbing miR-218 in bovine mammary epithelial cells. Food Funct. 2021, 12, 12047–12058. [Google Scholar] [CrossRef]
- Bionaz, M.; Loor, J.J. Identification of reference genes for quantitative real-time PCR in the bovine mammary gland during the lactation cycle. Physiol. Genom. 2007, 29, 312–319. [Google Scholar] [CrossRef]
- Loor, J.J.; Dann, H.M.; Everts, R.E.; Oliveira, R.; Green, C.A.; Guretzky, N.A.J.; Rodriguez-Zas, S.L.; Lewin, H.A.; Drackley, J.K. Temporal gene expression profiling of liver from periparturient dairy cows reveals complex adaptive mechanisms in hepatic function. Physiol. Genom. 2005, 23, 217–226. [Google Scholar] [CrossRef] [Green Version]
- Zhou, F.; Teng, X.; Wang, P.; Zhang, Y.; Miao, Y. Isolation, identification, expression and subcellular localization of PPARG gene in buffalo mammary gland. Gene 2020, 759, 144981. [Google Scholar] [CrossRef]
- Bionaz, M.; Loor, J.J. Gene networks driving bovine milk fat synthesis during the lactation cycle. BMC Genom. 2008, 9, 366. [Google Scholar] [CrossRef] [Green Version]
- Harvatine, K.J.; Boisclair, Y.R.; Bauman, D.E. Liver x receptors stimulate lipogenesis in bovine mammary epithelial cell culture but do not appear to be involved in diet-induced milk fat depression in cows. Physiol. Rep. 2014, 2, e00266. [Google Scholar] [CrossRef]
- Christofides, A.; Konstantinidou, E.; Jani, C.; Boussiotis, V.A. The role of peroxisome proliferator-activated receptors (PPAR) in immune responses. Metabolism 2021, 114, 154338. [Google Scholar] [CrossRef]
- Horita, M.; Farquharson, C.; Stephen, L.A. The role of miR-29 family in disease. J. Cell. Biochem. 2021, 122, 696–715. [Google Scholar] [CrossRef]
- Kurtz, C.L.; Peck, B.C.; Fannin, E.E.; Beysen, C.; Miao, J.; Landstreet, S.R.; Ding, S.; Turaga, V.; Lund, P.K.; Turner, S.; et al. MicroRNA-29 Fine-tunes the Expression of Key FOXA2-Activated Lipid Metabolism Genes and Is Dysregulated in Animal Models of Insulin Resistance and Diabetes. Diabetes 2014, 63, 3141–3148. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Xu, P.; Wu, M.; Chen, H.; Xu, J.; Wu, M.; Li, M.; Qian, F. Bioinformatics analysis of hepatitis C virus genotype 2a-induced human hepatocellular carcinoma in Huh7 cells. OncoTargets Ther. 2016, 9, 191–202. [Google Scholar] [CrossRef] [Green Version]
- Jeon, T.-I.; Osborne, T.F. SREBPs: Metabolic integrators in physiology and metabolism. Trends Endocrinol. Metab. 2012, 23, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Ru, P.; Hu, P.; Geng, F.; Mo, X.; Cheng, C.; Yoo, J.Y.; Cheng, X.; Wu, X.; Guo, J.Y.; Nakano, I.; et al. Feedback Loop Regulation of SCAP/SREBP-1 by miR-29 Modulates EGFR Signaling-Driven Glioblastoma Growth. Cell Rep. 2016, 16, 1527–1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bian, Y.; Lei, Y.; Wang, C.; Wang, J.; Wang, L.; Liu, L.; Liu, L.; Gao, X.; Li, Q. Epigenetic Regulation of miR-29s Affects the Lactation Activity of Dairy Cow Mammary Epithelial Cells. J. Cell. Physiol. 2015, 230, 2152–2163. [Google Scholar] [CrossRef] [PubMed]
- Semple, R.K.; Chatterjee, V.K.K.; O’Rahilly, S. PPAR and human metabolic disease. J. Clin. Investig. 2006, 116, 581–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kidani, Y.; Bensinger, S.J. Liver X receptor and peroxisome proliferator-activated receptor as integrators of lipid homeostasis and immunity. Immunol. Rev. 2012, 249, 72–83. [Google Scholar] [CrossRef] [Green Version]
- Kadegowda, A.K.; Bionaz, M.; Piperova, L.S.; Erdman, R.; Loor, J. Peroxisome proliferator-activated receptor-γ activation and long-chain fatty acids alter lipogenic gene networks in bovine mammary epithelial cells to various extents. J. Dairy Sci. 2009, 92, 4276–4289. [Google Scholar] [CrossRef] [Green Version]
- Moyes, K.M.; Drackley, J.K.; Morin, D.; Bionaz, M.; Rodriguez-Zas, S.L.; Everts, R.E.; Lewin, H.A.; Loor, J.J. Gene network and pathway analysis of bovine mammary tissue challenged with Streptococcus uberis reveals induction of cell proliferation and inhibition of PPARγ signaling as potential mechanism for the negative relationships between immune response and lipid metabolism. BMC Genom. 2009, 10, 542. [Google Scholar] [CrossRef] [Green Version]
- Thering, B.J.; Graugnard, D.E.; Piantoni, P.; Loor, J. Adipose tissue lipogenic gene networks due to lipid feeding and milk fat depression in lactating cows. J. Dairy Sci. 2009, 92, 4290–4300. [Google Scholar] [CrossRef] [Green Version]
- Swanton, E.M.; Saggerson, E.D. Glycerolipid metabolizing enzymes in rat ventricle and in cardiac myocytes. Biochim. Biophys. Acta (BBA) Lipids Lipid Metab. 1997, 1346, 93–102. [Google Scholar] [CrossRef]
- Buhman, K.K.; Smith, S.J.; Stone, S.J.; Repa, J.J.; Wong, J.S.; Knapp, F.; Burri, B.J.; Hamilton, R.L.; Abumrad, N.A.; Farese, R.V., Jr. DGAT1 Is Not Essential for Intestinal Triacylglycerol Absorption or Chylomicron Synthesis. J. Biol. Chem. 2002, 277, 25474–25479. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Zhao, Z.; Yu, X.; Li, J.; Lu, C.; Yang, R. Bovine lipid metabolism related gene GPAM: Molecular characterization, function identification, and association analysis with fat deposition traits. Gene 2017, 609, 9–18. [Google Scholar] [CrossRef]
- Yu, H.; Zhao, Y.; Iqbal, A.; Xia, L.; Bai, Z.; Sun, H.; Fang, X.; Yang, R.; Zhao, Z. Effects of polymorphism of the GPAM gene on milk quality traits and its relation to triglyceride metabolism in bovine mammary epithelial cells of dairy cattle. Arch. Anim. Breed. 2021, 64, 35–44. [Google Scholar] [CrossRef]
- Yu, H.; Iqbal, A.; Fang, X.; Jiang, P.; Zhao, Z. Transcriptome analysis of CRISPR/Cas9-mediated GPAM−/− in bovine mammary epithelial cell-line unravelled the effects of GPAM gene on lipid metabolism. Gene 2022, 834, 146574. [Google Scholar] [CrossRef]
Figure 1.
Detection of overexpression and interference efficiency of bta-miRNA29d-3p. Results are the mean ± SEM of three separate experiments. ** p < 0.01 versus control.
Figure 1.
Detection of overexpression and interference efficiency of bta-miRNA29d-3p. Results are the mean ± SEM of three separate experiments. ** p < 0.01 versus control.

Figure 2.
Effect of bta-miRNA29d-3p overexpression on genes related to lipid metabolism in BMECs. Panel (A): Effect of bta-miRNA29d-3p overexpression on the expression of transcription factors SREBF1, PPARG1, and LXRA. Panel (B): Effect of bta-miRNA29d-3p overexpression on the expression of fatty acid de novo synthesis and desaturation-related genes (ACACA, FASN, and SCD1). Panel (C): Effect of bta-miRNA29d-3p overexpression on TAG synthesis and expression of lipid droplet formation-related genes (DGAT1 and GPAM). Panel (D): Effect of bta-miRNA29d-3p overexpression on intracellular TAG content. Results are the mean ± SEM of three separate experiments. ** p < 0.01 versus control, * p < 0.05 versus control (Con miR).
Figure 2.
Effect of bta-miRNA29d-3p overexpression on genes related to lipid metabolism in BMECs. Panel (A): Effect of bta-miRNA29d-3p overexpression on the expression of transcription factors SREBF1, PPARG1, and LXRA. Panel (B): Effect of bta-miRNA29d-3p overexpression on the expression of fatty acid de novo synthesis and desaturation-related genes (ACACA, FASN, and SCD1). Panel (C): Effect of bta-miRNA29d-3p overexpression on TAG synthesis and expression of lipid droplet formation-related genes (DGAT1 and GPAM). Panel (D): Effect of bta-miRNA29d-3p overexpression on intracellular TAG content. Results are the mean ± SEM of three separate experiments. ** p < 0.01 versus control, * p < 0.05 versus control (Con miR).

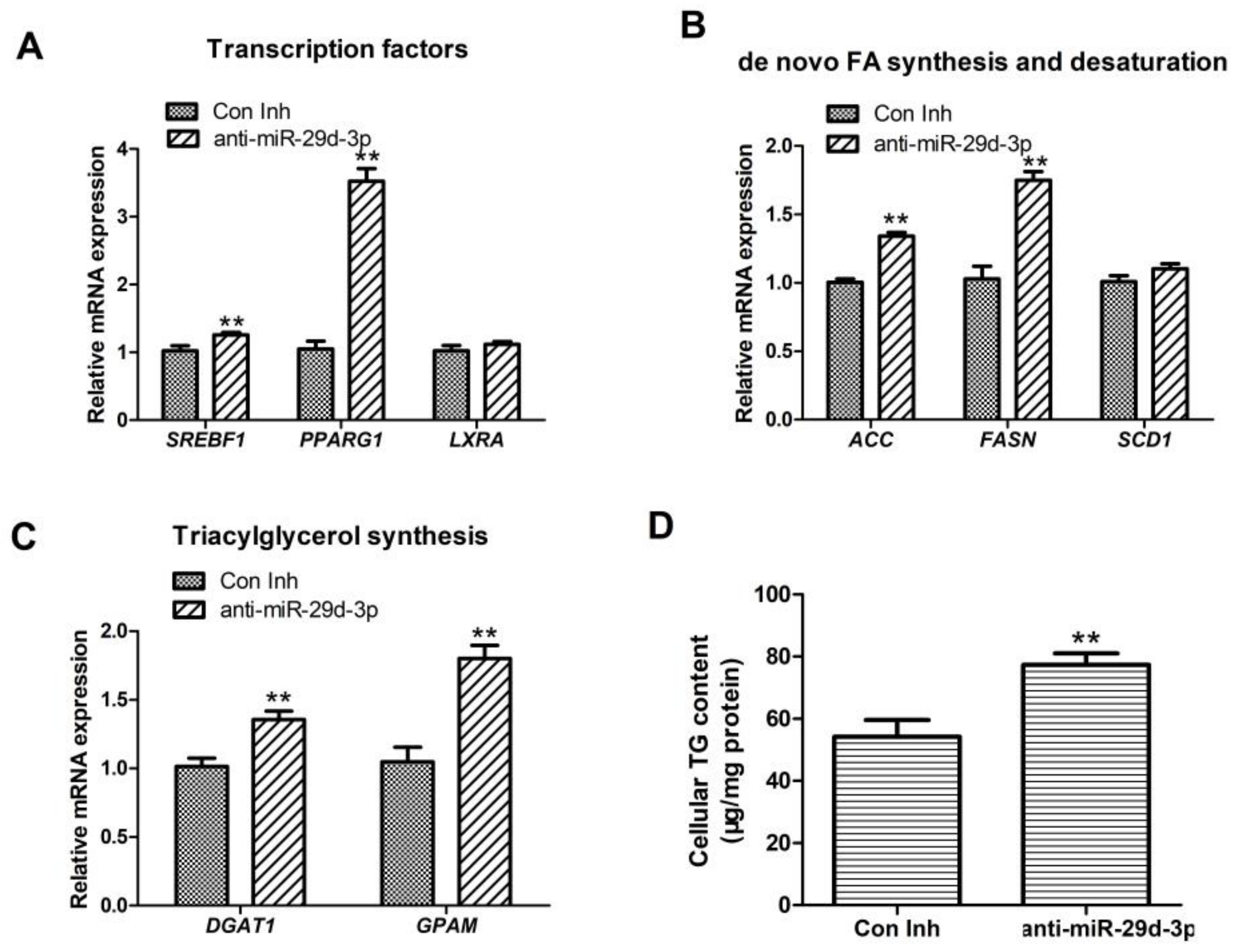
Figure 3.
Effect of interference with bta-miRNA29d-3p with respect to lipid metabolism-related genes in BMECs. Panel (A): Effect of interfering with bta-miRNA29d-3p on the expression of transcription factors SREBF1, PPARG1, and LXRA. Panel (B): Effect of interfering with bta-miRNA29d-3p with reference to the expression of fatty acid ab initio synthesis and desaturation-related genes (ACACA, FASN, and SCD1). Panel (C): Effect of interfering with bta-miRNA29d-3p on the analysis of TAG synthesis and lipid droplet formation-related genes (DGAT1 and GPAM) expression. Panel (D): Effect of interfering with bta-miRNA29d-3p on intracellular TAG content. Results are the mean ± SEM of three separate experiments. ** p < 0.01 versus control (Con Inh).
Figure 3.
Effect of interference with bta-miRNA29d-3p with respect to lipid metabolism-related genes in BMECs. Panel (A): Effect of interfering with bta-miRNA29d-3p on the expression of transcription factors SREBF1, PPARG1, and LXRA. Panel (B): Effect of interfering with bta-miRNA29d-3p with reference to the expression of fatty acid ab initio synthesis and desaturation-related genes (ACACA, FASN, and SCD1). Panel (C): Effect of interfering with bta-miRNA29d-3p on the analysis of TAG synthesis and lipid droplet formation-related genes (DGAT1 and GPAM) expression. Panel (D): Effect of interfering with bta-miRNA29d-3p on intracellular TAG content. Results are the mean ± SEM of three separate experiments. ** p < 0.01 versus control (Con Inh).

Figure 4.
Bta-miRNA29d-3p target gene prediction and dual luciferase assay. Panel (A): bta-miRNA29d-3p target gene prediction and conservativeness analysis. Panel (B): Effect of bta-miRNA29d-3p overexpression on dual luciferase activity. Panel (C): Effect of interfering with bta-miRNA29d-3p on dual-luciferase activity. Results are the mean ± SEM of three separate experiments. ** p < 0.01 versus control (Con miR, Con Inh).
Figure 4.
Bta-miRNA29d-3p target gene prediction and dual luciferase assay. Panel (A): bta-miRNA29d-3p target gene prediction and conservativeness analysis. Panel (B): Effect of bta-miRNA29d-3p overexpression on dual luciferase activity. Panel (C): Effect of interfering with bta-miRNA29d-3p on dual-luciferase activity. Results are the mean ± SEM of three separate experiments. ** p < 0.01 versus control (Con miR, Con Inh).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Characteristics of primers used of the RT-qPCR reaction.
| Gene1 | Accession No. | Primer Sequences (5′-3′) | Source |
|---|---|---|---|
| UXT | XM_005700842.1 | F: TGTGGCCCTTGGATATGGTT R: GGTTGTCGCTGAGCTCTGTG | [11] |
| MRPL39 | XM_005674737.1 | F: AGGTTCTCTTTTGTTGGCATCC R: TTGGTCAGAGCCCCAGAAGT | [11] |
| GPAM | AY515690 | F: GCAGGTTTATCCAGTATGGCATT R: GGACTGATATCTTCCTGATCATCTTG | [12] |
| PPARG1 | NM_001290893.1 | F: CGTTTCCTTAAACAAGTG R: TCCCTCAAAATAATAGTGC | [13] |
| DGAT1 | NM_174693 | F: CCACTGGGACCTGAGGTGTC R: GCATCACCACACACCAATTCA | [14] |
| LXRA | NM_001014861.1 | F: CATGCCTACGTCTCCATCCA R: TCACCAGTTTCATCAGCATCCT | [15] |
| SREBF1 | TC263657 | F: CCAGCTGACAGCTCCATTGA R: TGCGCGCCACAAGGA | [12] |
| FASN | CR552737 | F: ACCTCGTGAAGGCTGTGACTCA R: TGAGTCGAGGCCAAGGTCTGAA | [14] |
| ACACA | AJ132890 | F: CATCTTGTCCGAAACGTCGAT R: CCCTTCGAACATACACCTCCA | [14] |
| SCD1 | GU947654 | F: CCATCGCCTGTGGAGTCAC R: GTCGGATAAATCTAGCGTAGCA | [16] |
Table 2.
Primers used for site-directed mutation of bovine GPAM 3′UTR constructs.
| Primer Name | Primer Sequences (5′-3′) |
|---|---|
| GPAM-F GPAM-R | TTGCCCTTTCAGTTGGTTT TTGCCCTTTCAGTTGGTTT |
| MiR-29d-3p-binding site | TGGTGCTTTG |
| Mutation site | CACGTACCCT |
| Overlap-F | GTAATTAACCACGTACCCTAA |
| Overlap-R | GGTTTTAGGGTACGTGGTTAATTAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhao, X.; Li, J.; Zhao, S.; Chen, L.; Zhang, M.; Ma, Y.; Yao, D. Regulation of bta-miRNA29d-3p on Lipid Accumulation via GPAM in Bovine Mammary Epithelial Cells. Agriculture 2023, 13, 501. https://doi.org/10.3390/agriculture13020501
AMA Style
Zhao X, Li J, Zhao S, Chen L, Zhang M, Ma Y, Yao D. Regulation of bta-miRNA29d-3p on Lipid Accumulation via GPAM in Bovine Mammary Epithelial Cells. Agriculture. 2023; 13(2):501. https://doi.org/10.3390/agriculture13020501
Chicago/Turabian StyleZhao, Xin, Jun Li, Shuying Zhao, Lili Chen, Man Zhang, Yi Ma, and Dawei Yao. 2023. "Regulation of bta-miRNA29d-3p on Lipid Accumulation via GPAM in Bovine Mammary Epithelial Cells" Agriculture 13, no. 2: 501. https://doi.org/10.3390/agriculture13020501
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.