Genome Wide Identification, Characterization, and Expression Analysis of YABBY-Gene Family in WHEAT (Triticum aestivum L.)

 ,
, 
Abstract
:1. Introduction
2. Material and Methods
2.1. Identification of YABBY-Genes in Wheat
2.2. Phylogenetic Tree, Digital Expression and Motif Analysis
2.3. Chromosomal Location and Protein-Protein Interaction of YABBY Genes in ARABIDOSPSIS
2.4. Gene Structure and Conserved Motif Analysis
2.5. Gene Ontology and Cis-Elements Analysis of YABBY Genes
3. Results
3.1. Identification and Analysis of TaYABBY
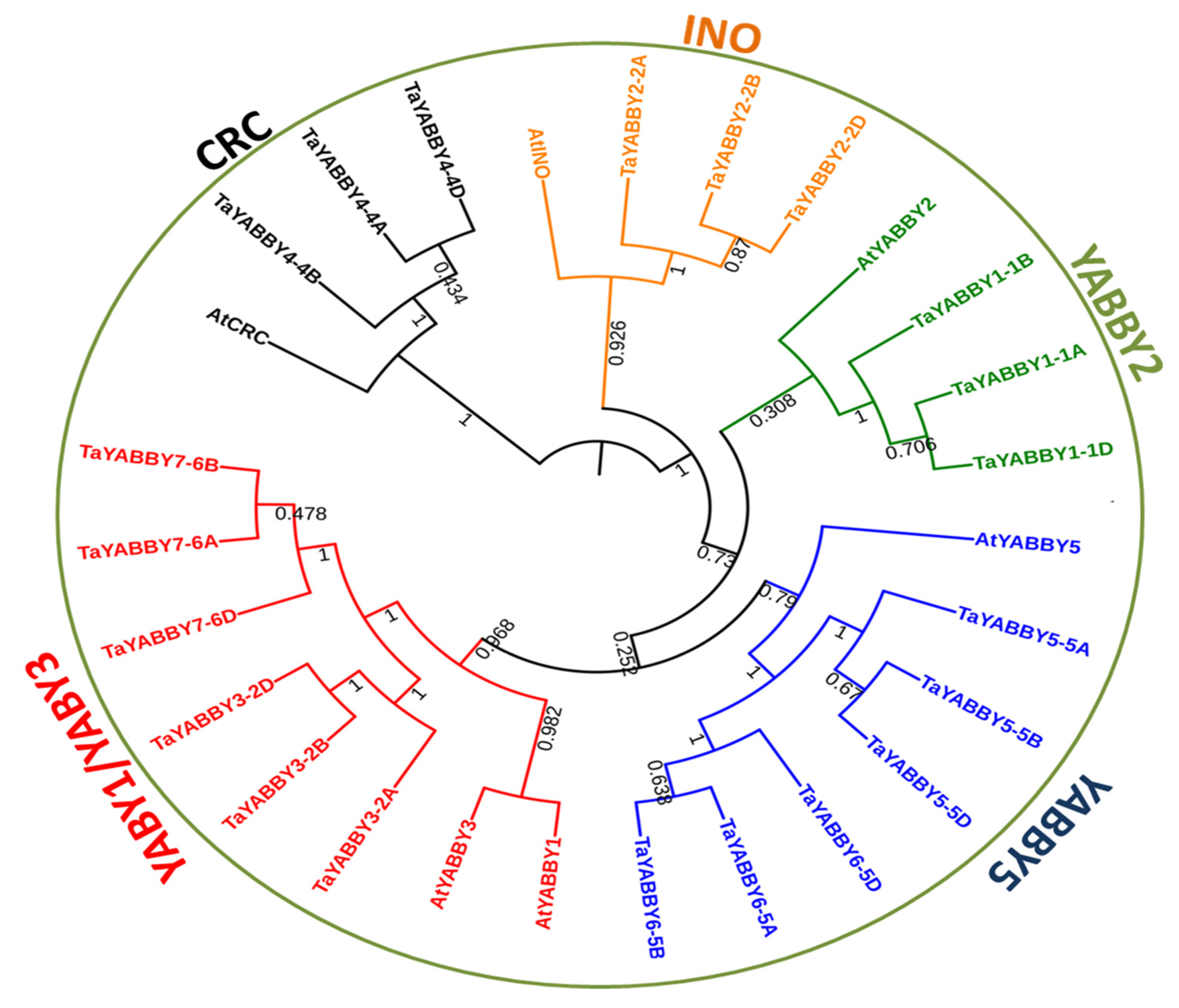
3.2. Evolutionary Relationship of TaYABBY
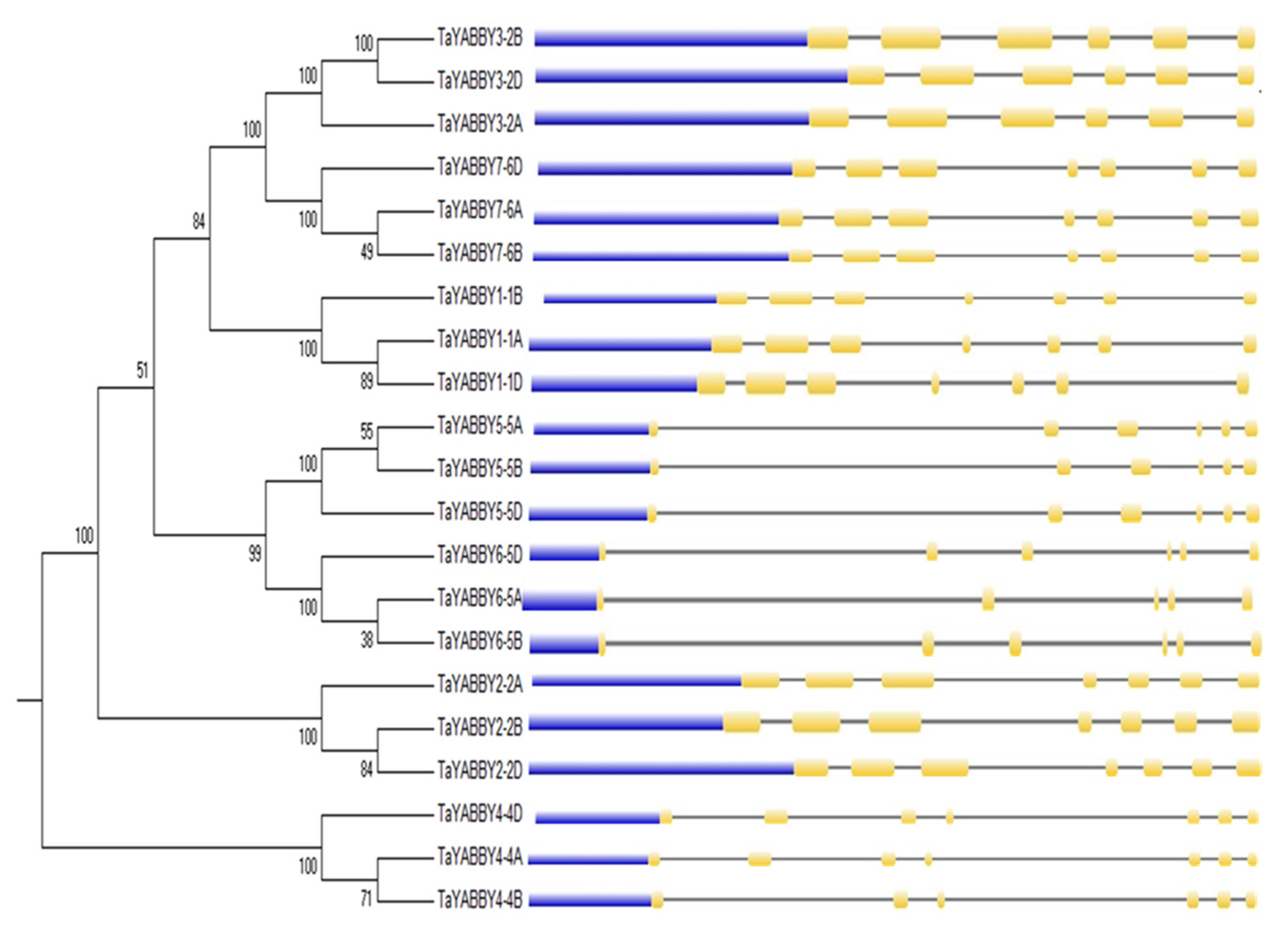
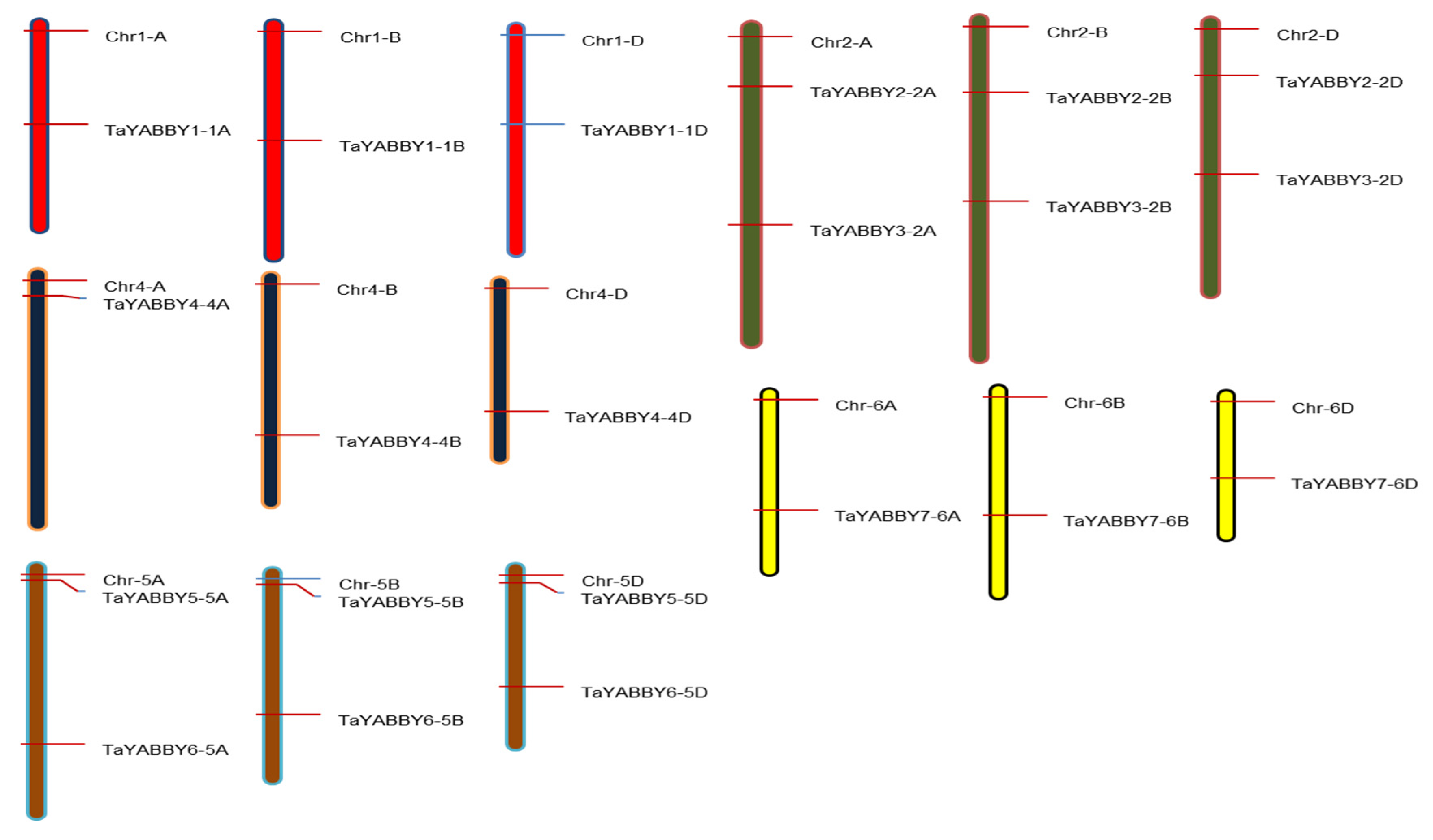
3.3. Conserved Motif Analysis, and Chromosomal Location of TaYABBY-Genes
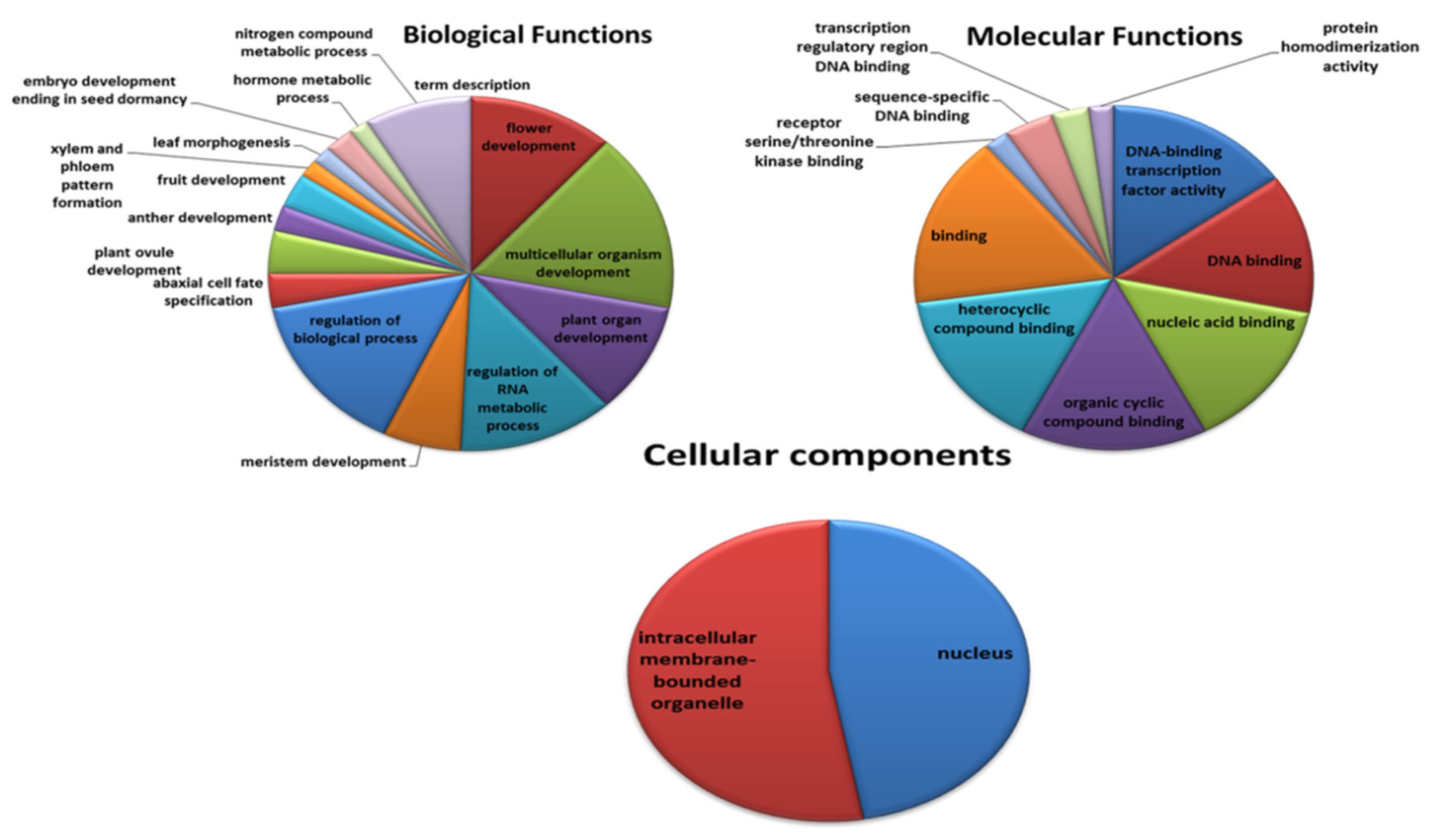
3.4. Gene Ontology of YABBY-Genes
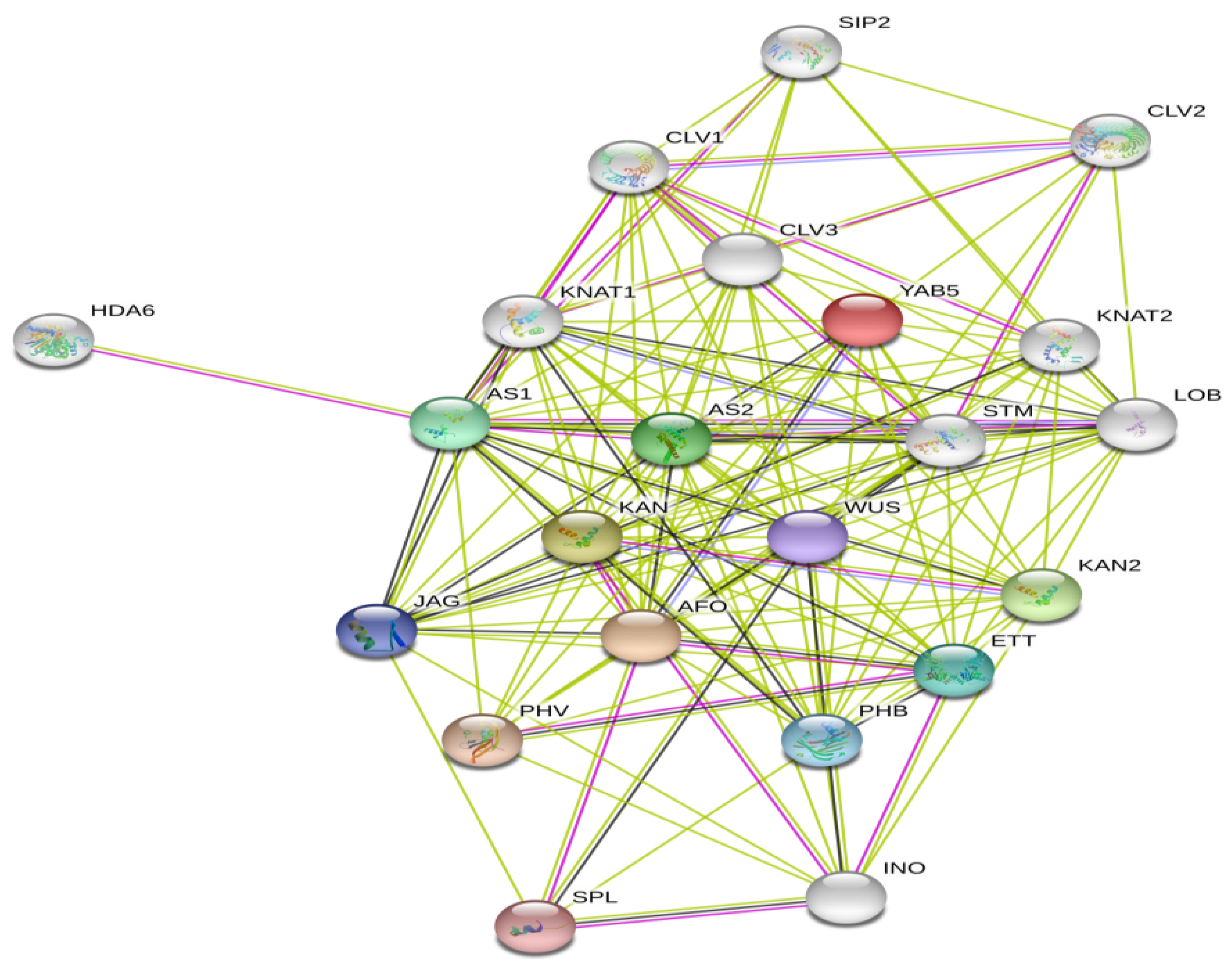
3.5. Protein-Protein Interaction of AtYABBY
3.6. Identified Cis-Regulatory Elements in TaYABBY Genes
3.7. Digital Expression Profiling of TaYABBY-Genes in Various Tissues in Normal and in Response to Stresses, at Different Tissues
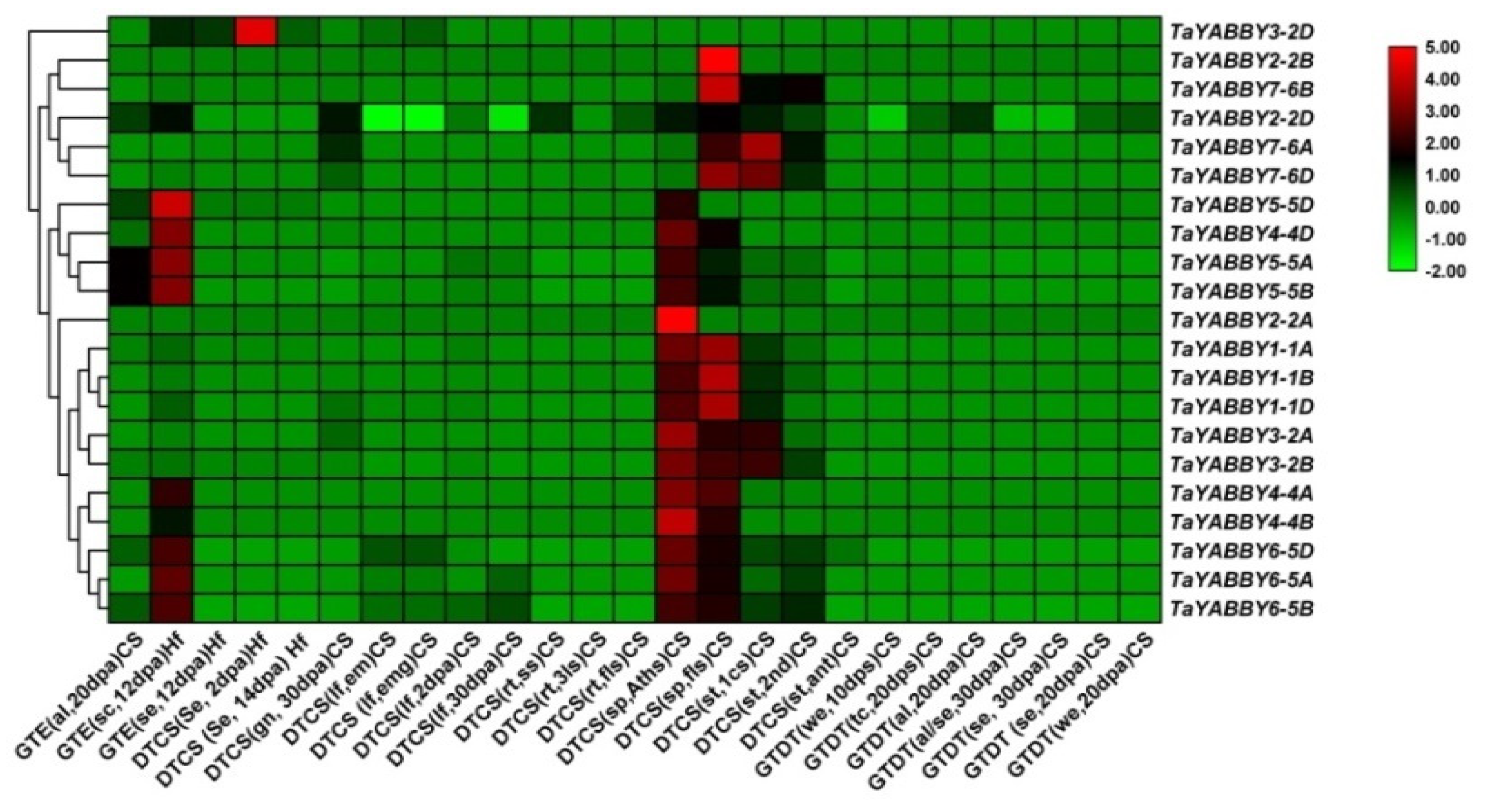
3.7.1. Reproductive and Vegetative Growth
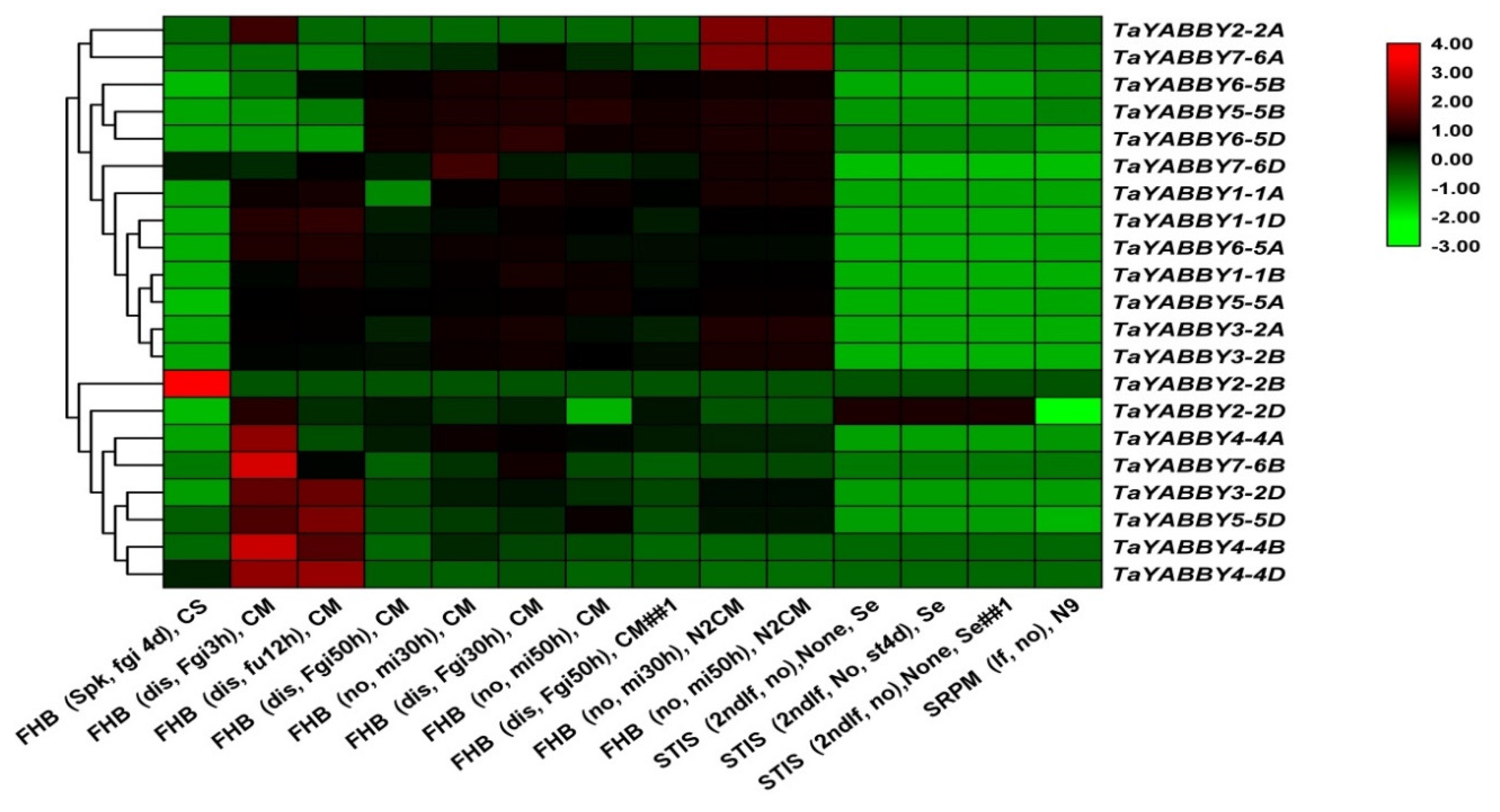
3.7.2. Biotic Stress
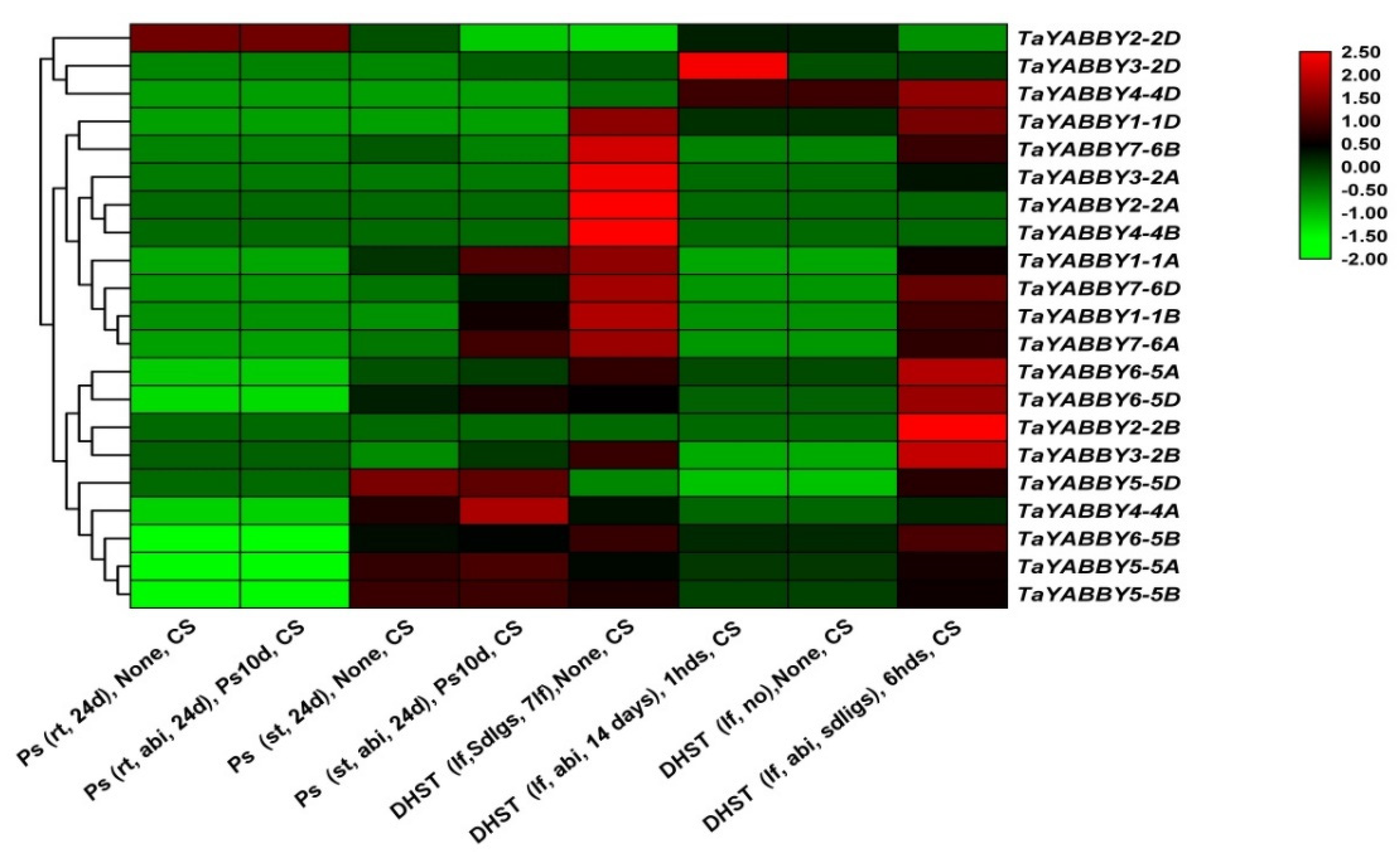
3.7.3. Abiotic Stress
4. Discussion
4.1. YABBY Genes Are Widely Distributed in Wheat
4.2. The Evolution of YABBY Proteins in Wheat
4.3. TaYABBY-Genes Digital Expression Profiling under Normal and Stresses during Developmental Processes of Triticum aestivum L.
4.3.1. TaYABBY-Genes Crucially Regulated the Wheat Growth and Developmental Activities
4.3.2. Biotic Stresses
4.3.3. Abiotic Stresses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sharif, R.; Xie, C.; Wang, J.; Cao, Z.; Zhang, H.; Chen, P.; Li, Y. Genome wide identification, characterization and expression analysis of HD-ZIP gene family in Cucumis sativus L. under biotic and various abiotic stresses. Int. J. Biol. Macromol. 2019. [Google Scholar] [CrossRef]
- Zhao, S.P.; Lu, D.; Yu, T.F.; Ji, Y.J.; Zheng, W.J.; Zhang, S.X.; Chai, S.C.; Chen, Z.Y.; Cui, X.Y. Genome-wide analysis of the YABBY family in soybean and functional identification of GmYABBY10 involvement in high salt and drought stresses. Plant Physiol. Biochem. 2017, 119, 132–146. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, G.; Cai, M.; Priyadarshani, S.V.G.N.; Aslam, M.; Zhou, Q.; Qin, Y. Genome-Wide Analysis of the YABBY Transcription Factor Family in Pineapple and Functional Identification of AcYABBY4 Involvement in Salt Stress. Int. J. Mol. Sci. 2019, 20, 5863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumaran, M.K.; Bowman, J.L.; Sundaresan, V. YABBY Polarity Genes Mediate the repression of KNOX homeobox genes in arabidopsis. Plant Cell 2002, 14, 2761–2770. [Google Scholar] [CrossRef] [PubMed]
- Bowman, J.L.; Smyth, D.R.; Meyerowitz, E.M. Genes directing flower development in Arabidopsis. Plant Cell 1989, 1, 37–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, C.M.; Jun, J.H.; Fletcher, J.C. Control of Arabidopsis leaf morphogenesis through regulation of the YABBY and KNOX families of transcription factors. Genetics 2010, 186, 197–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, S.; Hur, J.; Kim, S.J.; Han, M.J.; Kim, S.R.; An, G. Ectopic expression of OsYAB1causes extra stamens and carpels in rice. Plant Mol. Biol. 2004, 56, 133–143. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, L.; Sun, X.; Li, Y.; Yao, J.; Nocker, S.; Wang, X. Genome-wide Analysis of the YABBY Gene Gamily in Grapevine and Functional Characterization of VvYABBY4. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Bowman, J.L. The YABBY gene family and abaxial cell fate. Curr. Opin. Plant Biol. 2000, 3, 17–22. [Google Scholar] [CrossRef]
- Bowman, J.L.; Smyth, D.R. CRABS CLAW, a gene that regulates carpel and nectary development in Arabidopsis, encodes a novel protein with zinc finger and helix-loop-helix domains. Development 1999, 126, 2387–2396. [Google Scholar]
- Sawa, S.; Watanabe, K.; Goto, K.; Kanaya, E.; Morita, E.H.; Okada, K. Filamentous flower, a meristem and organ identify gene of Arabidopsis, encodes a protein with a zinc finger and HMG-related domains. Genes Dev. 1999, 13, 1079–1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.; Van Houten, J.; Gonzalez, G.; Xiao, H.; Van Der Knaap, E. Genome-wide identification, phylogeny and expression analysis of SUN, OFP and YABBY gene family in tomato. Mol. Genet. Genom. 2013, 288, 111–129. [Google Scholar] [CrossRef] [PubMed]
- Golz, J.F.; Roccaro, M.; Kuzoff, R.; Hudson, A. GRAMINIFOLIZ promotes growth and polarity of Antirrhinum leaves. Development 2004, 131, 3661–3670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juarez, M.T.; Twigg, R.W.; Timmermans, M.C.P. Specification of adaxial cell fate during maize leaf development. Development 2004, 131, 4533–4544. [Google Scholar] [CrossRef] [Green Version]
- Toriba, T.; Harada, K.; Takamura, A.; Nakamura, H.; Ichikawa, H.; Suzaki, T.; Hirano, H.Y. Molecular characterization the YABBY gene family in Oryza sativa and expression analysis of OsYABBY1. Mol. Genet. Genom. 2007, 277, 457–468. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Nagasawa, N.; Kawasaki, S.; Matsuoka, M.; Nagato, Y.; Hirano, H.Y. The YABBY Gene DROOPING LEAF Regulates Carpel Specification and Midrib Development in Oryza sativa. Plant Cell 2004, 16, 500–509. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, J.; Smyth, D.R. CRABS CLAW and SPATULA, two Arabidopsis genes that control carpel development in parallel with AGAMOUS. Development 1999, 126, 2377–2386. [Google Scholar]
- Liu, H.L.; Xu, Y.Y.; Xu, Z.H.; Chong, K. A rice YABBY gene, OsYABBY4, preferentially expresses in developing vascular tissue. Dev. Genes Evol. 2007, 217, 629–637. [Google Scholar] [CrossRef]
- Yang, C.; Ma, Y.; Li, J. The riceYABBY4gene regulates plant growth and development through modulating the gibberellin pathway. J. Exp. Bot. 2016, 67, 5545–5556. [Google Scholar] [CrossRef] [Green Version]
- Cong, B.; Barrero, L.S.; Tanksley, S.D. Regulatory change in YABBY-like transcription factor led to evolution of extreme fruit size during tomato domestication. Nat. Genet. 2008, 40, 800–804. [Google Scholar] [CrossRef]
- Huang, Z.; Knaap, E.V.D. Tomato fruit weight 11.3 maps close to fasciated on the bottom of chromosome 11. Theor. Appl. Genet. 2011, 123, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Shi, G.; Li, X.; Hu, D.; Cui, Y.; Hou, J.; Huang, F. Overexpression of a soybean YABBY gene, GmFILa, causes leaf curling in Arabidopsis thaliana. BMC Plant Biol. 2019, 19. [Google Scholar] [CrossRef] [PubMed]
- Juliana, P.; Poland, J.; Huerta-Espino, J.; Shrestha, S.; Crossa, J.; Crespo-Herrera, L.; Toledo, F.H.; Govindan, V.; Mondal, S.; Kumar, U.; et al. Improving grain yield, stress resilience and quality of bread wheat using large-scale genomics. Nat. Genet. 2019, 51, 1530–1539. [Google Scholar] [CrossRef] [PubMed]
- Lamesch, P.; Berardini, T.Z.; Li, D.; Swarbreck, D.; Wilks, C.; Sasidharan, R.; Muller, R.; Dreher, K.; Alexander, D.L.; Garcia-Hernandez, M. The Arabidopsis Information Resource (TAIR): Improved gene annotation and new tools. Nucleic Acids Res. 2011, 40, D1202–D1210. [Google Scholar] [CrossRef]
- International-Wheat-Genome-Sequencing-Consortium. A chromosome-based draft sequence of the hexaploid bread wheat (Triticum aestivum) genome. Science 2014, 345, 1251788. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.-S.; Cheng, C.-W.; Su, W.-C.; Chang, K.-C.; Huang, S.-W.; Hwang, J.-K.; Lu, C.-H. CELLO2GO: A web server for protein subCELlular LOcalization prediction with functional gene ontology annotation. PLoS ONE 2014, 9, e99368. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Yang, Y.; Luo, W.; Yang, C.; Ding, P.; Liu, Y.; Qiao, L.; Chang, Z.; Geng, H.; Wang, P.; et al. Genome-wide identification and analysis of the MADS-box gene family in bread wheat (Triticum aestivum L.). PLoS ONE 2017, 12, e0181443. [Google Scholar] [CrossRef]
- Borrill, P.; Ramirez-Gonzalez, R.; Uauy, C. expVIP: A customisable RNA-seq data analysis and visualization platform opens up gene expression analysis. Plant Physiol. 2016, 170, 2172–2186. [Google Scholar] [CrossRef] [Green Version]
- Guo, A.; Zhu, Q.; Chen, X.; Luo, J. GSDS: A gene structure display server. Hereditas 2007, 29, 1023–1026. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Bodén, M.; Buske, F.A.; Frith, M.C.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Ando, S.; Sasabe, M.; Machida, C.; Kurihara, D.; Higashiyama, T.; Machida, Y. Arabidopsis ASYMMETRIC LEAVES2 protein required for leaf morphogenesis consistently forms speckles during mitosis of tobacco BY-2 cells via signals in its specific sequence. J. Plant Res. 2012, 125, 661–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartholmes, C.; Hidalgo, O.; Gleissberg, S. Evolution of the YABBY gene family with emphasis on the basal eudicot Eschscholzia californica (Papaveraceae). Plant Biol. 2012, 14, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Siegfried, K.R.; Eshed, Y.; Baum, S.F.; Otsuga, D.; Drews, G.N.; Bowman, J.L. Members of the YABBY gene family specify abaxial cell fate in Arabidopsis. Development 1999, 126, 4117–4128. [Google Scholar]
- Buttar, Z.A.; Wu, S.N.; Arnao, M.B.; Wang, C.; Ullah, I.; Wang, C. Melatonin Suppressed the Heat Stress-Induced Damage in Wheat Seedlings by Modulating the Antioxidant Machinery. Plants 2020, 9, 809. [Google Scholar] [CrossRef]
- Zhang, X.L.; Yang, Z.P.; Zhang, J.; Zhang, L.G. Ectopic expression of BraYAB1–702, a member of YABBY gene family in Chinese cabbage, causes leaf curling, inhibition of development of shoot apical meristem and flowering stage delaying in Arabidopsis thaliana. Int. J. Mol. Sci. 2013, 14, 14872–14891. [Google Scholar] [CrossRef] [Green Version]
- Feldman, M.; Levy, A.A. Genome Evolution Due to Allopolyploidization in Wheat. Genetics 2012, 192, 763–774. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Gong, Q.; Wang, L.; Jin, Y.; Xi, J.; Li, Z.; Qin, W.; Yang, Z.; Lu, L.; Chen, Q.; et al. Genome-Wide Study of YABBY Genes in Upland Cotton and Their Expression Patterns under Different Stresses. Front. Genet. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Hou, H.; Wu, P.; Gao, L.; Zhang, C.; Hou, X. Characterization and expression profile analysis of YABBY family genes in Pak-choi (Brassica rapa ssp. chinensis) under abiotic stresses and hormone treatments. Plant Growth Regul. 2019. [Google Scholar] [CrossRef]
- Ge, M.; Lu, Y.D.; Zhang, T.F.; Li, T.; Zhang, X.L.; Zhao, H. Genome-wide identification and analysis of YABBY gene family in maize. Jiangsu J. Agric. Sci. 2014, 30, 1267. [Google Scholar]
- Sarojam, R.; Sappl, P.G.; Goldshmidt, A.; Efroni, I.; Floyd, S.K.; Eshed, Y.; Bowman, J.L. Differentiating Arabidopsis shoots from leaves by combined YABBY activities. Plant Cell 2010, 22, 2113–2130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gross, T.; Broholm, S.; Becker, A. CRABS CLAW Acts as a Bifunctional Transcription Factor in Flower Development. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villanueva, J.M.; Broadhvest, J.; Hauser, B.A.; Meister, R.J.; Schneitz, K.; Gasser, C.S. INNER NO OUTER regulates abaxial- adaxial patterning in Arabidopsis ovules. Genes Dev. 1999, 13, 3160–3169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawa, S.; Ito, T.; Shimura, Y.; Okada., K. FILAMENTOUS FLOWER controls the formation and development of Arabidopsis inflorescences and floral meristems. Plant Cell 1999, 11, 69–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Efroni, I.; Eshed, Y.; Lifschitz, E. Morphogenesis of simple and compound leaves: A critical review. Plant Cell 2010, 22, 1019–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhokane, D.; Karre, S.; Kushalappa, A.C.; McCartney, C. Integrated Metabolo-Transcriptomics Reveals Fusarium Head Blight Candidate Resistance Genes in Wheat QTL-Fhb2. PLoS ONE 2016, 11, e0155851. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Su, H.Y.; Song, J.; Zhao, X.Y.; Zhang, X.S. Ectopic expression of TaYAB1, a member of YABBY gene family in wheat, causes the partial abaxialization of the adaxial epidermises of leaves and arrests the development of shoot apical meristem in Arabidopsis. Plant Sci. 2006, 170, 364–371. [Google Scholar] [CrossRef]
- Li, Z.Y.; Xu, Z.S.; He, G.Y.; Yang, G.X.; Chen, M.; Li, L.C.; Ma, Y.Z. Overexpression of soybean GmCBL1 enhances abiotic stress tolerance and promotes hypocotyl elongation in Arabidopsis. Biochem. Biophys. Res. Commun. 2012, 427, 731–736. [Google Scholar] [CrossRef]
- Pearce, S.; Vazquez-Gross, H.; Herin, S.Y.; Hane, D.; Wang, Y.; Gu, Y.Q.; Dubcovsky, J. WheatExp: An RNA-seq expression database for polyploid wheat. BMC Plant Biol. 2015, 15. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Ding, P.; Qin, P.; Liu, Y.-X.; Xie, Q.; Chen, G.; Li, W.; Jiang, Q.; Chen, G.; Lan, X.-J.; et al. Structure and expression of the TaGW7 in bread wheat (Triticum aestivum L.). Plant Growth Regul. 2017, 82, 281–291. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Putative Gene Name | Accession ID | Subfamily | Protein Length | MW | pl | Gravy | Subcellular Location |
|---|---|---|---|---|---|---|---|
| TaYABBY1-1A | TraesCS1A02G176300.1 | YaBBY2 | 297 | 31404.47 | 8.62 | −0.349 | N |
| TaYABBY1-1B | TraesCS1B02G203800.1 | YaBBY2 | 297 | 31444.45 | 8.62 | −0.353 | N |
| TaYABBY1-1D | TraesCS1D02G162600.1 | YaBBY2 | 296 | 31280.33 | 8.62 | −0.328 | N |
| TaYABBY2-2A | TraesCS2A02G197200.1 | INO | 166 | 17846.33 | 5.62 | −0.352 | N |
| TaYABBY2-2B | TraesCS2B02G224700.1 | INO | 168 | 18102.65 | 5.64 | −0.376 | N |
| TaYABBY2-2D | TraesCS2D02G205100.1 | INO | 164 | 17758.29 | 5.92 | −0.347 | N |
| TaYABBY3-2A | TraesCS2A02G386200.1 | YaBY3/YABY1 | 262 | 28473.22 | 8.14 | −0.423 | N |
| TaYABBY3-2B | TraesCS2B02G403100.1 | YaBY3/YABY1 | 268 | 28855.63 | 6.74 | −0.302 | N |
| TaYABBY3-2D | TraesCS2D02G382700.1 | YaBY3/YABY1 | 269 | 28955.71 | 6.74 | −0.323 | N |
| TaYABBY4-4A | TraesCS4A02G058800.1 | CRC | 200 | 22304.67 | 8.98 | −0.532 | N |
| TaYABBY4-4B | TraesCS4B02G245900.1 | CRC | 200 | 22334.70 | 8.98 | −0.545 | N |
| TaYABBY4-4D | TraesCS4D02G245300.1 | CRC | 200 | 22318.70 | 8.98 | −0.531 | N |
| TaYABBY5-5A | TraesCS5A02G025900.1 | YaBBY5 | 207 | 22915.81 | 8.97 | −0.537 | N |
| TaYABBY5-5B | TraesCS5B02G025100.1 | YaBBY5 | 207 | 22663.48 | 9.13 | −0.514 | N |
| TaYABBY5-5D | TraesCS5D02G033700.1 | YaBBY5 | 204 | 22376.21 | 9.28 | −0.500 | N |
| TaYABBY6-5A | TraesCS5A02G371500.1 | YaBBY5 | 144 | 16610.89 | 9.35 | −0.724 | N |
| TaYABBY6-5B | TraesCS5B02G373600.1 | YaBBY5 | 186 | 21052.01 | 9.30 | −0.510 | N |
| TaYABBY6-5D | TraesCS5D02G380900.1 | YaBBY5 | 185 | 20968.88 | 9.30 | −0.551 | N |
| TaYABBY7-6A | TraesCS6A02G237700.1 | YaBY3/YABY1 | 250 | 26684.33 | 8.13 | −0.196 | N |
| TaYABBY7-6B | TraesCS6B02G266200.1 | YaBY3/YABY1 | 250 | 26652.34 | 8.13 | −0.167 | N |
| TaYABBY7-6D | TraesCS6D02G220400.1 | YaBY3/YABY1 | 250 | 26684.33 | 8.13 | −0.196 | N |
| Motif | E-Value | Site | Width | Sequence | Motif-Logo |
|---|---|---|---|---|---|
| 1 | 2.4e-783 | 21 | 44 | PEKRQRVPSAYNRFIKEEIQRIKANNPDITHREAFSAAAKNWAH |  |
| 2 | 1.8e-305 | 21 | 21 | EQLCYVHCNFCBTILAVSVPC |  |
| 3 | 6.4e-289 | 20 | 20 | SSLFKTVTVRCGHCANLLSVSLRGLLLPP |  |
| 4 | 7.8e-116 | 21 | 21 | FPHIHFGLMPDQGGK |  |
| 5 | 6.2e-065 | 17 | 17 | KPLMPMPSEKPAQQETEQHAR |  |
| 6 | 3.3e-062 | 12 | 12 | QGGDEDMLLKEGLYAAAAAAA |  |
| 7 | 1.3e-046 | 14 | 14 | PPAPLQPLPSLASPTSSDGSPRENSVVKP |  |
| 8 | 1.0e-042 | 9 | 9 | MSSSSSSEASFALDHLAEQQQ |  |
| 9 | 3.4e-028 | 3 | 3 | TTTTVAAESAASAVTTTTSPPPAAAAHHGQFHYPSSLNL |  |
| 10 | 1.1e-020 | 6 | 6 | EDDAAPAPKKIQGLY |  |
| Protein Name | Gene Family | Putative Function |
|---|---|---|
| KANADI (KAN) | Homeodomin-like superfamily protein | Regulates organ polarity in Arabidopsis |
| ASYMMETERIC LEAVE (AS1) | ASYMMETRIC LEAVE Proteins | Transcription factor required for normal cell differentiation. |
| ARFs ETTIN (ETT) | B3 family protein | Auxin-responsible factor AUX/IAA-related |
| PHABULOSA (PHB) | Homeobox-leucine zipper family protein | Lipid-binding START domain- containing protein |
| JAGGED (JAG) | C2H2 and C2HC zinc fingers superfamily protein | Controls the morphogenesis of lateral organs |
| WUSCHEL (WUS) | Homeodomain-like superfamily protein | Transcription factor that plays a central role during early embryogenesis. |
| SPOROCYTELESS (SPL) | Protein SPOROCYTELESS | Transcription factor that plays a central role during embryogenesis |
| PHAVOLUTA (PHV) | Homeobox-leucine zipper family protein | Lipid-binding START domain-containing protein |
| Site Name | Function | |
|---|---|---|
| Hormone | AuxRR-core | cis-acting regulatory element involved in auxin responsiveness |
| ABRE | cis-acting element involved in the abscisic acid responsiveness | |
| P-box | gibberellin-responsive element | |
| TGA-element | auxin-responsive element | |
| CGTCA-motif | cis-acting regulatory element involved in the MeJA-responsiveness | |
| GARE-motif | gibberellin-responsive element | |
| TGACG-motif | cis-acting regulatory element involved in the MeJA-responsiveness | |
| TCA-element | cis-acting element involved in salicylic acid responsiveness | |
| CCAAT-box | MYBHv1 binding site | |
| GC-motif | enhancer-like element involved in anoxic specific inducibility | |
| Stress and growth | AT1-motif | part of a light responsive module |
| AE-box | part of a module for light response | |
| A-box | cis-acting regulatory element | |
| Box 4 | part of a conserved DNA module involved in light responsiveness | |
| GCN4_motif | cis-regulatory element involved in endosperm expression | |
| I-box | part of a light responsive element | |
| MRE | MYB binding site involved in light responsiveness | |
| MSA-like | cis-acting element involved in cell cycle regulation | |
| Sp1 | light responsive element | |
| ARE | cis-acting regulatory element essential for the anaerobic induction | |
| HD-Zip 1 | element involved in differentiation of the palisade mesophyll cells | |
| MBSI | MYB binding site involved in flavonoid biosynthetic genes regulation | |
| CAAT-box | common cis-acting element in promoter and enhancer regions | |
| CAT-box | cis-acting regulatory element related to meristem expression | |
| chs-CMA2b | part of a light responsive element | |
| ATC-motif | part of a conserved DNA module involved in light responsiveness | |
| GATA-motif | part of a light responsive element | |
| ACE | cis-acting element involved in light responsiveness | |
| GA-motif | part of a light responsive element | |
| circadian | cis-acting regulatory element involved in circadian control | |
| TCCC-motif | part of a light responsive element | |
| TCT-motif | part of a light responsive element | |
| MBS | MYB binding site involved in drought-inducibility | |
| GT1-motif | light responsive element | |
| O2-site | cis-acting regulatory element involved in zein metabolism regulation |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buttar, Z.A.; Yang, Y.; Sharif, R.; Nan Wu, S.; Xie, Y.; Wang, C. Genome Wide Identification, Characterization, and Expression Analysis of YABBY-Gene Family in WHEAT (Triticum aestivum L.). Agronomy 2020, 10, 1189. https://doi.org/10.3390/agronomy10081189
Buttar ZA, Yang Y, Sharif R, Nan Wu S, Xie Y, Wang C. Genome Wide Identification, Characterization, and Expression Analysis of YABBY-Gene Family in WHEAT (Triticum aestivum L.). Agronomy. 2020; 10(8):1189. https://doi.org/10.3390/agronomy10081189
Chicago/Turabian StyleButtar, Zeeshan Ali, Yuan Yang, Rahat Sharif, Sheng Nan Wu, Yanzhou Xie, and Chengshe Wang. 2020. "Genome Wide Identification, Characterization, and Expression Analysis of YABBY-Gene Family in WHEAT (Triticum aestivum L.)" Agronomy 10, no. 8: 1189. https://doi.org/10.3390/agronomy10081189