Barley and wheat beta-glucan content influenced by weather, fertilization, and genotype
 Banafsheh Khaleghdoust1
Banafsheh Khaleghdoust1  Keyvan Esmaeilzadeh-Salestani1
Keyvan Esmaeilzadeh-Salestani1  Mailiis Korge1
Mailiis Korge1  Maarika Alaru1 Kaidi Möll1 Rando Värnik2
Maarika Alaru1 Kaidi Möll1 Rando Värnik2  Reine Koppel3 Ülle Tamm3 Max Kurg1,4
Reine Koppel3 Ülle Tamm3 Max Kurg1,4  Illimar Altosaar1
Illimar Altosaar1  Evelin Loit1*
Evelin Loit1*- 1Chair of Crop Science and Plant Biology, Institute of Agricultural and Environmental Sciences, Estonian University of Life Sciences, Tartu, Estonia
- 2Chair of Economics in Rural Economy, Institute of Agricultural and Environmental Sciences, Estonian University of Life Sciences, Tartu, Estonia
- 3Center of Estonian Rural Research and Knowledge, Jõgeva, Estonia
- 4BASF UAB, Vilnius, Lithuania
Introduction: Beta-glucan is one of the most important dietary fibers in cereal grains, having a great impact on human health and food quality. Wheat and barley are strategic crops and their fibers contribute toward their nutraceutical impact. However, the health of the environment is also important to ensure sustainable crop production. Hence the European decision makers have agreed to move toward organic cropping and to reduce the use of mineral fertilizers. Environment and agricultural management have a vast impact on the content of β-glucan in cereal grains. To exploit the valuable properties of β-glucan, the knowledge of factors increasing the amount of this component is crucial. We hypothesized that annual weather conditions, nitrogen fertilization, and cropping system affect the β-glucan amount in wheat and barley.
Methods: For this purpose, spring barley and winter wheat flour samples from an 8-year-long period (2014-2021) were characterized. The experiment included conventional and organic cropping systems with different nitrogen rates between 0 and 150 kg ha−1. In addition, the variation of β-glucan content in different varieties was analyzed.
Results and discussion: The results showed that the content of β-glucan in barley and wheat grains was affected mainly by weather conditions not by fertilization. The latter finding means that the regulation of crop nutrition is not going to impact dietary fiber content in our everyday food. Lower temperatures during tillering and higher 1,000 kernel weight and test weight showed a positive correlation with β-glucan content in barley grains, while precipitation during the grain filling period had a negative correlation. Our findings suggest that β-glucan can be obtained from low-input and organic systems as efficiently as from fertilized treatments. However, there is a need for adaptation strategies in industry, as β-glucan content can vary from year to year.
1 Introduction
Cereals are the most widely produced crops in the world, with multiple forms of use. They are considered the major source of calories and protein in the diets of both humans and livestock (Lafiandra et al., 2014). In Europe, in countries such as Germany, Poland, and the Baltic countries, the routine diet is based on cereal grains, and the use of cereal products is exclusively high (Lachman et al., 2012; Nogala-Kałucka et al., 2020). At the same time as wheat has become the most important source of food, barley on the other hand has been mostly grown for animal feed and brewing purposes. However, in recent years, the importance of barley grains as a nutraceutical ingredient has increased because of their high content of soluble fibers, especially as a rich source of β-glucan (beta-glucan; Kayal and Nirmala, 2014). But when considering that zero-waste production systems are what we are aiming for in the near future we should try to valorize cereal crops beyond their nutraceutical value. Considering barley the valorization of its hulls has been a focus of much research (Martínez-Subirà et al., 2021), but in addition the properties of cereal grain's dietary fibers have been studied extensively through the decades (Herforth et al., 2019). Cereal crops are important sources of dietary fiber. Arabinoxylans and β-glucans are primary components, accounting for ~70 and 20% of the total dietary fiber content of whole grain wheat, respectively, and 20–70% of the total dietary fiber content in barley grains, respectively (Prasadi and Joye, 2020). Paying more attention to the content of β-glucan in cereal crops could help to increase human dietary fiber consumption and alleviate the problems related to their deficiency.
Cereal β-glucans are linear homopolysaccharides of D-glucopyranosyl residues linked via β- (1–3) and β- (1–4) linkages (Lazaridou and Biliaderis, 2007), mainly found in the endosperm (75%) and aleurone layer (26%; Tiwari and Cummins, 2009; Mejía et al., 2020). Among cereals, β-glucans can be found in barley, oat, rye, and wheat (Maheshwari et al., 2017) with numeric values ranging from 2.5 to 11.3% in barley, 2.2–7.8% in oat, 1.2–2.0% in rye, and 0.4–1.4% in wheat (Lazaridou and Biliaderis, 2007). In barley, β-glucans are located throughout the starchy endosperm whereas in wheat the highest concentration is in the subaleurone layer with little in the rest of the endosperm (Izydorczyk and MacGregor, 2000; Lazaridou and Biliaderis, 2007).
Consumption of β-glucan revealed various positive effects on human health including regulation of blood glucose levels, and reduction of serum cholesterol, hypertension, and obesity (Lazaridou and Biliaderis, 2007; El Khoury et al., 2012). In the food industry, β-glucans are used as thickeners, texture enhancers, stabilizers, and fat substitutes (Mejía et al., 2020). Therefore, foods with dietary fibers such as β-glucans are considered healthier foods with lower calories and are fat-free (Elleuch et al., 2011). The importance of β-glucans for human health is supported also by the health claim authorized by the European Commission stating that barley β-glucans have been shown to lower blood cholesterol with a daily intake of 3 g of barley β-glucans (European Commission, 2012).
Content of β-glucan in grains is affected by different cropping systems (Tiwari and Cummins, 2009; Dickin et al., 2011). Nitrogen (N) availability in the soil is the main factor influencing β-glucan content in cereals (Güler, 2003b; Dvončová et al., 2010). It was reported that mineral N fertilizers increase the β-glucan content (Güler, 2003a; Noworolnik et al., 2014; Habiyaremye et al., 2021), particularly when they are fertilized with 100 kg N ha−1 (for wheat) and 80 kg N ha−1 (for barley; Güler, 2003a) regardless of the fact that β-glucan location in the grain among compared cereal grains is different (mostly in aleurone layer for wheat and endosperm for barley; Izydorczyk and MacGregor, 2000; Lazaridou and Biliaderis, 2007). On the other hand, β-glucan content of rye, wheat, barley, and oat obtained from organic farming was comparable with those from conventional farming (Menkovska et al., 2017), indicating that β-glucan content can be associated with grain size and yield (Güler, 2003b).
Weather and environmental conditions including temperature and precipitation are other factors affecting the content of β-glucans (Andersson and Börjesdotter, 2011; Dickin et al., 2011). Menkovska et al. (2017) stated that β-glucan content in oat grains depended on precipitation during the grain filling period. Drought stress decreases β-glucan concentration in barley grain (Coles et al., 1991), while Güler (2003a,b) demonstrated also a negative effect of increased levels of irrigation on barley β-glucan content. It has been found that rainy and cool weather during the period of grain establishment and flowering negatively affects β-glucan content in grain (Ehrenbergerová et al., 2008), while dry and warm growing conditions have a positive effect (Lazaridou and Biliaderis, 2007).
Genotype is another factor influencing the β-glucan content in cereal grains (Güler, 2003a; Dickin et al., 2011; Noworolnik et al., 2014; Choi et al., 2020; Habiyaremye et al., 2021). Waxy (high amylopectin; no or low amylose) barley genotypes as well as hull-less varieties have higher β-glucan content compared to genotypes with high amylose starch and hulled varieties (Ehrenbergerová et al., 2003; Izydorczyk and Dexter, 2008). Fan et al. (2009) found in the pot experiment with oat no genotype effect on grain β-glucan content, while Saastamoinen et al. (2004) stated that oat grain β-glucan depends significantly on cultivar. Tiwari and Cummins (2009) suggested that genotype is the most important factor in determining β-glucan content and found that initial levels in the grain genotype also play an important role in β-glucan levels at grain harvest. Choi et al. (2020) has found genotypic differences for barley β-glucan in interactions with year and location.
It is important to know whether β-glucan content in grains can be changed through agronomic management and varietal choice in field conditions and thereby develop action plans for plant breeding programs toward increased β-glucan content in grains. This study aimed to evaluate the effect of organic and mineral fertilizers on β-glucan content in winter wheat and spring barley grains, as well as to determine the impact of temperature and precipitation on β-glucan over 8 years. We hypothesized that the content of β-glucan in grains collected from intensively fertilized treatments is greater than in grains from organic treatments and it depends on weather conditions in specific growth stages. In addition, we characterized the content of β-glucan with respect to genotype variation.
2 Materials and methods
2.1 Experimental setup
In this study, being a part of a larger field experiment evaluating cropping systems' effect on crop production, two cereal grains, winter wheat and spring barley, were assessed based on their β-glucan content. Winter wheat “Fredis” variety and spring barley “Anni” variety grains were collected from 2014 to 2021 from the long-term field crop rotation experiment located at the Estonian University of Life Sciences in Tartu County (Eerika field). Experimental crop rotation consisting of barley (Hordeum vulgare L.) with under-sown red clover, red clover (Trifolium pratense L.), wheat (Triticum aestivum L.), pea (Pisum sativum L.), and potato (Solanum tuberosum L.) in four replications was established on Stagnic Luvisol soil. All species were cultivated organically (Org0, Org1, and Org2) and conventionally (N0, N1, N2, and N3) in parallel, as described in detail earlier by Alaru et al. (2014) and Keres et al. (2021). Organic treatments were organic control (Org0), off-season cover crops as green manure (Org1), and off-season cover crops with fully composted cattle manure (10 t ha−1 for barley + 10 t ha−1 for wheat and 20 t ha−1 for potato; Org2). Nitrogen amounts for barley and wheat in trial years were between 44 and 54 kg ha−1 depending on manure dry matter content. Conventional treatments were control N0 (N0P0K0), minerally (NH4NO3) fertilized N1 (N40 − 50P25K95), N2 (N80 − 100P25K95), and N3 (N120 − 150P25K95). Lower N rates (N40, N80, N120) were used for barley undersown with red clover, and higher rates (N50, N100, N150) for wheat.
Samples for the genotype analysis were collected in 2021 from the Estonian Crop Research Institute in Jõgeva County (50 km north of Tartu; Jõgeva field), the breeding institution of cereal grains. The soil type was Luvisol and fertilizer rates were N90P24K54 for spring barley and N151P36K75 for winter wheat. All studied spring barley varieties (Maali, Grace, Planet, Feedway, Tuuli, Breustar, Irina, Quench, Anneli, and Anni) were hulled two-row grains listed on Estonian Plant Variety Register. Most winter wheat varieties (Ada, Creator, Perenaise, Julie, Fredis, Kallas) are listed on Estonian Plant Variety Register (except for “Širvinta,” which is an Estonian heritage wheat grain).
After collection, all samples were milled [Perten Laboratory Mill 3100 (Perten Instruments AB, Sweden)]. The level of β-glucan in wheat (N = 179) and barley (N = 221) grains from the crop rotation experiment and barley (N = 10) and wheat (N = 7) varieties were determined using Mixed-linkage β-glucan assay kit (Megazyme, Ireland) based on the method published by Limberger-Bayer et al. (2014). This method is accepted by the Association of Official Analytical Chemists International (AOACI; Method 995.16) and the American Association of Cereal Chemists International (AACCI; Method 32-23). In brief, samples (100 mg) were suspended and hydrated in a sodium phosphate buffer (pH 6.5), incubated with purified lichenase enzyme, and an aliquot of filtrate was reacted with purified β-glucosidase enzymes. The glucose product was assayed using an oxidase/peroxidase reagent. The measurement of the content of β-glucan was performed in wheat and barley parallel assays, respectively, using a Helios Omega UV-Vis Spectrophotometer (Thermo Scientific) at 510 nm. The β-glucan content was calculated in dry weights and values of each sample were presented as mean ± standard deviation.
2.2 Weather conditions during the experiment
Air temperatures and precipitation during the study growing periods, which were collected from a weather station near the trial site, were fluctuating (Tables 1, 2). Tables represents the data from January to December from 2014 to 2021. The period between 2014 and 2017 was relatively colder (with an average temperature of 13.1°C) whereas the period between 2018 and 2021 was warmer (with an average temperature of 14.5°C). Precipitation was higher in the period of 2014–2017 (an average of 322 mm) when compared with the period of 2018–2021 (an average of 276 mm).
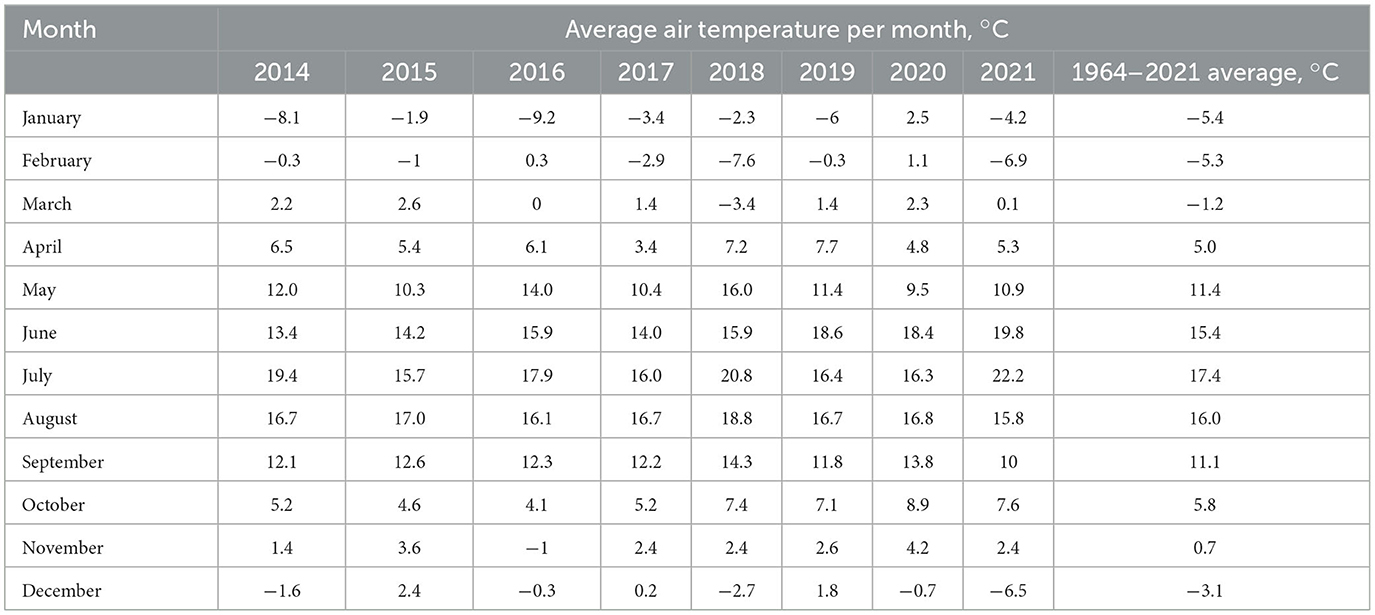
Table 1. Average air temperatures during the experimental period and their long-term averages.
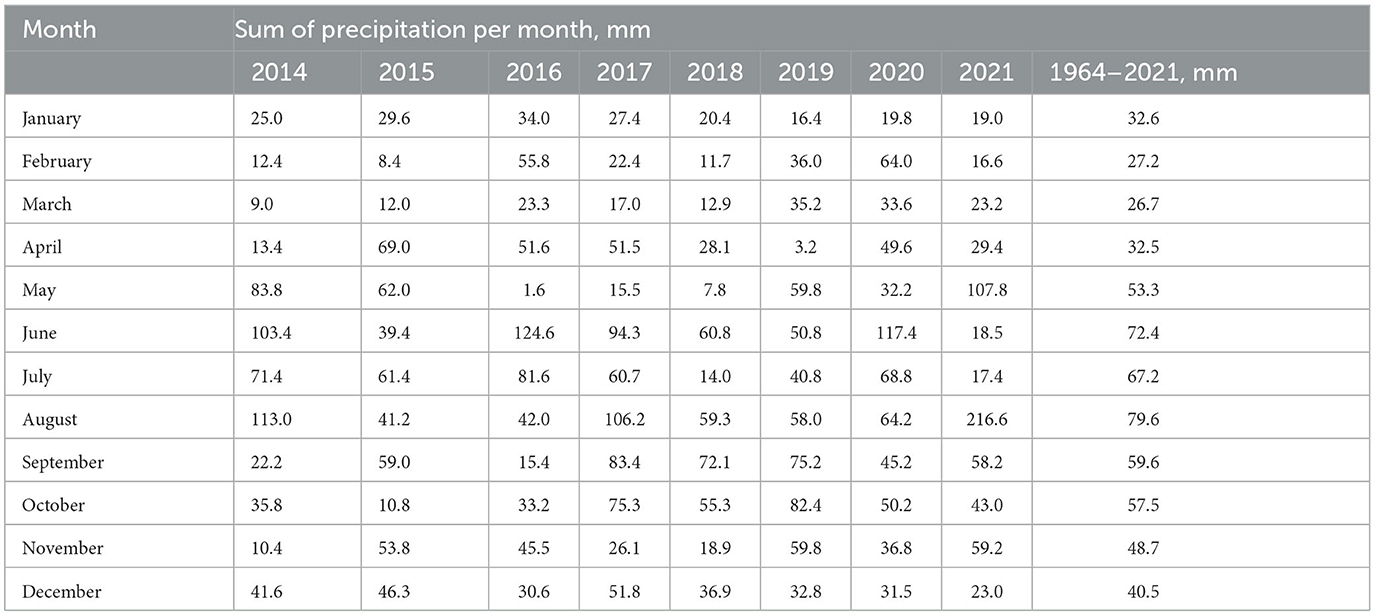
Table 2. Monthly precipitation during the experimental period and their long-term averages.
2.3 Statistical analysis
All β-glucan measurements were performed in two replicates and the results were expressed as means ± standard error. Results were analyzed using one-way and factorial analysis of variance (ANOVA), where factors were treated as fixed effects with the statistical software package Statistica version 13 (TIBCO Software Inc, USA). In case of significant (P < 0.05) differences, Fisher's least significant difference test (LSD post-hoc test) determined the differences among means. Pearson's correlation coefficient R was calculated for measuring the strength of a linear association between variables studied.
3 Results
3.1 Factors affecting the β-glucan content in barley and wheat kernels
3.1.1 Year conditions
Of the two factors investigated in the Eerika field experiment, only the year (weather) factor significantly influenced the β-glucan content of barley and wheat grains (R = 0.64, p < 0.001; R = 0.62, p < 0.001, respectively; Figures 1, 2). The proportion of variance was 90% for year and 2% for N treatment.
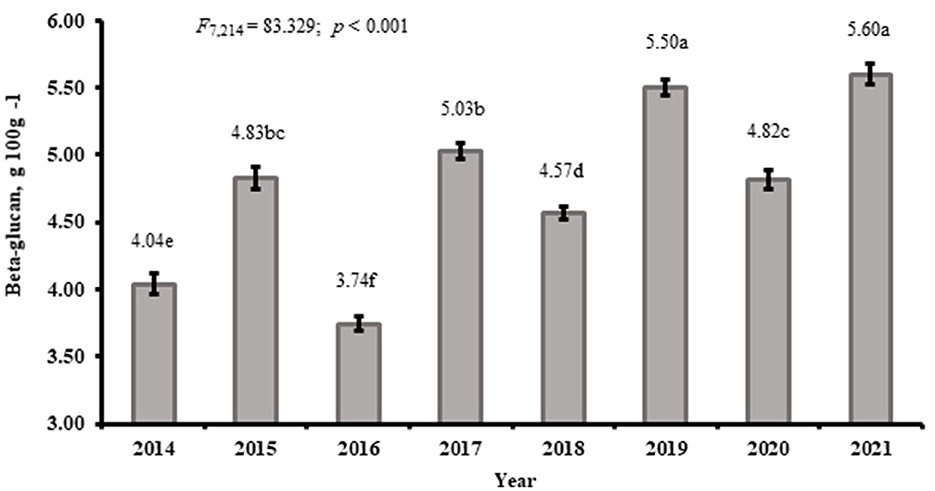
Figure 1. β-glucan content as an average of N treatments in spring barley “Anni” from Eerika field in different experimental years, g 100 g−1. Lower-case letter on the bars refer to significant differences (P < 0.05).

Figure 2. β-glucan content as an average of N treatments in winter wheat “Fredis” from Eerika field in different experimental years, g 100 g−1. Lower-case letter on the bars refer to significant differences (P < 0.05).
Mean β-glucan content of spring barley “Anni” over N treatments in different trial years ranged between 3.74 and 5.60 g 100 g−1, whereas the same data for winter wheat “Fredis” were 0.40–0.57 100 g−1, respectively. It should be taken into account that at this particular time there were two crop rotations, the first in 2014–2017 and the second in 2018–2021. In general, the second rotation was much warmer and the β-glucan content in the grains was much higher for barley (4.41 and 5.12 g 100 g−1 for 2014–2017 and 2018–2021, respectively) and also for wheat (0.49 and 0.56 g 100 g−1 for 2014–2017 and 2018–2021, respectively).
The content of β-glucan of wheat correlated negatively with the sum of precipitation in June and July (i.e., in the flowering and grain filling period of wheat; Figure 3A) and positively with air temperature in grain filling period (R = −0.23, p < 0.05; R = −0.35, p < 0.01 and R = 0.28, p < 0.05, respectively). Barley “Anni” β-glucan content correlated negatively with the sum of precipitation in June (flowering stage of barley; R = 0.67, p < 0.001) and July (grain filling period, R = 0.85, p < 0.001; Figure 3B). β-glucan content of barley correlated also negatively with air temperature in May (tillering and stem elongation stage of barley; R = 0.48, p < 0.01; Table 4).
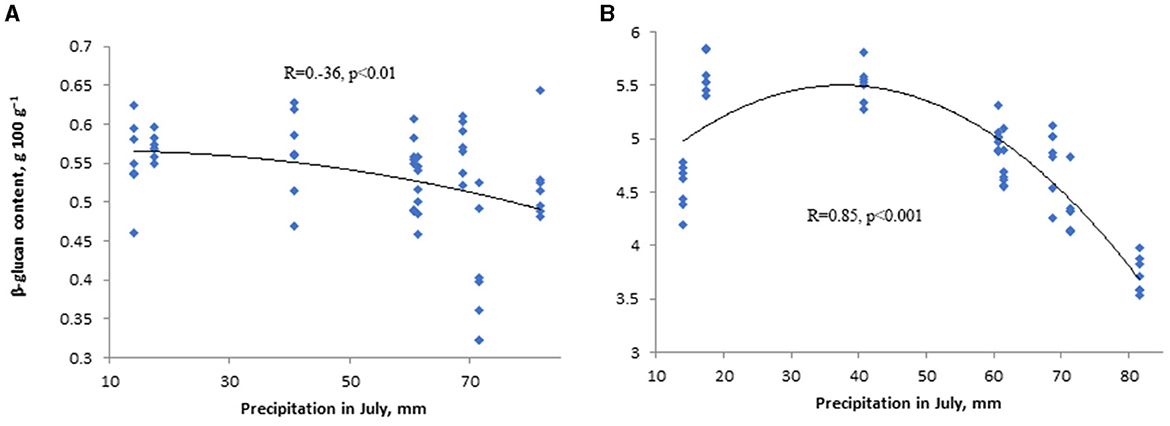
Figure 3. The influence of precipitation in grain filling period on β-glucan content of winter wheat “Fredis” (A) and spring barley “Anni” (B) of Eerika field.
3.1.2 Crop production system and N treatment
Data analysis of 8 years of results showed that the grain β-glucan content of barley and wheat was not significantly influenced by either crop production system or N treatments. Mean β-glucan content of barley grains over trial years fluctuated in different N treatments between 4.55 and 4.80 g 100 g−1 and in wheat from 0.51 to 0.56 g 100g−1.
3.1.3 Genotype
β-glucan content of barley “Anni” measured in the Eerika field experiment varied between 3.53 and 5.85 g 100 g−1. Such a large variation in the results was caused by different weather conditions and crop production systems (fertilizing with organic or mineral fertilizers) during the experimental period. Genotype of barley influenced the content of β-glucan in the Jõgeva field significantly, whereas the lowest value was obtained from “Maali” kernels and the highest from “Anni” kernels (Table 3). Differences between varieties reached up to 1.51 g 100 g−1.
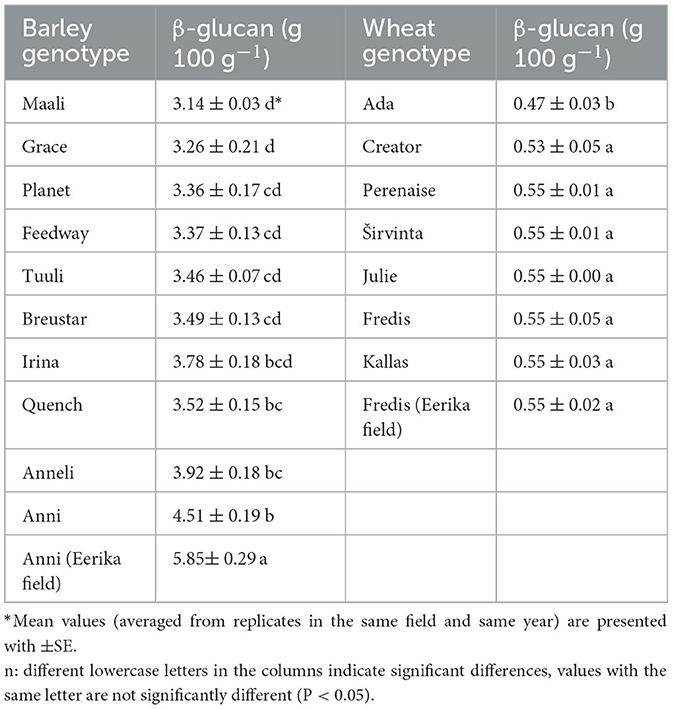
Table 3. Content of β-glucan (g 100 g−1) in spring barley and winter wheat genotypes in 2021 from Jõgeva field and Eerika field.
β-glucan content in wheat kernels measured was ~10 times lower than that of barley values. β-glucan content of wheat “Fredis” in the Eerika field fluctuated during eight trial years between 0.32 and 0.63 g 100 g−1. As with barley, genotype also affected wheat β-glucan values in the Jõgeva field experiment. Differences between wheat varieties reached up to 0.14 g 100 g−1, which was a relatively small difference, but still significant nonetheless.
The experiment showed that the effect of location on the β-glucan content of barley variety “Anni” was significant (p < 0.001; Table 3). We compared the β-glucan content of “Anni” variety obtained from the Eerika field with the data from the Jõgeva field (β-glucan values in 2021 at the same level of N quantities – 100 kg ha−1). Mean β-glucan values over repetitions were 5.85 ± 0.366 and 4.51 ± 0.192 g 100 g−1 in Eerika and Jõgeva fields, respectively. Higher 1,000 kernel weight values of barley resulted in higher β-glucan values (Figure 4).
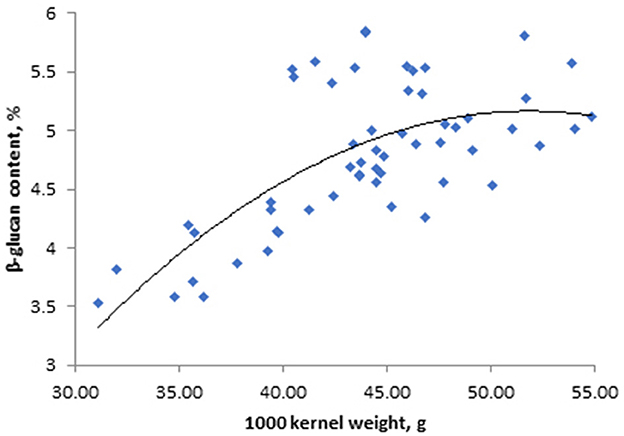
Figure 4. Relationship between 1,000 kernel weight and β-glucan content in spring barley “Anni” from Eerika field; R = 0.64, p < 0.001; n = 55.
The 1,000 kernel weight (TKW) values of barley and winter wheat cultivars were not significantly influenced by genotype. The TKW values of barley cultivars ranged between 37.6 (“Anneli”) and 43.6 (“Tuuli”) g, and for winter wheat cultivars between 37.6 (“Perenaise”) and 45.2 (“Julie”) g. Test weight of different spring barley and winter wheat cultivars also was not influenced by genotype; it ranged between 62 (“Anneli”) and 68 (“Grace”) and 75 (“Creator”) and 83 (“Ada”) kg hl−1, respectively. One thousand kernel weight values of barley “Anni” from Jõgeva (treatment N90) and Eerika (treatment N100) experimental field in 2021 were 40.3 ± 0.22 g and 44.0 ± 0.32 g, respectively. The β-glucan content of barley grains was positively correlated with grain TKW (Figure 4) and test weight values (R = 0.64, R = 0.54, respectively, p < 0.001, for both values), while the β-glucan content of winter wheat “Fredis” was not affected by either TKW or test weight (4).
4 Discussion
The quantity and quality of β-glucans are affected by fertilization, genotypes and their interaction with environment, including year and location (Güler, 2003b; Dvončová et al., 2010; Choi et al., 2020; Habiyaremye et al., 2021). Realization of β-glucan benefits requires an understanding of how those factors affect β-glucan concentration and how this variation affects the biological activity of cereals (Dickin et al., 2011). In order to optimize the quality of cereal grain, we should not only consider the traditional quality but also β-glucan concentration that may greatly benefit people facing high levels of blood-serum cholesterol. In the present experiment, β-glucan content was influenced by year (weather) conditions including air temperature and precipitation (Table 4). Previously, we showed that weather conditions had a greater impact on the content of arabinoxylan, another important polysaccharide, in barley and wheat than fertilization and cropping systems within the same experimental field (Korge et al., 2023). Our current result showed that there was a negative correlation between air temperature in May and the β-glucan content of barley grains. Conditions favorable to endosperm development would increase the accumulation of β-glucan in the grain, but β-glucan content may decrease under moist growing conditions and high temperatures in this period (Goudar et al., 2020). In Estonia's climate, barley is in the tillering/elongation stage in May, therefore, lower temperatures in May (9–10°C) extend the tillering and stem elongation phase accompanied by stronger rooting, higher number of shoots and formation of larger above ground biomass. Barley plants with higher above-ground biomass take up more nutrients to transport to the head during the grain filling period, resulting in higher 1,000 kernel weight and test weight. There was a positive correlation between barley “Anni” β-glucan and 1,000 kernel weight (Figure 4) as well as test weight (R = 0.64, p < 0.001 and R = 0.50, p < 0.01, respectively). Similarly, Yalçin et al. (2007) found a positive correlation between β-glucan content and 1,000 kernel weight in hull-less barleys grown in Turkey. They showed that dense and large grains have higher endosperm content compared to smaller grains, leading to a higher content of β-glucan. Test weight (TW), which is considered an indicator of grain density and properties, had a positive correlation with β-glucan content in barley grains in our experiment (Table 4). Since grains with a higher 1,000 kernel weight have a higher test weight, the β-glucan content of larger seeds is higher. The same correlations were not significant for wheat. This may be due to the fact that β-glucan is biosynthesized mainly in the endosperm of barley grains whereas in wheat grains, it is biosynthesized in the aleurone layer (Sapirstein, 2016).
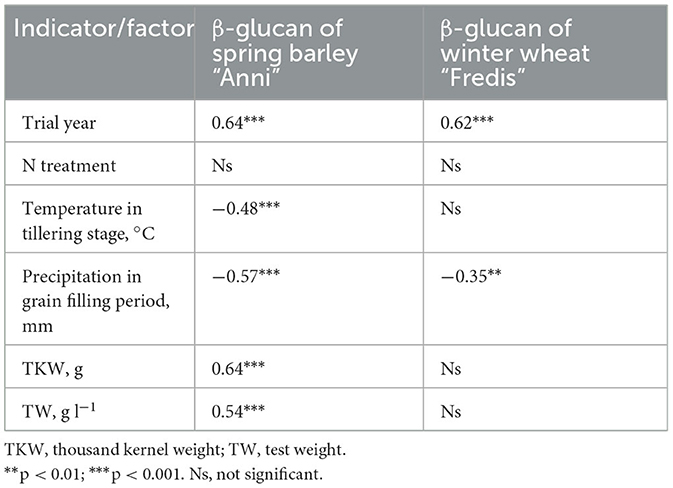
Table 4. Spring barley and winter wheat correlation coefficients (R) between β-glucan content and grains quality factors and environmental indicators (for Eerika field trial).
A negative correlation between the β-glucan content of barley and wheat grains and precipitation in June and July (Table 4) indicates that precipitation during the grain-filling period decreases the β-glucan content. While Choi et al. (2020) demonstrated a positive effect of rainfall on grain β-glucan content, current results are in accordance with Ehrenbergerová et al. (2008), who found that higher precipitation during barley flowering and grain filling decrease grain β-glucan content. Since β-glucan accumulates in the late stage of grain development, stress resulting from temperature and precipitation, which causes the end of early grain development, also reduces β-glucan concentration (Zhang et al., 2001; Tiwari and Cummins, 2009). The timing of water availability influences synthesis and deposition of main storage substances and their relative proportions in the grain (Havrlentová et al., 2023). The content of β-glucan in wheat grains was comparable in all studied years, except for 2014. High precipitation in June, July, and August (until harvesting) may be the reason for the low content of β-glucan in the year 2014. The lowest β-glucan content of barley grains was found for the year 2016, where the sum of precipitation for June and July was the highest among all studied years (2014–2021) and the highest for the year 2021, when we had the lowest precipitation in June and July compared to all other studied years. In Estonia's climate, barley is in flowering and grain-filling stages in June and July, respectively. Therefore, higher precipitation during this period can induce a dilution effect, which increases the water content in the grains and reduces the viscosity of the acid flour extract, leading to a decrease in β-glucan content (Aastrup, 1979; Güler, 2003a,b). In contrast, precipitation during the flowering time can lead to a reduction of the yield by disturbing the pollination process and decreasing the quality of grains including their β-glucan content (Aastrup, 1979).
We hypothesized that the content of β-glucan increases with higher mineral nitrogen rates compared to organic fertilizers. Previously, Güler (2003a) showed that the highest β-glucan content in their two-year experiment was observed when barley was fertilized with 80 kg N ha−1, which was the maximum N rate in their experiment. Our result showed that, in barley and wheat, fertilization up to 150 kg N ha−1 did not have a significant effect on β-glucan content throughout the 8 years (Table 4). Similarly, to our result Habiyaremye et al. (2021) did not find any effect of N fertilizers on barley β-glucan content in their three-year experiment with N rates varying from 0 to 162 kg ha−1. Although the β-glucan content of wheat grains is significantly less than barley grains, this result suggests that both crops can produce high β-glucan content without adding high mineral fertilizers into the soil. These results provide the opportunity for achieving strategies of the European Commission for increasing sustainability through organic farming.
However, the content of β-glucan can be influenced by locations and growing conditions rather than fertilization (Aastrup, 1979; Hesselman et al., 1981; Zhang et al., 2001). In our experiment, this was true for the barley variety “Anni,” in which β-glucan content was significantly different between the two locations (Table 3). In the crop rotation experiment, we had barley undersown with clover, which can affect the absorption of N fertilizer and make it easier and more accessible for the plant. Overall, undersowing legumes can be a beneficial practice for improving the absorption of N fertilizer and increasing the yield and quality of cereal grains (Lagerquist et al., 2022).
Genotype is another factor which significantly influences the content of β-glucan in grains of cereal (Choi et al., 2020). One reason can be that different genotypes have variations in sequence and transcript for genes engaged in the biosynthesis of β-glucan, which leads to a genotype-dependent production of β-glucan at grain development (Garcia-Gimenez et al., 2019). In addition, genotype directly affects the quality properties of grains such as 1,000 kernel weight (Yalçin et al., 2007). Similarly to Sinkovič et al. (2023) we found that β-glucan content is higher in barley grains compared to wheat and genotype has an impact on grain β-glucan content. This result shows that barley could have a greater role in human food as it can improve its physiological effect and provide health benefits (Izydorczyk and Dexter, 2008). In our experiment barley “Anni” had the highest content of β-glucan compared to varieties “Maali,” “Grace,” “Planet,” “Feedway,” “Tuuli,” and “Breustar.” This result indicates that the propensity of different genotypes for developing grains with higher content of β-glucan is different. Similarly, in wheat, the content of β-glucan was influenced by genotype, and variety Ada had the lowest β-glucan content compared to all other varieties. Unlike barley, we did not find a correlation between β-glucan content and 1,000 kernel weight of wheat grains in our experiment (Table 4), therefore, differences in β-glucan content may be the result of a genotype-environment interaction, which is the main factor deciding grain end use, as shown by Choi et al. (2020). Future studies should take a more detailed look on genotype and management interactions in conditions of changing climate.
5 Conclusions
The content of β-glucan in barley and wheat grains is affected mainly by year (weather) conditions in different plant growth stages, which supports our hypothesis. Lower temperatures during tillering and stem elongation and higher temperatures during grain filling period result in higher β-glucan content in barley grains. High amounts of precipitation during flowering and grain filling period reduce the content of β-glucan in grains. Future research could use this data for model development, to provide early information to industry for adoption. As there is a positive correlation between 1,000 kernel weight and β-glucan content then the conditions favoring 1,000 kernel weight increase also the β-glucan content in barley grains. This study did not support the idea that higher mineral fertilizer rates increase grain β-glucan content. The latter means that the regulation of crop nutrition is not going to impact dietary fiber content in our everyday food. These findings suggest that β-glucan can be obtained efficiently from systems managed organically or with low inputs. Genotype and growing location were other factors influencing β-glucan amount in our experiment. The content of β-glucan was 0.6–1.4 g 100 g−1 higher in “Anni” compared to other barley varieties, whereas variety “Ada” had the lowest β-glucan content in wheat. However, achieving cereal grains with higher quality including higher β-glucan content requires consideration of different factors, and different strategies may be needed for different crops and varieties.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
BK: Data curation, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing. KE-S: Writing – review & editing. MKo: Data curation, Formal analysis, Methodology, Supervision, Writing – review & editing. MA: Formal analysis, Writing – review & editing. KM: Methodology, Resources, Writing – review & editing. RV: Writing – review & editing. RK: Resources, Writing – review & editing. ÜT: Resources, Writing – review & editing. MKu: Data curation, Formal analysis, Investigation, Methodology, Writing – review & editing. IA: Supervision, Writing – review & editing. EL: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Supervision, Writing – review & editing.
Dedication
This year is the 110th Anniversary of the birth of Estonia's world-famous cereal biochemist. This cereal chemistry manuscript is dedicated to Commemorate the 110 years of Perten Instruments' Founder, Harald Perten.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by the Estonian Research Council project number RESTA28: Valorization of cereal and oil seed crops and grants PRG1260 and PRG1949. Support was also provided by ERA–NET CORE–ORGANIC Plus project FertilCrop and ERA Net Core Organic ALL–Organic.
Acknowledgments
We are thankful to Rõhu Experimental Station, who have contributed toward the experiment management.
Conflict of interest
MKu was employed by BASF UAB.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Aastrup, S. (1979). The effect of rain on β-glucan content in barley grains. Carlsberg Res. Commun. 44, 381–393.
Alaru, M., Alaru, M., Talgre, L., Eremeev, V., Tein, B., Luik, A., et al. (2014). Crop yields and supply of nitrogen compared in conventional and organic farming systems. Agric. Food Sci. 23, 317–326. doi: 10.23986/afsci.46422
Andersson, A. A. M., and Börjesdotter, D. (2011). Effects of environment and variety on content and molecular weight of β-glucan in oats. J. Cereal. Sci. 54, 122–128. doi: 10.1016/j.jcs.2011.03.003
Choi, H., Esser, A., and Murphy, K. M. (2020). Genotype × environment interaction and stability of β-glucan content in barley in the Palouse region of eastern Washington. Crop Sci. 60, 2500–2510. doi: 10.1002/csc2.20181
Coles, G. D., Jamieson, P. D., and Haslemore, R. M. (1991). Effect of moisture stress on malting quality in Triumph barley. J. Cereal. Sci. 14, 161–177. doi: 10.1016/S0733-5210(09)80137-9
Dickin, E., Steele, K., Frost, G., Edwards-Jones, G., and Wright, D. (2011). Effect of genotype, environment and agronomic management on β-glucan concentration of naked barley grain intended for health food use. J. Cereal. Sci. 54, 44–52. doi: 10.1016/j.jcs.2011.02.009
Dvončová, D., Havrlentová, M., Hlinková, A., and Hozlár, P. (2010). Effect of fertilization and variety on the β-glucan content in the grain of oats. Sci. Technol. Qual. 17, 108–116. doi: 10.15193/zntj/2010/70/108-116
Ehrenbergerová, J., Brezinová Belcredi, N., Psota, V., Hrstková, P., Cerkal, R., and Newman, C. W. (2008). Changes caused by genotype and environmental conditions in beta-glucan content of spring barley for dietetically beneficial human nutrition. Plant Foods Hum. Nutr. 63, 111–117. doi: 10.1007/s11130-008-0079-7
Ehrenbergerová, J., Vaculová, K., Psota, V., Havlová, P, and Šerhantová, V. (2003). Effects of cropping system and genotype on variability in important phytonutrients content of the barley grain for direct food use. Plant Soil Environ. 249, 443–450. doi: 10.17221/4155-PSE
El Khoury, D., Cuda, C., Luhovyy, B. L., and Anderson, G. H. (2012). Beta glucan: health benefits in obesity and metabolic syndrome. J. Nutr. Metab. 2012, 851362. doi: 10.1155/2012/851362
Elleuch, M., Bedigian, D., Roiseux, O., Besbes, S., Blecker, C., and Attia, H. (2011). Dietary fibre and fibre-rich by-products of food processing: characterisation, technological functionality and commercial applications: a review. Food Chem. 124, 411–421. doi: 10.1016/J.FoodChem.2010.06.077
European Commission (2012). Commission Regulation (EU) 1048/2012. Available online at: https://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2012:310:0038:0040:EN:PDF (accessed November 30, 2023).
Fan, M., Zhang, Z., Wang, F., Li, Z., and Hu, Y. (2009). Effect of nitrogen forms and levels on β-glucan accumulation in grains of oat (Avena sativa L.) plants. J. Plant Nutr. Soil Sci. 172, 861–866. doi: 10.1002/jpln.200800094
Garcia-Gimenez, G., Russell, J., Aubert, M. K., Fincher, G. B., Burton, R. A., Waugh, R., et al. (2019). Barley grain (1,3;1,4)-β-glucan content: effects of transcript and sequence variation in genes encoding the corresponding synthase and endohydrolase enzymes. Sci. Rep. 9, 17250. doi: 10.1038/s41598-019-53798-8
Goudar, G., Sharma, P., Janghu, S., and Longvah, T. (2020). Effect of processing on barley β-glucan content, its molecular weight and extractability. Int. J. Biol. Macromol. 162, 1204–1216. doi: 10.1016/j.ijbiomac.2020.06.208
Güler, M. (2003a). Barley grain β-glucan content as affected by nitrogen and irrigation. Field Crops Res. 84, 335–340. doi: 10.1016/S0378-4290(03)00100-X
Güler, M. (2003b). Nitrogen and irrigation effects on β-glucan content of wheat grain. Acta Agric. Scand. B Soil Plant Sci. 53, 156–160. doi: 10.1080/09064710310005527
Habiyaremye, C., Schroeder, K. L., Reganold, J. P., White, D., Packer, D., and Murphy, K. M. (2021). Effect of nitrogen and seeding rate on β-glucan, protein, and grain yield of naked food barley in no-till cropping systems in the palouse region of the pacific northwest. Front. Sustain. Food Syst. 5, 663445. doi: 10.3389/fsufs.2021.663445
Havrlentová, M., Dvoráček, V., Jurkaninová, L., and Gregusová, V. (2023). Unraveling the potential of β-D-glucans in poales: from characterization to biosynthesis and factors affecting the content. Life 13, 6. doi: 10.3390/life13061387
Herforth, A., Arimond, M., Álvarez-Sánchez, C., Coates, J., Christianson, K., and Muehlhoff, E. (2019). A global review of food-based dietary guidelines. Adv. Nutr. 10, 590–605. doi: 10.1093/advances/nmy130
Hesselman, K., Elwinger, K., Nilsson, M., and Thomke, S. (1981). The effect of β-glucanase supplementation, stage of ripeness, and storage treatment of barley in diets fed to broiler chickens, Poultry Sci. 60, 2664–2671. doi: 10.3382/ps.0602664
Izydorczyk, M. S., and Dexter, J. E. (2008). Barley β-glucans and arabinoxylans: molecular structure, physicochemical properties, and uses in food products-a Review. Food Res. Int. 41, 850–868. doi: 10.1016/j.foodres.2008.04.001
Izydorczyk, M. S., and MacGregor, A. W. (2000). Evidence of intermolecular interactions of b-glucans and arabinoxylans. Carbohydrate Polym. 41, 417–420. doi: 10.1016/S0144-8617(99)00151-4
Kayal, V. V., and Nirmala, M. J. (2014). Study on estimation, extraction and analysis of barley beta-glucan. Int. J. Sci. Res. 3, 10.
Keres, I., Alaru, M., Koppel, R., Altosaar, I., Tosens, T., and Loit, E. (2021). The combined effect of nitrogen treatment and weather conditions on wheat protein-starch interaction and dough quality. Agriculture 11, 121232. doi: 10.3390/agriculture11121232
Korge, M., Alaru, M., Keres, I., Khaleghdoust, B., Möll, K., Altosaar, I., and Loit, E. (2023). The influence of cropping system, weather conditions and genotype on arabinoxylan content in wheat and barley grains. J. Cereal Sci. 110, 103650. doi: 10.1016/j.jcs.2023.103650
Lachman, J., Hamouz, K., Orsák, M., Pivec, V., Hejtmánková, K., Pazderu, K., et al. (2012). Impact of selected factors—cultivar, storage, cooking and baking on the content of anthocyanins in coloured-flesh potatoes. Food Chem. 133, 1107–1116. doi: 10.1016/j.foodchem.2011.07.077
Lafiandra, D., Riccardi, G., and Shewry, P. R. (2014). Improving cereal grain carbohydrates for diet and health. J. Cereal. Sci. 59, 312–326. doi: 10.1016/j.jcs.2014.01.001
Lagerquist, E., Menegat, A., Dahlin, A. S., Parsons, D., Watson, C., Ståhl, P., et al. (2022). Temporal and spatial positioning of service crops in cereals affects yield and weed control. Agriculture 12, 91398. doi: 10.3390/agriculture12091398
Lazaridou, A., and Biliaderis, C. G. (2007). Molecular aspects of cereal β-glucan functionality: physical properties, technological applications and physiological effects. J. Cereal. Sci. 46, 101–118. doi: 10.1016/j.jcs.2007.05.003
Limberger-Bayer, V. M., De Francisco, A., Chan, A., Oro, T., Ogliari, P. J., and Barreto, P. L. M. (2014). Barley β-glucans extraction and partial characterization. Food Chem. 154, 84–89. doi: 10.1016/j.foodchem.2013.12.104
Maheshwari, G., Sowrirajan, S., and Joseph, B. (2017). Extraction and isolation of β-glucan from grain sources-a review. J. Food Sci. 82, 1535–1545. doi: 10.1111/1750-3841.13765
Martínez-Subirà, M., Romero, M. P., Macià, A., Puig, E., Romagosa, I., and Moralejo, M. (2021). Bioactive compounds and antioxidant capacity in pearling fractions of hulled, partially hull-less and hull-less food barley genotypes. Foods 10, 3. doi: 10.3390/foods10030565
Mejía, S. M. V., de Francisco, A., and Bohrer, B. M. (2020). A comprehensive review on cereal β-glucan: extraction, characterization, causes of degradation, and food application. Crit. Rev. Food Sci. Nutr. 60, 3693–3704. doi: 10.1080/10408398.2019.1706444
Menkovska, M., Damjanovski, D., Levkov, V., Gjorgovska, N., Knezevic, D., and Nikolova, N. (2017). Content of β–glucan in cereals grown by organic and conventional farming. Banats. J. Biotechnol. 8, 39–47. doi: 10.7904/2068-4738-viii(16)-39
Nogala-Kałucka, M., Kawka, A., Dwiecki, K., and Siger, A. (2020). Evaluation of bioactive compounds in cereals. Study of wheat, barley, oat and selected grain. Acta Sci. Pol. Technol. Aliment. 19, 405–423. doi: 10.17306/J.AFS.2020.0858
Noworolnik, K., Wirkijowska, A., and Mikos-Szymanska, M. (2014). Effect of genotype and nitrogen fertilization on grain yield and quality of spring barley intended for health food use. Bulg. J. Agri. Sci. 20, 576–580.
Prasadi, N. V. P., and Joye, I. J. (2020). Dietary fibre from whole grains and their benefits on metabolic health. Nutrients 12, 1–20. doi: 10.3390/nu12103045
Saastamoinen, M., Hietaniemi, V., Pihlava, J.-M., Eurola, M., Kontturi, M., Tuuri, H., Niskanen, M., and Kangas, A. (2004). β-Glucan contents of groats of different oat cultivars in official variety, in organic cultivation, and in nitrogen ferilization trials in Finland. Agri and Food Scie. 13, 68–79.
Sapirstein, H. D. (2016). Bioactive compounds in wheat bran. Encycl. Food Grains 2–4, 268–276. doi: 10.1016/B978-0-12-394437-5.00109-1
Sinkovič, L., Rakszegi, M., Pipan, B., and Meglič, V. (2023). Compositional traits of grains and groats of barley, oat and spelt grown at organic and conventional fields. Foods 12, 51054. doi: 10.3390/foods12051054
Tiwari, U., and Cummins, E. (2009). Simulation of the factors affecting β-glucan levels during the cultivation of oats. J. Cereal. Sci. 50, 175–183. doi: 10.1016/j.jcs.2009.04.014
Yalçin, E., Çelik, S., Akar, T., Sayim, I., and Köksel, H. (2007). Effects of genotype and environment on β-glucan and dietary fiber contents of hull-less barleys grown in Turkey. Food Chem. 101, 171–176. doi: 10.1016/j.foodchem.2006.01.010
Keywords: beta-glucan, nitrogen, dietary fiber, cropping system, organic, conventional, genotype
Citation: Khaleghdoust B, Esmaeilzadeh-Salestani K, Korge M, Alaru M, Möll K, Värnik R, Koppel R, Tamm Ü, Kurg M, Altosaar I and Loit E (2024) Barley and wheat beta-glucan content influenced by weather, fertilization, and genotype. Front. Sustain. Food Syst. 7:1326716. doi: 10.3389/fsufs.2023.1326716
Received: 23 October 2023; Accepted: 11 December 2023;
Published: 09 January 2024.
Edited by:
Matteo Balderacchi, Independent Researcher, Piacenza, ItalyReviewed by:
Lekshmi K. Edison, University of Florida, United StatesEvan Berry Karas Craine, The Land Institute, United States
Copyright © 2024 Khaleghdoust, Esmaeilzadeh-Salestani, Korge, Alaru, Möll, Värnik, Koppel, Tamm, Kurg, Altosaar and Loit. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Evelin Loit, evelin.loit@emu.ee