
Amino Acid Treatments Induce Adventitious Root Formation in Two Different Genotypes of Campanula portenschlagiana †



Department of Food Science, Aarhus University, 8200 Aarhus, Denmark
*
Authors to whom correspondence should be addressed.
†
Presented at the 1st International Electronic Conference on Horticulturae, 16–30 April 2022; Available online: https://sciforum.net/event/IECHo2022 .
Biol. Life Sci. Forum 2022, 16(1), 8; https://doi.org/10.3390/IECHo2022-12498
Published: 15 April 2022
(This article belongs to the Proceedings of The 1st International Electronic Conference on Horticulturae)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Campanula portenschlagiana is an ornamental plant vegetatively propagated from mother plants’ shoot explants. In this study, we worked with two different genotypes of C. portenschlagiana, Deep Blue Ocean and White Ocean. We found different root abilities between the two genotypes, where Deep Blue Ocean showed a higher percentage of rooted explants when compared with the White Ocean genotype. Plant hormone analyses showed that Deep Blue Ocean has a higher amount of endogenous auxins when compared with the White Ocean genotype. In the White Ocean genotype, L-glutamate and L-tryptophan treatments increased the percentage of rooted explants by 80% and 40% compared with in the control, respectively. L-glutamate and L-tryptophan treatments increased the root biomass of the Deep Blue Ocean genotype. Taken together, our results showed that the amino acids L-glutamate and L-tryptophan could be required in adventitious root formation in C. portenschlagiana. Furthermore, the differences between genotypes can be partially explained by endogenous hormone concentration.
1. Introduction
Campanula portenschlagiana is an ornamental plant originally from the Dalmatian Mountains in Croatia. Like many potted flowering plants, C. portenschlagiana is vegetatively propagated from mother plants’ shoot explants. In C. portenschlagiana explants, adventitious root (AR) formation success depends on the genetic background, where rooting ability can be classified in a range from easy-to-root to difficult-to-root genotypes.
Plant hormones play an important role in the control of AR formation, with auxin being known as the central player in the hormonal network that regulates the AR formation process [1]. Indole-3-acetic acid accumulates at the basal cut site and induces AR formation by interacting with many endogenous factors and exogenous stimuli [2]. Although a large number of studies tried to understand AR formation, most of them were made in Arabidopsis and tomato, and little is known about AR formation in C. portenschlagiana.
Here, we selected two genotypes of C. portenschlagiana to investigate their ability in AR formation. We also analyzed the endogenous concentration of plant hormones at the basal cut site. Finally, we treated both genotypes with amino acids to evaluate their response to form AR through exogenous stimulation. Our results showed dissonant efficiency to form AR among genotypes, which can be somewhat explained by distinct endogenous hormone amounts at the basal cut site. Moreover, the AR formation in the difficult-to-root genotype can be stimulated with exogenous amino acid treatments.
2. Methods
2.1. Experimental Setup
Shoot explants from both Deep Blue Ocean and White Ocean stock plant genotypes of Campanula portenschlagiana were placed in an aeroponic system, consisting of three closed irrigation systems, one for each treatment. For each cultivar, 20 cuttings per treatment per replication were used. The cuttings were cultivated in a nutrient solution, which contained 1 mM CaSO4, 1 mM K2HPO4, 1 mM KH2PO4, 2 mM MgSO4, and micronutrients with the following concentrations: 30 µmol B, 15 µmol Fe, 10 µmol Mn, 5 µmol Zn, 0.75 µmol Cu, and 0.5 µmol Mo, and 1 mM of L-glutamate or 1 mM of L-tryptophan was added in the nutrient solution. As a control, we used the nutrient solution without any amino acids added. The plants were grown in a photoperiod of 16/8 h (day/night), at approximately 23/18 °C (day/night), relative humidity of 70–80%, and 150 μmol m− 2 s− 1 of light intensity.
2.2. Morphological and Physiological Analyses
After 21 days of treatment, cuttings from the Deep Blue Ocean and White Ocean genotypes were evaluated for fresh and dry weight of the root, number of rooted cuttings, length of the root, and number of roots per cutting. The number of rooted cuttings was counted every day from day 0 to day 21. For the dry weight measurement, fresh cuttings were dried at 65 °C in a constant flux oven for 48 h.
2.3. Plant Hormone Analyses
The 1 cm fragments at the basal cut site were collected and immediately frozen in liquid nitrogen. Twenty milligrams of the homogenized sample material were extracted with 80% methanol containing 1% acetic acid. The samples were placed in an ultrasound bath for 45 min. After extraction, the samples were centrifugated at 1500 rpm for 10 min at 20 °C. The supernatant was transferred to another vial, and the pellet was re-extracted with the same process. After the second extraction, the supernatant was combined with the first supernatant obtained. The samples were diluted 4 times and filtered through a syringe filter (0.22 µm).
The standard solutions for the calibration curve were prepared from the mixed stock solution containing indole-3-acetic acid, indole-3-butyric acid, abscisic acid, gibberellic acid 3, and trans-zeatin 6-purine, in the range of 0.095–400 ng/mL. The content of all hormones was detected by QTRAP 4500 LC/LC-MS platform. Analyst Software (version 1.6.1, Concord, ON, Canada) was used for instrument control, data acquisition, and quantification. Three independent biological replicates were analyzed.
2.4. Statistical Analyses
The data were collected from three independent experiments, and an analysis of variance (ANOVA) was performed in R (version 3.3.1, R Core Development Team, 2017, Vienna, Austria). Pairwise comparisons were performed with a t-test. Multiple comparisons were performed with Tukey’s honestly significant difference test after ANOVA.
3. Results and Discussion
3.1. Deep Blue Ocean and White Ocean Explants Have Different Efficiencies When Generating Adventitious Root
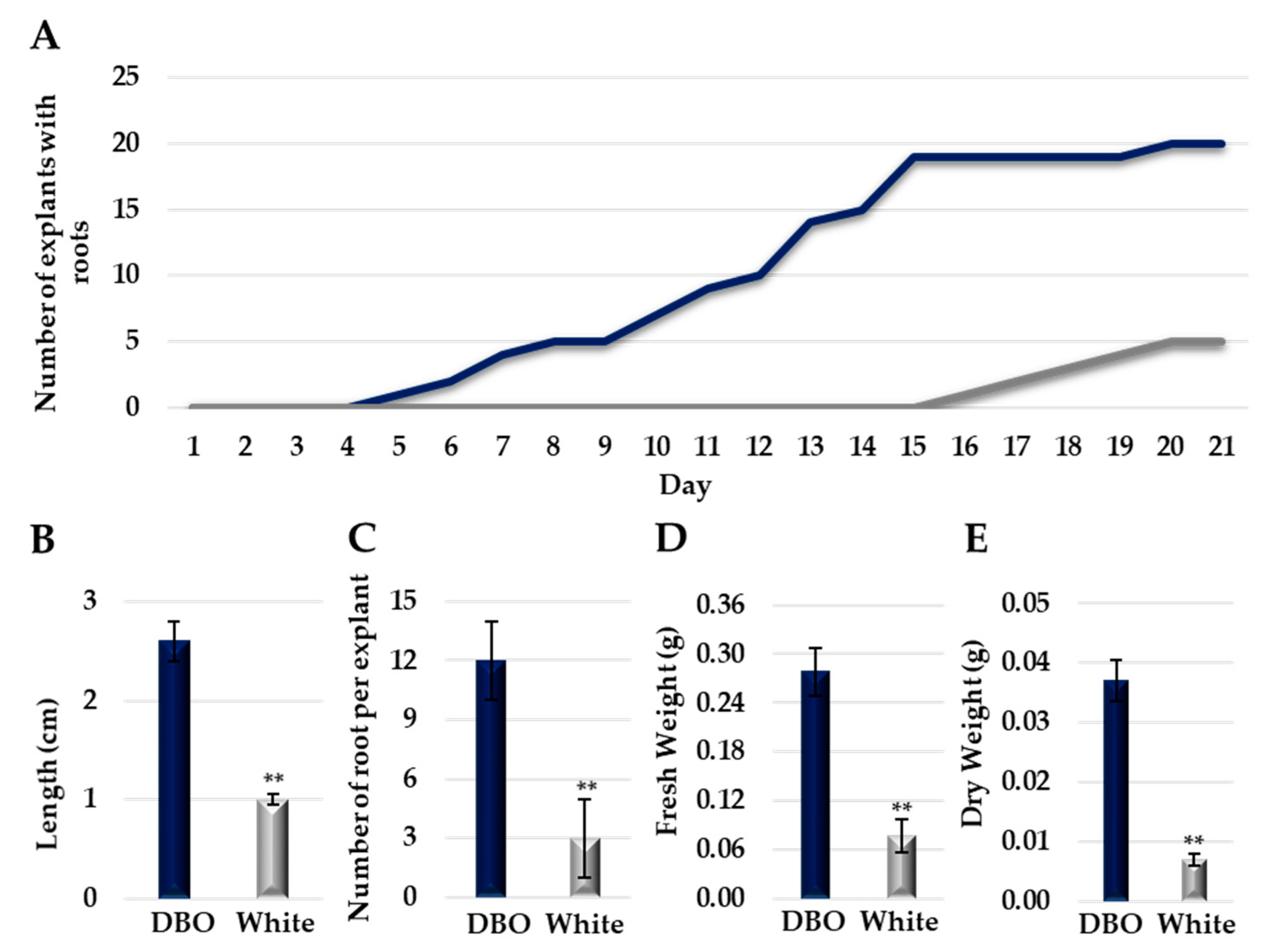
To investigate the ability to form adventitious roots (AR) in Campanula portenschlagiana, both the Deep Blue Ocean and White Ocean genotypes explants were set up in an aeroponic system and root formation was monitored since the first day of the experiment. Deep Blue Ocean explants started to form AR after 4 days of growth in the aeroponic system, while AR formation in White Ocean explants started after 15 days (Figure 1A). After 21 days, 100% of the Deep Blue Ocean explants and 25% of the White Ocean explants formed AR. The average root length in the Deep Blue Ocean genotype was 1.6 times higher than the root length in the White Ocean (Figure 1B). The Deep Blue Ocean genotype showed 4 times more root per explant than the White Ocean genotype (Figure 1C). When we analyzed the root biomass, we found that the fresh and dry weight of the White Ocean genotype was 3.6 and 5.2 times lower than the Deep Blue Ocean genotype, respectively (Figure 1D,E). Our results show that the Deep Blue Ocean genotype has a greater ability to form AR when compared with the White Ocean genotype.
3.2. Deep Blue Ocean and White Ocean Explants Show Contrasting Concentrations of Endogenous Plant Hormones
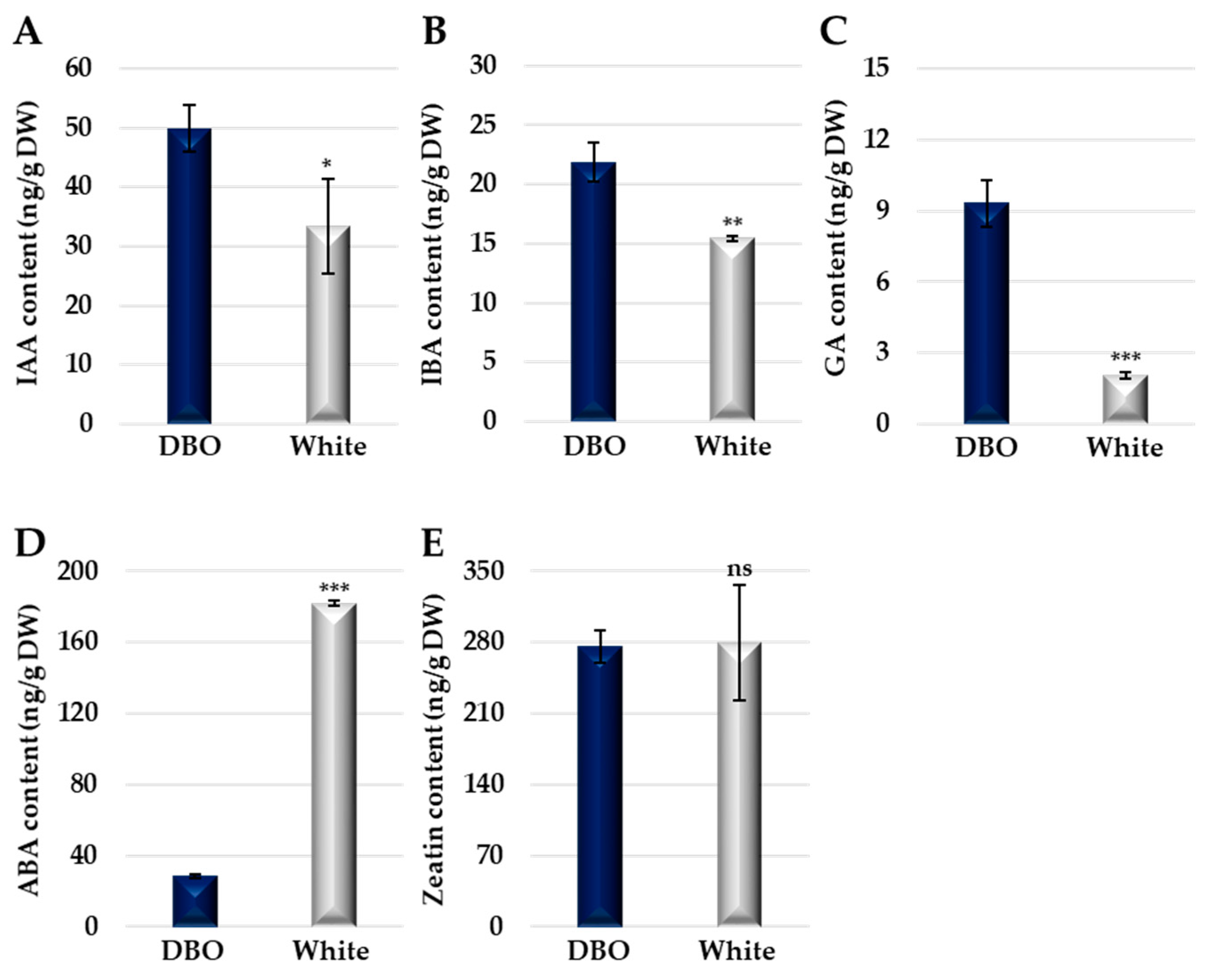
To better understand the differences in AR formation between Deep Blue Ocean and White Ocean, we performed plant hormone analyses at the basal cut site of both genotypes. Our analyses showed that two forms of auxin, indole-3-acetic acid (IAA) and indole-3-butyric acid (IBA), have higher concentrations in the Deep Blue Ocean genotype when compared with the White Ocean genotype (Figure 2A,B). The free level of IAA at the basal cut site in explants was largely demonstrated to be essential to AR formation [3]. Additionally, the conversation of IBA to IAA seems to be an important step in AR formation in many species [4]. The differences in IAA and IBA content between Deep Blue Ocean and White Ocean can partially explain the discrepancy in the AR formation capacity.
Our results showed that the gibberellic acid 3 (GA) content is 4.6 times higher in the Deep Blue Ocean genotype when compared with the White Ocean genotype (Figure 2C). The role of GA in AR formation is largely unclear and contradictory. In Arabidopsis and Populus, studies indicate that GA is a negative regulator of AR formation [5,6], probably interfering with the IAA gradient at the basal cut site. In rice, GA stimulates AR formation via a mechanism that requires ethylene [7]. Although we do not know if GA triggers AR formation in C. portenschlagiana, our results can be indicative that GA participates in this process.
The content of abscisic acid (ABA) was 6.4 times higher in the White Ocean genotype when compared with the Deep Blue Ocean genotype (Figure 2D), suggesting that a lower level of ABA may facilitate AR formation in C. portenschlagiana. In Vaccinium corymbosum, treatment with exogenous auxin increased the AR formation and decreased ABA endogenous content [8]. ABA was shown to inhibit AR emergence in rice and tomato [7,9].
3.3. Amino Acid Treatments Can Increase AR Formation in C. portenschlagiana
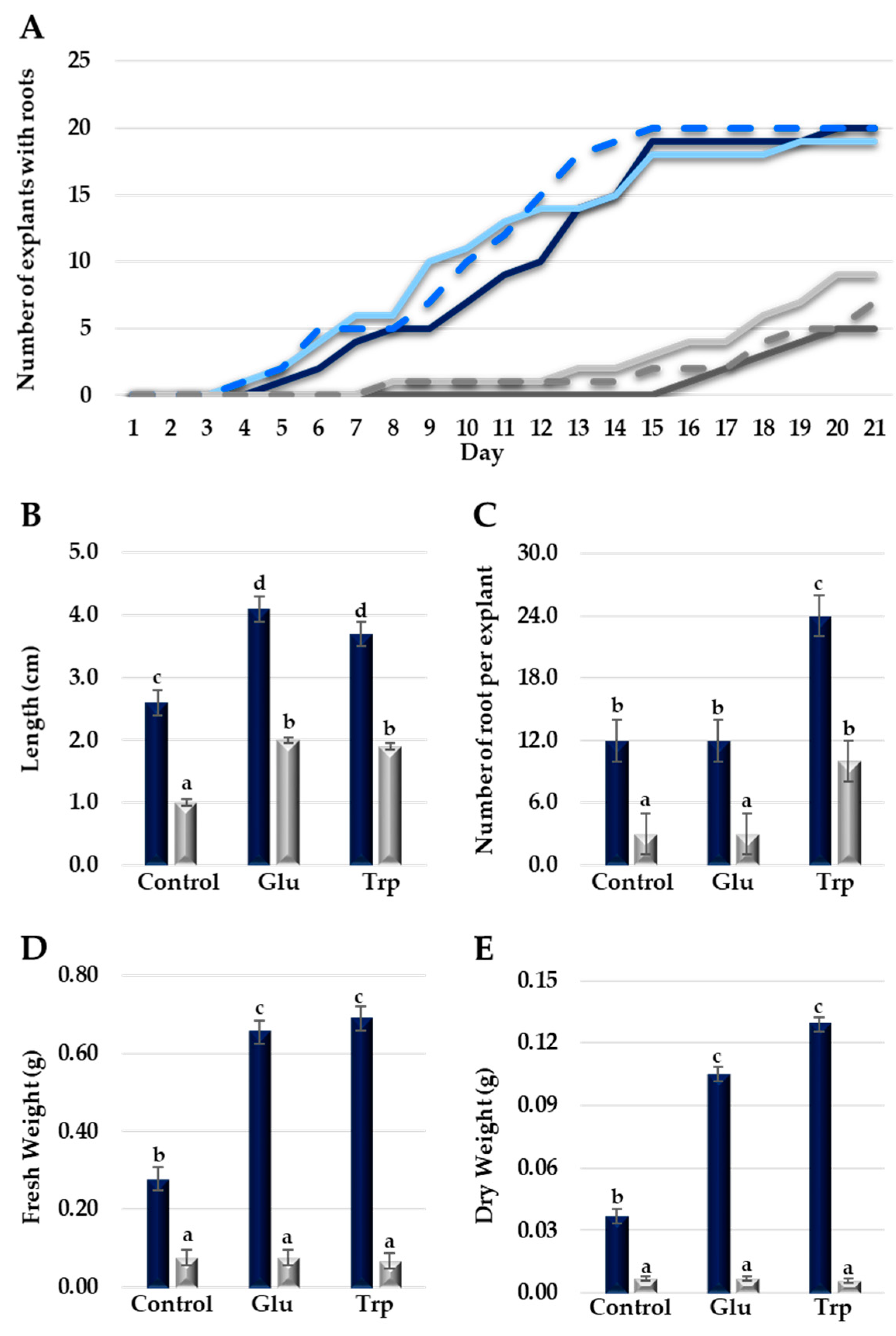
In order to explore the AR formation ability in both the Deep Blue Ocean and White Ocean genotypes, we treated the explants with two different amino acids: L-glutamate and L-tryptophan. L-glutamate and L-tryptophan increased the number of rooted explants in the White Ocean genotype by 80% and 40%, respectively (Figure 3A), whereas in the Deep Blue Ocean genotype, the control and treatments with amino acids showed a 100% increase in rooted explants.
Both the L-glutamate and L-tryptophan treatments increased the root length in the Deep Blue Ocean and White Ocean genotypes (Figure 3B). Compared with the control, L-glutamate did not change the number of roots per explant in both the Deep Blue Ocean and White Ocean genotypes (Figure 3C). Root biomass in the White Ocean genotype did not change in both amino acid treatments when compared with the control (Figure 3D,E). The Deep Blue Ocean genotype showed a significant increment in root biomass in both amino acid treatments when compared with the control (Figure 3D,E). The glutamate and tryptophan amino acids seem to have different pathways to enhance AR formation in plants. Glutamate is an essential source of N remobilization, which is important in lateral root and AR formation [11]. Tryptophan participates in one of the two routes of IAA biosynthesis, and the conversion of tryptophan to IAA can occur via three different pathways [12]. Taken together, our results demonstrate that amino acid treatments can increase AR formation in explants from both easy-to-root and difficult-to-root genotypes in C. portenschlagiana.
4. Conclusions
In the present study, we investigated the adventitious root (AR) formation of two genotypes of Campanula portenschlagiana by using an aeroponic system. We found that differences in endogenous plant hormones could explain the disparity in the AR formation ability between genotypes. We successfully established treatments that increased the AR formation in both genotypes, without using any exogenous plant hormones. The results provide additional evidence for the regulation of the AR formation process in C. portenschlagiana.
Author Contributions
Conceptualization, T.B. and I.A.P.; methodology, T.B. and I.A.P.; software, T.B.; validation, T.B.; formal analysis, T.B.; investigation, T.B.; resources, I.A.P.; data curation, T.B.; writing—original draft preparation, T.B.; writing—review and editing, I.A.P.; visualization, T.B.; supervision, I.A.P.; project administration, T.B. and I.A.P.; funding acquisition, I.A.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Green Development and Demonstration Program (GUDP) grant FUTFLOR (Project number: 34009-20-1752).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors thank Katrine Heinsvig Kjær and Inge Ulsted Sørensen for providing advice, and Jan Uglebjerg Storm and Jacob Morten Andersen for providing the explants of Campanula.
Conflicts of Interest
The authors declare no conflict of interest.
References
- De Klerk, G.J.; Van der Krieken, W.; De Jong, J.C. Review—The formation of adventitious roots: New concepts, new possibilities. Vitr. Cell. Dev. Biol. Plant 1999, 35, 189–199. [Google Scholar] [CrossRef]
- Kang, C.; Zhai, H.; Xue, L.; Zhao, N.; He, S.; Liu, Q. A lycopene β-cyclase gene, IbLCYB2, enhances carotenoid contents and abiotic stress tolerance in transgenic sweetpotato. Plant Sci. 2018, 272, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Fattorini, L.; Falasca, G.; Kevers, C.; Mainero Rocca, L.; Zadra, C.; Altamura, M.M. Adventitious rooting is enhanced by methyl jasmonate in tobacco thin cell layers. Planta 2009, 231, 155–168. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Tong, J.; Xiao, L.; Ruan, Y.; Liu, J.; Zeng, M.; Huang, H.; Wang, J.W.; Xu, L. YUCCA-mediated auxin biogenesis is required for cell fate transition occurring during de novo root organogenesis in Arabidopsis. J. Exp. Bot. 2016, 67, 4273–4284. [Google Scholar] [CrossRef] [PubMed]
- Busov, V.; Meilan, R.; Pearce, D.W.; Rood, S.B.; Ma, C.; Tschaplinski, T.J.; Strauss, S.H. Transgenic modification of gai or rgl1 causes dwarfing and alters gibberellins, root growth, and metabolite profiles in Populus. Planta 2006, 224, 288–299. [Google Scholar] [CrossRef] [PubMed]
- Mauriat, M.; Petterle, A.; Bellini, C.; Moritz, T. Gibberellins inhibit adventitious rooting in hybrid aspen and Arabidopsis by affecting auxin transport. Plant J. 2014, 78, 372–384. [Google Scholar] [CrossRef] [PubMed]
- Steffens, B.; Rasmussen, A. The physiology of adventitious roots. Plant Physiol. 2016, 170, 603–617. [Google Scholar] [CrossRef] [PubMed]
- An, H.; Zhang, J.; Xu, F.; Jiang, S.; Zhang, X. Transcriptomic profiling and discovery of key genes involved in adventitious root formation from green cuttings of highbush blueberry (Vaccinium corymbosum L.). BMC Plant Biol. 2020, 20, 182. [Google Scholar] [CrossRef] [PubMed]
- McAdam, S.A.M.; Brodribb, T.J.; Ross, J.J. Shoot-derived abscisic acid promotes root growth. Plant Cell Environ. 2016, 39, 652–659. [Google Scholar] [CrossRef]
- Gonin, M.; Bergougnoux, V.; Nguyen, T.D.; Gantet, P.; Champion, A. What makes adventitious root? Plants 2019, 8, 240. [Google Scholar] [CrossRef]
- Druege, U.; Hilo, A.; Pérez-Pérez, J.M.; Klopotek, Y.; Acosta, M.; Shahinnia, F.; Zerche, S.; Franken, P.; Hajirezaei, M.R. Molecular and physiological control of adventitious rooting in cuttings: Phytohormone action meets resource allocation. Ann. Bot. 2019, 123, 929–949. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y. Auxin biosynthesis. Arab. Book 2014, 12, e0173. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Adventitious root formation of Deep Blue Ocean and White Ocean genotypes. (A) Number of explants of Deep Blue Ocean (dark blue) and White Ocean (light gray) with adventitious root by day after cutting. (B) Average root length (cm) of adventitious root of Deep Blue Ocean (DBO) and White Ocean (white). (C) Number of roots per explant. (D) Root fresh weight (g). (E) Root dry weight (g). Error bars represent standard deviation. t-test significance: ** p < 0.01; n = 20.
Figure 1.
Adventitious root formation of Deep Blue Ocean and White Ocean genotypes. (A) Number of explants of Deep Blue Ocean (dark blue) and White Ocean (light gray) with adventitious root by day after cutting. (B) Average root length (cm) of adventitious root of Deep Blue Ocean (DBO) and White Ocean (white). (C) Number of roots per explant. (D) Root fresh weight (g). (E) Root dry weight (g). Error bars represent standard deviation. t-test significance: ** p < 0.01; n = 20.

Figure 2.
Plant hormone quantification of Deep Blue Ocean and White Ocean genotypes. (A) Indole-3-acetic acid (IAA) content of Deep Blue Ocean (DBO; dark blue) and White Ocean (White; light gray) in ng/g of dry weight (DW). (B) Indole-3-butyric acid (IBA) content. (C) Gibberellic acid 3 (GA) content. (D) Abscisic acid (ABA) content. (E) Trans-zeatin 6-purine (Zeatin) content. Error bars represent standard deviation. t-test significance: * p < 0.05, ** p < 0.01, *** p < 0.001, ns = no significance; n = 3.
Figure 2.
Plant hormone quantification of Deep Blue Ocean and White Ocean genotypes. (A) Indole-3-acetic acid (IAA) content of Deep Blue Ocean (DBO; dark blue) and White Ocean (White; light gray) in ng/g of dry weight (DW). (B) Indole-3-butyric acid (IBA) content. (C) Gibberellic acid 3 (GA) content. (D) Abscisic acid (ABA) content. (E) Trans-zeatin 6-purine (Zeatin) content. Error bars represent standard deviation. t-test significance: * p < 0.05, ** p < 0.01, *** p < 0.001, ns = no significance; n = 3.

Figure 3.
Effects of L-glutamate and L-tryptophan treatments on Deep Blue Ocean and White Ocean genotypes. (A) Number of explants of Deep Blue Ocean (dark blue) and White Ocean (dark gray), treated with L-glutamate (Deep Blue Ocean: light blue; White Ocean: light gray) or L-tryptophan (Deep Blue Ocean: blue dashed line; White Ocean: gray dashed line) with adventitious roots by day after cutting. (B) Average root length (cm) of adventitious roots of Deep Blue Ocean (dark blue) and White Ocean (light gray) treated with L-glutamate (Glu) or L-tryptophan (Trp). (C) Number of roots per explant. (D) Root fresh weight (g). (E) Root dry weight (g). Error bars represent standard deviation. Statistics were calculated through ANOVA, where mean values with different letters are significantly different at p < 0.05; n = 20.
Figure 3.
Effects of L-glutamate and L-tryptophan treatments on Deep Blue Ocean and White Ocean genotypes. (A) Number of explants of Deep Blue Ocean (dark blue) and White Ocean (dark gray), treated with L-glutamate (Deep Blue Ocean: light blue; White Ocean: light gray) or L-tryptophan (Deep Blue Ocean: blue dashed line; White Ocean: gray dashed line) with adventitious roots by day after cutting. (B) Average root length (cm) of adventitious roots of Deep Blue Ocean (dark blue) and White Ocean (light gray) treated with L-glutamate (Glu) or L-tryptophan (Trp). (C) Number of roots per explant. (D) Root fresh weight (g). (E) Root dry weight (g). Error bars represent standard deviation. Statistics were calculated through ANOVA, where mean values with different letters are significantly different at p < 0.05; n = 20.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bergonci, T.; Paponov, I.A. Amino Acid Treatments Induce Adventitious Root Formation in Two Different Genotypes of Campanula portenschlagiana . Biol. Life Sci. Forum 2022, 16, 8. https://doi.org/10.3390/IECHo2022-12498
AMA Style
Bergonci T, Paponov IA. Amino Acid Treatments Induce Adventitious Root Formation in Two Different Genotypes of Campanula portenschlagiana . Biology and Life Sciences Forum. 2022; 16(1):8. https://doi.org/10.3390/IECHo2022-12498
Chicago/Turabian StyleBergonci, Tábata, and Ivan A. Paponov. 2022. "Amino Acid Treatments Induce Adventitious Root Formation in Two Different Genotypes of Campanula portenschlagiana " Biology and Life Sciences Forum 16, no. 1: 8. https://doi.org/10.3390/IECHo2022-12498