
Improving Leaf Photosynthetic Performance of Apple through a Novel Root-Zone Irrigation in the Loess Plateau

College of Water Resource Science and Engineering, Taiyuan University of Technology, Taiyuan 030024, China
*
Author to whom correspondence should be addressed.
Agriculture 2022, 12(9), 1362; https://doi.org/10.3390/agriculture12091362
Submission received: 14 August 2022
/
Revised: 27 August 2022
/
Accepted: 30 August 2022
/
Published: 1 September 2022
(This article belongs to the Section Agricultural Water Management)
Abstract
:As freshwater becomes an increasingly scarce and expensive natural resource, novel water-saving irrigation methods for dwarfing apple orchards are needed in the Loess Plateau. However, studies are lacking on the effects of novel root-zone irrigation technology on leaf-level photosynthesis, which directly determines the yield and survival ability of trees. In this study, the leaf gas characteristics and water status of five-year-old dwarfing apple trees in the Loess Plateau of China were monitored during the 2016–2017 growing seasons under water storage pit irrigation (WSPI) and surface irrigation (SI) treatments. Under WSPI, the leaf water potential (Ψm), net photosynthesis rate (Pn), stomatal conductance (gs), and chlorophyll content (Chl) were significantly higher than those under SI (by 12.21–28.36%), while non-photochemical quenching, superoxide dismutase, and sucrose were lower. Compared with SI, WSPI improved dwarfing apple yield by 25.4% and 26.7% in 2016 and 2017, respectively. WSPI increased the photosystem II (PSII) activity by increasing the chlorophyll fluorescence features (the potential quantum yield of PSII (Fv/Fm), actual quantum yield (ΦII), photochemical quenching (qP), and electron transport rate (ETR)). Principal component analysis showed that Ψm, Chl, the chlorophyll fluorescence features (qP, ΦII, and ETR), Pn, and gs could represent the leaf photosynthetic difference between WSPI and SI treatments. Results indicated that apple under WSPI could improve its yield through enhancing leaf photosynthetic performance, and water storage pit irrigation is an effective root-zone irrigation method for apple orchards on the Loess Plateau.
1. Introduction
In the main apple regions, there has been an increasing tendency to grow dwarfing apples using high-density planting [1,2], as this cultivation pattern both increases fruit yield and decreases labor costs. Nonetheless, these dwarfing apple orchards in the Loess Plateau of China are characterized by poor irrigation water quality and small and discontinuous land use [3], which has led to high costs associated with equipment and difficulty in operating drip- and micro-irrigation systems in small orchards. For these reasons, surface irrigation (SI), such as border irrigation and flood irrigation, is still the most conventional method [4]. However, the use of SI results in substantial water loss due to evaporation from the soil surface and soil erosion risk increase due to soil compaction [5], which means that it is necessary to develop new water-saving irrigation techniques to improve both water-use efficiency and yields in these small-holding orchards.
Water storage pit irrigation (WSPI) is a novel root-zone irrigation technology, which has been tested in rainfed dwarfing apple orchards on the Loess Plateau [6,7]. WSPI allows irrigation water or rainfall runoff to directly infiltrate into the subsurface soil through special emitters (water storage pits) dug around half of the radius of a tree canopy [8]. To promote the radial movement of soil moisture and to avoid the possibility of deep percolation caused by pit irrigation, each storage pit has an impermeable bottom and an infiltrating pit wall [9]. These strategies combine the advantages of both pit irrigation [10], which is inexpensive and can easily be employed by an orchardist; and subsurface irrigation, which increases the soil water available to the deep root zone [11]. Although WSPI has been applied to improve apple yield and water-use efficiency [12,13], most research has concentrated on the plant–soil interface and the underlying mechanism, such as root growth and soil water distribution [7,14], while little is known about the leaf physiological characteristics under WSPI.
Leaf photosynthesis is the fundamental physiological process that a plant uses for growth and development, and is the ultimate basis of yield formation [15,16]. Irrigation would affect the leaf photosynthetic efficiency of the apple by influencing the soil–plant–atmosphere continuum (SPAC) water potential and root distribution [17,18,19]. Numerous measurements have been performed to determine the leaf photosynthetic performance for varied apple rootstocks under different irrigation levels and irrigation frequencies [20,21,22,23], and most have focused on drip irrigation [24] and partial root-zone drying irrigation [25]. However, few studies have investigated these variations in the dwarfing apple under nonuniform soil water content and root distributions. More studies on the effects of WSPI on leaf photosynthetic characteristics are needed to further clarify the carbon accumulation strategy and formulate reasonable irrigation systems in dwarfing apple orchards.
We previously found that the total fine root length was increased under WSPI, as well as increasing soil water storage in deep soil compared to SI [14,26]. In addition, we found that the depth of root water uptake for apple trees changed from 20 cm to 100 cm under WSPI, while SI mainly used 0–40 cm soil depth of soil water [27]. Deep-rooted plants would influence leaf water status and stomatal regulation [28,29], and then might lead to differences in the photosynthetic rate of apple trees. However, there are few reports on the leaf photosynthetic response to WSPI. Previous studies have shown that the leaf gas exchange characteristics in young or mature apple trees varied for WSPI at different pit depths (20 cm, 40 cm, and 60 cm) and soil water content limits, but it could not indicate whether there was a difference between WSPI and SI. To provide further insight into the relationship between net photosynthetic rate variation and leaf water status under WSPI compared to SI, a two-year field study (2016–2017) was conducted in a dwarfing apple orchard in the Loess Plateau of China. Leaf water potential, soil water content, and stomatal and nonstomatal parameters were measured to determine the differences in carbon accumulation between WSPI and SI. It was hypothesized that WSPI would improve leaf photosynthesis, resulting in an increased apple yield as compared to SI. This study aims to contribute to the scientific basis for the accelerated generalization and adaptation of irrigation theory and technologies under WSPI in the Loess Plateau.
2. Materials and Methods
2.1. Study Site
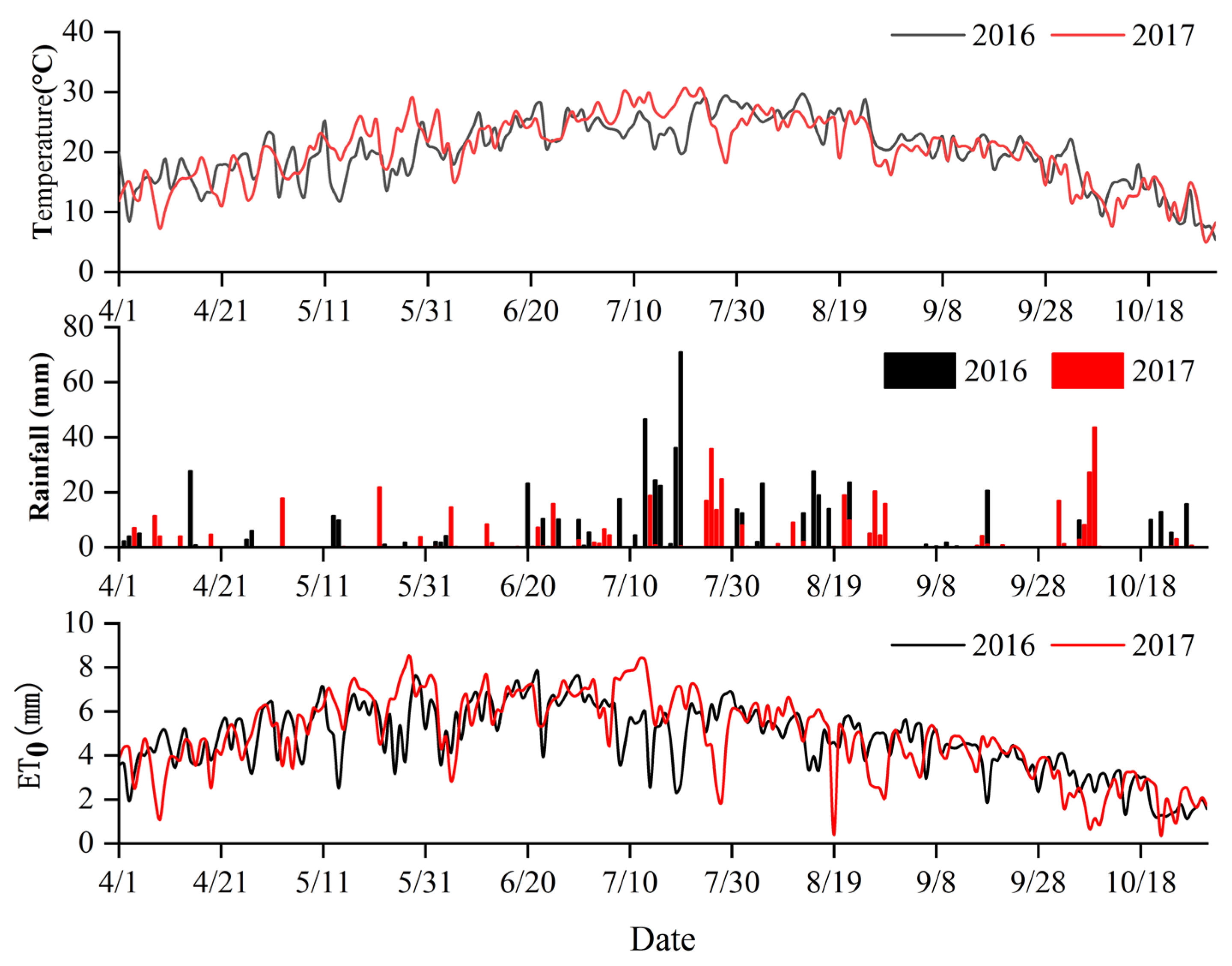
Experiments were conducted in the dwarfing apple interstock orchard at the Shanxi Academy of Agricultural Sciences, located in Taigu County, Jinzhong City, in the Shanxi Province (37°23′ N, 112°32′ E, 781.9 m elevation). The site has a semiarid climate with a mean annual air temperature of approximately 9.8 °C and a mean annual rainfall of 460 mm. The soil texture of 0–100 cm depth is mainly silt loam; the average soil bulk density is 1.47 g cm−3. The soil moisture at the field capacity is 0.30 cm3 cm−3, and at the saturated water content is 50%. Air temperature, rainfall, and reference evapotranspiration (ET0, calculated using the Hargreaves equation) over the course of the experiment from April to October in 2016 and 2017 are shown in Figure 1. Table 1 is the main meteorological factors in the experiment zone.
2.2. Experimental Design
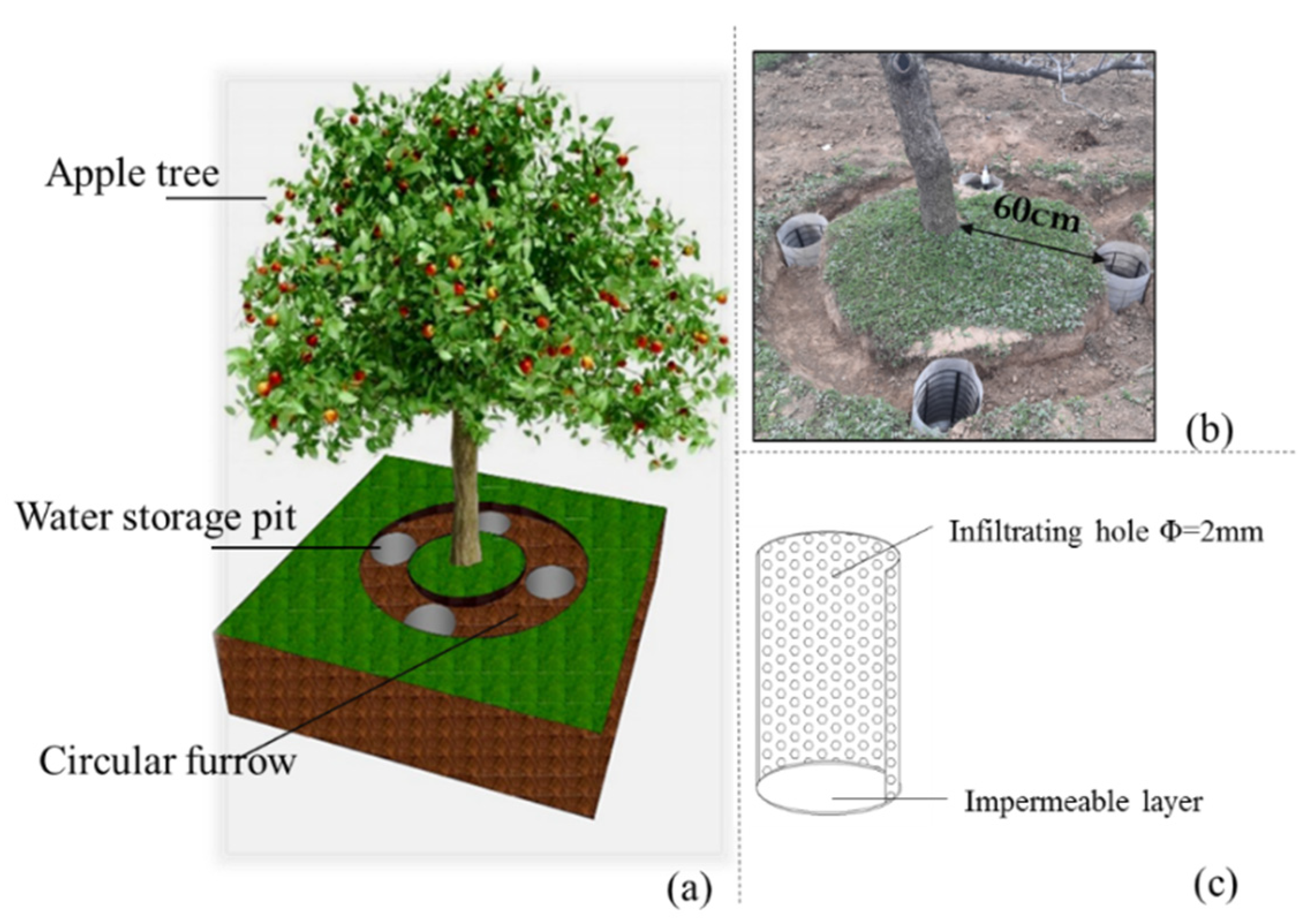
Five-year-old ‘Red Fuji’ (Malus domestica Borkh.) apple trees were scion-grafted onto an SH interstock and crabapple (Malus robusta Rehd) rootstock, then transplanted with a row spacing of 4 m and a plant spacing of 2 m, considering future tree growth and diameter. In April 2016, two irrigation treatments, WSPI (pit diameter 30cm and depth 40 cm, Figure 2) and SI (traditional surface border irrigation, equipped with PE pipe), were randomly applied across the middle of the experimental site (100 × 12 m2), with six replicate trees per treatment. The irrigation date and amount were the same for both treatments. According to local rainfall and farmer’s irrigation habits [27], irrigation was performed on 21 April, 1 June, and 6 September in 2016 and on 19 April, 17 May, 19 June, 19 July, and 16 August in 2017, respectively (Table 1). The total irrigation amount was 135 mm and 225 mm in 2016 and 2017, respectively, which accounted for 61% and 64% of the calculated crop evapotranspiration (ETc). The crop coefficient (Kc) for apple trees and ETc were calculated according to the methodology described by Jaouhari et al. [30]. Each year, the trees were fertilized with 600 kg ha−1 of urea, 150 kg ha−1 of superphosphate and 100 kg ha−1 of potassium sulphate [13]. Other management options such as pruning branches and weeding were kept constant throughout the experiment.
2.3. Data Collection
2.3.1. Gas Exchange Parameters, Leaf Water Potential, and Soil Water Content
For one sunny day of each month in 2016 and 2017, during the growth stage of the apple trees, three fully expanded and sunlight-exposed leaves in the middle of new shoots were sampled randomly for gas exchange parameters from each tree (three randomly selected trees, n = 9). Between 9:00 and 11:00 am, the net photosynthesis rate (Pn), stomatal conductance (gs), intercellular CO2 concentration (Ci), and transpiration rate (Tr) were measured using a LI-COR 6400XT portable photosynthesis system (LI-COR Inc., Lincoln, NE, USA) with a 6400-02B LED source. Water-use efficiency (WUE) was calculated as Pn divided by Tr. The photosynthetic photon flux was set at 1500 μmol m−2 s−1, leaf temperature was 25 ± 2 °C, and relative humidity was 65 ± 5%.
On the same dates, predawn leaf water potential (ΨPL) and midday leaf water potential (ΨM) were measured using a chilled-mirror dewpoint potentiometer (WP4C, Decagon Devices, Inc., Pullman, WA, USA). After measuring Pn, the same leaves were sampled between 11:30 am and 1:00 pm, detached without the veins, and stored in a sealed chamber. The chamber temperature was set as to 25 °C, on fast mode. Weekly, both before and after irrigation, soil water content was measured using a time-domain reflectometry with intelligent microelements (TRIME, IMKO, Ettlingen, Germany). The measurements were performed in 20 cm increments at depths of 0–100 cm. The sites of water determination were installed at a distance of 50 cm from each tree trunk under the SI treatment and through the center of two adjacent pits under the WSPI treatment.
2.3.2. Leaf Chlorophyll Content and Chlorophyll Fluorescence Features
For each month of 2017, leaf chlorophyll (Chl) was measured according to Porra et al. [31]. Undamaged and fully expanded leaves were sampled between 9:00 am and 11:00 pm. A total of 0.2 g of fresh leaf mass was homogenized, filtered, and diluted with 80% acetone. The absorbance of the extract was measured at 645 and 664 nm using a spectrophotometer. Chlorophyll content was calculated according to the following formula:
Chla = 12.72 × D663 × 2.59 × D645
Chlb = 22.88 × D645 × 4.67 × D663
Chl = Chla + Chlb
During the same period, mature leaves from the upper part of three trees per treatment were selected to measure chlorophyll fluorescence. Maximum (Fm) and minimum (F0) fluorescence yields of dark-adapted leaves were recorded in the predawn hours (between 5:00 and 6:00 am), using a LI-COR 6400XT portable photosynthesis system (LI-COR Inc., Lincoln, NE, USA) with an integrated leaf chamber fluorimeter (Li-6400-40, LI-COR Inc.). The maximum light-adapted fluorescence (Fm′) and steady-state light-adapted fluorescence (Fs′) were assessed diurnally. Minimal fluorescence under light conditions (F0′) was calculated using the following:
F0′ = F0/(Fv/Fm+F0/Fm′)
The potential quantum yield of PSII (Fv/Fm) was estimated, where Fv is the maximum variable fluorescence (Fv= Fm − F0). Other data-derived parameters included the actual quantum yield, ΦII = (Fm′ − Fs)/Fm’; photochemical quenching, qP = (Fm′ − Fs)/(Fm′ − F0′); non-photochemical quenching, NPQ = (Fm − Fm′)/Fm′, and electron transport rate, ETR = ΦII × 0.5 × PPFD × 0.85.
2.3.3. Antioxidant Enzyme and Soluble Sugar Content
Simultaneously, 15 undamaged leaves from each tree were collected at midday from both WSPI and SI treatments (three replicates for each treatment). The midrib was removed from the leaves, before they were immediately frozen in liquid nitrogen, lyophilized, and kept at −80 °C until analysis. For sucrose and fructose assays, 0.5 g of leaf dry mass was ground to a fine powder with a mortar and pestle, and extraction was performed three times with 10 mL 80% ethanol in a water bath at 80 °C for 10 min, followed by centrifugation at 4000× g. The pooled supernatants were extracted, combined, and allowed to evaporate until dry at 100 °C, at which point 5 mL of distilled water was added and the solution was filtered (0.45 μm). The content of sucrose and fructose was analyzed using resorcinol colorimetric assay [32].
To measure the amount of the antioxidant enzyme, superoxide dismutase (SOD), 5 g frozen leaf samples were ground to a fine powder in liquid nitrogen and homogenized on ice in 50 mM phosphate buffer (pH 7). The homogenate was centrifuged at 4 °C and 12,000× g for 30 min, to obtain the enzyme extract as the supernatant. A 3 mL total reaction mixture contained 50 mM phosphate buffer (pH 7.8), 13 mM methionine, 75 μmM nitro blue tetrazolium (NBT), 0.1 mM EDTA, 2 μM riboflavin, and 0–200 μL enzyme extract. SOD activity was measured on the basis of its ability to inhibit the photochemical reduction in NBT in the presence of riboflavin in 560 nm light [33]. One unit of SOD activity was defined as the amount of enzyme required to cause a 50% inhibition of NBT photoreduction at 25 °C.
2.4. Data Analysis
Analysis of variance (ANOVA) and correlation analysis were performed using the software package SPSS (IBM SPSS Statistics 26, SPSS Inc., Chicago, IL, USA). The least significant difference (LSD) test was used to compare means, and differences were considered significant at a level of p < 0.05. Principal component analysis (PCA) was used to analyze the best linear combination of variables (including leaf water potential, gas exchange parameters, chlorophyll content, chlorophyll fluorescence features, SOD, sucrose and fructose, total 17 traits in 2017, and n = 204), which would account for the greatest total variance in the data.
3. Results
3.1. Soil Water Content and Leaf Water Potential
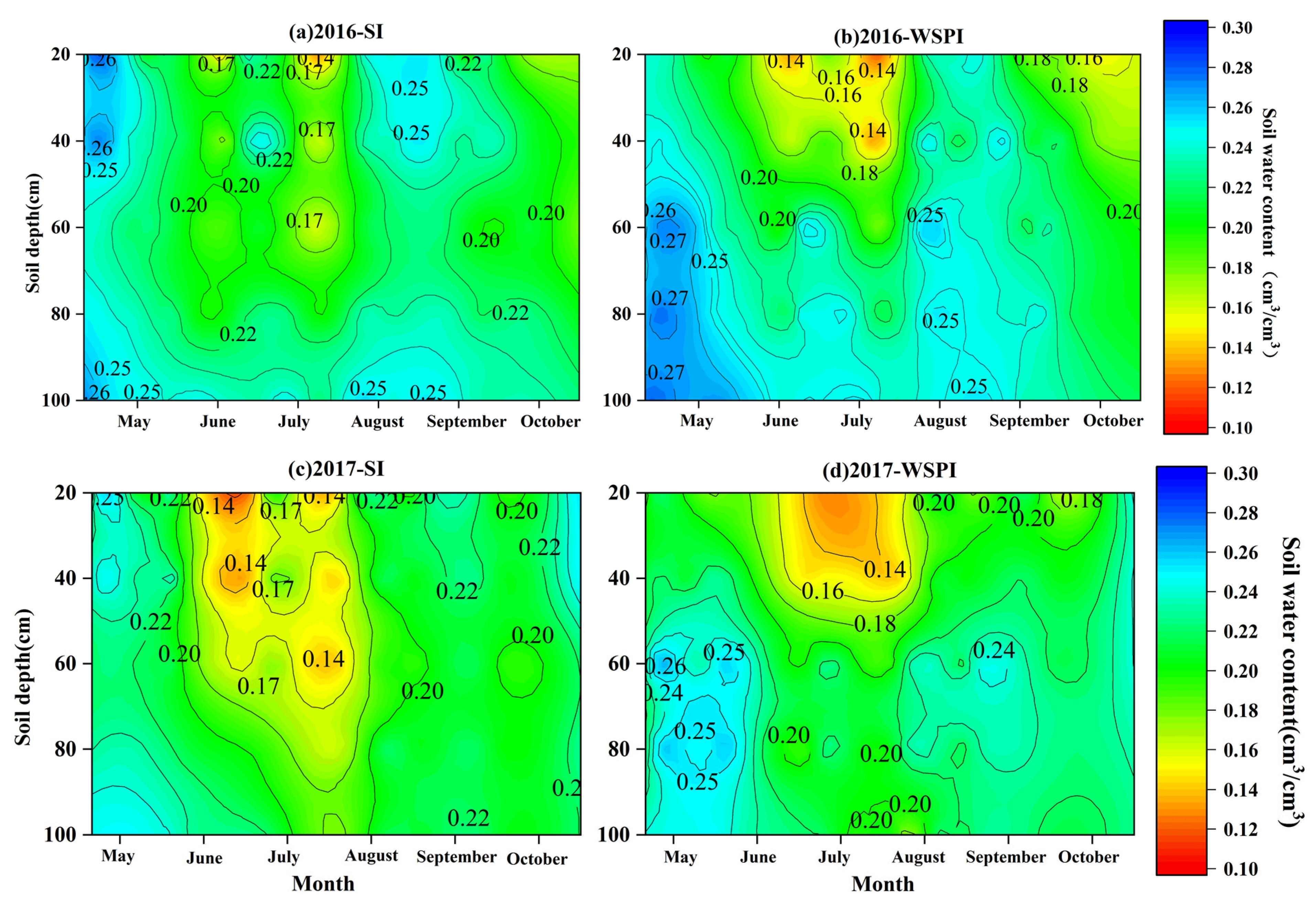
In terms of the 0–100 cm profile, the peak area of soil moisture content under SI was concentrated in 40 cm, while the high-value area under WSPI was basically located at 60–80 cm (Figure 3). In 2016 and 2017, the mean soil moisture content of 0–20 cm under SI was 21.2% and 20.29%, which were 1.12 and 1.13 times higher than those under WSPI, respectively. However, WSPI significantly increased the water content of the 40–80 cm soil layer (p = 0.0221) and kept the soil water content at approximately 23% (76% of field water capacity). Compared with SI, the soil moisture content of 40–80 cm under WSPI increased by 1.09 and 1.14 times in 2016 and 2017, respectively.
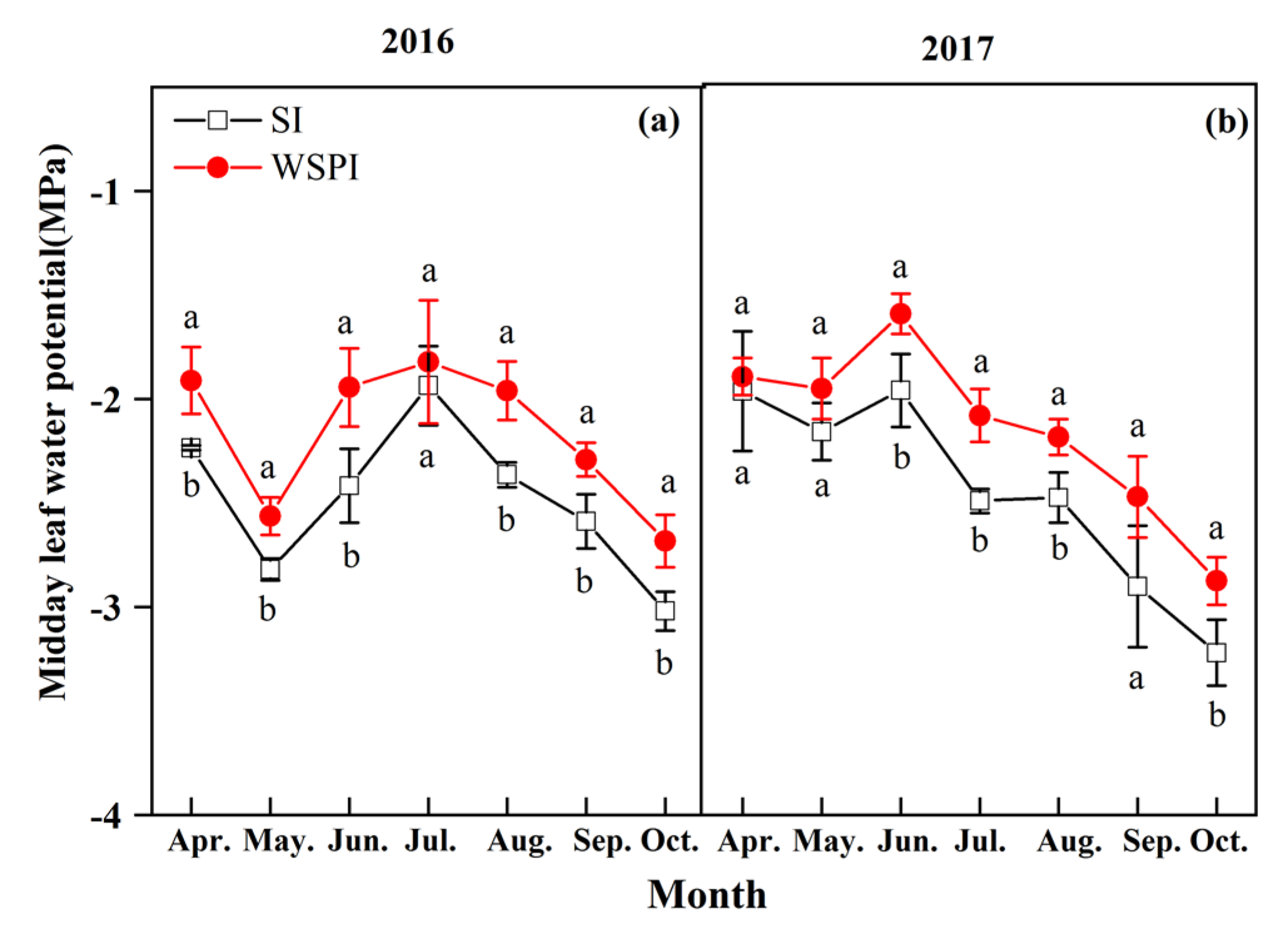
The seasonal variations in midday leaf water potential (Ψm) under different treatments during the period 2016–2017 are shown in Figure 4. There was a significant difference in Ψm between WSPI and SI treatment for both years (p < 0.05), except for July of 2016, for bud development and flowering (April and May) and September of 2017. With the same amount of irrigation, the mean value of Ψm from the WSPI treatment was 12.4 and 11.6% higher in 2016 and 2017, respectively, than that from SI.
3.2. Gas Exchange Characteristics
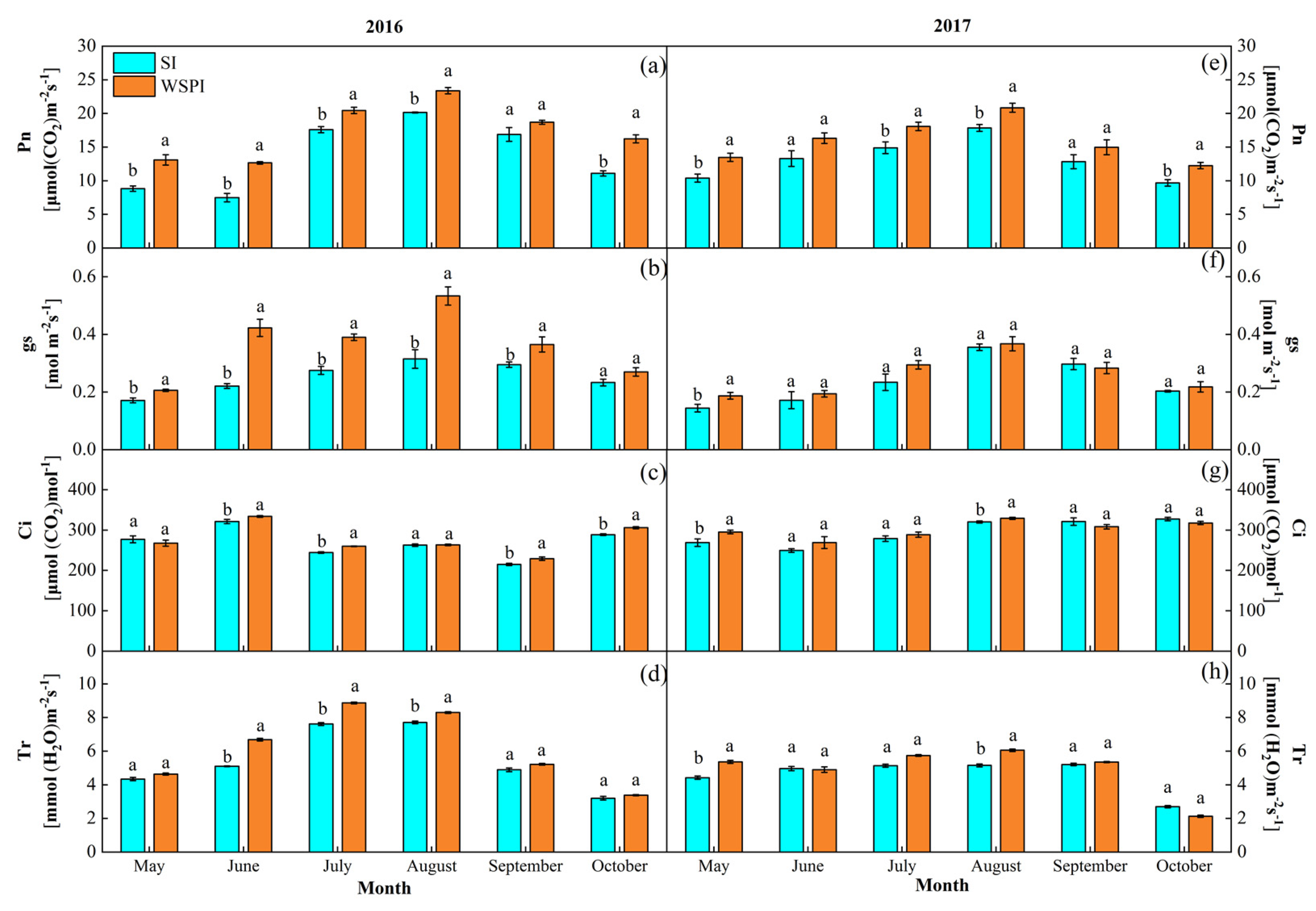
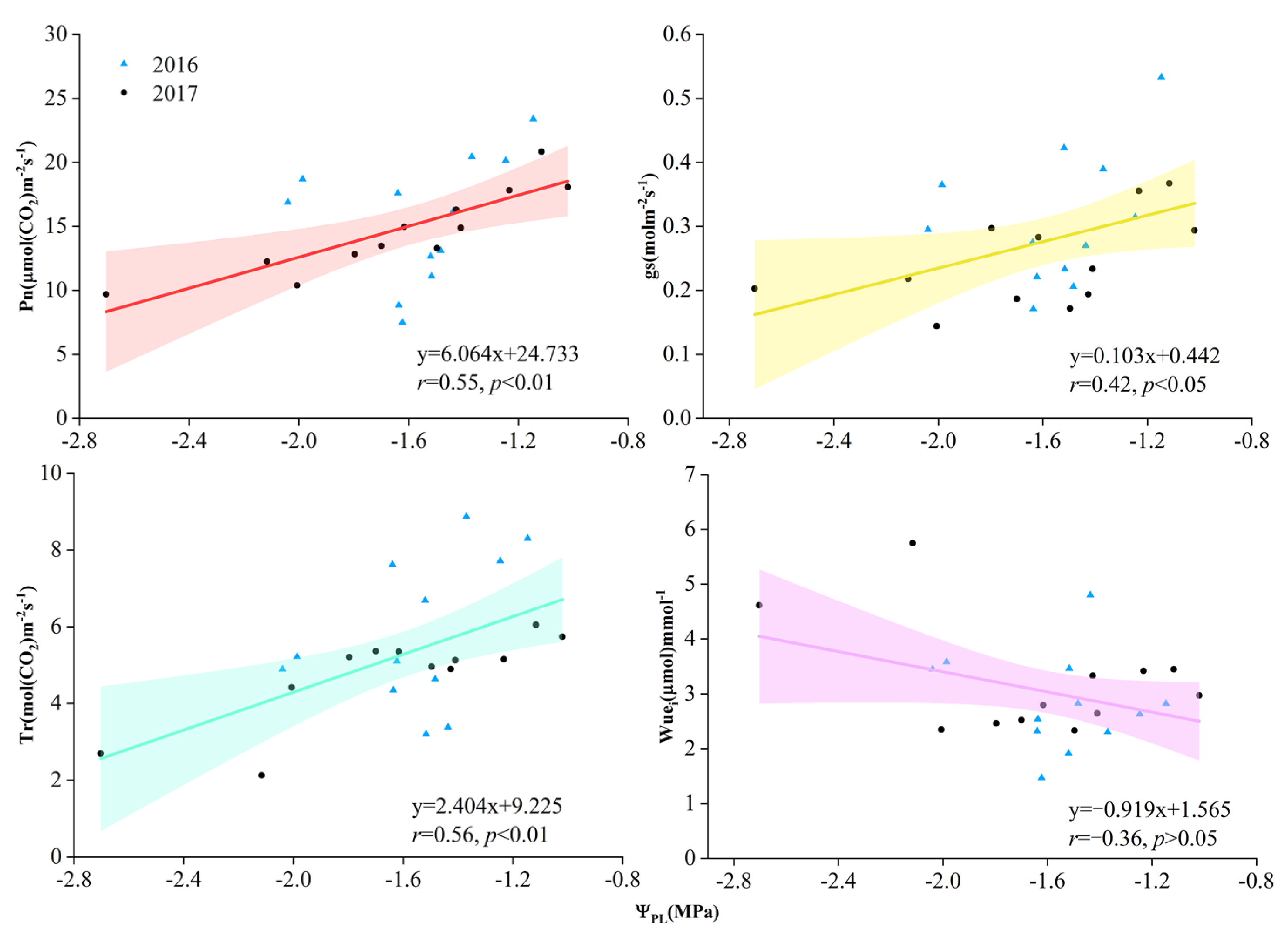
The most significant differences between treatment effects were found during the hottest time of each growing season, usually from July to August (Figure 5). The WSPI treatment exhibited significantly higher Pn than the SI treatment in both years, and the single peak in August was 16.1% and 16.8% greater in 2016 and 2017, respectively (Figure 5a,e). The gs, Tr, and Ci for WSPI were higher than those of SI, and showed a more significant difference in 2016. The mean value of gs, Tr, and Ci for WSPI was greater than that of SI by 28%, 8.7%, and 2.7%, respectively. Significant linear correlations (p < 0.05) were found between leaf gas exchange and predawn leaf water potential (ΨPL) during the growing season in both years (Figure 6), as a higher ΨPL was accompanied by increased A, gs, and Tr. However, the relationship between Wue and ΨPL was not significant.
3.3. Chlorophyll Content and Fluorescence Features of Leaves
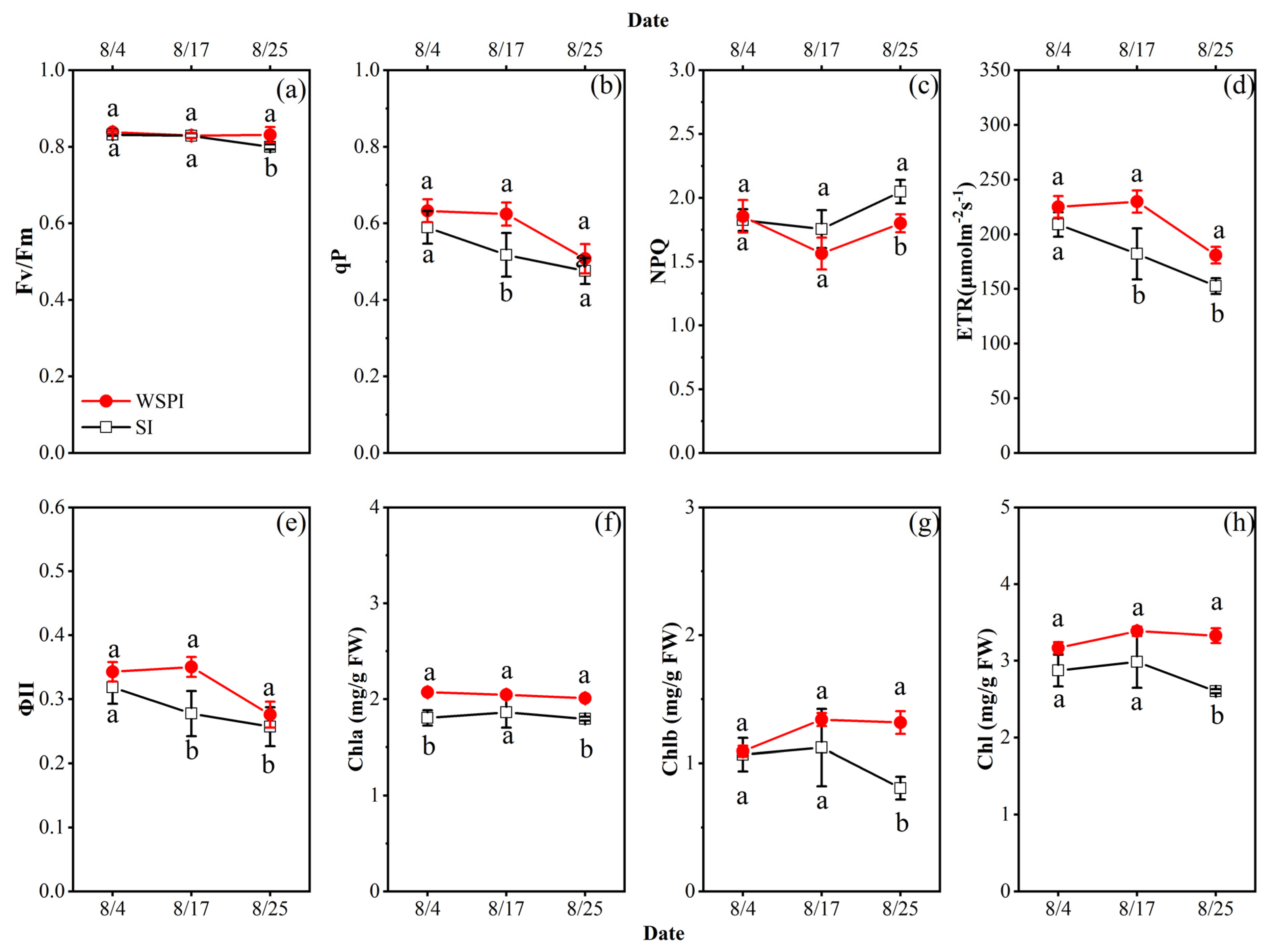
The analysis of the Chl content of leaves in early, mid-, and at the end of August allowed us to study the physiological changes under different treatments (Figure 7). Compared with SI, WSPI treatment increased the content of Chla, Chlb, and total chlorophyll in August of 2017 (Figure 7f–h), as those values were higher by an average of 12.2 (p = 0.001), 25.1 (p = 0.049), and 16.8% (p = 0.014), respectively. The WSPI treatment exhibited a higher Fv/Fm than the SI treatment (p = 0.019), as well as a higher qP (p = 0.008), ΦII (p = 0.008), and ETR (p = 0.002) (Figure 7a,b,d,e). There was no significant difference in the Fv/Fm between WSPI and SI treatments in mid-August (17 August), as both treatments were irrigated (16 August) and not affected by water stress. However, the qP, ΦII, and ETR of leaves in the WSPI treatment were significantly higher at the mid and final measurement points of August than those of the SI. The values of Fv/Fm, qP, ΦII, and ETR decreased during August without considering irrigation treatments, but the magnitude of the decrease in SI was greater than that in WSPI. There was no significant difference in NPQ at the early and mid-points of August between treatments, but at the final point in August, the NPQ of leaves for SI was higher than that of the WSPI treatment.
3.4. Antioxidant Enzyme, Soluble Sugar, and Yield
To explore whether WSPI could delay leaf senescence and reduce the extent of stress, we measured the antioxidant enzyme activity and the content of soluble sugar in August and October of 2017 (the second year after the implementation of WSPI). In October, the leaves entered senescence, showing higher SOD activity than in August (Table 2). Compared with SI, WSPI significantly decreased the activity of SOD in both August and October, by 36.5% and 29.7%, respectively. There was no significant difference in fructose content between WSPI and SI; however, WSPI significantly decreased the sucrose content in August. Compared with SI, WSPI improved dwarfing apple yield by 25.4% and 26.7% in 2016 and 2017, respectively.
3.5. Principal Component Analysis (PCA)
The 17 measured parameters across the different irrigation treatments were analyzed using PCA, and the results are summarized in Table 3. Based on the factor loadings of these traits, the three principal components (PCs) could represent the effects of different irrigation treatments on leaf photosynthetic performance, which explained 86.7% of the total variance. Principle component 1 (PC1) was characterized by positive scores for Ψm, qP, ETR, and ΦII, while PC2 was characterized by positive scores for Pn, gs, and Chl. NPQ was characterized by positive scores for PC3. The leaf water status, Chl fluorescence features, leaf stomata status, and chlorophyll content were sensitive indicators in the dwarfing apple response to WSPI.
4. Discussion
Maintaining a relatively high Ψm was crucial for leaf photosynthesis [34,35]. Our observations were comparable to the literature values of −0.7–−4.0 MPa in apple [36,37,38,39], and based on the results of Sircelj et al. [40], apple trees would experience severe stress when Ψm was −3 MPa or lower. In this study, Ψm did not decrease to this stress level for WSPI treatment, but it did for SI. This result was due to the following factors: (1) Soil water is an important factor that limits the plant water status. It has been demonstrated that Ψm is high when soil water is maintained at a high level due to irrigation [25,41]. In this study, the mean amount of SWC in a 0–100 cm soil profile under WSPI was 1.2 times that under SI treatment. (2) The availability of soil water in deeper soil profiles is indispensable for improving the leaf water content of plants and yield [42]. In this study, more irrigation water was stored at a 40–80 cm soil depth. Previous research at the same site has also shown that compared to SI, WSPI could significantly improve the soil water content of the root zone in the medium soil layer (40–150 cm) [14] and reduce surface evaporation. (3) Deeper plant roots have the ability to reduce the effect of drought stress in the Loess Plateau of China [43]. Then, the development of deep roots could have enabled the shoots to maintain a higher water status [44]. Compared with SI, the fine root length density for WSPI was improved by 10.32% at soil depths from 40 to 100 cm [14], and the average stem flow of apple trees under WSPI was 1.18–1.68 times higher from May to October [45].
The leaf stomatal strategy is regulated by the leaf water status [46], and the gs was directly related to the Pn [47]. To the best of our knowledge, this was the first investigation of the differences between stomatal regulation and photosynthetic capacity between WSPI and SI. We found significant linear correlations between Pn, gs, and ΨPL (the value of correlation coefficients were from 0.42 to 0.56, p < 0.01), which indicated that the variations in Pn for different irrigation treatments were mainly due to stomatal regulation caused by soil moisture and leaf water status [48]. In this study, the Pn, gs, and Tr of leaves for WSPI were greater than those for SI. This observation indicated that stomatal limitation was the major limiting factor in photosynthesis [49]. However, the Ci value for WSPI and SI did not necessarily decrease with gs (r = 0.01, p > 0.05). Based on the physiological mechanism of plant photosynthesis [50], Ci would increase when the gs value increases, while an increase in photosynthetic capacity would consume CO2, thus reducing the correlation between Ci and gs [51,52]. These relationships imply that there are also nonstomatal factors (such as Chl content, chlorophyll fluorescence, and enzymatic activity) affecting the photosynthesis of apple under different irrigation methods [53].
Chlorophyll is the key pigment for photosynthesis [54]. In general, healthy leaves have greater chlorophyll content and a higher photosynthetic rate. In this study, WSPI improved the Chl content of leaves during the growth stage (August) when compared to SI. The Chl content was increased by Ψm, and improved soil nutrients [55]. Li et al. [56] found that WSPI decreased the cumulative NH3 volatilization compared with SI. The significant increase in chlorophyll content could effectively increase the light capture and conversion efficiency [57,58]. Beyond these parameters, the increase in Fv/Fm, qP, ΦII, and ETR under the WSPI treatment indicated that the PSII activity of leaves was improved compared with SI. When subjected to water or temperature stress, the plant would maintain normal photosynthesis through the xanthophyll cycle [53,59,60]. In this study, the rise in NPQ for SI treatment suggested that apple trees under SI went though increasing energy dissipation to protect the photosynthetic system. Furthermore, the rate of chlorophyll degradation for WSPI slowed down when compared to SI (Figure 7h), in effect extending the functional period of effective photosynthetic time.
Under soil water stress or during a period of senescence, oxidative stress was induced by increasing the production of reactive oxygen species (ROS) [61]. SOD, the first step in the system that scavenges excess ROS, plays a central role in protecting cells against photodamage to PSII [62]. The activity of SOD under WSPI was significantly lower than that of SI due to the lower oxidative stress. In this study, the activity of SOD was increased in October when compared to August for both treatments. This increase in activity was an adaptation response to water stress, high light levels, and the lower temperatures in autumn [63]. However, this study found no significant difference in sucrose and fructose levels under WSPI and SI, with the exception of sucrose in August. Unlike other plants, sorbitol is the main soluble sugar of apple leaves and accounted for 60-70% of the photosynthesis in leaves [64]. The content of sucrose and fructose was measured in this study, considering that sorbitol is converted to fructose by sorbitol dehydrogenase, and more than 80% of the total carbon flux via fructose [65]. Further studies are needed to investigate the impacts of WSPI on sorbitol levels in apple leaves.
Under the same total amount of irrigation, WSPI increased the apple yield when compared with SI (Table 2). This is consistent with the results of Zhao et al. [14], in that they also found that the average yield for WSPI with 40 cm of pit depth was 1.17 times higher than that of surface irrigation. Li et al. [10] showed that apple yield for pit irrigation using less total water was not significantly lower than yields using furrow irrigation. The PCA analysis of leaf photosynthetic performance showed that WSPI improved apple yield by affecting the leaf water status, chlorophyll content and fluorescence features, and leaf stomata status. However, other biochemical indices are also important for interpreting the holistic photosynthesis mechanism, such as root growth, leaf area, and trunk diameter [66,67].
5. Conclusions
Based on our results, water storage pit irrigation could improve the leaf photosynthetic performance of dwarfing apple compared with SI treatment, via increasing the soil water content at the middle soil depth, leaf water potential, stomatal conductance, chlorophyll content, and PSII activity. The accumulation of SOD and sucrose was significantly reduced under WSPI than that under SI. These findings contribute to gaining a more in-depth understanding of water-saving mechanisms when using water storage pit irrigation.
Author Contributions
L.Z. edited the original draft; J.M., X.S. and X.G. reviewed and edited the draft; L.Z. and J.M. provided the funding. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financially supported by the National Natural Science Foundation of China (52109061, 52079085), and the Key Research and Development Projects of Shanxi Province (201903D211011).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kviklys, D.; Samupliene, G. Relationships among the rootstock, crop load, and sugar hormone signaling of apple tree, and their effects on biennial bearing. Front. Plant Sci. 2020, 11, 1213. [Google Scholar] [CrossRef]
- Zhao, D.; Yuan, J.; Xu, K.; Cheng, C.; Li, H. Selection of morphological, physiological and biochemical indices: Evaluating dwarfing apple interstocks in cold climate zones. N. Z. J. Crop Hortic. Sci. 2016, 44, 291–311. [Google Scholar] [CrossRef]
- Song, X.; Gao, X.; Zhao, X.; Wu, P.; Dyck, M. Spatial distribution of soil moisture and fine roots in rain-fed apple orchards employing a rainwater collection and infiltration (rwci) system on the loess plateau of china. Agric. Water Manag. 2017, 184, 170–177. [Google Scholar] [CrossRef]
- Zheng, L.; Ma, J.; Sun, X.; Guo, X.; Cheng, Q.; Shi, X. Estimating the root water uptake of surface-irrigated apples using water stable isotopes and the hydrus-1D model. Water 2018, 10, 1624. [Google Scholar] [CrossRef]
- Gao, X.; Zhao, X.; Wu, P.; Yang, M.; Ye, M.; Tian, L.; Zou, Y.; Wu, Y.; Zhang, F.; Siddique, K. The economic-environmental trade-off of growing apple trees in the drylands of China: A conceptual framework for sustainable intensification. J. Clean. Prod. 2021, 296, 126497. [Google Scholar] [CrossRef]
- Guo, X.; Lei, T.; Sun, X.; Ma, J.; Zheng, L.; Zhang, S.; He, Q. Modelling soil water dynamics and root water uptake for apple trees under water storage pit irrigation. Int. J. Agric. Biol. Eng. 2019, 12, 126–134. [Google Scholar] [CrossRef]
- Meng, W.; Sun, X.; Ma, J.; Guo, X.; Lei, T.; Li, R. Measurement and simulation of the water storage pit irrigation trees evapotranspiration in the Loess Plateau. Agric. Water Manag. 2020, 226, 105804. [Google Scholar] [CrossRef]
- Sun, X.; Ma, J.; Guo, X. Water storage pit irrigation technique. In Proceedings of the International Conference on Effective Utilization of Agricultural Soil & Water Resources and Protection of Environment and the 4th Annual Academic Conference of Chinese Society of Agricultural Soil and Water Engineering, Nanjing, China, 16–18 August 2006; Hohai University Press: Nanjing, China, 2007; pp. 211–214. [Google Scholar]
- Sun, X. Effect of water storage pit irrigation on soil and water conservation. In Proceedings of the 12th ISCO Conference, Beijing, China, 26–31 May 2002; Tsinghua University Press: Beijing, China, 2002; Volume 2, pp. 367–369. [Google Scholar]
- Li, T.; Zhang, J. Effect of pit irrigation on soil water content, vigor, and water use efficiency within vineyards in extremely arid regions. Sci. Hortic. 2017, 218, 30–37. [Google Scholar] [CrossRef]
- Nazari, E.; Besharat, S.; Zeinalzadeh, K.; Mohammadi, A. Measurement and simulation of the water flow and root uptake in soil under subsurface drip irrigation of apple tree. Agric. Water Manag. 2021, 255, 106972. [Google Scholar] [CrossRef]
- Zhang, B.; Hu, Y.; Hill, R.; Wu, S.; Song, X. Combined effects of biomaterial amendments and rainwater harvesting on soil moisture, structure and apple roots in a rainfed apple orchard on the Loess Plateau, China. Agric. Water Manag. 2021, 248, 106776. [Google Scholar] [CrossRef]
- Cheng, Q.; Ma, J.; Ren, R.; Zheng, L.; Guo, X.; Sun, X. Effects of fertilization management under WSPI on soil nitrogen distribution and nitrogen absorption in apple orchard in Loess Plateau. Agronomy 2020, 10, 1386. [Google Scholar] [CrossRef]
- Zhao, Y.; Ma, J.; Sun, X.; Guo, X. Spatial distribution of soil moisture and fine roots of apple trees under water storage pit irrigation. J. Irrig. Drain. Eng. 2014, 140, 333–340. [Google Scholar] [CrossRef]
- Ashraf, M.; Harris, P. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Chen, Z.; Tao, X.; Khan, A.; Tan, D.; Luo, H. Biomass accumulation, photosynthetic traits and root development of cotton as affected by irrigation and nitrogen-fertilization. Front. Plant Sci. 2018, 9, 173. [Google Scholar] [CrossRef]
- Kucukyumuk, C.; Kacal, E.; Ertek, A.; Ozturk, G.; Kurttas, Y.S.K. Pomological and vegetative changes during transition from flood irrigation to drip irrigation: Starkrimson Delicious apple variety. Sci. Hortic. 2012, 136, 17–23. [Google Scholar] [CrossRef]
- Ghafari, H.; Hassanpour, H.; Jafari, M.; Besharat, S. Effect of partial root zone irrigation on physiology, water use efficiency, fruit yield, phenolic compounds and antioxidant capacity of apple. Arch. Agron. Soil Sci. 2020, 67, 1521–1538. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, G.; Gu, S.; Xia, J.; Zhao, J. Critical responses of photosynthetic efficiency of goldspur apple tree to soil water variation in semiarid loess hilly area. Photosynthetica 2010, 48, 589–595. [Google Scholar] [CrossRef]
- Sircelj, H.; Tausz, M.; Grill, D.; Batič, F. Detecting different levels of drought stress in apple trees (Malus domestica, Borkh.) with selected biochemical and physiological parameters. Sci. Hortic. 2007, 113, 362–369. [Google Scholar] [CrossRef]
- Fujisawa, H.; Moriya, Y. Influence of rootstock and soil moisture on leaf photosynthesis of apple trees on JM1, JM7, JM8, and M.9 rootstocks. Acta Hortic. 2012, 932, 441–446. [Google Scholar] [CrossRef]
- Bolat, I.; Dikilitas, M.; Ercisli, S.; Ikinci, A.; Tonkaz, T. The effect of water stress on some morphological, physiological, and biochemical characteristics and bud success on apple and quince rootstocks. Sci. World J. 2014, 2014, 769732. [Google Scholar] [CrossRef]
- Tworkoski, T.; Fazio, G.; Glenn, D. Apple rootstock resistance to drought. Sci. Hortic. 2016, 204, 70–78. [Google Scholar] [CrossRef]
- Bhusal, N.; Han, S.; Yoon, T. Impact of drought stress on photosynthetic response, leaf water potential, and stem sap flow in two cultivars of bi-leader apple trees (Malus × domestica Borkh.). Sci. Hortic. 2019, 246, 535–543. [Google Scholar] [CrossRef]
- Zegbe, J.; Behboudian, M.; Clothier, B. Reduced irrigation maintains photosynthesis, growth, yield, and fruit quality in ‘pacific rose’ apple. J. Sustain. Agric. 2007, 30, 125–136. [Google Scholar] [CrossRef]
- Guo, X.; Sun, X.; Ma, J.; Lei, T.; Zheng, L. Effects of water storage pit irrigation on root distribution and root distribution fuction of apple trees. Fresen. Environ. Bull. 2019, 28, 326–338. [Google Scholar]
- Zheng, L.; Ma, J.; Sun, X.; Guo, X.; Li, Y.; Ren, R.; Cheng, Q. Effective root growth zone of apple tree under water storage pit irrigation using stable isotope methodology. Arch. Agron. Soil Sci. 2019, 65, 1521–1535. [Google Scholar] [CrossRef]
- Rouhi, V.; Samson, R.; Lemeur, R.; Van Damme, P. Photosynthetic gas exchange characteristics in three different almond species during drought stress and subsequent recovery. Environ. Exp. Bot. 2007, 59, 117–129. [Google Scholar] [CrossRef]
- Glenn, D. Canopy gas exchange and water use efficiency of ’Empire’ apple in response to particle Film, irrigation, and microclimatic factors. J. Am. Soc. Hortic. Sci. 2010, 135, 25–32. [Google Scholar] [CrossRef]
- Jaouhari, N.; Abouabdillah, A.; Bouabid, R.; Bourioug, M.; Aleya, L.; Chaoui, M. Assessment of sustainable deficit irrigation in a moroccan apple orchard as a climate change adaptation strategy. Sci. Total Environ. 2018, 642, 574–581. [Google Scholar] [CrossRef]
- Porra, R.; Thompson, W.; Kriedmann, P. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim. Biophys. Acta. 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Tian, J.; Hu, Y.; Gan, X.; Zhang, Y.; Hu, X.; Gou, L.; Luo, H.; Zhang, W. Effects of increased night temperature on cellulose synthesis and the activity of sucrose metabolism enzymes in cotton fiber. J. Integr. Agric. 2013, 12, 979–988. [Google Scholar] [CrossRef]
- Dhindsa, R.; Plumb-Dhindsa, P.; Thorpe, T. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Williams, L.; Araujo, F. Correlations among predawn leaf, midday leaf, and midday stem water potential and their correlations with other measures of soil and plant water status in vitis vinifera. J. Am. Soc. Hortic. 2002, 127, 448–454. [Google Scholar] [CrossRef]
- Zhou, B.; Sun, J.; Liu, S.; Jin, W.; Zhang, Q.; Wei, Q. Dwarfing apple rootstock responses to elevated temperatures: A study on plant physiological features and transcription level of related genes. J. Integr. Agric. 2016, 15, 1025–1033. [Google Scholar] [CrossRef]
- Atkinson, C.; Policarpo, M.; Webster, A.; Kingswell, G. Drought tolerance of clonal malus determined from measurements of stomatal conductance and leaf water potential. Tree Physiol. 2000, 20, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Naor, A.; Gal, Y.; Peres, M. The inherent variability of water stress indicators in apple, nectarine and pear orchards, and the validity of a leaf-selection procedure for water potential measurements. Irrig. Sci. 2006, 24, 129–135. [Google Scholar] [CrossRef]
- O’Connell, M.; Goodwin, I. Responses of ‘pink lady’ apple to deficit irrigation and partial rootzone drying: Physiology, growth, yield, and fruit quality. Aust. J. Agric. Res. 2007, 58, 1068–1076. [Google Scholar] [CrossRef]
- Eyles, A.; Bound, S.; Oliver, G.; Corkrey, R.; Hardie, M.; Green, S.; Close, D. Impact of biochar amendment on the growth, physiology and fruit of a young commercial apple orchard. Trees 2015, 29, 1817–1826. [Google Scholar] [CrossRef]
- Sircelj, H.; Tausz, M.; Grill, D.; Batic, F. Biochemical responses in leaves of two apple tree cultivars subjected to progressing drought. J. Plant Physiol. 2005, 162, 1308–1318. [Google Scholar] [CrossRef]
- Egea, G.; Dodd, I.; Gonzálezreal, M.; Domingo, R.; Baille, A. Partial rootzone drying improves almond tree leaf-level water use efficiency and afternoon water status compared with regulated deficit irrigation. Funct. Plant Biol. 2011, 38, 372–385. [Google Scholar] [CrossRef]
- Guo, Z.; Yu, Z.; Wang, D.; Shi, Y.; Zhang, Y. Photosynthesis and winter wheat yield responses to supplemental irrigation based on measurement of water content in various soil layers. Field Crop. Res. 2014, 166, 102–111. [Google Scholar] [CrossRef]
- Li, H.; Si, B.; Ma, X.; Wu, P. Deep soil water extraction by apple sequesters organic carbon via root biomass rather than altering soil organic carbon content. Sci. Total Environ. 2019, 670, 662–671. [Google Scholar] [CrossRef] [PubMed]
- Black, M.; Patterson, K.; Minchin, P.; Gould, K.; Clearwater, M. Hydraulic responses of whole vines and individual roots of kiwifruit (actinidia chinensis) following root severance. Tree Physiol. 2011, 31, 508. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sun, X.; Ma, J.; Guo, X.; An, J.; Li, R. Daily variation of stem flow rate of apple tree and influence factors under water storage pit irrigation. Water Saving Irri. 2017, 5, 11–15. [Google Scholar]
- Solari, L.; Johnson, S.; Dejong, T. Relationship of water status to vegetative growth and leaf gas exchange of peach (prunus persica) trees on different rootstocks. Tree Physiol. 2006, 26, 1333–1341. [Google Scholar] [CrossRef]
- Sun, X.; Yan, H.; Ma, P.; Liu, B.; Zou, Y.; Liang, D.; Ma, F.; Li, P. Responses of young ‘pink lady’ apple to alternate deficit irrigation following long-term drought: Growth, photosynthetic capacity, water-use efficiency, and sap flow. Photosynthetica 2012, 50, 501–507. [Google Scholar] [CrossRef]
- Ma, X.; Jacoby, P.; Sanguinet, K. Improving net photosynthetic rate and rooting depth of grapevines through a novel irrigation strategy in a semi-arid climate. Front. Plant Sci. 2020, 11, 575303. [Google Scholar] [CrossRef]
- Hamed, S.; Lefi, E.; Chaieb, M. Physiological responses of pistacia vera, l. versus pistacia atlantica desf.to water stress conditions under arid bioclimate in tunisia. Sci. Hortic. 2016, 203, 224–230. [Google Scholar] [CrossRef]
- Farquhar, G.; Ehleringer, J.; Hubick, K. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Phys. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Garcia-Sanchez, F.; Syvertsen, J.; Gimeno, V.; Botia, P.; Perez-Perez, J. Responses to flooding and drought stress by two citrus rootstock seedlings with different water-use efficiency. Physiol. Plant. 2007, 130, 532–542. [Google Scholar] [CrossRef]
- Yuan, J.; Xu, M.; Duan, W.; Fan, P.; Li, S. Effects of whole-root and half-root water stress on gas exchange and chlorophyll fluorescence parameters in apple trees. J. Am. Soc. Hortic. Sci. 2013, 138, 395–402. [Google Scholar] [CrossRef]
- Wang, Z.; Li, G.; Sun, H.; Ma, L.; Guo, Y.; Zhao, Z.; Gao, H.; Mei, L. Effects of drought stress on photosynthesis and photosynthetic electron transport chain in young apple tree leaves. Biol. Open 2018, 7, bio035279. [Google Scholar] [CrossRef] [PubMed]
- Sayyad-Amin, P.; Jahansooz, M.; Borzouei, A.; Ajili, F. Changes in photosynthetic pigments and chlorophyll-a fluorescence attributes of sweet-forage and grain sorghum cultivars under salt stress. J. Biol. Phys. 2016, 42, 601–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afonso, S.; Ribeiro, C.; Bacelar, E.; Ferreira, H.; Oliveira, I.; Silva, A.; Goncalves, B. Influence of training system on physiological performance, biochemical composition and antioxidant parameters in apple tree (Malus domestica Borkh.). Sci. Hortic. 2017, 225, 394–398. [Google Scholar] [CrossRef]
- Li, J.; Chen, P.; Sun, X.; Liu, P. Effects of fertigation strategies on water and nitrogen distribution under water storage pit irrigation for orchards. Int. J. Agric. Biol. Eng. 2018, 11, 165–171. [Google Scholar] [CrossRef]
- Baker, N. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89. [Google Scholar] [CrossRef]
- Urban, L.; Aarrouf, J.; Bidel, L. Assessing the effects of water deficit on photosynthesis using parameters derived from measurements of leaf gas exchange and of chlorophyll a fluorescence. Front. Plant Sci. 2017, 8, 2068. [Google Scholar] [CrossRef]
- Ballester, C.; Zarco-Tejada, P.; Nicolas, E.; Alarcon, J.; Fereres, E.; Intrigliolo, D.; Gonzalez-Dugo, V. Evaluating the performance of xanthophyll, chlorophyll and structure-sensitive spectral indices to detect water stress in five fruit tree species. Precis. Agric. 2018, 19, 178–193. [Google Scholar] [CrossRef]
- Petridis, A.; Therios, I.; Samouris, G.; Koundouras, S.; Giannakoula, A. Effect of water deficit on leaf phenolic composition, gas exchange, oxidative damage and antioxidant activity of four greek olive (olea europaea L.) cultivars. Plant Physiol. Biochem. 2012, 60, 1–11. [Google Scholar] [CrossRef]
- Abid, G.; M’hamdi, M.; Mingeot, D.; Aouida, M.; Aroua, I.; Muhovski, Y.; Khaled, S.; Souissi, F.; Mannai, K.; Jebara, M. Effect of drought stress on chlorophyll fluorescence, antioxidant enzyme activities and gene expression patterns in faba bean (Vicia faba L.). Arch. Agron. Soil Sci. 2017, 63, 536–552. [Google Scholar] [CrossRef]
- Silva, E.; Ferreira-Silva, S.; Fontenele, A.; Ribeiro, R.; Viégas, R.; Silveira, J. Photosynthetic changes and protective mechanisms against oxidative damage subjected to isolated and combined drought and heat stresses in jatropha curcas plants. J. Plant Physiol. 2010, 167, 1157–1164. [Google Scholar] [CrossRef]
- Pina, J.; Moraes, R. Gas exchange, antioxidants and foliar injuries in saplings of a tropical woody species exposed to ozone. Ecotoxicol. Environ. Saf. 2010, 73, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Feng, F.; Cheng, L. Expression patterns of genes involved in sugar metabolism and accumulation during apple fruit development. PLoS ONE 2012, 7, e33055. [Google Scholar] [CrossRef] [Green Version]
- Samuoliene, G.; Viškelienė, A.; Sirtautas, R.; Kviklys, D. Relationships between apple tree rootstock, crop-load, plant nutritional status and yield. Sci. Hortic. 2016, 211, 167–173. [Google Scholar] [CrossRef]
- Hernandez-Santana, V.; Fernández, J.; Cuevas, M.; Perez-Martin, A.; Diaz-Espejo, A. Photosynthetic limitations by water deficit: Effect on fruit and olive oil yield, leaf area and trunk diameter and its potential use to control vegetative growth of super-highdensity olive orchards. Agric. Water Manag. 2017, 184, 9–18. [Google Scholar] [CrossRef]
- Yang, Q.; Zhang, F.; Li, F. Effect of different drip irrigation methods and fertilization on growth, physiology and water use of young apple tree. Sci. Hortic. 2011, 129, 119–126. [Google Scholar] [CrossRef]
Figure 1.
Air temperature, rainfall, and reference evapotranspiration (ET0) in the growing seasons of apple in 2016 and 2017.
Figure 1.
Air temperature, rainfall, and reference evapotranspiration (ET0) in the growing seasons of apple in 2016 and 2017.

Figure 2.
Diagram of water storage pit irrigation (WSPI) systems. (a) Field three-dimensional schematic. (b) Actual location of water storage pit arrangement, and the pit edge was 60cm from the trunk. (c) Water storage pit.
Figure 2.
Diagram of water storage pit irrigation (WSPI) systems. (a) Field three-dimensional schematic. (b) Actual location of water storage pit arrangement, and the pit edge was 60cm from the trunk. (c) Water storage pit.

Figure 3.
Seasonal variations of soil water content under surface irrigation (a,c) and water storage pit irrigation (b,d) in 2016 and 2017. WSPI means water storage pit irrigation, and SI means surface irrigation. Color changes indicate the magnitude of soil moisture content.
Figure 3.
Seasonal variations of soil water content under surface irrigation (a,c) and water storage pit irrigation (b,d) in 2016 and 2017. WSPI means water storage pit irrigation, and SI means surface irrigation. Color changes indicate the magnitude of soil moisture content.

Figure 4.
Seasonal course of midday leaf water potential under water storage pit irrigation (WSPI red line) and surface irrigation (SI, black line) in 2016 (a) and 2017 (b). Bars indicate standard deviation of the mean (n = 3); the same letters in figure indicate nonsignificant differences between the treatments, whereas different letters show significant differences (p < 0.05).
Figure 4.
Seasonal course of midday leaf water potential under water storage pit irrigation (WSPI red line) and surface irrigation (SI, black line) in 2016 (a) and 2017 (b). Bars indicate standard deviation of the mean (n = 3); the same letters in figure indicate nonsignificant differences between the treatments, whereas different letters show significant differences (p < 0.05).

Figure 5.
Net photosynthesis rate (Pn), stomatal conductance (gs), intercellular CO2 concentration (Ci), and transpiration rate (Tr) of apple leaves under water storage pit irrigation (WSPI) and surface irrigation (SI) in 2016 (a–d) and 2017 (e–h). Bars indicate standard errors of the mean (n = 9); the same letters in figure indicate nonsignificant differences between the treatments, whereas different letters show significant differences (p < 0.05).
Figure 5.
Net photosynthesis rate (Pn), stomatal conductance (gs), intercellular CO2 concentration (Ci), and transpiration rate (Tr) of apple leaves under water storage pit irrigation (WSPI) and surface irrigation (SI) in 2016 (a–d) and 2017 (e–h). Bars indicate standard errors of the mean (n = 9); the same letters in figure indicate nonsignificant differences between the treatments, whereas different letters show significant differences (p < 0.05).

Figure 6.
Relationship between leaf gas exchange and predawn stem water potential (ΨPL) during the growing season in 2016 and 2017. r indicates correlation coefficients between each pair of tested parameters. Pn means net photosynthesis rate, gs means stomatal conductance, Tr means transpiration rate, and Wue means water-use efficiency.
Figure 6.
Relationship between leaf gas exchange and predawn stem water potential (ΨPL) during the growing season in 2016 and 2017. r indicates correlation coefficients between each pair of tested parameters. Pn means net photosynthesis rate, gs means stomatal conductance, Tr means transpiration rate, and Wue means water-use efficiency.

Figure 7.
The potential quantum yield of PSII (Fv/Fm, shown in a), photochemical quenching (qP, b), non-photochemical quenching (NPQ, c), actual quantum yield (ΦII, d), (e) electron transport rate (ETR, e), and chlorophyll content (f–h) of apple leaves under water storage pit irrigation (WSPI) and surface irrigation (SI) during the August of 2017. The Bars indicate standard errors of the mean (n = 3 for chlorophyll content parameters, and n = 9 for chlorophyll fluorescence parameters); the same letters in figure indicate nonsignificant differences between the treatments, whereas different letters show significant differences (p < 0.05).
Figure 7.
The potential quantum yield of PSII (Fv/Fm, shown in a), photochemical quenching (qP, b), non-photochemical quenching (NPQ, c), actual quantum yield (ΦII, d), (e) electron transport rate (ETR, e), and chlorophyll content (f–h) of apple leaves under water storage pit irrigation (WSPI) and surface irrigation (SI) during the August of 2017. The Bars indicate standard errors of the mean (n = 3 for chlorophyll content parameters, and n = 9 for chlorophyll fluorescence parameters); the same letters in figure indicate nonsignificant differences between the treatments, whereas different letters show significant differences (p < 0.05).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Monthly average solar radiation (Rs), mean relative humidity (RH), mean atmosphere temperature (Ta), mean vapor pressure deficit (VPD), total precipitation (P), total evapotranspiration (ET0), and irrigation amount (I) in 2016 and 2017.
Table 1.
Monthly average solar radiation (Rs), mean relative humidity (RH), mean atmosphere temperature (Ta), mean vapor pressure deficit (VPD), total precipitation (P), total evapotranspiration (ET0), and irrigation amount (I) in 2016 and 2017.
| Year | Month | Rs (W m−1) | RH (%) | Ta (℃) | P (mm) | VPD (kPa) | ET0 (mm) | I (mm) |
|---|---|---|---|---|---|---|---|---|
| April | 239.55 | 55.52 | 16.26 | 48.6 | 0.79 | 130.78 | 45 | |
| May | 234.67 | 59.85 | 18.17 | 24.0 | 0.81 | 168.79 | ||
| June | 265.69 | 73.35 | 23.19 | 76.0 | 0.71 | 187.17 | 45 | |
| 2016 | July | 233.92 | 87.91 | 25.05 | 262.8 | 0.35 | 170.89 | |
| August | 240.54 | 87.64 | 25.24 | 134.2 | 0.35 | 155.29 | ||
| September | 224.33 | 82.37 | 20.18 | 24.2 | 0.39 | 123.81 | 45 | |
| October | 155.75 | 83.39 | 12.95 | 58.5 | 0.21 | 74.19 | ||
| April | 237.13 | 52.63 | 14.69 | 31 | 0.80 | 124.57 | 45 | |
| May | 258.58 | 49.91 | 21.25 | 43.6 | 1.18 | 191.65 | 45 | |
| June | 241.54 | 72.87 | 23.07 | 51.6 | 0.72 | 187.47 | 45 | |
| 2017 | July | 228.24 | 79.65 | 26.75 | 125.4 | 0.67 | 194.26 | 45 |
| August | 214.65 | 88.24 | 23.91 | 95 | 0.33 | 151.73 | 45 | |
| September | 199.51 | 81.01 | 20.29 | 6.6 | 0.42 | 125.15 | ||
| October | 106.32 | 91.23 | 12.08 | 104.2 | 0.12 | 65.83 |
Table 2.
Effects of water storage pit irrigation (WSPI) and surface irrigation (SI) on superoxide dismutase (SOD), sucrose, fructose, yield, and single fruit weight in 2016 and 2017. Different letters show significant differences (p < 0.05).
Table 2.
Effects of water storage pit irrigation (WSPI) and surface irrigation (SI) on superoxide dismutase (SOD), sucrose, fructose, yield, and single fruit weight in 2016 and 2017. Different letters show significant differences (p < 0.05).
| SOD (U/g FW) | Sucrose (mg/g DW) | Fructose (mg/g DW) | Yield (kg/tree) | Single Fruit Weight (g) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| August | October | August | October | August | October | 2016 | 2017 | 2016 | 2017 | |
| SI | 150.24 a | 175.67 a | 11.34 a | 11.41 a | 0.78 a | 0.23 a | 10.28 b | 11.47 b | 183.72 b | 170.64 b |
| WSPI | 95.39 b | 123.56 b | 6.98 b | 11.71 a | 0.77 a | 0.27 a | 12.89 a | 14.53 a | 234.39 a | 213.03 a |
Table 3.
Eigen vectors and eigen values of the first three principal components for different irrigation treatments. The numbers in bold indicate values of factor loading > 0.8.
Table 3.
Eigen vectors and eigen values of the first three principal components for different irrigation treatments. The numbers in bold indicate values of factor loading > 0.8.
| Traits | PC1 | PC2 | PC3 |
|---|---|---|---|
| Ψm | 0.969 | 0.055 | 0.029 |
| Pn | 0.405 | 0.885 | −0.080 |
| gs | −0.354 | 0.869 | 0.212 |
| Ci | −0.708 | 0.476 | 0.313 |
| Tr | 0.574 | 0.629 | 0.415 |
| WUE | −0.849 | 0.038 | 0.291 |
| Fv/Fm | 0.714 | 0.559 | −0.123 |
| qP | 0.966 | 0.065 | 0.159 |
| NPQ | −0.131 | −0.295 | 0.882 |
| ETR | 0.942 | 0.225 | −0.109 |
| ΦII | 0.941 | 0.226 | −0.108 |
| Chla | −0.597 | 0.722 | −0.303 |
| Chlb | −0.498 | 0.782 | −0.148 |
| Chl | −0.554 | 0.803 | −0.223 |
| SOD | 0.053 | −0.706 | −0.298 |
| Sucrose | −0.770 | −0.169 | 0.178 |
| Fructose | 0.685 | 0.220 | 0.452 |
| Eigen value | 7.991 | 5.023 | 1.731 |
| Cumulative% | 47.01 | 76.56 | 86.74 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zheng, L.; Ma, J.; Sun, X.; Guo, X. Improving Leaf Photosynthetic Performance of Apple through a Novel Root-Zone Irrigation in the Loess Plateau. Agriculture 2022, 12, 1362. https://doi.org/10.3390/agriculture12091362
AMA Style
Zheng L, Ma J, Sun X, Guo X. Improving Leaf Photosynthetic Performance of Apple through a Novel Root-Zone Irrigation in the Loess Plateau. Agriculture. 2022; 12(9):1362. https://doi.org/10.3390/agriculture12091362
Chicago/Turabian StyleZheng, Lijian, Juanjuan Ma, Xihuan Sun, and Xianghong Guo. 2022. "Improving Leaf Photosynthetic Performance of Apple through a Novel Root-Zone Irrigation in the Loess Plateau" Agriculture 12, no. 9: 1362. https://doi.org/10.3390/agriculture12091362
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.