 Liping Fan1
Liping Fan1 Jinxing Xia1
Jinxing Xia1 Jilong Shen2Qiang Fang3,4Hui Xia3,4Meijuan Zheng1Jin-Hee Han5Eun-Taek Han5
Jilong Shen2Qiang Fang3,4Hui Xia3,4Meijuan Zheng1Jin-Hee Han5Eun-Taek Han5 Bo Wang1*
Bo Wang1* Yuanhong Xu1*
Yuanhong Xu1*- 1Department of Clinical Laboratory, the First Affiliated Hospital of Anhui Medical University, Anhui, China
- 2The Key Laboratories of Parasitology and Zoonoses Anhui and Department of Parasitology, Anhui Medical University, Anhui, China
- 3Department of Microbiology and Parasitology, Bengbu Medical College, Anhui, China
- 4Anhui Key Laboratory of Infection and Immunity, Bengbu Medical College, Bengbu, China
- 5Department of Medical Environmental Biology and Tropical Medicine, School of Medicine, Kangwon National University, Chuncheon, South Korea
Background: Plasmodium tryptophan-rich (TR) proteins have been proposed as potential vaccine candidate antigens. Among them, P. vivax tryptophan-rich antigens (PvTR-Ags), which have positionally conserved tryptophan residues in a TR domain, are highly antigenic in humans. Several of these antigens, including PvTRAg-26, have exhibited erythrocyte-binding activities.
Methods: Subclasses of IgG antibodies against PvTRAg-26 were detected by enzyme-linked immunosorbent assay in 35 P. vivax infected patients and mice immunized with the recombinant antigen to characterize its antigenicity and immunogenicity. Moreover, the antigen-specific immune responses and Th1/Th2-type cytokine patterns of splenocytes from the immunized animals were determined in vitro. The subcellular localization of PvTRAg-26 in ring-stage parasites was also detected by indirect immunofluorescence assay.
Results: The IgG1 and IgG3 levels in P. vivax-infected patients were significantly higher than those in uninfected individuals. In the PvTRAg-26-immunized mice, elevated levels of antigen-specific IgG antibodies were observed, dominated by the IgG1 subclass, and Th1-type cytokines were remarkably increased compared with Th2-type cytokines. Additionally, the subcellular location of the PvTRAg-26 protein was closely associated with the caveola-vesicle complex on the infected-erythrocyte membrane in the early ring stage of P. vivax.
Conclusions: PvTRAg-26, a P. vivax TR antigen, with high antigenicity and immunogenicity, induces Th1-cytokine response and increases production of IgG1 antibodies. This immune profiling study provided a substantial evidence that PvTRAg-26 may be a potential candidate for P. vivax vaccine development.
Introduction
Plasmodium vivax is the predominant malaria parasite epidemic in Asian and South American countries, which affects millions of people each year (1). In most cases, the parasite causes benign malaria. However, it may give rise to a severe, even fatal infection (2–4). It has been well established that malaria parasites have presented relative resistance to commonly used anti-malarial drugs. Thus the identification of novel anti-malarial drugs and the development of vaccines are urgently needed for effective control of the disease.
In a long time, development of P. vivax vaccines has been hindered by the absence of a continuous in vitro culture system and low-level parasitemia of patients (5). Therefore, the majority of P. vivax vaccine studies are focused on orthologous antigens of P. falciparum; circumsporozoite surface proteins (CSPs), thrombospondin-related adhesive protein (TRAP) of the pre-erythrocyte stage, apical membrane antigen-1 (AMA-1), Duffy-binding protein (DBP), rhoptry-associated proteins, merozoite surface proteins of the erythrocyte stage, and Pvs25 and Pvs28 from sexual stage of the parasite (6–10). Among them, only three antigens of CSPs, TRAP, and Pvs25 of P. vivax have been extensively investigated in clinical vaccine trials (11). However, the novel practical vaccine molecules of P.vivax remain undiscovered.
Plasmodium tryptophan-rich antigens (TR-Ags) have been proposed as a group of potential vaccine candidates. The TR-Ags were first identified in the murine malaria parasite of P. yoelii. Mice immunized with the recombinant TR-Ags produced highly protective immunity against P. yoelii infection (12, 13). Similarly, TR-Ags of P. falciparum could inhibit the invasion of erythrocytes by its merozoites (14). The genome of P. vivax encodes more TR-Ags than that of any other Plasmodium species. So far, fifteen TR-Ags have been found to be able to evoke significant cellular and humoral immune responses in P. vivax-exposed individuals (15). A recent study showed that TR-Ags could bind to normal human erythrocytes and the process could be inhibited by the sera of malaria patients (16). Our previous research also revealed that the conserved TR motifs exist in most PvTR-Ags which have high antigenicity in P. vivax infection, even in patients from low-endemic regions. We recently demonstrated that there are five proteins that are associated with the caveola-vesicle complex (CVC) structure, a unique structure of P. vivax-infected erythrocytes (17).
Among the five PvTR-Ags, PvTRAg-26 is an erythrocyte-binding protein (16). Although the antigenicity of PvTRAg-26 was partially tested in the previous study in P. vivax patients, the nature of the IgG subclass response to PvTRAg-26 in patients and the immunogenicity of PvTRAg-26 remain unclarified either in vitro cell experiments or in vivo animal experiments. Moreover, the membrane-associated subcellular localization needs to be investigated. In the present study, we tested the antigenicity and immunogenicity of PvTRAg-26 in the serum samples collected from symptomatic P. vivax patients as well as PvTRAg-26 immunized mice. Total IgG antibody and its subclasses were detected in the blood and the antigen-specific immune response and Th1/Th2-type cytokines of splenocytes were measured. Additionally, the subcellular localization of the PvTRAg-26 antigen on the membrane of P. vivax-infected erythrocytes was also performed.
Materials and Methods
Human Serum Samples
Serum samples of 35 malaria patients were collected in the hospitals of Bengbu and Hefei, Anhui province of China, all of them showing positive P. vivax parasite by microscopy. Simultaneously, fifteen serum samples of the individuals from malaria non-endemic areas were taken as control. The positive or negative sera were confirmed by both microscopy and nested PCR methods (18).
Expression and Purification of Recombinant PvTRAg-26
Genomic DNAs were prepared from P. vivax isolates and used as templates for PCR amplifications. PvTRAg-26 coding genes were amplified with primers of PvTRAg-26-F (5′-CCTTCACTTATAGATAAGTACGATGCT-3′) and PvTRAg-26-R (5′-TTATATTTTTGAATTCTTCCACTGAATCC-3′) and inserted into pET-28a (+)-His vector (Sango Biotech, Shanghai, China). The inserted DNA fragments were sequenced on an ABI 3730 X 1 DNA Analyzer (Applied Biosystems, Foster City, CA, USA) by Sango Biotech Co. Ltd. Purified plasmid DNAs were prepared with a TIANprep Mini Plasmid Kit (TIANGEN, Beijing, China). The recombinant protein was affinity-purified by using a Ni-Sepharose column (Sango Biotech) as described previously (17). Recombinant PvTRAg-26 was then denatured with β-mercaptoethanol in sample buffer and analyzed by 13% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), followed by immunoblotting assay with an anti-His tag antibody (Qiagen, Hilden, Germany).
Animal Immunization With Recombinant PvTRAg-26
Female BALB/c mice, 6-8 weeks old, were purchased from Vital River Laboratory Animal Technology Co, Ltd (Beijing, China). The mice were treated following the Guidelines for the Care and Use of Research Animals established by Anhui Medical University. Two groups of mice, 5 in each, were immunized subcutaneously (SC) with 50 μg of PvTRAg-26 in phosphate-buffered saline (PBS) or Freund's complete adjuvant (Sigma-Aldrich, San Francisco, CA, USA), for four times in a 3-wk interval. Boost injections were given after 3, 6, and 9 weeks of the priming with the same amount of antigen together with Freund's incomplete adjuvant (Sigma-Aldrich). The mouse sera were collected 2 weeks after the final boost and antibodies against PvTRAg-26 were measured as described previously (19).
Enzyme-Linked Immunosorbent Assay (ELISA)
To investigate the prevalence of IgG subclasses against PvTRAg-26, serum samples from 35 P. vivax-infected patients and 15 uninfected individuals were selected. The ELISA was performed following the manufacture's instructions. Briefly, 5 μg/mL of PvTRAg-26 in coating buffer (0.05 M NaHCO3, pH 9.6) was incubated in 96-well ELISA plates (Corning-Costar, Corning, NY, USA) overnight at 4°C. The plates were incubated with 5% skimmed milk in PBS/T (0.05% Tween-20) for 1 h at 37°C to block nonspecific binding sites, and then incubated with 100 μL of individual sera diluted 1:50 in PBS/T. For IgG subclasses, the plates were washed and then incubated with horseradish peroxidase (HRP)-conjugated anti-human IgG1, IgG2, IgG3, and IgG4 antibodies (ImmunoWay, Plano, TX, USA) in dilution of 1:2000 in PBS/T. Chromogenic reactions were developed and measured based on the previous description (19).
To identify and compare the levels of total and subclasses of IgG antibodies against PvTRAg-26 in immunized mice sera, we coated ELISA plates with the recombinant antigen (1.25 μg/mL). The plates were blocked and incubated with mouse sera (1:2000 dilution in PBS/T) at 37°C for 45 min. For total IgG antibody measurements, the plates were washed and then incubated with HRP-conjugated anti-mouse IgG (H + L) (Invitrogen, Waltham, MA, USA) at a 1:50,000 dilution at 37°C for 45 min, whereas for IgG subclasses the plates were incubated with HRP-conjugated anti-mouse IgG1 (Invitrogen, MA, USA), IgG2a (Invitrogen), IgG2b (Abcam, Cambridge, MA, USA), and IgG3 (Abcam) antibodies at 1:30,000, 1:1000, 1:2000, and 1:1000 dilutions, respectively. Chromogenic reactions were developed and determined as previously described (19).
Splenocyte Proliferation and Cytokine Assays
Spleens were removed from mice 2 weeks after the fourth immunization. Splenocytes obtained from PvTRAg-26 immunized or control mice were resuspended at concentrations of 5 × 106 cells/mL in complete RPMI 1640 supplemented with 10% FBS. One hundred microliters of the cell suspension, and 100 μL of PvTRAg-26 proteins were added to 96-well culture plates at final concentrations of 2.5, 5, 10, or 20 μg/mL, respectively. Concanavalin A (Con A; Sigma-Aldrich) or lipopolysaccharide (LPS; Sigma-Aldrich) at final concentrations of 5 μg/mL or 10 μg/mL were used as positive control and PBS, as negative control. After a 72 h culture (37°C and 5% CO2), 100 μL of supernatants per well was collected and stored at −20°C for cytokine assays. Viable cells were measured using a Cell Counting Kit-8 (CCK-8 or WST-8) assay following the commercial kit protocols. Cytokines of interferon (IFN)-γ, interleukin (IL)-2, IL-4, and IL-10 were examined in culture supernatants of immunized mice using BD Cytometric Bead Array (CBA) Flex Set kit (BD Biosciences, San Diego, CA, USA) according to the manufacturer's instructions. The results were obtained by flow cytometry (FACS Calibur, BD Biosciences, San Jose, CA, USA) and analyzed using Flow Cytometric Analysis Program (FCAP) array software (Soft Flow, Kedves, Hungary).
Indirect Immunofluorescence Assay (IFA)
IFA was performed with 4% paraformaldehyde-fixed parasites (17). Slides were incubated with the following primary antibodies: mouse anti-PvTRAg-26 sera (1:100) and rabbit anti-PvPHIST/CVC-8195 sera (1:100) or mouse anti-PvTRAg-26 sera (1:100) and rabbit anti-Band 3 antibody (1:200). After primary antibody reactions, the samples were then treated with secondary antibodies, Alexa Fluor 488-conjugated goat anti-mouse IgG (1:500, Invitrogen) or Alexa Fluor 568-conjugated goat anti-mouse IgG (1:500, Invitrogen), and 4, 6-diamidino-2-phenylindole (DAPI) (1:1000, Invitrogen) was used to stain the nuclei. The slides were then mounted with Prolong Gold anti-fade reagent (Invitrogen), and visualized with confocal laser-scanning microscopy (FV1000; Olympus, Tokyo, Japan) under oil immersion. Images were edited using Adobe Photoshop CS5 (Adobe Systems, San Jose, CA, USA).
Statistical Analysis
Data were analyzed using GraphPad Prism (GraphPad Software, San Diego, CA, USA). Student's t-tests were used for comparing the difference between the means in each group. P < 0.05 was considered statistically significant.
Ethical Considerations
The protocols of the study were approved by and carried out following the recommendations of the Life Ethics Committee of Anhui Medical University (No. 20160118) and the Animal Ethics Committee of Anhui Medical University (LLSC20160161). All subjects gave their written informed consents as per the Declaration of Helsinki.
Results
Expression of Recombinant PvTRAg-26
The complete PvTRAg-26 (PlasmoDB accession No. PVX_112660) protein sequence in the sal-1 strain consists of 223 amino acids (26 kDa), rich in tryptophan residues (5.8%). The entire exon-2 (450 bp, encoding the tryptophan-rich domain) of pvtrag-26 was amplified, cloned, and expressed in Escherichia coli (Figure 1A). The recombinant protein was purified under non-denaturing conditions, as shown in Figure 1B. The corresponding immunoblots were probed with an anti-His tag monoclonal antibody. Sera of P. vivax-infected patients and PvTRAg-26-immunized mice revealed a similar and specific migration pattern in PvTRAg-26 blotting (Figure 1C). Serum samples from uninfected individuals or normal mice were used as negative controls (data not shown).
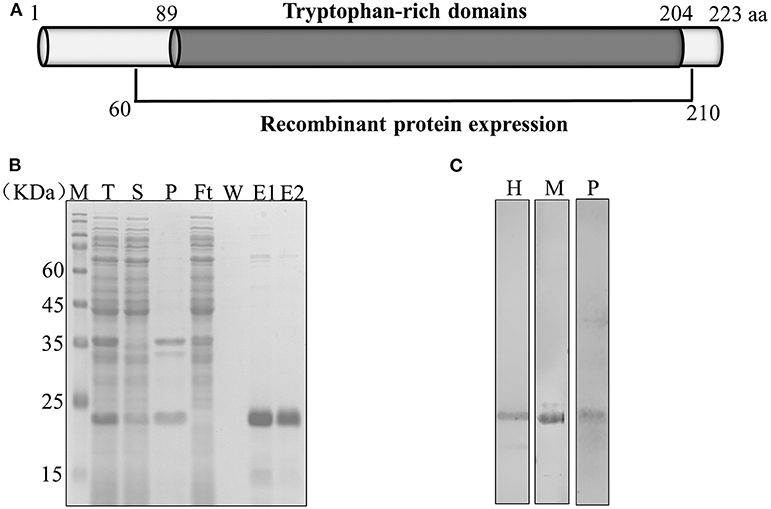
Figure 1. Schematic diagram showing the expression of PvTRAg-26. (A) Diagram of the gene structure of pvtrag-26; aa, amino acids. (B) Expression and purification of recombinant PvTRAg-26. T: total translation mix, S: supernatant, P: precipitate, Ft: flow through, W: wash; E: elution treated with reducing buffer. (C) Recombinant PvTRAg-26 protein under reducing conditions was probed with an anti-His tag antibody, P. vivax malaria patient serum and immune mouse serum. H: anti-His tag antibody, M: immune mouse serum, P: P. vivax-infected patients' pooled sera.
IgG Subclasses Recognizing PvTRAg-26 in Malaria Patients
We evaluated the prevalence of each IgG subclass antibodies against PvTRAg-26. The results showed that the mean levels and the hierarchy of IgG subclasses were as follows: IgG3 > IgG1 > IgG4 > IgG2 (Figure 2). IgG1 and IgG3 subclasses were the predominant antibodies compared to the others (P < 0.0001). Titres of the cytophilic antibodies (IgG3 and IgG1) were significantly higher than the non-cytophilic antibodies (IgG2 and IgG4) (P < 0.05). The concentrations of IgG1 and IgG3 in P. vivax patients were markedly elevated in comparison with those of uninfected controls.
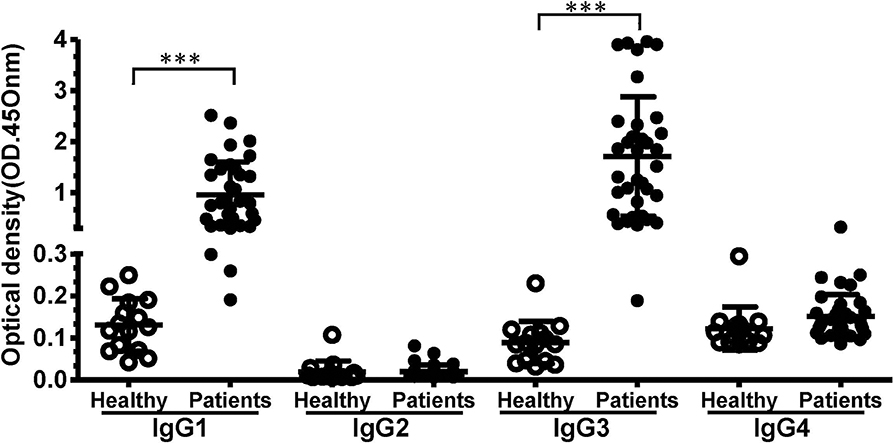
Figure 2. The levels of IgG subclasses recognizing PvTRAg-26 in sera from P. vivax-infected patients and uninfected individuals. Differences between IgG subclass levels in the negative and positive groups were analyzed using Student's t-tests. ***P < 0.001. The P values for IgG1 and IgG3 were <0.001, and those for IgG2 or IgG4 were >0.05. P < 0.05 was considered to indicate a significant difference.
Anti-PvTRAg-26 IgG and Its Subclasses in the Sera of Immunized Mice
The serum levels of antigen-specific total IgG and its subclasses in response to PvTRAg-26 were determined. The level of total PvTRAg-26 specific IgG antibodies in antigen-immunized mice was notably increased in comparison with that in the control (P < 0.001) (Figure 3A). IgG1 subclass dominated compared to IgG2b and IgG3 in the immunized mice (Figure 3B).
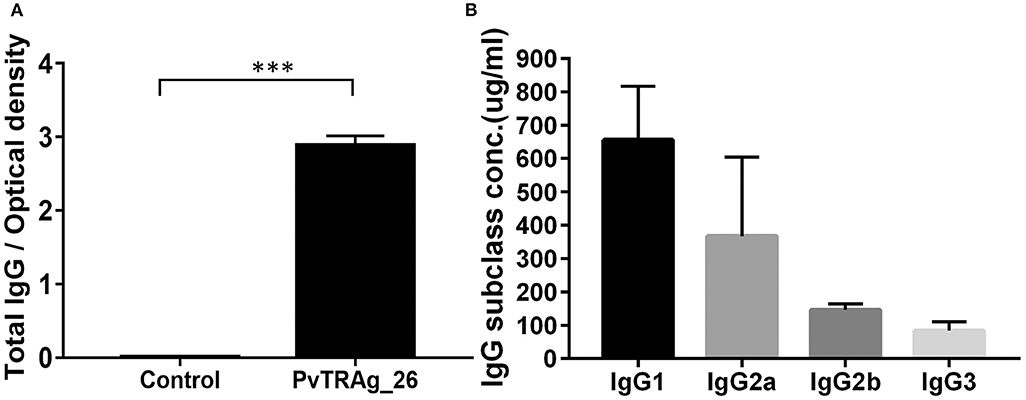
Figure 3. Levels of serum IgG and IgG subclasses in immunized mice. (A) Antigen-specific IgG levels were detected by ELISA in the sera of mice, as indicated after the final immunizations with PvTRAg-26, ***P < 0.001. (B) Serum IgG subclass pattern in PvTRAg-26-immunized mice, ***P < 0.001.
Antigen-Specific Immune Cell Responses and Cytokine Release
Splenocyte proliferation assays and CBAs were performed to assess the antigen-specific response and the secretion of cytokines in immunized BALB/c mice. Splenocytes from the animals immunized with PvTRAg-26 and controls were stimulated with various concentrations of the PvTRAg-26 antigen, Con A, or LPS for 72 h. The cultural supernatants were harvested for CBA and cell proliferation assay. The splenocyte proliferation in PvTRAg-26-immunized mice showed a notable proliferative response compared to those of the control group (P < 0.01, Figure 4A). Additionally, cytokine determinations demonstrated a biased Th1-type response, with an elevated level of IFN-γ and IL-2 secretions in the splenocytes of mice immunized with PvTRAg-26. By contrast, production of Th2-type cytokines of IL-4 and IL-10 were remarkably dampened (Figure 4B).
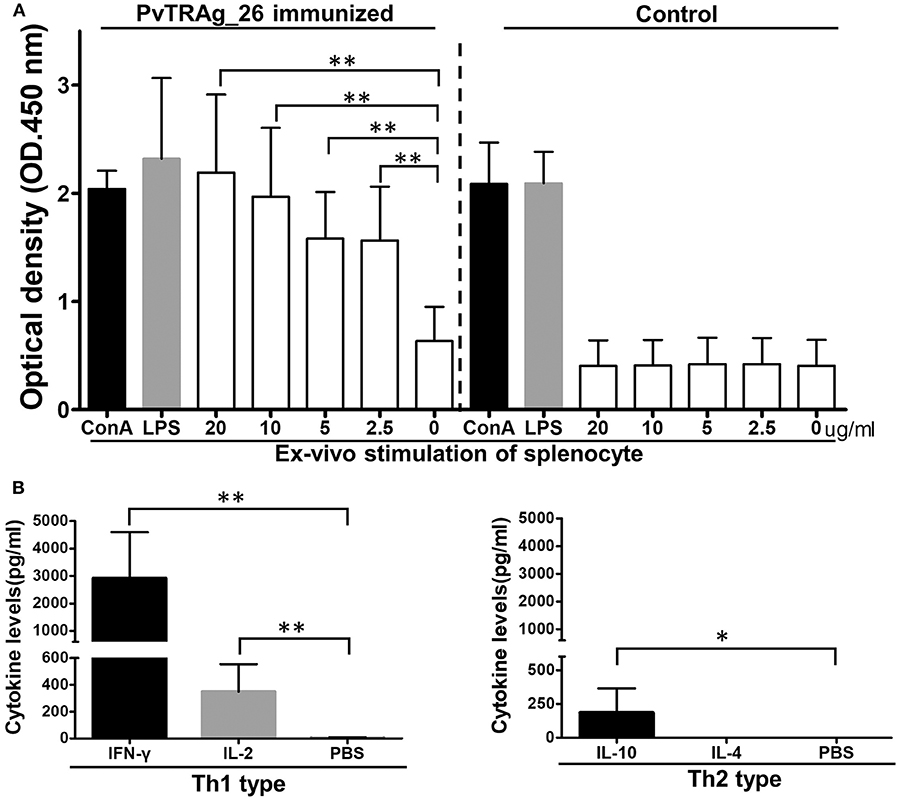
Figure 4. Proliferation index and cytokine secretion of splenocytes from mice immunized with PvTRAg-26. (A) Splenocytes from mice immunized with or without PvTRAg-26 were stimulated with various concentrations of PvTRAg-26 for 72 h before testing, as indicated. The splenocytes were stimulated with Con A or LPS as positive controls, as indicated, *P < 0.05, **P < 0.01. (B) Cytokine secretion profile of the splenocytes from the antigen-immunized mice.
Association of PvTRAg-26 With CVC and the Erythrocyte Membrane in the Early Ring Stage
Immunofluorescent assay was conducted by using anti-Band 3 (an erythrocyte membrane marker) and anti-PvPHIST/CVC-8195 (a CVC marker) sera. In the early ring or trophozoite stage of the parasite, PvTRAg-26 signals were merged (at least partially) with Band 3 and PvPHIST/CVC-8195. Specific fluorescence was visualized on the parasitophorous vacuolar membrane (PVM) (Figure 5) but the pre-immunized mouse sera did not show any signals (data not shown).
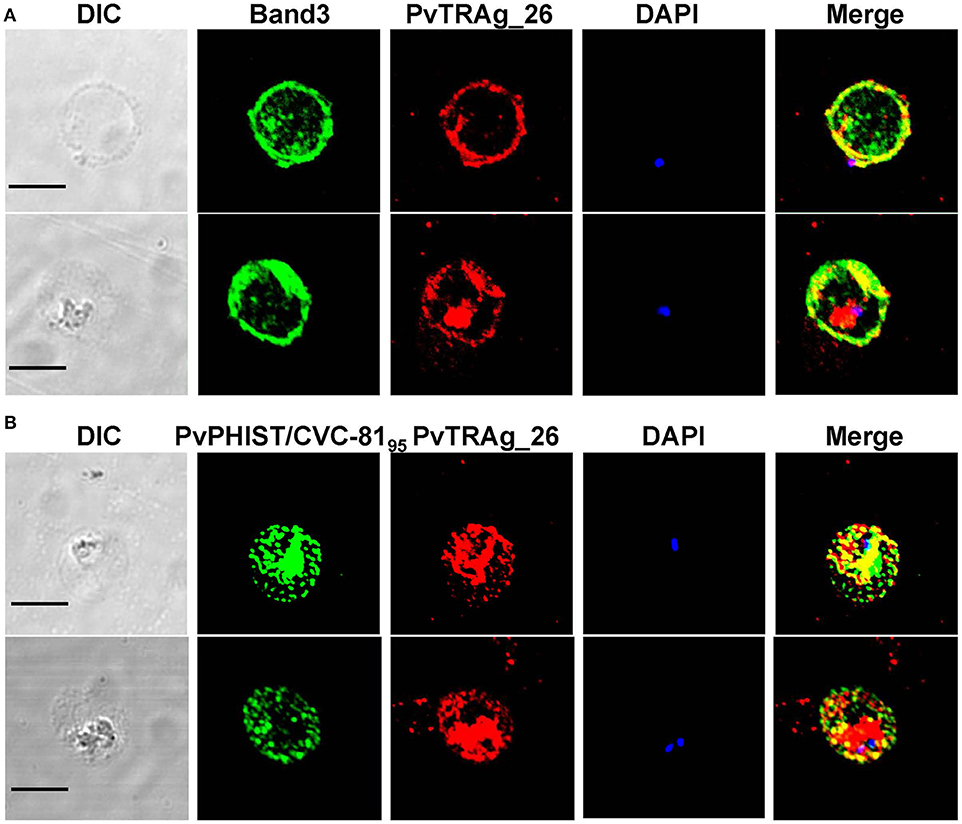
Figure 5. Localization of PvTRAg-26 in the early ring/trophozoite stage. (A) Ring stage parasites were double-labeled with mouse antisera against PvTRAg-26 (red) and a rabbit anti-Band 3 antibody (green). (B) Ring/trophozoite stage parasites were dual-labeled with mouse antisera against PvTRAg-26 (red) and rabbit antisera against PvCVC8195 (CVC marker, green). Nuclei were visualized with DAPI in merged images. The bar represents 5 μm.
Discussion
The TR-Ags of different Plasmodium species have been noted to have potential in malaria vaccine candidate screening (12, 16, 20–22) due to their parasite growth inhibition activity (23–25). P. vivax contains more abundant TR-Ags than any other human malaria parasites. Among them, PvTRAg-26, which contains positionally conserved tryptophan residues in a TR domain, could elicit a high level of protective IgG antibodies even in low malaria-endemic areas (17). Studies demonstrated that PvTRAg-26 possesses erythrocyte-binding ability (16). However, its immunogenic properties have not been fully explored. Here, we analyzed the cellular and humoral immune responses to PvTRAg-26 antigen in patients and immune mouse serum samples.
Antibodies play a crucial role in mediating acquired immunity to malaria during the intra-erythrocytic development stage of the parasites (26). The limited polymorphism of PvTR-Ags may contribute to their high immunogenicity. Our previous study exhibited that five PvTR-Ags, including PvTRAg-26, produced an elevated level of IgG antibodies in the sera of vivax patients (17). Definition of the IgG subclass response to PvTRAg-26 is important because the function of immune effectors varies in different subclasses (27, 28). Investigation of the antibody subclass response may provide further insight into the functions of antibodies and their roles in immune protection. Previous studies reported that cytophilic subclasses of IgG1 and IgG3 promote opsonic phagocytosis of merozoites or neutrophil-mediated killing, inducing a protection from malaria (29–31). Similarly, we noted that cytophilic antibody subclasses, IgG1 and IgG3, were predominant in host response to PvTRAg-26 antigen stimulation. Augmentation of humoral immune response mediated by IgG1 and IgG3 antibodies has been believed to play a pivotal role in reducing the risk of clinical malaria and parasitemia (32). During this process, complement activation mediated by IgG1 and IgG3 would be essential for inhibition of parasite invasion to host erythrocytes (33). Thus, further studies are needed to elucidate the functional activities of antibodies and their relationship with host protective immunity to vivax malaria.
Immunization with recombinant PvTRAg-26 resulted in a high level of IgG antibody response in mice, in which IgG1 subclass was predominant, followed by IgG2a. Similar results were also seen in the studies of mice with other malaria vaccine candidates, such as PvMSP119, PfMSP119, and PvMSP9 (34–36). The IgG1 and IgG3 antibodies are non-cytophilic and responsible for Th2-biased response (37), while the IgG2a and IgG2b are cytophilic and link to Th1 response in mice. Importantly, IgG2a antibodies in mice are considered to be most efficacious in complement activation and in activation of antibody-dependent cellular cytotoxic mechanism, in addition to ameliorating parasitemia caused by P. yoelii (31). We speculate that PvTRAg-26 antigen may induce a comprehensive Th1-Th2 protective response. Other studies have also shown the protective immunity in the presence of elevated levels of IgG1 and IgG2a with a combined Th1-Th2 immune responses to P. yoelii infection (38–44).
The mechanism of cellular immunity is closely associated with activation of phagocytes, antigen-specific cytotoxic T-lymphocytes and release of cytokines against infectious protozoan parasites (45). Cytokines are generally responsible for direct or indirect restriction of pathogenesis of infectious diseases (15, 46–48). It has been known that CD4+ T cells play a crucial role in protection against Plasmodium infection both in humans and in animals (49–51). The phenotype indicators of CD4+ T cells mainly include IFN-γ and IL-2 for Th1 response, and IL-4 and IL-10 for Th2 response (52, 53). Several studies demonstrated that PvTR-Ags elicit a combined Th1 and Th2 response in vivax malaria patients (51, 54). Here, we also observed a simultaneous up-regulation of cytokines of Th1 (IFN-γ and IL-2) and Th2 (IL-10) types, suggesting a systemic immune response of mice to PvTRAg-26 stimuation. The Th1 cytokines, e.g., IFN-γ, play an essential part in controlling malaria parasitemia during the early stages of infection (48, 55) and provide host with an effective protection from malaria (56, 57). IL-2 is a crucial T cell cytokine associated with proliferation, homeostasis, and differentiation of CD4+ and CD8+ T cells (58), and regulates the balance between effector Th1 cells and regulatory T cells in control of blood-stage malaria infection (59, 60). Contrarily, IL-10, a Th2 type cytokine, is known to be able to modulate the immune response to malaria parasites and to be involved in deterioration of parasitemia in Plasmodium infection (61, 62).
As visualized by IFA, PvTRAg-26 was transported from the parasite to the erythrocyte membrane through the CVC structure in the early ring stage. PvTRAg-26 was detectable on the PVM and its signal might be merged with Band 3 and CVC proteins. The function of the CVC largely remains unknown. It is hypothesized that the CVC may link to the transportation of materials from the parasite to the outside medium through the red blood cell cytoplasm (63–65). Similar to binding of PypAg-1/PypAg-3 to the membrane of red blood cells in rosette formation of P. yoelii (12), the co-localization of PvTRAg-26 with the CVC on the surface of infected erythrocytes suggests the transportation of PvTRAg-26 to the surface of the host cells, which may help promote the invasion process of P. vivax parasites. Further approaches in vivo are needed to determine the efficacy of PvTRAg-26 as a promising vaccine candidate.
Conclusions
PvTRAg-26 possesses high antigenicity and immunogenicity and can induce potent Th1 and Th2 responses in patients and immunized mice. The recombinant PvTRAg-26 antigen has the potential in development of a novel molecular vaccine for prevention of P. vivax infection.
Data Availability Statement
All datasets generated for this study are included in the article/supplementary material.
Ethics Statement
The studies involving human participants were reviewed and approved by the Ethics Committee of Anhui Medical University (20160118), Anhui, China. The patients/participants provided their written informed consent to participate in this study. The animal study was reviewed and approved by the Animal Ethics Committee, Anhui Medical University (LLSC20160161).
Author Contributions
BW, YX, and E-TH conceived and designed the experiments. LF, JX, and MZ designed the research protocol and performed the experiments. LF, JS, HX, and QF performed data acquisition and analysis. LF, JX, BW, YX, JS, J-HH and E-TH contributed to the interpretation of results and assisted in writing the manuscript. All authors read and approved the final manuscript.
Funding
This study was funded by the National Natural Science Foundation of China (No. 81601446) (BW), the Natural Science Foundation of Anhui Province (1708085QH210) (BW), the National Research Foundation of Korea (NRF) funded by the Korean government (MSIP) (NRF-2017R1A2A2A05069562) (E-TH), (2015R1A4A1038666) (E-TH), and the Foundation of the Anhui Science and Technology Department (201904a07020049) (YX). The funders had no role in the study design, data collection, and analysis, decision to publish, or preparation of the manuscript.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank Dr. Patchanee Chootong, and Mr. Obed Cudjoe and Mr. Paresh Vishwasrao for critical review of the manuscript.
Abbreviations
P. vivax, Plasmodium vivax; TRAg, tryptophan-rich antigen; CVC, caveola-vesicle complex; PCR, polymerase chain reaction; PBS, phosphate-buffered saline; SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis; ELISA, enzyme-linked immunosorbent assay; Con A, concanavalin A; LPS, lipopolysaccharide; IFN-γ, interferon-γ; IL, interleukin; IFA, indirect immunofluorescence assay; PHIST, helical interspersed subtelomeric; PVM, parasitophorous vacuolar membrane.
References
2. Kochar DK, Saxena V, Singh N, Kochar SK, Kumar SV, Das A. Plasmodium vivax malaria. Emerg Infect Dis. (2005) 11:132–4. doi: 10.3201/eid1101.040519
3. Kochar DK, Das A, Kochar SK, Saxena V, Sirohi P, Garg S, et al. Severe Plasmodium vivax malaria: a report on serial cases from Bikaner in northwestern India. Am J Trop Med Hyg. (2009) 80:194–8. doi: 10.4269/ajtmh.2009.80.194
4. Mehndiratta S, Rajeshwari K, Dubey AP. Multiple-organ dysfunction in a case of Plasmodium vivax malaria. J Vector Borne Dis. (2013) 50:71-3.
5. Pico de Coana Y, Rodriguez J, Guerrero E, Barrero C, Rodriguez R, Mendoza M, et al. A highly infective Plasmodium vivax strain adapted to Aotus monkeys: quantitative haematological and molecular determinations useful for P. vivax malaria vaccine development. Vaccine. (2003) 21:3930-7. doi: 10.1016/S0264-410X(03)00278-0
6. Zakeri S, Abouie Mehrizi A, Djadid ND, Snounou G. Circumsporozoite protein gene diversity among temperate and tropical Plasmodium vivax isolates from Iran. Trop Med Int Health. (2006) 11:729–37. doi: 10.1111/j.1365-3156.2006.01613.x
7. Bueno LL, Fujiwara RT, Soares IS, Braga EM. Direct effect of Plasmodium vivax recombinant vaccine candidates AMA-1 and MSP-119 on the innate immune response. Vaccine. (2008) 26:1204–13. doi: 10.1016/j.vaccine.2007.12.031
8. Tran TM, Oliveira-Ferreira J, Moreno A, Santos F, Yazdani SS, Chitnis CE, et al. Comparison of IgG reactivities to Plasmodium vivax merozoite invasion antigens in a Brazilian Amazon population. Am J Trop Med Hyg. (2005) 73:244–55. doi: 10.4269/ajtmh.2005.73.244
9. Wickramarachchi T, Illeperuma RJ, Perera L, Bandara S, Holm I, Longacre S, et al. Comparison of naturally acquired antibody responses against the C-terminal processing products of Plasmodium vivax Merozoite Surface Protein-1 under low transmission and unstable malaria conditions in Sri Lanka. Int J Parasitol. (2007) 37:199–208. doi: 10.1016/j.ijpara.2006.09.002
10. Xainli J, Baisor M, Kastens W, Bockarie M, Adams JH, King CL. Age-dependent cellular immune responses to Plasmodium vivax Duffy binding protein in humans. J Immunol. (2002) 169:3200–7. doi: 10.4049/jimmunol.169.6.3200
11. Mueller I, Shakri AR, Chitnis CE. Development of vaccines for Plasmodium vivax malaria. Vaccine. (2015) 33:7489–95. doi: 10.1016/j.vaccine.2015.09.060
12. Burns JM, Adeeku EK, Belk CC, Dunn PD. An unusual tryptophan-rich domain characterizes two secreted antigens of Plasmodium yoelii-infected erythrocytes. Mol Biochem Parasitol. (2000) 110:11–21. doi: 10.1016/S0166-6851(00)00252-8
13. Burns JM Jr, Dunn PD, Russo DM. Protective immunity against Plasmodium yoelii malaria induced by immunization with particulate blood-stage antigens. Infect Immun. (1997) 65:3138–45. doi: 10.1128/IAI.65.8.3138-3145.1997
14. Ntumngia FB, Bouyou-Akotet MK, Uhlemann AC, Mordmuller B, Kremsner PG, Kun JF. Characterisation of a tryptophan-rich Plasmodium falciparum antigen associated with merozoites. Mol Biochem Parasitol. (2004) 137:349–53. doi: 10.1016/j.molbiopara.2004.06.008
15. Zeeshan M, Bora H, Sharma YD. Presence of memory T cells and naturally acquired antibodies in Plasmodium vivax malaria exposed individuals against a group of tryptophan-rich antigens with conserved sequences. J Infect Dis. (2013) 207:175–85. doi: 10.1093/infdis/jis650
16. Zeeshan M, Tyagi RK, Tyagi K, Alam MS, Sharma YD. Host-parasite interaction: selective numbers of 'pv-fam-a' family proteins of Plasmodium vivax Bind to a restricted number of human erythrocyte receptors. J Infect Dis. (2015) 211:1111–20. doi: 10.1093/infdis/jiu558
17. Wang B, Lu F, Cheng Y, Chen JH, Jeon HY, Ha KS, et al. Immunoprofiling of the tryptophan-rich antigen family in Plasmodium vivax. Infect Immun. (2015) 83:3083–95. doi: 10.1128/IAI.03067-14
18. Wang B, Han SS, Cho C, Han JH, Cheng Y, Lee SK, et al. Comparison of microscopy, nested-PCR, and real-Time-PCR assays using high-throughput screening of pooled samples for diagnosis of malaria in asymptomatic carriers from areas of endemicity in Myanmar. J Clin Microbiol. (2014) 52:1838–45. doi: 10.1128/JCM.03615-13
19. Cheng Y, Shin EH, Lu F, Wang B, Choe J, Tsuboi T, et al. Antigenicity studies in humans and immunogenicity studies in mice: an MSP1P subdomain as a candidate for malaria vaccine development. Microbes Infect. (2014) 16:419–28. doi: 10.1016/j.micinf.2014.02.002
20. Curtidor H, Ocampo M, Rodriguez LE, Lopez R, Garcia JE, Valbuena J, et al. Plasmodium falciparum TryThrA antigen synthetic peptides block in vitro merozoite invasion to erythrocytes. Biochem Biophys Res Commun. (2006) 339:888–96. doi: 10.1016/j.bbrc.2005.11.089
21. Tyagi RK, Sharma YD. Erythrocyte binding activity displayed by a selective group of Plasmodium vivax tryptophan rich antigens is inhibited by patients' antibodies. PLoS ONE. (2012) 7:e50754. doi: 10.1371/journal.pone.0050754
22. Uhlemann AC, Oguariri RM, McColl DJ, Coppel RL, Kremsner PG, Anders RF, et al. Properties of the Plasmodium falciparum homologue of a protective vaccine candidate of Plasmodium yoelii. Mol Biochem Parasitol. (2001) 118:41–8. doi: 10.1016/S0166-6851(01)00370-X
23. Alam MS, Choudhary V, Zeeshan M, Tyagi RK, Rathore S, Sharma YD. Interaction of Plasmodium vivax tryptophan-rich antigen PvTRAg38 with band 3 on human erythrocyte surface facilitates parasite growth. J Biol Chem. (2015) 290:20257–72. doi: 10.1074/jbc.M115.644906
24. Rathore S, Dass S, Kandari D, Kaur I, Gupta M, Sharma YD. Basigin Interacts with Plasmodium vivax tryptophan-rich antigen PvTRAg38 as a second erythrocyte receptor to promote parasite growth. J Biol Chem. (2017) 292:462–76. doi: 10.1074/jbc.M116.744367
25. Alam MS, Zeeshan M, Mittra P, Choudhary V, Sharma YD. Receptor specific binding regions of Plasmodium vivax tryptophan rich antigens and parasite growth inhibition activity of PvTRAg35.2. Microbes Infect. (2016) 18:550–8. doi: 10.1016/j.micinf.2016.04.006
26. Cohen S, Mc GI, Carrington S. Gamma-globulin and acquired immunity to human malaria. Nature. (1961) 192:733–7. doi: 10.1038/192733a0
27. Wiener E, Atwal A, Thompson KM, Melamed MD, Gorick B, Hughes-Jones NC. Differences between the activities of human monoclonal IgG1 and IgG3 subclasses of anti-D(Rh) antibody in their ability to mediate red cell-binding to macrophages. Immunology. (1987) 62:401–4.
28. Irani V, Guy AJ, Andrew D, Beeson JG, Ramsland PA, Richards JS. Molecular properties of human IgG subclasses and their implications for designing therapeutic monoclonal antibodies against infectious diseases. Mol Immunol. (2015) 67(2 Pt A):171–82. doi: 10.1016/j.molimm.2015.03.255
29. Hill DL, Eriksson EM, Li Wai Suen CS, Chiu CY, Ryg-Cornejo V, Robinson LJ, et al. Opsonising antibodies to P. falciparum merozoites associated with immunity to clinical malaria. PLoS ONE. (2013) 8:e74627. doi: 10.1371/journal.pone.0074627
30. Osier FH, Mackinnon MJ, Crosnier C, Fegan G, Kamuyu G, Wanaguru M, et al. New antigens for a multicomponent blood-stage malaria vaccine. Sci Transl Med. (2014) 6:247ra102. doi: 10.1126/scitranslmed.3008705
31. White WI, Evans CB, Taylor DW. Antimalarial antibodies of the immunoglobulin G2a isotype modulate parasitemias in mice infected with Plasmodium yoelii. Infect Immun. (1991) 59:3547–54. doi: 10.1128/IAI.59.10.3547-3554.1991
32. Bouharoun-Tayoun H, Druilhe P. Plasmodium falciparum malaria: evidence for an isotype imbalance which may be responsible for delayed acquisition of protective immunity. Infect Immun. (1992) 60:1473–81. doi: 10.1128/IAI.60.4.1473-1481.1992
33. Boyle MJ, Reiling L, Feng G, Langer C, Osier FH, Aspeling-Jones H, et al. Human antibodies fix complement to inhibit Plasmodium falciparum invasion of erythrocytes and are associated with protection against malaria. Immunity. (2015) 42:580–90. doi: 10.1016/j.immuni.2015.02.012
34. Dutta S, Ware LA, Barbosa A, Ockenhouse CF, Lanar DE. Purification, characterization, and immunogenicity of a disulfide cross-linked Plasmodium vivax vaccine candidate antigen, merozoite surface protein 1, expressed in Escherichia coli. Infect Immun. (2001) 69:5464–70. doi: 10.1128/IAI.69.9.5464-5470.2001
35. Li G, Basagoudanavar SH, Gowda DC. Effect of GPI anchor moiety on the immunogenicity of DNA plasmids encoding the 19-kDa C-terminal portion of Plasmodium falciparum MSP-1. Parasite Immunol. (2008) 30:315–22. doi: 10.1111/j.1365-3024.2008.01027.x
36. Oliveira-Ferreira J, Vargas-Serrato E, Barnwell JW, Moreno A, Galinski MR. Immunogenicity of Plasmodium vivax merozoite surface protein-9 recombinant proteins expressed in E. coli. Vaccine. (2004) 22:2023–30. doi: 10.1016/j.vaccine.2003.07.021
37. Grey HM, Hirst JW, Cohn M. A new mouse immunoglobulin: IgG3. J Exp Med. (1971) 133:289–304. doi: 10.1084/jem.133.2.289
38. Ahlborg N, Ling IT, Holder AA, Riley EM. Linkage of exogenous T-cell epitopes to the 19-kilodalton region of Plasmodium yoelii merozoite surface protein 1 [MSP1(19)] can enhance protective immunity against malaria and modulate the immunoglobulin subclass response to MSP1(19). Infect Immun. (2000) 68:2102–9. doi: 10.1128/IAI.68.4.2102-2109.2000
39. Near KA, Stowers AW, Jankovic D, Kaslow DC. Improved immunogenicity and efficacy of the recombinant 19-kilodalton merozoite surface protein 1 by the addition of oligodeoxynucleotide and aluminum hydroxide gel in a murine malaria vaccine model. Infect Immun. (2002) 70:692–701. doi: 10.1128/IAI.70.2.692-701.2002
40. Hirunpetcharat C, Wipasa J, Sakkhachornphop S, Nitkumhan T, Zheng YZ, Pichyangkul S, et al. CpG oligodeoxynucleotide enhances immunity against blood-stage malaria infection in mice parenterally immunized with a yeast-expressed 19 kDa carboxyl-terminal fragment of Plasmodium yoelii merozoite surface protein-1 [MSP1(19)] formulated in oil-based Montanides. Vaccine. (2003) 21:2923–32. doi: 10.1016/S0264-410X(03)00132-4
41. Tamborrini M, Stoffel SA, Westerfeld N, Amacker M, Theisen M, Zurbriggen R, et al. Immunogenicity of a virosomally-formulated Plasmodium falciparum GLURP-MSP3 chimeric protein-based malaria vaccine candidate in comparison to adjuvanted formulations. Malar J. (2011) 10:359. doi: 10.1186/1475-2875-10-359
42. Dodoo D, Theisen M, Kurtzhals JA, Akanmori BD, Koram KA, Jepsen S, et al. Naturally acquired antibodies to the glutamate-rich protein are associated with protection against Plasmodium falciparum malaria. J Infect Dis. (2000) 181:1202–5. doi: 10.1086/315341
43. Oeuvray C, Theisen M, Rogier C, Trape JF, Jepsen S, Druilhe P. Cytophilic immunoglobulin responses to Plasmodium falciparum glutamate-rich protein are correlated with protection against clinical malaria in Dielmo, Senegal. Infect Immun. (2000) 68:2617–20. doi: 10.1128/IAI.68.5.2617-2620.2000
44. Hisaeda H, Saul A, Reece JJ, Kennedy MC, Long CA, Miller LH, et al. Merozoite surface protein 3 and protection against malaria in Aotus nancymai monkeys. J Infect Dis. (2002) 185:657–64. doi: 10.1086/339187
45. Suzuki Y. The immune system utilizes two distinct effector mechanisms of T cells depending on two different life cycle stages of a single pathogen, Toxoplasma gondii, to control its cerebral infection. Parasitol Int. (2019) 76:102030. doi: 10.1016/j.parint.2019.102030
46. Phillips S. Effector mechanisms against asexual erythrocytic stages of Plasmodium. Immunol Lett. (1994) 41:109–14. doi: 10.1016/0165-2478(94)90117-1
47. Su Z, Stevenson MM. Central role of endogenous gamma interferon in protective immunity against blood-stage Plasmodium chabaudi AS infection. Infect Immun. (2000) 68:4399–406. doi: 10.1128/IAI.68.8.4399-4406.2000
48. Angulo I, Fresno M. Cytokines in the pathogenesis of and protection against malaria. Clin Diagn Lab Immunol. (2002) 9:1145–52. doi: 10.1128/CDLI.9.6.1145-1152.2002
49. Fonseca L, Seixas E, Butcher G, Langhorne J. Cytokine responses of CD4+ T cells during a Plasmodium chabaudi chabaudi (ER) blood-stage infection in mice initiated by the natural route of infection. Malar J. (2007) 6:77. doi: 10.1186/1475-2875-6-77
50. Oliveira GA, Kumar KA, Calvo-Calle JM, Othoro C, Altszuler D, Nussenzweig V, et al. Class II-restricted protective immunity induced by malaria sporozoites. Infect Immun. (2008) 76:1200–6. doi: 10.1128/IAI.00566-07
51. Zeeshan M, Tyagi K, Sharma YD. CD4+ T cell response correlates with naturally acquired antibodies against Plasmodium vivax tryptophan-rich antigens. Infect Immun. (2015) 83:2018–29. doi: 10.1128/IAI.03095-14
52. Scholzen A, Mittag D, Rogerson SJ, Cooke BM, Plebanski M. Plasmodium falciparum-mediated induction of human CD25Foxp3 CD4 T cells is independent of direct TCR stimulation and requires IL-2, IL-10 and TGFbeta. PLoS Pathog. (2009) 5:e1000543. doi: 10.1371/journal.ppat.1000543
53. Torre D, Speranza F, Giola M, Matteelli A, Tambini R, Biondi G. Role of Th1 and Th2 cytokines in immune response to uncomplicated Plasmodium falciparum malaria. Clin Diagn Lab Immunol. (2002) 9:348–51. doi: 10.1128/CDLI.9.2.348-351.2002
54. Mittra P, Singh N, Sharma YD. Plasmodium vivax: immunological properties of tryptophan-rich antigens PvTRAg 35.2 and PvTRAg 80.6. Microbes Infect. (2010) 12:1019–26. doi: 10.1016/j.micinf.2010.07.004
55. Langhorne J, Ndungu FM, Sponaas AM, Marsh K. Immunity to malaria: more questions than answers. Nat Immunol. (2008) 9:725–32. doi: 10.1038/ni.f.205
56. Patterson PS, Bosshardt SC, Udhayukumar V, Xiao L, Kidd M, Hunter RL, et al. Prolonged expression of IFNgamma induced by protective blood-stage immunization against Plasmodium yoelii malaria. Vaccine. (1999) 18:173–80. doi: 10.1016/S0264-410X(99)00217-0
57. Coccia M, Collignon C, Herve C, Chalon A, Welsby I, Detienne S, et al. Cellular and molecular synergy in AS01-adjuvanted vaccines results in an early IFN gamma response promoting vaccine immunogenicity. Npj Vaccines. (2017) 2. doi: 10.1038/s41541-017-0027-3
58. Smith KA. Interleukin-2: inception, impact, and implications. Science. (1988) 240:1169–76. doi: 10.1126/science.3131876
59. Finney OC, Riley EM, Walther M. Regulatory T cells in malaria–friend or foe? Trends Immunol. (2010) 31:63–70. doi: 10.1016/j.it.2009.12.002
60. Scholzen A, Minigo G, Plebanski M. Heroes or villains? T regulatory cells in malaria infection. Trends Parasitol. (2010) 26:16–25. doi: 10.1016/j.pt.2009.10.004
61. Niikura M, Inoue S, Kobayashi F. Role of interleukin-10 in malaria: focusing on coinfection with lethal and nonlethal murine malaria parasites. J Biomed Biotech. (2011) 2011:383962. doi: 10.1155/2011/383962
62. Nakamae S, Kimura D, Miyakoda M, Sukhbaatar O, Inoue SI, Yui K. Role of IL-10 in inhibiting protective immune responses against infection with heterologous Plasmodium parasites. Parasitol Int. (2019) 70:5–15. doi: 10.1016/j.parint.2019.01.003
63. Aikawa M, Miller LH, Rabbege J. Caveola–vesicle complexes in the plasmalemma of erythrocytes infected by Plasmodium vivax and P cynomolgi. Unique structures related to Schuffner's dots. Am J Pathol. (1975) 79:285–300.
64. Aikawa M. Morphological changes in erythrocytes induced by malarial parasites. Biol Cell. (1988) 64:173–81. doi: 10.1016/0248-4900(88)90077-9
Keywords: malaria, Plasmodium vivax, tryptophan-rich antigens, immunogenicity, vaccine candidate
Citation: Fan L, Xia J, Shen J, Fang Q, Xia H, Zheng M, Han J-H, Han E-T, Wang B and Xu Y (2020) An Erythrocyte Membrane-Associated Antigen, PvTRAg-26 of Plasmodium vivax: A Study of Its Antigenicity and Immunogenicity. Front. Public Health 8:148. doi: 10.3389/fpubh.2020.00148
Received: 20 December 2019; Accepted: 09 April 2020;
Published: 28 April 2020.
Edited by:
Sunil Dhiman, Defence Research and Development Establishment (DRDE), IndiaReviewed by:
Takafira Mduluza, University of Zimbabwe, ZimbabweTarek Adnan Ahmad, Bibliotheca Alexandrina, Egypt
Copyright © 2020 Fan, Xia, Shen, Fang, Xia, Zheng, Han, Han, Wang and Xu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bo Wang, wangbo@ahmu.edu.cn; Yuanhong Xu, xyhong1964@163.com