
Influence of Lighting and Laser Irradiation on the Germination of Caper Seeds

1
Plant Production Department, Universitat Politècnica de València, 46022 Valencia, Spain
2
Horticulture and Floriculture, Agriculture Faculty, National University of Cuyo, Mendoza M5528AHB, Argentina
*
Author to whom correspondence should be addressed.
Agriculture 2022, 12(10), 1612; https://doi.org/10.3390/agriculture12101612
Submission received: 8 September 2022
/
Revised: 29 September 2022
/
Accepted: 30 September 2022
/
Published: 4 October 2022
(This article belongs to the Special Issue Impact of Light on Horticultural Crops)
Abstract
:Caper seeds present difficulties in their germination, which has been studied by several research teams. It is known that light can release dormancy in some seeds, but its effect on caper seed germination has not yet been deeply studied. The main aim of this study was to analyze the response of caper seeds germination to light exposure. The study analyzed the germination response of seeds to lighting with different wavelengths (white, red, blue, red + blue and darkness) and to the He-Ne laser light, using both dry seeds and seeds that had been previously soaked in water. Overall, it could be stated that caper seeds are insensitive to light during the germination process. Thus, germination could be carried out in lightness or darkness, so germination in nurseries could be carried out in the darkness, leading to substantial energy savings. Caper seed irradiation with a He-Ne laser during short exposure times improved the germination percentage for the seeds previously soaked in water, germinating all viable seeds. However, applying a solution of gibberellic acid was always required in all the cases studied.
1. Introduction
The caper (Capparis spinosa L.) is a deciduous creeping shrub native to Asia, which spreads throughout the Mediterranean basin, where it grows in dry lands. It is currently cultivated in Spain, Italy, France, Greece, and North Africa, as well as in South America [1,2]. It is mainly cultivated for the floral buds, called capers; however, their fruits and, to a lesser extent, their vegetative shoots are also consumed pickled or salted. As Shahrajabian et al. [3] have stated, different parts of the plant are rich sources of antioxidants and bioactive compounds beneficial to health. Furthermore, the flowers have a high ornamental value; thus, caper plants are included in gardening, particularly in xeriscape [4,5].
In a recent review article, Sottile et al. [1] stated that, as a crop, caper should not be considered a difficult crop to propagate. Pascual et al. [6] reported acceptable percentages of success in the rooting of cuttings, and Foschi et al. [7,8,9] obtained high germination percentages of dry seeds using gibberellic acid and without the need for any treatment in fresh seeds. These germination tests were carried out in a growth chamber under a photoperiod of 12 h.
Light is one of the main environmental signals for plants [10], being an important factor in breaking seed dormancy [11]. To respond to environmental signals, plants have developed several families of photoreceptors, which are photosensitive pigments capable of being activated by photons of specific wavelengths and, in turn, activatingsignal translation pathways, providing the ability to respond to light stimuli [12]. These photoreceptors include the following: (i) Phytochromes, which are red (600–700 nm) and far red (700–800 nm) light photoreceptors; (ii) Cryptochromes, which are photoreceptors for blue (400–500 nm) and ultraviolet A (320–400 nm) light; (iii) Phototropins, another group of blue and UV-A light photoreceptors [10,13].
Butler et al. [14] obtained a photo-reversible pigment from etiolated shoots of maize and named it phytochrome. Later, in studies carried out on Arabidopsis thaliana (L.) Heynh., Sharrock and Quail [15] identified sequences that showed that small families of genes encode phytochromes. Specifically, these authors hypothesized that the minimum number of phytochrome genes present in higher plants could be determined in studies in A. thaliana. They stated that phytochromes in this plant were encoded by a small gene family consisting of at least three genes and probably four or five. Subsequent studies have shown that A. thaliana contains five different phytochromes (phyA, phyB, phyC, phyD and phyE) [10] that are encoded by five genes (PHYA, PHYB, PHYC, PHYD and PHYE). Cryptochromes were first identified in A. thaliana [16]. Different organisms have different numbers of cryptochromes; plants have at least two types of cryptochromes, and this number can range up to six, as in soybean (Glycine max L.) [16].
According to sensitivity to white light, seeds have been classified into three categories [17]: (i) positive photoblastic (the seeds that germinate only under white light); (ii) negative photoblastic (the seeds that germinate only in the dark); (iii) light insensitive (the seeds that germinate both under white light and in darkness). A Photoblastic Index (PI; Equation (1)) has been utilized to evaluate the photoblastic responses [18]:
PI = (GD − GL)/(GD + GL)
GD is the germination (%) in darkness, and GL is the germination (%) under light. This index ranges from 1 (negative photoblastism) to −1 (positive photoblastism); PI = 0 indicates that germination is not dependent on light.
Another index expressing a light requirement is the Relative Light Germination (RLG; Equation (2) [19]):
RLG = (GL)/(GD + GL)
GD is the germination (%) in darkness, and GL is the germination (%) under light. The RLG values vary from 0 (only seeds exposed to darkness germinate) to 1 (only seeds exposed to light germinate), with values close to 0.5 in seeds germinating both in light and darkness.
Takaki [20] proposed to replace the term photoblastism with forms of phytochrome that control germination, proposing a classification based on three mechanisms, depending on the level of fluence (total energy received by a seed in a period of time, J m−2 [21]) to saturate the responses [20,22]:
- Low Fluency Responses (LFRs), which represent the classic reversible red/far red responses, in which Pfr (the active form of phytochrome, which absorbs light of 735 nm) production promotes plant responses and removal of Pfr reverses the response. These seeds have phyB controlling the germination process through LFR. The saturation of the response frequently occurs at low levels of Pfr/Ptotal (10−2−0.87 Pfr/Ptotal) and intermediate fluences (1–1000 µmol m−2 s−1).
- High Irradiance Responses (HIRs), which represent responses produced by prolonged high irradiation, which do not show reciprocity or reversibility. These seeds have phyA controlling germination through HIR, and the maximum reaction generally occurs at wavelengths that maintain low Pfr levels for long periods of time, such as occurs in far-red-rich environments.
- Very low fluence responses (VLFRs), which represent the saturation of responses by very low fluences, with reciprocity but no reversibility because the photo-equilibrium maintained by far-red light (or even safe dim green light used in experiments) produces enough Pfr to saturate these responses, which occur at low levels of Pfr (10−6−10−3 Pfr/Ptotal). These seeds have phyA controlling the germination process through VLFR.
LED (Light Emitting Diode) lights are an alternative to incandescent lamps and cold white fluorescent tubes for growing plants. They have several advantages due to their small size, long lifespan, low emission temperature, high efficiency in energy conversion, and the possibility of selecting specific wavelengths [23,24]. In the last two decades, several studies have shown that LEDs of different wavelengths can modify the germination, growth, and development of seedlings in many species [11,25,26,27,28,29,30]. In this type of light, which is mainly monochromatic, it is essential to know the optimal light spectrum and the intensity required by the different species at each phenological stage to optimize yield and quality [28].
The irradiation of seeds with laser light (Light Amplification by Stimulated Emission of Radiation) can also be an alternative to improve the germination or growth and development of seedlings of various species based on the bio-stimulant effect of laser light [31,32,33]. Laser irradiation effects depend on many laser parameters, such as wavelength, irradiation duration, power, dose, and method (constant/pulse) [31]. However, seed properties are also important, particularly their genetic traits and physiological properties (health status, seed quality), and even their orientation during irradiation [31,32].
Among the different lasers used in agriculture, the helium-neon laser (He-Ne) is the most used [31], considering that its wavelength of 632.4 nm corresponds to the red light which is responsible for phytochrome activation [32]. The improvement and acceleration of germination have been related to an induction of the enzymatic activities, a change of thermodynamic parameters, and an acceleration of physiological and biochemical metabolism of seeds, increasing, in some cases, the levels of gemination-promoting growth hormones. such as gibberellic acid (GA3). and decreasing inhibitors, such as abscisic acid [34,35,36,37,38,39].
Light and gibberellins can release dormancy in some seeds [40], specifically in those that present coat dormancy, promoting their germination [41]. Caper seeds have a non-deep physiological dormancy, specifically a coat-imposed dormancy due to a mechanical characteristic, which can be released by adding GA3 to the germination substrate [7]. As far as is known, the effect of light on caper seed germination has not yet been deeply studied. The main aim of this study was to analyze the response of caper seeds germination to light exposure. Particularly, the study analyzes the germination response of seeds to lighting with different wavelengths (white, red, blue, red + blue and darkness) and to the He-Ne laser light.
2. Materials and Methods
2.1. Plant Material
Caper (Capparis spinosa L.) seeds were extracted from ripe fruits produced by adult plants grown in Llíria, Valencia, Spain (39°38′54.2′′ N, 0°37′3.5′′ W). The fruit collection was carried out over the first fortnight of September 2019, 2020, and 2021, each constituting a different lot.
After the extraction, mature seeds were selected using the flotation method [42] with tap water. The seeds were disinfected by soaking them in a 25% sodium hypochlorite solution and then rinsing with tap water. They were dried for 15 days at room temperature in the shade and kept in hermetically sealed glass containers at 7 ± 0.5 °C in a domestic refrigerator (Beko, Beko Electronics España, Barcelona, Spain). At the beginning of each germination test, the three seed lots were within the recommended storage period to not affect their viability [7,43].
2.2. Viability and Germination Tests
The seed lots’ viability was determined by the tetrazolium test, as Foschi et al. [7] reported (four replicates of fifty seeds each), according to the International Rules for Seed Testing [44].
Germination tests were carried out with the Between Paper method, placing 100 seeds per Petri dish of 9 cm diameter [45]. In all cases, four replicates were performed per treatment. Ultrapure water (Wasserlab G.R type II analytical grade water system; from now on referred to as water) or a solution of 500 mg L−1 of GA3 (Semefil L, Nufarm L.) were used to wet the substrate. The Petri dishes were then placed under controlled conditions in a growth chamber (model Zimbueze, Seville, Spain) at 30 ± 1/20 ± 1 °C, 85 ± 1% relative humidity for a photoperiod of 12 h (cold white fluorescent tubes Philips TL-D 36W/54), providing a photosynthetic photon flux density (PPFD) of 81.1 ± 1.7 μmol m−2 s−1, unless stated otherwise.
The germination test lasted 120 days, and germinated seeds were counted and removed periodically. Seeds were considered germinated when the radicle protruded from the testa and the micropylar endosperm, reaching a length of approximately 2 mm. Results of germination tests were fitted to the logistic function [46,47], defined as a particular case of Richards’ function ([48] Equation (3)):
G = A/1 + e(β − kt)
G is the cumulative germination (%), A represents the final germination percentage, t is the germination time (d), and β and k are function parameters used to determine the time required to reach 50% of G (Gt50 = β/k; d) and the mean relative cumulative germination rate (k/2; d−1).
2.3. Experiment 1
This experiment evaluated the effect of light and darkness on the germination of the two caper seed lots (corresponding to seeds produced in 2019 and 2020). The experiment started in March 2021. The seeds were placed in the germination chamber with a photoperiod of 12 h of white light or continuous darkness. Light exposure was provided by cool white fluorescent tubes, as previously stated, and seeds that germinated in the dark were placed in closed opaque boxes. The Photoblastic Index (PI) and the Relative Light Germination (RLG) were determined using Equations (1) and (2).
Eight combinations of the three sources of variation were tested: 2 seed lots, light-darkness and 2 wetting solutions.
2.4. Experiment 2
The second experiment evaluated the effect of lighting with different wavelengths on caper seed germination. It started in July 2021. In light of the results obtained in the first experiment, tests were performed under white, red, blue, and red + blue lights. Color lighting was achieved with LED lights (AMZLAB GmbH), consisting of 80 LEDs (52 red, wavelength range 600–700 nm, and 28 blue, wavelength range 400–500 nm) placed 25 cm above the Petri dishes. The maximum power, when using the full spectrum of lights, was 30 W. Four types of light were tested: white was provided by fluorescent tubes (Philips TL-D 36W/54) with a power of 36 W, a distribution of the color spectrum ranging from wavelengths of 300 to 800 nm and with photosynthetically active radiation of 81.1 μmol m−2 s−1; red (52 red LEDs; 102.5 μmol m−2 s−1), blue (28 blue LEDs; 80.1 μmol m−2 s−1) and red + blue (52 red LEDs + 28 blue LEDs; 125.4 μmol m−2 s−1). Darkness was applied as in Experiment 1, placing the seeds in closed opaque boxes.
Table 1 shows the percentages and wavelengths for the color spectra of the lamps, measured with a Thorlabs spectrometer, model CCS200/M. The rates of each type of wavelength in each light were calculated using the ImageJ program [49], measuring the areas between the desired ranges of the wavelengths of the color spectrum.
This experiment analyzed 20 combinations of the three sources of variation: 2 seed lots, 5 types of lighting and 2 wetting solutions.
2.5. Experiment 3
The third experiment evaluated the influence of He-Ne laser irradiation, with different exposure times, on germination of caper seeds. Seeds from the lot harvested in 2021 were used in this experiment, which started in December 2021.
The seeds were irradiated with a He-Ne laser (JDS Uniphase model 1145), with an output emission power of 22.5 mW and wavelength of 632.8 nm, belonging to the red band of the spectrum. It had a circular beam of 0.7 mm in diameter.
To define the exposure times (and therefore energies) to be analyzed in this experiment, a preliminary study was carried out to analyze the effect on seed viability of the following exposure times: 0, 1, 5, 15, 30, 60, 120 and 180 s, for which 40 seeds were irradiated during each of the times. These assayed exposure times expanded the range Juan [50] tested with the same laser. Neither of the analyzed exposure times decreased (p ≤ 0.05) the seed viability (data not shown), thus, maximum and minimum thresholds were included in the experiment, as well as three intermediate levels: 0, 1, 15, 60 and 180 s, corresponding to 0, 22.5, 337.5, 1350, 4050 mJ applied energy levels, respectively. Intact seeds were irradiated one by one. After irradiation, the germination test was performed as previously indicated.
A total of 10 combinations of two sources of variation were tested: 5 exposure times and 2 wetting solutions.
2.6. Experiment 4
The fourth experiment evaluated the effect on germination of soaking the seeds in water before irradiation with the He-Ne laser with two timings of exposure, 1 and 15 s, which led to the best results in Experiment 3. As in the previous experiment, seeds of the 2021 lot were used and the experiment started in February 2022.
Caper seeds were soaked in water for four days before laser irradiation by a He-Ne laser for 0, 1 and 15 s. The germination test was performed as previously indicated. The germination substrate was only wetted with the GA3 solution, as this experiment was not analyzing the effect of the wetting solution, since it haf already been stated that the GA3 solution was required to get an acceptable germination percentage that allowed adjusting the logistic model.
This experiment tested six combinations of two sources of variation: 3 exposure times and soaked/dry seeds.
2.7. Data Analysis
All the tests verified that the tolerance required by the ISTA standards [45] was met, either between the replicates or between the germination tests. The statistical analysis program Statgraphics [51] was used to perform multi-way analyses of variance (ANOVA; p ≤ 0.05) and verify the normality of the data. Mean separations were performed where appropriate, using Fisher’s smallest significance difference (LSD test) at p ≤ 0.05.
3. Results and Discussion
3.1. Experiment 1
The viability of the two seed lots was very high (82.5 + 2.5% and 90.0 + 3.1% in the seeds produced in 2019 and 2020, respectively), and, as a consequence, among other factors of careful harvesting, cleaning, and drying of the seeds, and according to that reported by Foschi et al. [9], there were no differences (p < 0.01) between both values.
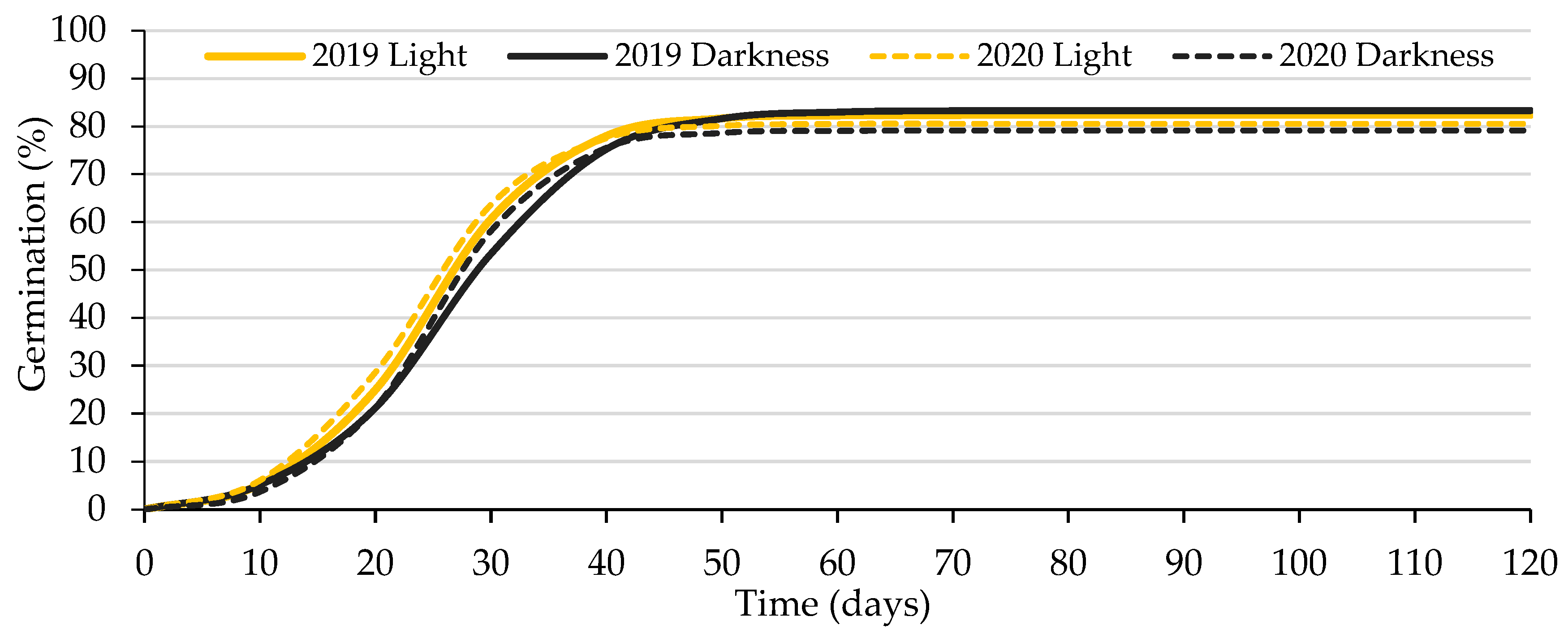
Germination percentages obtained when water was used to wet the germination substrate were very low (specifically 9.5% in light and 6.8% in darkness, on average, for the two lots). These values were significantly lower (p < 0.05) than those obtained with the addition of GA3 (81.5% in light and 81.2% in darkness, on average for the two lots). The low germination percentages obtained with water did not allow adjusting the logistic model; thus, the statistical analysis (Table 2) was only carried out for the seeds wetted with the GA3 solution. The high germination values obtained with the GA3 addition in the two lots are worth noting, accounting for 92% and 97% of the viable seeds. The 16 curves were adjusted to the germination logistic model with a determination coefficient higher than 99.6%, which allowed using the variable A (instead of G), as well as the other variables derived from the logistic function, as occurred in previous studies of caper seed germination carried out by Pascual et al. [52] and Foschi et al. [8] It applied to all the experiments reported in this manuscript. Figure 1 shows the logistic model adjusted to the average curves of accumulated germination of caper seeds from this experiment.
Neither the seed lot and the seeds’ exposure to light–darkness, nor their interaction, influenced (p < 0.05) any of the determined germination parameters. These results coincided with those obtained by Germanà and Chiancone [53], in the sense that no significant difference was obtained when incubating mechanically scarified caper seeds in lightness and darkness. The non-statistical significance between the mean values of A obtained in light (81.5%) and darkness (81.2%) indicated that the caper seeds germinated equally in light and darkness, which was corroborated by the values of the photoblastism (PI = −0.002) and Relative Light Germination (RLG = 0.5) indices. PI and RLG were calculated only for seeds germinated in substrate wetted with GA3 solution and not with water to exclude calculations of both indices based on a small number of seeds, as Milberg et al. [19] did.
As Takaki [20] reported, light insensitive seeds present phyA, corresponding to the so-called very low fluence responses (VLFR [21]). They saturate the responses by very low fluences because the photoequilibrium maintained by far-red light or extremely low light fluences in most regions of the visible spectrum produces enough Pfr to saturate this response [20,22].
3.2. Experiment 2
With the low germination percentages obtained with the seeds in water (on average, 3%, data not shown), it was not possible to fit the logistic model, so these data were not included in the subsequent statistical analysis. The germination percentage obtained with the GA3 addition did significantly (p ≤ 0.01) fit the logistic function, obtaining determination coefficients greater than 98.6%.
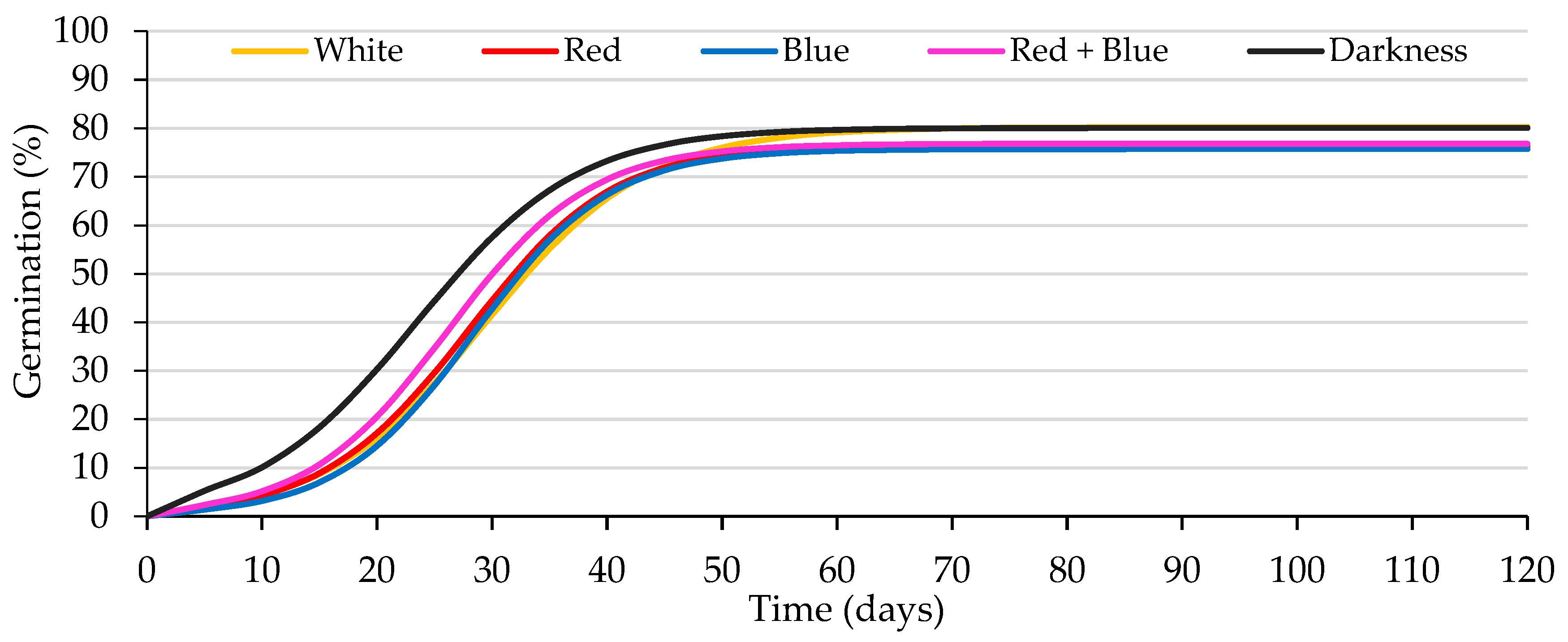
The seed lot did not affect any germination parameters (p ≤ 0.05; Table 3); thus, in Figure 2, to facilitate its interpretation, average values for both lots are presented. As seen in Table 3 and Figure 2, the light wavelength did not influence (p ≤ 0.05) the germination of the seeds, not differing from permanent darkness. In all cases, these germination percentages were high, comparable to those obtained in the first experiment; the difference between the germination values obtained in this and the previous experiment (both for white light and darkness) did not exceed the tolerance level established by ISTA Rules [45]. Neither the Gt50 nor the k/2 was affected by the different wavelengths. The interaction between the seed lot and the exposure to different wavelengths did not affect (p ≤ 0.05) any of the germination parameters.
No significant differences were obtained between the A values obtained in white light (80.3%) and in darkness (80.1%), indicating that the caper seeds germinated equally in white light and darkness, which was corroborated by the values of the Photoblastic Index (PI = −0.001) and Relative Light Germination (RLG = 0.5). These results confirmed those obtained in the previous experiment, that caper seeds are insensitive to white light; thus, nurseries could save the energy needed to illuminate the seeds during germination.
Caper seeds were also insensitive to blue light, even though all plants had cryptochromes [16], photoreceptors of blue light (400–500 nm; [13]). They were also insensitive to red light, even though phytochromes are photoreceptors of red light (600–700 nm).
Neither Marín [54], nor Moreno et al. [55], obtained significant differences (p ≤ 0.05) when applying white, red, and blue light in the Serrano variety of pepper (Capssicumm annuum L.) nor orchids Encyclia, respectively. Paniagua et al. [27] and Cho et al. [26] analyzed the effect of LED lights of different wavelengths on broccoli (Brassica oleracea Plenck var. italica), with none of them obtaining any statistical difference (p ≤ 0.05) in the final germination percentage. Aguado and Álvarez [25] obtained no differences in the germination and emergence of lettuce (Lactuca sativa L.), basil (Ocimum basilicum L.), and tomato (Solanum lycopersicum L.) seedlings exposed to LED lights with spectra differing on the proportions of red and blue lights.
However, Enache and Livadariu [29], with the use of red LED lights in Artemisia dracunculus L.. obtained a 10% greater germination percentage than with white light, which was, in turn, higher than those obtained under blue and green LEDs. However, the authors did not statistically compare these results.
3.3. Experiment 3
The viability of the 2021 lot was 85 + 4%, according to what was obtained for the 2019 and 2020 lots. The germination obtained in the seeds wetted with water was very low, on average 6.5% (6.2% in irradiated and 8% in unirradiated seeds). The low germination data did not adjust to the logistic model; thus, they were not included in the analysis of variance, as in Experiments 1 and 2.
Germination data for seeds wetted with the GA3 solution was fitted (p ≤ 0.01) to the logistic function, presenting coefficients of determination for the 20 curves greater than 98.7%. Figure 3 shows the cumulative germination curves fitted to the logistic model obtained for the average values of each irradiation duration, the final germination percentages ranging between 69% and 82%. The tested laser irradiation durations (0, 1, 15, 60 and 180 s) did not significantly affect (p ≤ 0.05) A or k/2, but an increase in Gt50 was observed in seeds irradiated for 180 s in relation to the control seeds (7 days delay; Table 4) and to those irradiated for 1 and 15 s (up to 10 days delay).
The results coincided with those that Juan, [50], obtained with the same type of laser used, in the sense that laser irradiation did not improve the germination of caper seeds. Similar results were obtained by Álvarez et al. [56] in tomato seeds irradiated with a He-Ne laser, which did not improve the germination percentages.
On the other hand, other studies on many species found that irradiation with laser light improved germination parameters, including wheat (Triticum aestivum L. [36]), lupine (Lupinus albus L.) and bean (Vicia faba L.) [38], radish (Raphanus sativus L. [57]), soybean (Glycine max L. [58]), safflower (Carthamus tinctorius L. [59]), sunflower (Helianthus annuus L. [37]) and Chinese woad (Isatis indigotica L. [34]). The laser light can affect the thermodynamic parameters of the seeds by increasing their internal energy, affecting the enzymatic activity (mainly of amylases, proteases and glucosidases), which may positively influence the germination [36]. Another He-Ne laser effect is the acceleration of seed metabolism through increased levels of germination-promoting hormones, such as GA3, and decreased inhibitors, such as abscisic acid, as stated by Soliman and Harith [39] in Acacia farnesiana L. and by Swathy et al. [33] in eggplant (Solanum melongena L.).
The success of bio-stimulation caused by monochromatic laser light depends on the wavelength, irradiation duration, power, dose and method (constant or pulse), but also on the seed physiological properties and even the seed position during the laser irradiation [31]. Krawiec et al. [32] related the greatest response of seeds irradiated with a laser beam to the fact that these seeds had been previously soaked in water; thus, it was decided to analyze the effect of soaking the seeds in water before their irradiation, as presented in Experiment 4.
3.4. Experiment 4
Germination data were fitted (p ≤ 0.01) to the logistic function, presenting coefficients of determination for the 24 curves greater than 98.1%. Figure 4 shows the germination curves adjusted to the logistic model in which germination was higher in seeds irradiated when they had been previously soaked in water compared to dry seeds and control. These differences (p ≤ 0.05) are shown in Table 5, where it can also be seen that germination was affected (p ≤ 0.01) by the irradiation duration. Soaking the seeds in water before irradiation represented 48% of the variation in the data, while the irradiation duration represented 26% of the variability. Figure 5 presents the significant interaction (p ≤ 0.01) between the irradiation duration and the seed soaking before the irradiation. Germination percentages were comparable to those obtained in Experiment 3 with the same seed lot; the difference between the germination values obtained in this and the previous experiment for non-irradiated seeds, did not exceed the tolerance level established by ISTA Rules [45]. It can be seen that soaking the seeds before laser irradiation significantly increased (p ≤ 0.05) the germination percentage, germinating all viable seeds. Neither Gt50 nor k/2 was affected by the analyzed (p ≤ 0.05) factors.
As already mentioned in Experiment 3, the laser light can affect the thermodynamic parameters of the seeds [36]. It can also increase seed metabolism through increased levels of germination-promoting hormones (GA3) and decreased inhibitors (abscisic acid), positively affecting germination [33]. In this experiment, irradiating the seeds once the germination metabolism had started, due to the prior seed soaking, improved the effectiveness of the laser irradiation in relation to that obtained in Experiment 3, resulting in significant differences (p ≤ 0.05). This was in accordance with that reported by Perveen et al. [37,59] for sunflower and safflower, respectively, who obtained good germination results irradiating seeds that had been previously soaked in water with a He-Ne laser.
Future research will focus on studying the photoreceptors (phytochromes, cryptochromes and phototropins) present in caper seeds, as well as on analyzing the enzymatic activities and the levels of germination promoting and inhibiting hormones in seed irradiated with laser light.
4. Conclusions
Caper seeds are insensitive to exposure to white, red, blue, and red + blue lights during the germination process, not showing differences between the germination response in lightness in relation to darkness. Thus, germination can be carried out in lightness or darkness, and, therefore, germination in nurseries could be carried out in darkness, leading to important energy savings. Caper seed irradiation with a He-Ne laser during short exposure times improved the germination percentage when the seeds had been previously soaked in water, germinating all viable seeds. However, applying a solution of gibberellic acid was always required in all the cases studied.
Author Contributions
Conceptualization, M.J. and B.P.; methodology, M.L.F.; software, M.L.F.; validation, M.L.F., M.J., B.P. and N.P.-S.; formal analysis, M.L.F., M.J., B.P. and N.P.-S.; investigation, M.L.F. and N.P.-S.; resources, M.J.; data curation, M.L.F., B.P. and N.P.-S.; writing—original draft preparation, M.L.F., B.P. and N.P.-S.; writing—review and editing, M.L.F., M.J., B.P. and N.P.-S.; visualization, M.L.F., M.J., B.P. and N.P.-S.; supervision, B.P. and N.P.-S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Not applicable.
Acknowledgments
Authors thank Vicente Ferrando, from the Applied Physics Department (UPV), for the technical support in the measurements with the spectrometer for the wavelength of the lamps used in this research.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sottile, F.; Caltagirone, C.; Peano, C.; Del Signore, M.B.; Barone, E. Can the caper (Capparis spinosa L.) still be considered a difficult-to-propagate crop? Horticulturae 2021, 7, 316. [Google Scholar] [CrossRef]
- Rhizopoulou, S.; Ioannidi, E.; Alexandredes, N.; Argiropoulos, A. A study on functional and structural traits of the nocturnal flowers of Capparis spinosa L. J. Arid Environ. 2006, 66, 635–647. [Google Scholar] [CrossRef]
- Shahrajabian, M.H.; Sun, W.; Cheng, Q. Plant of the millennium, caper (Capparis spinosa L.), chemical composition and medicinal uses. Bull. Natl. Res. Cent. 2021, 45, 131. [Google Scholar] [CrossRef]
- Rivera, D.; Inocencio, C.; Obón, C.; Alcaraz, F. Review of food and medicinal uses of Capparis L. subgenus Capparis (Capparidaceae). Econ. Bot. 2003, 57, 515–534. [Google Scholar] [CrossRef]
- Fici, S. A taxonomic revision of the Capparis spinosa group (Capparaceae) from the Mediterranean to Central Asia. Phytotaxa 2014, 174, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Pascual, B.; San Bautista, A.; Pascual-Seva, N.; García-Molina, R.; López Galarza, S.; Maroto, J.V. Estudio del enraizamiento de estaquillas de madera suave en alcaparra (Capparis spinosa L.). Actas Hortic. 2008, 50, 89–93. [Google Scholar]
- Foschi, M.L.; Juan, M.; Pascual, B.; Pascual-Seva, N. The imbibition, viability, and germination of caper seeds (Capparis spinosa L.) in the first year of storage. Plants 2022, 11, 202. [Google Scholar] [CrossRef]
- Foschi, M.L.; Juan, M.; Pascual, B.; Pascual-Seva, N. Water uptake and germination of caper (Capparis spinosa L.) seeds. Agronomy 2020, 10, 838. [Google Scholar] [CrossRef]
- Foschi, M.L.; Juan, M.; Pascual, B.; Pascual-Seva, N. Collection guidelines to achieve a viable caper commercial propagation. Agronomy 2022, 12, 74. [Google Scholar] [CrossRef]
- Takano, M.; Inagaki, N.; Xie, X.; Kiyota, S.; Baba-Kasai, A.; Tanabata, T.; Shinomura, T. Phytochromes are the sole photoreceptors for perceiving red/far-red light in rice. Proc. Natl. Acad. Sci. USA 2009, 106, 14705–14710. [Google Scholar] [CrossRef] [Green Version]
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.M.; Nonogaki, H. Seeds: Physiology of Development, Germination and Dormancy, 3rd ed.; Springer: New York, NY, USA, 2013; ISBN 978-1-4614-4692-7. [Google Scholar]
- Meisel, L.A.; Urbina, D.C.; Pinto, M.E. Fotorreceptores y respuestas de plantas a señales lumínicas. Fisiol. Veg. 2011, 18, 1–10. [Google Scholar]
- Casal, J.J. Fotomorfogénesis: La luz como factor regulador del crecimiento. In Fundamentos de Fisiología Vegetal; Azcón-Bieto, J., Talón, M., Eds.; McGraw-Hill–Interamericana, S.L.: Madrid, Spain; Publicacions i Edicions de la Universitat De Barcelona: Barcelona, Spain, 2013; pp. 467–481. ISBN 978-84-481-5168-3. [Google Scholar]
- Butler, W.L.; Norris, K.H.; Siegelman, H.W.; Hendricks, S.B. Detection, assay, and preliminary purification of the pigment controlling photoresponsive development of plants. Proc. Natl. Acad. Sci. USA 1959, 45, 1703–1708. [Google Scholar] [CrossRef]
- Sharrock, R.A.; Quail, P.H. Novel phytochrome sequences in Arabidopsis thaliana: Structure, evolution, and differential expression of a plant regulatory photoreceptor family. Genes Dev. 1989, 3, 1745–1757. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Lin, C. Mechanisms of cryptochrome-mediated photoresponses in plants. Annu. Rev. Plant Biol. 2020, 71, 103–129. [Google Scholar] [CrossRef] [Green Version]
- Orozco-Segovia, A. Fisiología y ecología del fitocromo: Su función en las semillas. Bot. Sci. 1989, 49, 71–84. [Google Scholar] [CrossRef] [Green Version]
- Castillo, M.L.C.; Bustamante, R.O.; Peña-Gómez, F.T.; Gutiérrez, V.L.; Reyes, C.A.; Arredondo-Núñez, A.; Marey, M. Negative photoblastism in the invasive species Eschscholzia californica Cham. (Papaveraceae): Patterns of altitudinal variation in native and invasive range. Gayana Bot. 2013, 70, 331–337. [Google Scholar] [CrossRef] [Green Version]
- Milberg, P.; Andersson, L.; Thompson, K. Large-seeded spices are less dependent on light for germination than small-seeded ones. Seed Sci. Res. 2000, 10, 99–104. [Google Scholar] [CrossRef] [Green Version]
- Takaki, M. New proposal of classification of seeds based on forms of phytochrome instead of photoblastism. Rev. Bras. Fisiol. Veg. 2001, 13, 104–108. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Academic Press: San Diego, CA, USA, 2014; ISBN 978-0-12-416683-7. [Google Scholar]
- Mathews, S. Phytochrome-mediated development in land plants: Red light sensing evolves to meet the challenges of changing light environments. Mol. Ecol. 2006, 15, 3483–3503. [Google Scholar] [CrossRef]
- Lejeune, P.; Fratamico, A.; Bouché, F.; Huerga-Fernández, S.; Tocquin, P.; Périlleux, C. LED color gradient as a new screening tool for rapid phenotyping of plant responses to light quality. Gigascience 2022, 11, 1–13. [Google Scholar] [CrossRef]
- He, D.; Kozai, T.; Niu, G.; Zhang, X. Light-Emitting Diodes for horticulture. In Light-Emitting Diodes, Solid State Lighting Technology and Application; Li, J., Zhang, G.Q., Eds.; Springer International Publishing AG, Part of Springer Nature: Cham, Germany, 2019; pp. 513–547. ISBN 978-3-319-99211-2. [Google Scholar]
- Aguado, G.D.; Álvarez, M. INTA Mendoza Realiza Ensayos Sobre los Efectos de Iluminación LED en Plantines Hortícolas. Available online: https://inta.gob.ar/noticias/inta-mendoza-realiza-ensayos-sobre-los-efectos-de-iluminacion-led-en-plantines-horticolas-0 (accessed on 6 September 2022).
- Cho, J.Y.; Son, D.M.; Kim, J.M.; Seo, B.S.; Yang, S.Y.; Bae, J.H.; Heo, B.G. Effect of LED as light quality on the germination, growth and physiological activities of broccoli sprouts. J. Bio-Environ. Control 2008, 17, 116–123. [Google Scholar]
- Paniagua, G.; Hernández, C.; Rico, F.; Domínguez, F.A.; Martínez, E.; Martínez, C.L. Effect of high intensity LED light on the germination and growth of broccoli seedlings (Brassica oleracea L.). Polibotánica 2015, 40, 199–212. [Google Scholar] [CrossRef]
- Paradiso, R.; Proietti, S. Light-quality manipulation to control plant growth and photomorphogenesis in greenhouse horticulture: The state of the art and the opportunities of modern LED systems. J. Plant Growth Regul. 2022, 41, 742–780. [Google Scholar] [CrossRef]
- Enache, I.M.; Livadariu, O. Preliminary results regarding the testing of treatments with Light-Emitting Diode (LED) on the seed germination of Artemisia dracunculus L. Sci. Bull. Ser. F Biotechnol. 2016, 20, 51–56. [Google Scholar]
- Tang, D.S.; Hamayun, M.; Khan, A.L.; Jan, S.; Nawaz, Y.; Irshad, M.; Na, Y.E.; Lee, I.J. Exposure to red light, temperature and exogenous gibberellins influenced germination of some winter weeds. Afr. J. Biotechnol. 2012, 11, 273–279. [Google Scholar] [CrossRef]
- Hernandez, A.C.; Dominguez, P.A.; Cruz, O.A.; Ivanov, R.; Carballo, C.A.; Zepeda, B.R. Laser in agriculture. Int. Agrophysics 2010, 24, 407–422. [Google Scholar]
- Krawiec, M.; Dziwulska-Hunek, A.; Kornarzyński, K. The use of physical factors for seed quality improvement of horticultural plants. J. Hortic. Res. 2018, 26, 81–94. [Google Scholar] [CrossRef] [Green Version]
- Swathy, P.S.; Kiran, K.R.; Joshi, M.B.; Mahato, K.K.; Muthusamy, A. He–Ne laser accelerates seed germination by modulating growth hormones and reprogramming metabolism in brinjal. Sci. Rep. 2021, 11, 7948. [Google Scholar] [CrossRef]
- Chen, Y.P.; Liu, Y.J.; Wang, X.L.; Ren, Z.Y.; Yue, M. Effect of microwave and He-Ne laser on enzyme activity and biophoton emission of Isatis indigotica Fort. J. Integr. Plant Biol. 2005, 47, 849–855. [Google Scholar] [CrossRef]
- Chen, Y.P.; Yue, M.; Wang, X.-L. Influence of He–Ne laser irradiation on seeds thermodynamic parameters and seedlings growth of Isatis indogotica. Plant Sci. 2005, 168, 601–606. [Google Scholar] [CrossRef]
- Jamil, Y.; Perveen, R.; Ashraf, M.; Ali, Q.; Iqbal, M.; Ahmad, M.R. He–Ne laser-induced changes in germination, thermodynamic parameters, internal energy, enzyme activities and physiological attributes of wheat during germination and early growth. Laser Phys. Lett. 2013, 10, 045606. [Google Scholar] [CrossRef]
- Perveen, R.; Ali, Q.; Ashraf, M.; Al-Qurainy, F.; Jamil, Y.; Raza Ahmad, M. Effects of different doses of low power continuous wave He-Ne laser radiation on some seed thermodynamic and germination parameters, and potential enzymes involved in seed germination of sunflower (Helianthus annuus L.). Photochem. Photobiol. 2010, 86, 1050–1055. [Google Scholar] [CrossRef] [PubMed]
- Podleśny, J.; Stochmal, A.; Podleśna, A.; Misiak, L.E. Effect of laser light treatment on some biochemical and physiological processes in seeds and seedlings of white lupine and faba bean. Plant Growth Regul. 2012, 67, 227–233. [Google Scholar] [CrossRef]
- Soliman, A.S.; Harith, M.A. Effects of laser biostimulation on germination of Acacia farnesiana (L.) Willd. Acta Hortic. 2009, 854, 41–50. [Google Scholar] [CrossRef]
- Demotes-Mainard, S.; Péron, T.; Corot, A.; Bertheloot, J.; Le Gourrierec, J.; Pelleschi-Travier, S.; Crespel, L.; Morel, P.; Huché-Thélier, L.; Boumaza, R.; et al. Plant responses to red and far-red lights, applications in horticulture. Environ. Exp. Bot. 2016, 121, 4–21. [Google Scholar] [CrossRef]
- Finch-Savage, W.E.; Leubner-Metzger, G. Seed dormancy and the control of germination. New Phytol. 2006, 171, 501–523. [Google Scholar] [CrossRef]
- Davies, F.T.; Geneve, R.L.; Wilson, S.B. Hartmann and Kester’s Plant Propagation Principles and Practices, 9th ed.; Pearson Education: New York, NY, USA, 2018; ISBN 13: 978-0-13-448089-3. [Google Scholar]
- Pascual, B.; San Bautista, A.; López-Galarza, S.; Alagarda, J.; Maroto, J.V. Germination behavior after storage of caper seeds. Seed Sci. Technol. 2006, 34, 151–159. [Google Scholar] [CrossRef] [Green Version]
- ISTA. ISTA Working Sheets on Tetrazolium Testing. Tree and Shrub Species, 1st ed.; The International Seed Testing Association (ISTA): Bassersdorf, Switzerland, 2003; Volume II, ISBN 3-906549-41-1. [Google Scholar]
- ISTA. International Seed Testing Association (ISTA); ISTA Rules; ISTA: Bassersdorf, Switzerland, 2018; ISBN 3906549275. ISSN 0251-0952. [Google Scholar]
- Pascual, B.; San Bautista, A.; Ferreros, N.; López-Galarza, S.; Maroto, J.V. Analysis of germination of caper seeds as influenced by the position of fruit on the mother plant, fruit maturation stage and fruit weight. J. Hortic. Sci. Biotechnol. 2003, 78, 73–78. [Google Scholar] [CrossRef]
- Torres, M.; Frutos, G. Logistic function analysis of germination behaviour of aged fennel seeds. Environ. Exp. Bot. 1990, 30, 383–390. [Google Scholar] [CrossRef]
- Causton, D.R.; Venus, J.C. Single leaf growth and the Richards function: Methodology. In The Biometry of Plant Growth; Causton, D.R., Venus, J.C., Eds.; Edward Arnold: London, UK, 1981; pp. 87–143. ISBN 978-0-7131-2812-3. [Google Scholar]
- Rasband, W.; Ferreira, T. ImageJ. Programa de Procesamiento de Imágenes Diseñado para Imágenes Científicas; National Institutes of Health: Bethesda, MD, USA, 2016.
- Juan, M. Estudio para la Mejora de las Técnicas de Propagación de la Alcaparra (Capparis spinosa L.). Ph.D. Thesis, Universitat Politècnica de València, València, España, 2017. [Google Scholar]
- Statgraphics Statistical Graphics Statgraphics Centurion XVIII; Statistical Graphics Corporation: Rockville, MD, USA, 2018.
- Pascual, B.; San Bautista, A.; Pascual Seva, N.; García Molina, R.; López-Galarza, S.; Maroto, J.V. Effects of soaking period and gibberellic acid addition on caper seed germination. Seed Sci. Technol. 2009, 37, 33–41. [Google Scholar] [CrossRef]
- Germanà, M.A.; Chiancone, B. In vitro germination and seedling development of caper (Capparis spinosa L.) mature seeds. Acta Hortic. 2009, 839, 181–186. [Google Scholar] [CrossRef]
- Marín, L. Estudio de la Influencia de Diferentes Longitudes de onda de luz LED en la Germinación de una Orquídea Encyclia sp. Bachelor’s Thesis, Universidad Federal de Integración Latino-Americana, Foz do Iguaçu, Brasil, 2016. [Google Scholar]
- Moreno, A.; Loza, S.; Ortiz, M. Efecto de luz LED sobre semillas de Capsicum annuum L. var. serrano. Biotecnol. Veg. 2017, 17, 145–151. [Google Scholar]
- Álvarez, A.; Ramírez, R.; Chávez, L.; Camejo, Y.; Licea, L.; Porras, E.; García, B. Efecto del tratamiento de semillas con láser de baja potencia, sobre el crecimiento y rendimiento en plantas de tomate (Solanum lycopersicum L.). ITEA Inf. Tec. Econ. Agrar. 2011, 107, 290–299. [Google Scholar]
- Krawiec, M.; Dziwulska-Hunek, A.; Kornarzyński, K.; Palonka, S. Wpływ wybranych czynników fizycznych na kiełkowanie nasion rzodkiewki (Raphanus sativus L.). Acta Agrophysica 2012, 19, 737–748. [Google Scholar]
- Klimek-Kopyra, A.; Dłużniewska, J.; Ślizowska, A.; Dobrowolski, J.W. Impact of coherent laser irradiation on germination and mycoflora of soybean seeds—Innovative and prospective seed quality management. Agriculture 2020, 10, 314. [Google Scholar] [CrossRef]
- Perveen, R.; Wang, X.; Jamil, Y.; Ali, Q.; Ali, S.; Zakaria, M.Q.; Afzaal, M.; Kasana, R.A.; Saleem, M.H.; Fiaz, S. Quantitative determination of the effects of He–Ne laser irradiation on seed thermodynamics, germination attributes and metabolites of safflower (Carthamus tinctorius L.) in relation with the activities of germination enzymes. Agronomy 2021, 11, 1411. [Google Scholar] [CrossRef]
Figure 1.
Logistic model adjusted to the curves of accumulated germination of caper seeds from Experiment 1. Average values of the combination of the seed lot (2019 and 2020) and the exposure to light-darkness of the seeds. A 500 mg L−1 GA3 solution was used to wet the substrate.
Figure 1.
Logistic model adjusted to the curves of accumulated germination of caper seeds from Experiment 1. Average values of the combination of the seed lot (2019 and 2020) and the exposure to light-darkness of the seeds. A 500 mg L−1 GA3 solution was used to wet the substrate.

Figure 2.
Logistic model adjusted to the curves of accumulated germination of caper seeds from Experiment 2. Average values for the seed exposure to white, red, blue or red + blue lights and to darkness. A 500 mg L−1 GA3 solution was used to wet the substrate.
Figure 2.
Logistic model adjusted to the curves of accumulated germination of caper seeds from Experiment 2. Average values for the seed exposure to white, red, blue or red + blue lights and to darkness. A 500 mg L−1 GA3 solution was used to wet the substrate.

Figure 3.
Logistic model adjusted to the curves of accumulated germination of caper seeds from Experiment 3. Average values of the He-Ne laser irradiation during 0, 1, 15, 60 and 180 s. A 500 mg L−1 GA3 solution was used to wet the substrate.
Figure 3.
Logistic model adjusted to the curves of accumulated germination of caper seeds from Experiment 3. Average values of the He-Ne laser irradiation during 0, 1, 15, 60 and 180 s. A 500 mg L−1 GA3 solution was used to wet the substrate.

Figure 4.
Logistic model adjusted to the curves of accumulated germination of caper seeds from Experiment 4. Average values corresponding to the combination of the exposure time of the seeds to the He-Ne laser irradiation (0, 1 or 15 s) after being soaked in water or not. A 500 mg L−1 GA3 solution was used to wet the substrate.
Figure 4.
Logistic model adjusted to the curves of accumulated germination of caper seeds from Experiment 4. Average values corresponding to the combination of the exposure time of the seeds to the He-Ne laser irradiation (0, 1 or 15 s) after being soaked in water or not. A 500 mg L−1 GA3 solution was used to wet the substrate.

Figure 5.
Analysis of the significant interactions of the analysis of variance in Table 5 between irradiation time and seed soaking prior to irradiation on the final germination. Average values of four replicates. Different letters indicate significant differences according to the LSD test. Error bars represent the LSD (p ≤ 0.05).
Figure 5.
Analysis of the significant interactions of the analysis of variance in Table 5 between irradiation time and seed soaking prior to irradiation on the final germination. Average values of four replicates. Different letters indicate significant differences according to the LSD test. Error bars represent the LSD (p ≤ 0.05).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Relative percentage distribution of the different wavelengths for white light (cold white fluorescent lamp) and blue, red, and red + blue LED lights.
Table 1.
Relative percentage distribution of the different wavelengths for white light (cold white fluorescent lamp) and blue, red, and red + blue LED lights.
| Wavelength | White Light | Blue LED | Red LED | Red + Blue LED |
|---|---|---|---|---|
| Violet 300–400 nm | 1.51% | 0.00% | 0.00% | 0.00% |
| Blue 400–500 nm | 30.33% | 97.48% | 2.98% | 30.25% |
| Green 500–600 nm | 43.32% | 1.05% | 7.88% | 5.27% |
| Red 600–700 nm | 22.27% | 1.47% | 80.55% | 58.41% |
| Far Red 700–800 nm | 2.56% | 0.00% | 8.58% | 6.06% |
| R/FR | 8.69 | 0.00 | 9.39 | 9.63 |
Table 2.
Effect of the seed lot and the exposure to light–darkness on the germination parameters: final germination percentage (A, %), time required to reach 50% of final germination (Gt50, d), and average germination rate (k/2; d−1); average values from Experiment 1. A 500 mg L−1 GA3 solution was used to wet the substrate.
Table 2.
Effect of the seed lot and the exposure to light–darkness on the germination parameters: final germination percentage (A, %), time required to reach 50% of final germination (Gt50, d), and average germination rate (k/2; d−1); average values from Experiment 1. A 500 mg L−1 GA3 solution was used to wet the substrate.
| A | Gt50 | k/2 | |
|---|---|---|---|
| Seed lot (L) | |||
| 2019 | 79.87 | 24.33 | 0.098 |
| 2020 | 82.88 | 25.24 | 0.088 |
| Exposure to (E) | |||
| Light | 81.50 | 23.91 | 0.094 |
| Darkness | 81.25 | 25.68 | 0.092 |
| Analysis of Variance | |||
| Factors (degrees of freedom) | % Sum of squares | ||
| L (1) | 16.30 NS | 5.41 NS | 10.18 NS |
| E (1) | 0.12 NS | 20.32 NS | 0.61 NS |
| L × E (1) | 2.49 NS | 3.37 NS | 5.14 NS |
| Residual (12) | 81.09 | 70.90 | 84.08 |
| Standard deviation (+) | 3.88 | 1.92 | 0.02 |
NS: Not significant differences (p ≤ 0.05) according to the LSD test. (+) The standard deviation has been calculated as the square root of the residual mean square.
Table 3.
Effect of the seed lot and the exposure to white, red, blue or red + blue lights and to darkness on the germination parameters: final germination percentage (A, %), time required to reach 50% of final germination (Gt50; d), and average germination rate (k/2; d−1); average values from Experiment 2. A 500 mg L−1 GA3 solution was used to wet the substrate.
Table 3.
Effect of the seed lot and the exposure to white, red, blue or red + blue lights and to darkness on the germination parameters: final germination percentage (A, %), time required to reach 50% of final germination (Gt50; d), and average germination rate (k/2; d−1); average values from Experiment 2. A 500 mg L−1 GA3 solution was used to wet the substrate.
| A | Gt50 | k/2 | |
|---|---|---|---|
| Seed Lot (L) | |||
| 2019 | 78.0 | 28.1 | 0.077 |
| 2020 | 77.7 | 26.1 | 0.080 |
| Exposure to Lighting (EL) | |||
| White | 80.3 | 29.6 | 0.073 |
| Red | 76.3 | 27.7 | 0.083 |
| Blue | 75.7 | 28.5 | 0.085 |
| Red + Blue | 76.8 | 26.2 | 0.081 |
| Darkness | 80.1 | 23.4 | 0.073 |
| Analysis of variance | |||
| Factors (degrees of freedom) | % Sum of squares | ||
| L (1) | 0.1 NS | 5.2 NS | 0.7 NS |
| EL (4) | 12.3 NS | 23.2 NS | 13.0 NS |
| L × EL (4) | 2.5 NS | 4.5 NS | 9.5 NS |
| Residual (30) | 85.2 | 67.1 | 76.7 |
| Standard deviation (+) | 5.9 | 4.2 | 0.01 |
NS: Not significant differences (p ≤ 0.05) according to the LSD test. (+) The standard deviation has been calculated as the square root of the residual mean square.
Table 4.
Effect of the He-Ne laser irradiation during 0, 1, 15, 60 and 180 s on the germination parameters: final germination percentage (A, %), time required to reach 50% of final germination (Gt50; d), and average germination rate (k/2; d−1); average values from Experiment 3. A 500 mg L−1 GA3 solution was used to wet the substrate.
Table 4.
Effect of the He-Ne laser irradiation during 0, 1, 15, 60 and 180 s on the germination parameters: final germination percentage (A, %), time required to reach 50% of final germination (Gt50; d), and average germination rate (k/2; d−1); average values from Experiment 3. A 500 mg L−1 GA3 solution was used to wet the substrate.
| A | Gt50 | k/2 | |
|---|---|---|---|
| Time (T) | |||
| 0 s | 72.81 | 23.05 bc | 0.069 |
| 1 s | 78.69 | 20.43 c | 0.071 |
| 15 s | 81.96 | 22.37 c | 0.089 |
| 60 s | 69.86 | 27.41 ab | 0.088 |
| 180 s | 69.10 | 30.68 a | 0.061 |
| Analysis of variance | |||
| Factors (degrees of freedom) | % Sum of squares | ||
| T (4) | 35.58 NS | 66.05 ** | 26.31 NS |
| Residual (15) | 64.42 | 33.95 | 73.69 |
| Standard deviation (+) | 7.82 | 3.08 | 0.021 |
Different letters in the same column within each factor indicate significant differences (p ≤ 0.05) according to the LSD test. **: significance level p ≤ 0.01; NS: Not significant. (+) The standard deviation has been calculated as the square root of the residual mean square.
Table 5.
Effect of He-Ne laser irradiation during 0, 1 and 15 s applied to dry and soaking the seeds in water, for 4 days, on the germination parameters: final germination percentage (A, %), time required to reach 50% of final germination (Gt50; d), and average germination rate (k/2; d−1); average values from Experiment 4. A 500 mg L−1 GA3 solution was used to wet the substrate.
Table 5.
Effect of He-Ne laser irradiation during 0, 1 and 15 s applied to dry and soaking the seeds in water, for 4 days, on the germination parameters: final germination percentage (A, %), time required to reach 50% of final germination (Gt50; d), and average germination rate (k/2; d−1); average values from Experiment 4. A 500 mg L−1 GA3 solution was used to wet the substrate.
| A | Gt50 | k/2 | |
|---|---|---|---|
| Time (T) | |||
| 0 s | 66.0 b | 23.8 | 0.055 |
| 1 s | 77.6 a | 22.9 | 0.051 |
| 15 s | 74.6 a | 21.8 | 0.054 |
| Soaking (S) | |||
| Soaked seeds | 79.4 a | 21.6 | 0.058 |
| Dry seeds | 65.9 b | 24.1 | 0.049 |
| Analysis of variance | |||
| Factors (degrees of freedom) | % Sum of squares | ||
| T (2) | 25.6 ** | 3.4 NS | 1.9 NS |
| S (1) | 48.2 ** | 8.2 NS | 11.3 NS |
| T × S (2) | 13.8 ** | 3.8 NS | 1.3 NS |
| Residual (18) | 12.4 | 84.6 | 85.5 |
| Standard deviation (+) | 4.0 | 4.6 | 0.014 |
Different letters in the same column within each factor indicate significant differences (p ≤ 0.05) according to the LSD test. **: significance level p ≤ 0.01, NS: Not significant. (+) The standard deviation has been calculated as the square root of the residual mean square.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Foschi, M.L.; Juan, M.; Pascual, B.; Pascual-Seva, N. Influence of Lighting and Laser Irradiation on the Germination of Caper Seeds. Agriculture 2022, 12, 1612. https://doi.org/10.3390/agriculture12101612
AMA Style
Foschi ML, Juan M, Pascual B, Pascual-Seva N. Influence of Lighting and Laser Irradiation on the Germination of Caper Seeds. Agriculture. 2022; 12(10):1612. https://doi.org/10.3390/agriculture12101612
Chicago/Turabian StyleFoschi, María Laura, Mariano Juan, Bernardo Pascual, and Nuria Pascual-Seva. 2022. "Influence of Lighting and Laser Irradiation on the Germination of Caper Seeds" Agriculture 12, no. 10: 1612. https://doi.org/10.3390/agriculture12101612
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.