 Guoxing Xue1
Guoxing Xue1 Yue Fan2Chunyu Zheng2Hao Yang3Liang Feng4Xingyu Chen1Yanqi Yang1Xin Yao1
Yue Fan2Chunyu Zheng2Hao Yang3Liang Feng4Xingyu Chen1Yanqi Yang1Xin Yao1 Wenfeng Weng1Lingyan Kong5Chuang Liu6
Wenfeng Weng1Lingyan Kong5Chuang Liu6 Jianping Cheng1
Jianping Cheng1 Jingjun Ruan1*
Jingjun Ruan1*- 1College of Agriculture, Guizhou University, Guiyang, Guizhou, China
- 2College of Food Science and Engineering, Xinjiang Institute of Technology, Aksu, China
- 3Agricultural Service Center of Langde Town, Kaili, Guizhou, China
- 4Chengdu Institute of Food Inspection, Chengdu, Sichuan, China
- 5The First Senior Middle School of Yuanyang County, Xinxiang, Henan, China
- 6Henan Institute of Technology, Xinxiang, Henan, China
The second-largest transcription factor superfamily in plants is that of the basic helix-loop-helix (bHLH) family, which plays an important complex physiological role in plant growth, tissue development, and environmental adaptation. Systematic research on the Chenopodium quinoa bHLH family will enable a better understanding of this species. Herein, authors used a variety of bioinformatics methods and quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) to explore the evolution and function of the 218 CqbHLH genes identified. A total of 218 CqbHLH transcription factor genes were identified in the whole genome, located on 18 chromosomes. A phylogenetic tree was constructed using the CqbHLH and AtbHLH proteins to determine their homology, and the members were divided into 20 subgroups and one unclustered gene. Authors also analyzed 218 CqbHLH genes, conservative motifs, chromosome diffusion, and gene replication. The author constructed one Neighbor-Joining (NJ) tree and a collinearity analysis map of the bHLH family in C. quinoa and six other plant species to study the evolutionary relationship and homology among multiple species. In addition, the expression levels of 20 CqbHLH members from different subgroups in various tissues, different fruit developmental stages, and six abiotic stresses were analyzed. Authors identified 218 CqbHLH genes and studied their biological functions, providing a basis for better understanding and further studying the bHLH family in quinoa.
1 Introduction
The bHLH transcription factor (TF) family is one of the most important in plants and plays a vital role in complex physiological processes (Murre et al., 1989). The bHLH domain comprises approximately 60 amino acids. The first part mainly comprises 13-17 amino acids, the basic N-terminal residues. The second component, the HLH domain, is mainly composed of a helix-loop-helix region of approximately 40 hydrophobic amino acids called the C-terminus, which allows bHLH proteins to form homologous or isomeric dimers (Atchley et al., 1999). According to the DNA binding and developmental characteristics of bHLH proteins, they can be divided into six groups: A, B, C, D, E, and F (Ledent and Vervoort, 2001); there are specific differences in the core domain sequences in different groups (Dang et al., 1992; Phillips, 1994; Crozatier et al., 1996; Crews, 1998; Fisher and Caudy, 1998; Ledent and Vervoort, 2001; Dang et al., 2011; Hardin, 2004).
Honkanen et al. (2018) found that the bHLH transcription factor encoded by the RSLI-like gene is necessary for regulating root cells; it is conserved in all terrestrial plants, indicating that the mechanism may be ancient and originate from a common ancestor. Pires and Dolan (2010a) found that most bHLH subgroups in angiosperms (such as Arabidopsis and rice) also exist in early differentiation populations of terrestrial plants; therefore, the diversity has been maintained for a long time, and these genes have undergone a long evolutionary process. Pires and Dolan (2010b) studied the whole genomes of some terrestrial plants and algae. They found that the diversity of bHLH proteins was established in the early terrestrial plants 440 million years ago, which is highly conserved. These characteristics may increase environmental adaptability and support morphological changes in plants.
Wang et al. (2018b) found that eight Setaria italica bHLH genes significantly changed expression levels under drought stress. The drought response elements have also been found in the cis-acting element analysis of these genes. Li et al. (2020) found that CsbHLH041, a bHLH transcription factor gene in Cucumis sativus, is an important regulator of salt and ABA (abscisic acid) tolerance. Jiang et al. (2009) also found that transcription factor AtbHLH92 in Arabidopsis thaliana plays a vital regulatory role in salt and osmotic stress tolerance. AtbHLH112, another member of the gene family, is also important in regulating plant drought resistance (Li et al., 2021). Zhao et al. (2020) found that MdbHLH130 is an important regulator of water stress, and CsbHLH18 was found by Geng and Liu (2018) to play a significant role in regulating cold stress. At present, bHLH transcription factor research has revealed a crucial regulatory role in response to various abiotic stresses (drought, cold, salt, etc.). According to Zhai et al. (2016), the expression level of TabHLH39 in Triticum aestivum changed significantly in the roots, stems, and leaves at the seedling stage, as well as under abiotic stress (drought, salt, and cold). This indicates that bHLH transcription factors play an important role in abiotic stress resistance and plant growth.
bHLH transcription factors are closely involved in plant activities, especially in growth, fruit development, and environmental adaptation (Duek and Fankhauser, 2005; Feller et al., 2011). According to Wang et al. (2021), the overexpression of BvbHLH93 in sugar beets can enhance antioxidant enzyme production and reduce reactive oxygen species production, significantly improving salt tolerance. AtNHX1 and AtNHX6 regulate salt tolerance in Arabidopsis, while AtMYC2 and AtbHLH122 regulate the expression of AtNHX1 and AtNHX6 under the guidance of ABA (Luo et al., 2009; Krishnamurthy et al., 2019). MfbHLH38 from Myrothamnus flabellifolia was transduced into Arabidopsis, improving drought and salt tolerance (Qiu et al., 2020). bHLH transcription factors also directly or indirectly regulate tolerance to salt and drought in rice (Li et al., 2010; Ikeda et al., 2013; Liu et al., 2022). PalbHLH1 and PalMYB90 are at high expression in poplar, thereby enhancing the activity of antioxidant enzymes, releasing H2O2, and improving its resistance to Botrytis cinerea. bHLH transcription factors also play an essential role in biomass synthesis; MrbHLH1 is an indispensable synergistic gene that regulates MrMYB1 anthocyanin synthesis. The two genes can form the MrMYB1-MrbHLH1 complex to regulate anthocyanin synthesis (Liu et al., 2013). DhbHLH1 and DhMYB2 interact to regulate the production of anthocyanins in Dendrobium hybrid petals, and both DhMYB2 and DhbHLH1 can also induce white petals to produce anthocyanins. bHLH transcription factors also regulate lignin synthesis and have a role in hormone signal transduction, involving ABA (Li et al., 2017), jasmonic acid, brassinosteroids, salicylic acid, and ethylene (Pires and Dolan (2010a); Feller et al., 2011; Dang et al., 2011).
In 2017, the latest C. quinoa genome sequence was published (https://www.cbrc.kaust.edu.sa/chenopodiumdb/) (Jarvis et al., 2017), enabling us to study the evolution, development, and gene function of quinoa better. Because of the important role of bHLH transcription factors in plants, a large number of gene families have been discovered in Arabidopsis thaliana (Toledo-Ortiz et al., 2003), Oryza sativa (Li et al., 2006), Solanum tuberosum (Wang et al., 2018c), Brassica rapa (Song et al., 2014), Solanum lycopersicum (Sun et al., 2015), S. italica (Fan et al., 2021a), Zea mays (Zhang et al., 2018a), T. aestivum (Wei and Chen, 2018), Capsicum annuum (Zhang et al., 2020), Fagopyrum tataricum (Sun et al., 2020), Sorghum bicolor (Fan et al., 2021b), Brachypodium distachyon (Niu et al., 2017), and Vitis vinifera (Wang et al., 2018a). In this study, we identified 218 C. quinoa bHLH family members in the genome, divided them into 20 subgroups, and analyzed their gene structure, conserved motifs, gene distribution, phylogeny, and homology. We also examined the bHLH gene expression levels under abiotic stress and during stages of growth and development, which helps study the gene function and evolution in C. quinoa.
2 Materials and methods
2.1 Gene identification
The latest C. quinoa genome sequence was downloaded from https://www.cbrc.kaust.edu.sa/chenopodiumdb/. The bHLH family members were identified in two BLASTp searches (Altschul et al., 1997; Liu et al., 2018). First, all possible bHLH proteins were identified from all the sequences of C. quinoa using BLASTp (score ≥ 100, e ≤ 1e10) and the Pfam Protein Family Database (http://pfam.sanger.ac.uk/). A consistent Hidden Markov model (HMM) profile for the bHLH domain (PF00011) was obtained. We used HMMER 3.0 software (with default parameters) with a cutoff value of 0.01 (http://plants.ensembl.org/hmmer/index.html) and SMART software (http://smart.emblheidelberg.de/) to determine the existence of the bHLH domains (Bateman et al., 2000; Finn et al., 2011; Letunic and Bork, 2018), verify the results, and deduce the C. quinoa bHLH family members. In addition, from the ExPasy website (http://web.expasy.org/protparam/), the basic characteristics of the members of the CqbHLH gene family were identified: protein length, coding sequence length (CDS), isoelectric point (pI), molecular weight (MW) and subcellular localization.
2.2 bHLH gene structure
ClusterW with the default parameters was used to create multiple protein sequence alignments between 218 CqbHLH genes and Arabidopsis bHLH proteins (Thompson et al., 2003). GeneDoc software was used to manually adjust the deduced amino acid sequence of the bHLH domain. The exon/intron structure of each CqbHLH gene was analyzed using the gene structure display server GSDS (http://GSDS.cbi.pku.edu.cn) (Guo et al., 2007). The bHLH protein motifs were identified using the MEME server (http://meme-suite.org/tools/meme) with the following settings: 10 motifs and 6–200 residues (Bailey et al., 2009; Liu et al., 2018; Xie et al., 2018; Liu M. et al., 2019).
2.3 Chromosome distribution and gene replication
Using Circos and the C. quinoa database (https://www.cbrc.kaust.edu.sa/chenopodiumdb/) (Krzywinski et al., 2009), all CqbHLH genes were mapped to the 18 chromosomes and Chr00 of C. quinoa. The MCScan X tool kit was used to analyze gene replication events with the default parameters (Wang et al., 2012). We used TBtools’ Dual Syntony Plotter (https://github.com/CJ-Chen/TBtools) to analyze homology within six species (A. thaliana, V. vinifera, S. lycopersicum, B. detachyon, O. sativa, and Z. mays). The Ka/Ks Calculator 2. Zero was used to calculate the non-synonymous (Ka) and synonymous (Ks) values of each duplicate bHLH gene (Wang et al, 2010).
2.4 Phylogenetic analysis and classification of the CqbHLH gene family
All identified CqbHLH genes were grouped according to the classification of bHLHTFs in A. thaliana. The Jukes–Cantor model of MEGA 7.0 was used to build an NJ tree with a bootstrap value of 1000, and the BLOSUM62 cost matrix was used to allocate Genetic R11. The UniProt database (https://www.uniprot.org/) and published journals were used to obtain three dicotyledons, A. thaliana, V. vinifera, and S. lycopersicum, and three monocotyledons, B. distachyon, O. sativa, and Z. mays; protein sequences were combined with 218 CqbHLH sequences for phylogenetic analysis.
2.5 Bicolor plant materials, growth conditions, and abiotic stresses
Wild C. quinoa (2n = 4x = 36) is the latest sequenced C. quinoa variety and is widely researched. In 2021, C. quinoa was planted at Guizhou University. C. quinoa Material I was planted in a pot plant mixed with soil and vermiculite (1:1); the relative humidity in the growth chamber was 75%, sunlight lasted 16 h per day (25°C), and darkness lasted 8 h per day (20°C). When the plants grew to the seedling stage (on the 21st day), we took five samples with good growth and in similar condition, collected roots, stems, and leaves, and quickly stored them in liquid nitrogen at −80° C for further use. They were then subjected to six abiotic stress treatments, including high temperature (40°C), low temperature (4°C), flooding (of the whole plant), drought (30% PEG6000), salt (5% NaCl), and ultraviolet radiation (70 µW/cm, 220 V, 30 W). The roots, stems, and leaves samples were taken after 0, 2, and 24 h, respectively, and quickly stored in liquid nitrogen at −80°C for further use. The C. quinoa material II was planted in the experimental field of Guizhou University, and the cultivation and management measures were consistent with the field practice. Five quinoa plants of similar growth status and length were selected to take tissue samples of roots, stems, leaves, flowers, and fruits (middle of grain-filling stage) samples were taken at maturity and immediately placed in liquid nitrogen, and stored at −80°C. When the fruit begins to appear, fifteen plants with uniform growth were selected for mixed sampling on days 7, 14, 21, 28, and 35 when the fruits appeared. Immediately after sampling, place the sample in liquid nitrogen and store it at -80°C.
2.6 Total RNA extraction, cDNA reverse transcription, and qRT PCR analysis
Total RNA was extracted using a plant RNA extraction kit (TaKaRa Bio) and treated with DNase I without the RNA enzyme to remove trace DNA. qRT-PCR primers were designed using Primer Premier 5.0 software (Table S8) (Singh et al.,1998). The internal control was the GAPDH gene (Sudhakar Reddy et al., 2016). qRT-PCR was performed at least three times using SYBR Premix Ex TaqII (TaKaRa Bio). We used 2−(ΔΔCt) to calculate the relative gene expression (Livak and Schmittgen, 2001).
2.7 Statistical and analysis
We used JMP6. Zero software (SAS Institute) to analyze all of the above data through ANOVA. The least significant difference (LSD) was compared, with a significance level of p<0.05. Origin 8.0 software was used to draw a histogram.
3 Results
3.1 Identification of bHLH genes in C. quinoa
We used two domain identification methods to find all the possible bHLH family members in the C. quinoa genome. To distinguish the genes, they were named CqbHLH1 to CqbHLH218 according to their chromosomal position. The characteristics of these genes, including the amino acid number, molecular weight (Mw), isoelectric point (pI), coding sequence length (CDS), domain information, and subcellular localization, are provided (Table S1). Among the 218 proteins, CqbHLH32 was the smallest, with only 84 amino acids, and CqbHLH86 was the largest, containing 962 amino acids; the average amino acid number was 324. The Mw of the 218 CqbHLHs ranged from 9.84 kDa (CqbHLH32) to 104.51 kDa (CqbHLH86), with an average of 35.86 kDa. The pI ranged from 4.6 (CqbHLH215) to 11.84 (CqbHLH93), with an average value of 6.78. Of these 218 CqbHLH proteins, 2 contained only the bHLH-MYC_N domain (CqbHLH16 and CqbHLH167), and 192 had only the HLH domain. According to the predicted subcellular localization, 183 CqbHLHs were located in the nucleus, 13 in the chloroplast, 13 in the cytoplasm, two in the Golgi apparatus (CqbHLH62, CqbHLH70), two in the peroxisome (CqbHLH16, CqbHLH66), two in the plasma membrane (CqbHLH86, CqbHLH116), one in the mitochondria (CqbHLH177), one in the endoplasmic reticulum (CqbHLH151), and one in the extracellular matrix (CqbHLH40). The proportion of CqbHLH genes in C. quinoa to the total gene number was approximately 0.49%, similar to that in O. sativa (0.44%) and S. lycopersicum (0.46%) and less than that in A. thaliana (0.59%) (Toledo-Ortiz et al., 2003; Li et al., 2006; Sun et al., 2015).
3.2 Multiple sequence alignment, phylogenetic analysis, and classification of CqbHLH proteins
Using the 218 identified CqbHLH proteins and other 61 Arabidopsis bHLH proteins, we constructed a phylogenetic tree with a bootstrap value of 1000 through Neighbor-Joining (NJ) (Figure 1A; Tables S1, S2). Based on the topological structure of the tree and the classification method proposed by Pires and Toledo-Ortiz (Pires and Dolan (2010a); Toledo-Ortiz et al., 2003), we divided the 218 CqbHLH proteins into 20 subgroups and one ungrouped protein (CqbHLH135), consistent with the classification group of bHLH proteins in Arabidopsis, indicating that they were retained during long-term evolution. However, subgroup 20 was absent from the bHLH family of C. quinoa, indicating that it may have been lost or undifferentiated in the long-term evolution of C. quinoa. Among the 20 subgroups of C. quinoa, subgroup 18 had the most members (23 CqbHLHs), while subgroups 13 and 21 had the least members (2 CqbHLHs). The phylogenetic tree showed that some CqbHLHs were closely bound to AtbHLHs (bootstrap support ≥ 80). The collinearity analysis of C. quinoa and A. thaliana revealed the homology of CqbHLHs with AtbHLHs, speculated to have similar functions.
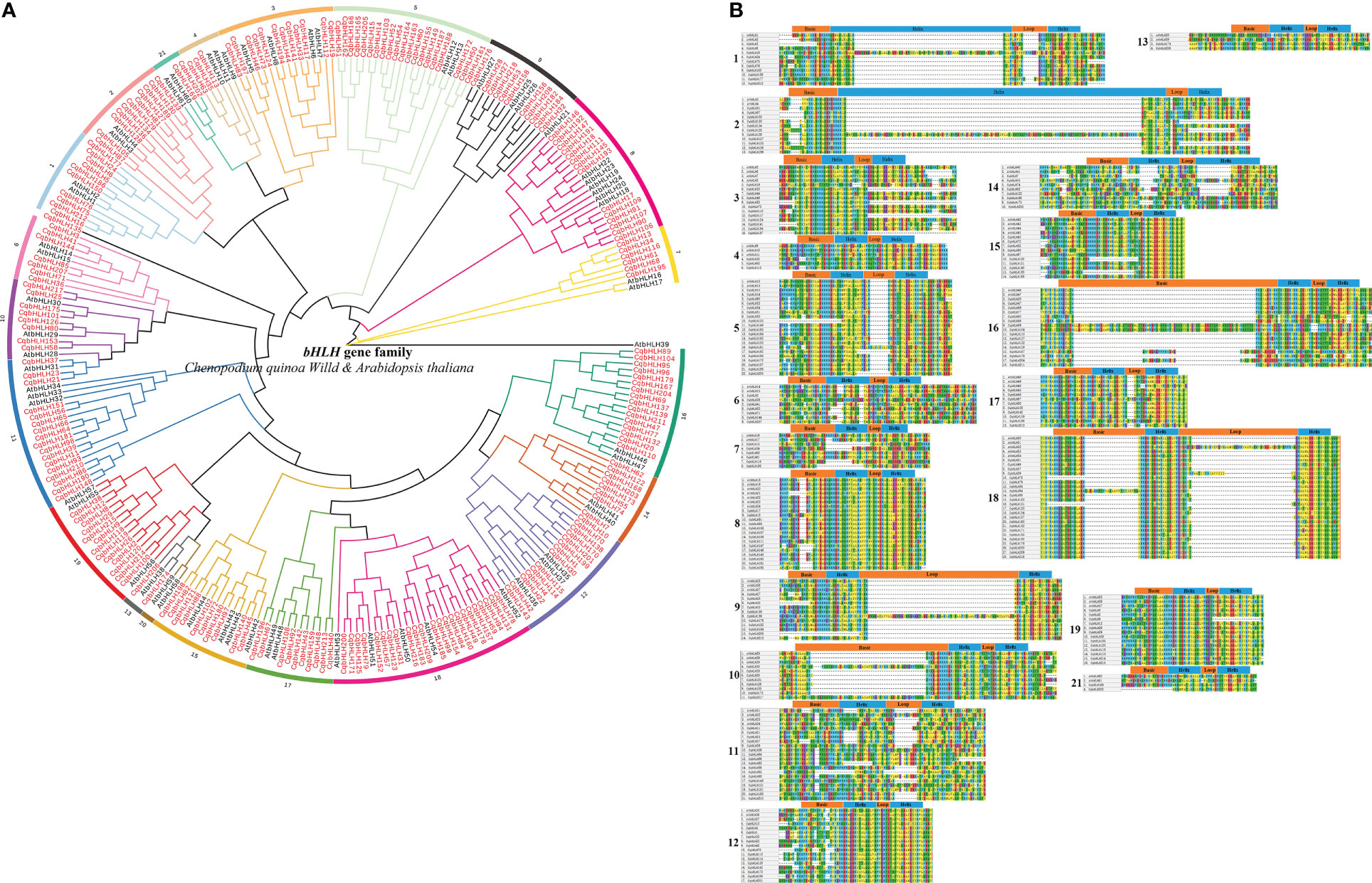
Figure 1 (A) A rootless phylogenetic tree showing the relationship between the bHLH domains of C. quinoa and A. thaliana, where AtbHLH represents a bHLH protein of A. thaliana. (B) Multiple sequence alignment between C. quinoa and 20 subgroup of A. thaliana. C. quinoa lacks the subgroups 20. The boundary and location of the basic helix loop helix are described at the top of the subgroup.
The bHLH domains of the selected A. thaliana and C. quinoa family members were sequenced (Figure 1B); most of the bHLH domains in C. quinoa span approximately 53 amino acids. There were 17 amino acids in the basic region, 15 in the helix region, and six in the loop region (Toledo-Ortiz et al., 2003). Meanwhile, some C. quinoa bHLH family members’ conservative domains also showed a significant amino acid number increase (such as the helix region of CqbHLH16 and CqbHLH129). The loop is the most divergent region of this domain, especially in subgroups 1, 5, 9, 11, 17, and 18. The same is true for bHLH proteins in other plants, including A. thaliana (Toledo-Ortiz et al., 2003), S. lycopersicum (Sun et al., 2015), S. italic (Fan et al., 2021a) and F. tataricum (Sun et al., 2020).
3.3 Analysis of conserved domains, gene structure, and cis-acting elements of CqbHLH genes
Understanding the intron-exon structures of CqbHLH genes will help explore their physiological functions. A comparison of the number and location of exons and introns shows that the 218 CqbHLHs genes have between one and 19 exons (Figures 2A, B; Tables S1, S8); 14 genes (6.42%) contain one exon, and the rest have two or more exons. The 14 intron-free genes belong to three subgroups (8, 14, and 19) but are mainly found in subgroup 8 and 19, among the 42 genes with introns, four are the most common. CqbHLH27 has the most introns (18), while members of Group 8 had no introns or only one intron. Group 9 had the most variation in exon number: one intron in CqbHLH32) and 18 introns in CqbHLH27. Through comparative analysis, we found that subgroup 11 showed the greatest diversity in intron numbers.

Figure 2 Phylogenetic tree, gene structure, and motif distribution of 218 bHLH genes in C. quinoa. (A) The phylogenetic tree is constructed using the NJ (Neighbor- Joining) method, with 1000 replicates on each node. (B) Pink area: UTR (untranslated area), yellow area: CDS (coding sequence), and yellow/dark green area: bHLH domain (HLH, bHLH-MYC_N). (C) The colored area represents the conservative motifs (motifs 1–10) of 218 CqbHLHs, and the black line represents the relative protein length.
To further study the characteristic regions of the 218 CqbHLH proteins, we used the online tool MEME (https://meme-suiteorg/meme/tools/meme). The conservative motifs of the 218 CqbHLH proteins were analyzed, and ten were selected. Figure 2C shows that motifs 1 and 2 were widely distributed and the most numerous in CqbHLHs, whereas motif 7 was the least distributed in the CqbHLHs. CqbHLH members in the same subgroup usually have similar motif compositions. For example, the members in subgroup 5 mainly include motifs 1, 2, 4, and 5. Subgroup 8 mainly includes motifs 1, 2, 6, and 9. Some motifs mostly existed in specific subgroups, such as motif 9 in subgroup 18. Motifs 1 and 2 were found in most of the subgroups. After further sequence analysis, we found that the motifs have specific arrangement characteristics and positional relationships; for example, motif 6 is only found at the end of subgroups 1, 8, 9, 10, 11, 12, and 21; in the subgroup 18, there is a specific arrangement of motifs 3, 1, 2 and 9. Studying the conserved motifs of proteins will help us to research their functions systematically. The functions of most of these CqbHLH proteins are yet to be proven, but their phylogenetic results support the groupings of the C. quinoa bHLH gene subgroups.
We analyzed the cis-acting elements of 218 CqbHLH gene initiation codons, 2000 bp upstream (Table S3). Analysis of the cis-acting elements of the 218 CqbHLHs genes showed that light- and hormone-related elements were the most abundant; ABA response elements were the most abundant among the hormone-related elements. The hormone-related elements include gibberellin, salicylic acid, auxin, and methyl jasmonate response elements. Concerning environmental pressures, the CqbHLH145 promoter contained low-temperature response elements, and CqbHLH145 also showed high expression levels under cold stress (Figure 3A), indicating direct or indirect regulation of the physiological metabolism of quinoa at low temperatures.
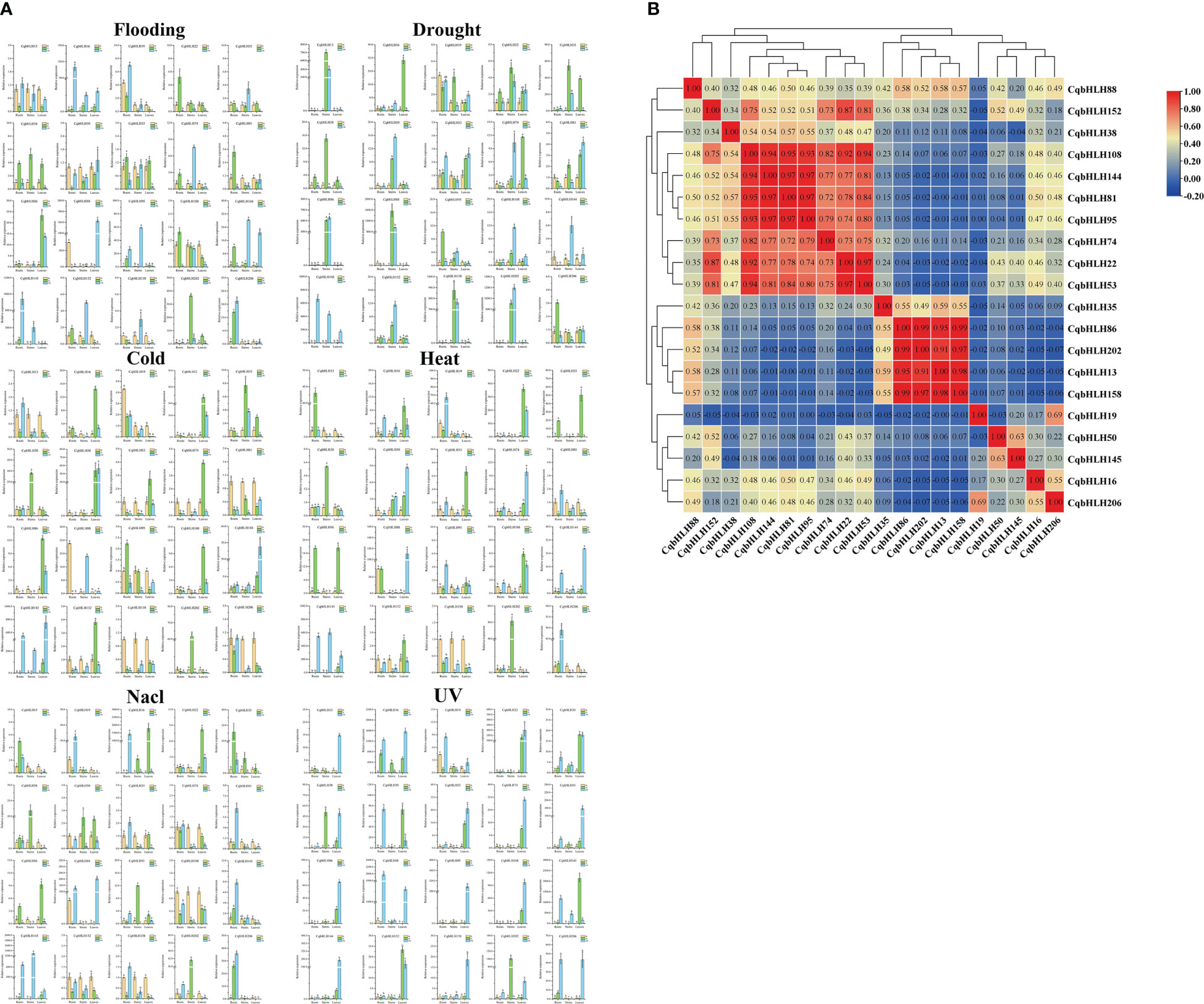
Figure 3 (A) The gene expression patterns of 20 CqbHLH genes in the roots, stems, and leaves of young C. quinoa seedlings treated with 0 h, 2 h, and 24 h abiotic stresses (UV, flooding, drought, NaCl, heat, and cold) were detected by qRT-PCR. The error value is obtained from the three measurements. The lowercase letters above the bar graph represent significant differences between different letters (p < 0.05, LSD). (B) Number > 0: positive correlation; Number < 0: negative correlation; 0.05 represents a significant correlation.
3.4 CqbHLH chromosome distribution and gene replication
A physical location map of CqbHLH was created (Figure 4A; Table S4). The 218 CqbHLH genes were unevenly distributed on the 18 chromosomes (Chrs) and Chr00. Each gene is named according to its physical location from top to bottom on Chr1 to Chr18 (Chr00 comprises the segments that could not be located on chromosomes). Among these, Chr07 had the largest number (26 genes, ~11.93%), followed by Chr01 (21 genes, ~9.63%), and Chr04 had the lowest content (4 genes, ~1.83%). Chr16 contained 18 (~8.26%) CqbHLH genes; Chr11 and Chr10 contained 15 (~6.88%) CqbHLH genes; Chr14 contained 14 (~6.42%) CqbHLH genes; Chr08 and Chr06 contained 13 (~5.96%) CqbHLH genes; Chr15 contained 12 (~5.50%) CqbHLH genes; Chr15 contained 11 (~5.05%) CqbHLH genes; Chr09 and Chr02 contained 9 (~4.13%) CqbHLH genes; Chr00 contained 8 (~3.67%) CqbHLH genes; Chr05 contained 7 (~3.21%) CqbHLH genes; Chr03, Chr12, and Chr13 each contained 6 (~2.75%) CqbHLH genes; Chr18 contains five (~2.29%) CqbHLH genes. Most of the CqbHLH genes were distributed on the 18 chromosomes and at the end of Chr00. We observed many CqbHLH gene replication events; chromosomal regions within 200 kb of two or more identical genome regions were defined as tandem duplication events (Chen et al., 2017). Twelve tandem repeats were found on Chr01, Chr02, Chr07, Chr08, Chr09, Chr11, Chr13, Chr15 and Chr16, involving 26 CqbHLH genes. Among them, CqbHLH128, CqbHLH129, CqbHLH130, CqbHLH133, CqbHLH134, and CqbHLH135 on Chr11 had two tandem repeat events (CqbHLH129 and CqbHLH128/CqbHLH130; CqbHLH134 and CqbHLH133/CqbHLH135). All genes with tandem repeats belonged to the same subgroup (except for CqbHLH135). For example, CqbHLH5 and CqbHLH6 are tandem repeats clustered in subgroup 1.
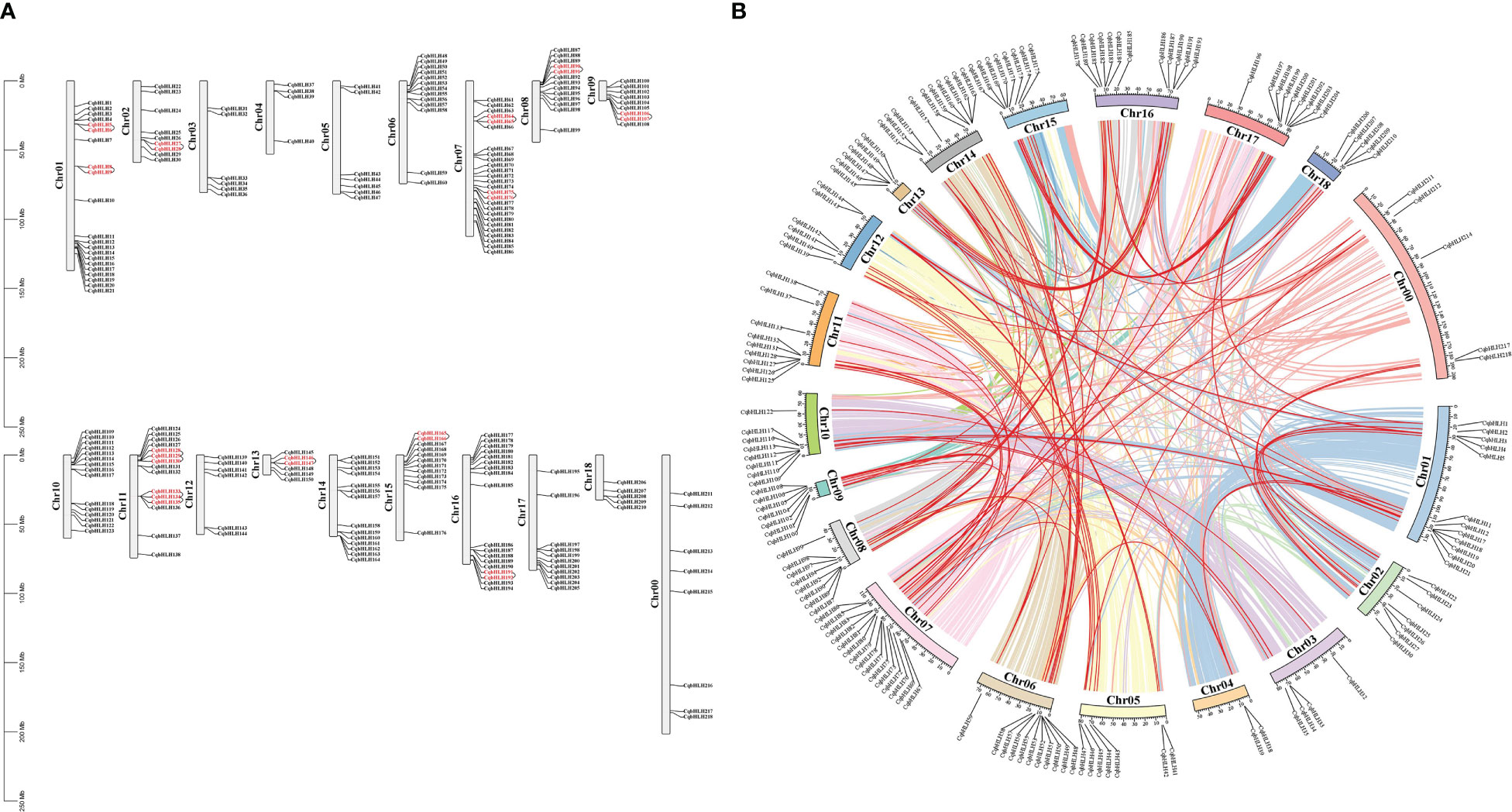
Figure 4 (A) Location map of 218 bHLH genes on C. quinoa chromosomes. The scale on the left side of the chromosome indicates its length. To the left side of the chromosome is its number; to the right side is the C. quinoa bHLH gene contained in the chromosome; the red CqbHLH is a tandem repeat gene. (B) The schematic diagram of the distribution of C. quinoa genes on the chromosomes and the relationship between chromosomes. Different color lines inside represent gene pairs between different chromosomes; red lines represent C. quinoa bHLH gene pairs; the C. quinoa bHLH genes are indicated outside the chromosome, inside of which is the chromosome number.
In addition, the gene fragment repeat analysis showed that there were 152 similar sites in CqbHLH, and 95 pairs of fragments were duplicated. A total of 152 (69.72%) homologous genes were identified in the CqbHLH gene family, indicating an evolutionary relationship between bHLH members (Figure 4B; Table S5). The CqbHLH gene was unevenly distributed among the 19 C. quinoa linkage groups (LGs). Some LGs (LG07) had more CqbHLH genes than others (LG04 and LG03). The CqbHLH gene of LG07 is the most (15), whereas that of LG04 is the least (2). Further analysis of the genes showed that fragments of repeated CqbHLHs were connected within their subgroups, except for CqbHLH13/CqbHLH22, CqbHLH7/CqbHLH143, CqbHLH32/CqbHLH122, CqbHLH42/CqbHLH208, and CqbHLH184/CqbHLH47. These results indicate that some CqbHLH genes may result from replication events, which play an essential role in the evolution of C. quinoa and amplify the CqbHLH gene family.
3.5 Collinearity analysis of CqbHLH
To further determine the phylogenetic mechanisms of the bHLH family, we constructed a collinearity diagram of C. quinoa and six representative species (three representative dicotyledon plants: A. thaliana, S. lycopersicum, and V. vinifera, and three representative monocotyledon plants: B. distachyon, O. sativa, and Z. mays) (Figure 5; Table S6). A total of 147 CqbHLH genes were found to have a collinear relationship with the genes of A. thaliana (82), V. vinifera (90), S. lycopersicum (96), B. distachyon (33), O. sativa (28) and Z. mays (15). At the same time, the homologous logarithms for the six species are A. thaliana (135), V. vinifera (184), S. lycopersicum (191), B. detachyon (57), O. sativa (48) and Z. mays (23). Some CqbHLH genes are associated with at least four synthetic gene pairs (particularly C. quinoa and B. detachyon), such as CqbHLH33 and CqbHLH117. CqbHLH and A. thaliana (52.0%), B. detachyon (52.63%), O. sativa (56.25%), and S. lycopersicum (59.68%) contained two or more synthetic gene pairs, accounting for more than 50% of the total homologous logarithm, suggesting that these genes are essential in evolution.

Figure 5 The collinearity diagram of bHLH genes in quinoa and six other representative plants: three dicotyledons (tomato, Arabidopsis, and grape) and three monocotyledons (two spike grass, rice, and corn). The gray line represents the collinear gene pairs between the two species, and the red line represents the bHLH gene pair.
As expected, some CqbHLH genes only had homologous gene pairs with three dicotyledons, such as CqbHLH55, CqbHLH58, CqbHLH71, and CqbHLH72. They all have at least one pair of homologous genes with three selected representative dicotyledons, indicating that these genes may have gradually formed after the independent differentiation of dicotyledons. We also found that some of the CqbHLH genes pair with at least one homologous gene within the six species, such as CqbHLH139, CqbHLH30, CqbHLH22, CqbHLH74, CqbHLH117, CqbHLH47, CqbHLH1, and CqbHLH33, indicating that they may be essential primitive genes that have not been lost or shown strong differentiation during the long-term evolution of C. quinoa.
3.6 Evolutionary analysis of bHLH Proteins in C. quinoa and six other species
To analyze the evolutionary relationship between the C. quinoa protein triple helix family and six other plants (A. thaliana, B. distachyon, S. lycopersicum, V. vinifera, O. sativa, and Z. mays), we used the MEME web server and the Genes R11 method to construct a rootless NJ tree with ten conserved motifs from the amino acid sequences of the 218 CqbHLH proteins and the bHLH proteins (355) from the other six plants (Figure 6; Tables S7, S8). The distribution of CqbHLHs in phylogenetic trees is relatively scattered, while the CqbHLH protein is more inclined to S. lycopersicum. The bHLH protein aggregation of S. lycopersicum is shown in Figure 3. The proteins of the seven plants mainly contained motifs 1, 2, and 3. Motifs 1, 2, and 3 were almost all arranged in the same way in the seven species. Subgroup 8 contained the most: Motifs 9, 7, 8, 1, 3, 2, 5, and 4. bHLH proteins from the same branch of rootless NJ trees usually had similar motifs and motif arrangements; compared with the other four selected representative plants, bHLH proteins tended to be on the same branch in B. detachyon and S. lycopersicum, showing that their relationship with CqbHLH proteins may be close, compared with the other four plants.
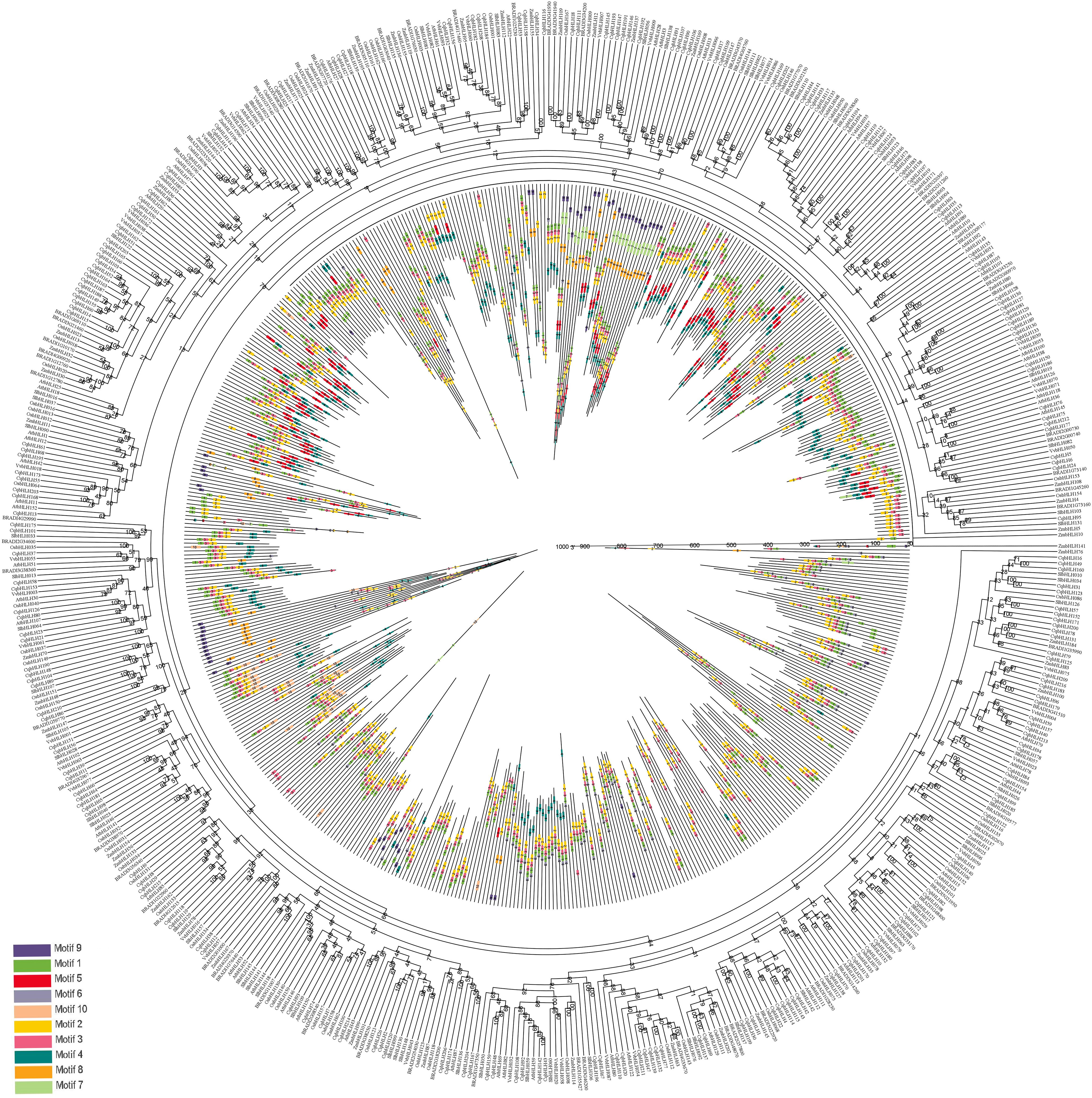
Figure 6 Phylogenetic tree and conserved motifs (1–10) bHLH proteins in quinoa and six representative plants (three dicotyledons representing S. lycopersicum, A. thaliana, and V. vinifera; three monocotyledons representing B. distachyon, O. sativa, and Z. mays). The outer circle is the quinoa and six species bHLH rootless phylogenetic tree, and the inner circle is the distribution of conservative motifs in and the relative length of bHLH proteins.
3.7 CqbHLH expression patterns in different plant organs and at different times in fruit stage
To study the role of CqbHLHs in the growth period of C. quinoa, we selected 20 representative genes from the CqbHLH gene family according to the homologous relationship between CqbHLHs and AtbHLH genes and the evolutionary relationship of NJ tree. We used qRT-PCR (quantitative real-time polymerase chain reaction) technology to detect the gene relative expression of these members in different organs during the appropriate development stages of C. quinoa plants (Figures 7A, B). The results showed different expression patterns in the selected 20 CqbHLH genes. At different times of fruit development (Figure 7A), it was found that among the 20 CqbHLH genes, most of them (CqbHLH16, CqbHLH19, CqbHLH22, CqbHLH38, CqbHLH50, CqbHLH53, CqbHLH74, CqbHLH81, CqbHLH108, CqbHLH144, CqbHLH152, CqbHLH158 and CqbHLH206) showed the highest expression in 21 D (day). Three genes (CqbHLH13, CqbHLH86, and CqbHLH88) were found to have the highest expression in 35 D. No genes were found to have the highest expression at 7 D. In the different organs (Figure 7B), it was found that most of the 20 CqbHLH genes (CqbHLH16, CqbHLH38, CqbHLH50, CqbHLH53, CqbHLH74, CqbHLH81, CqbHLH108, CqbHLH144, CqbHLH152, CqbHLH158 and CqbHLH206) were highest expressed in fruits. In the stem, four genes (CqbHLH13, CqbHLH35, CqbHLH86, and CqbHLH88) showed the highest expression. Some genes have been found to exhibit typical tissue expression specificity, which may be related to the physiological pathways in which they participate.
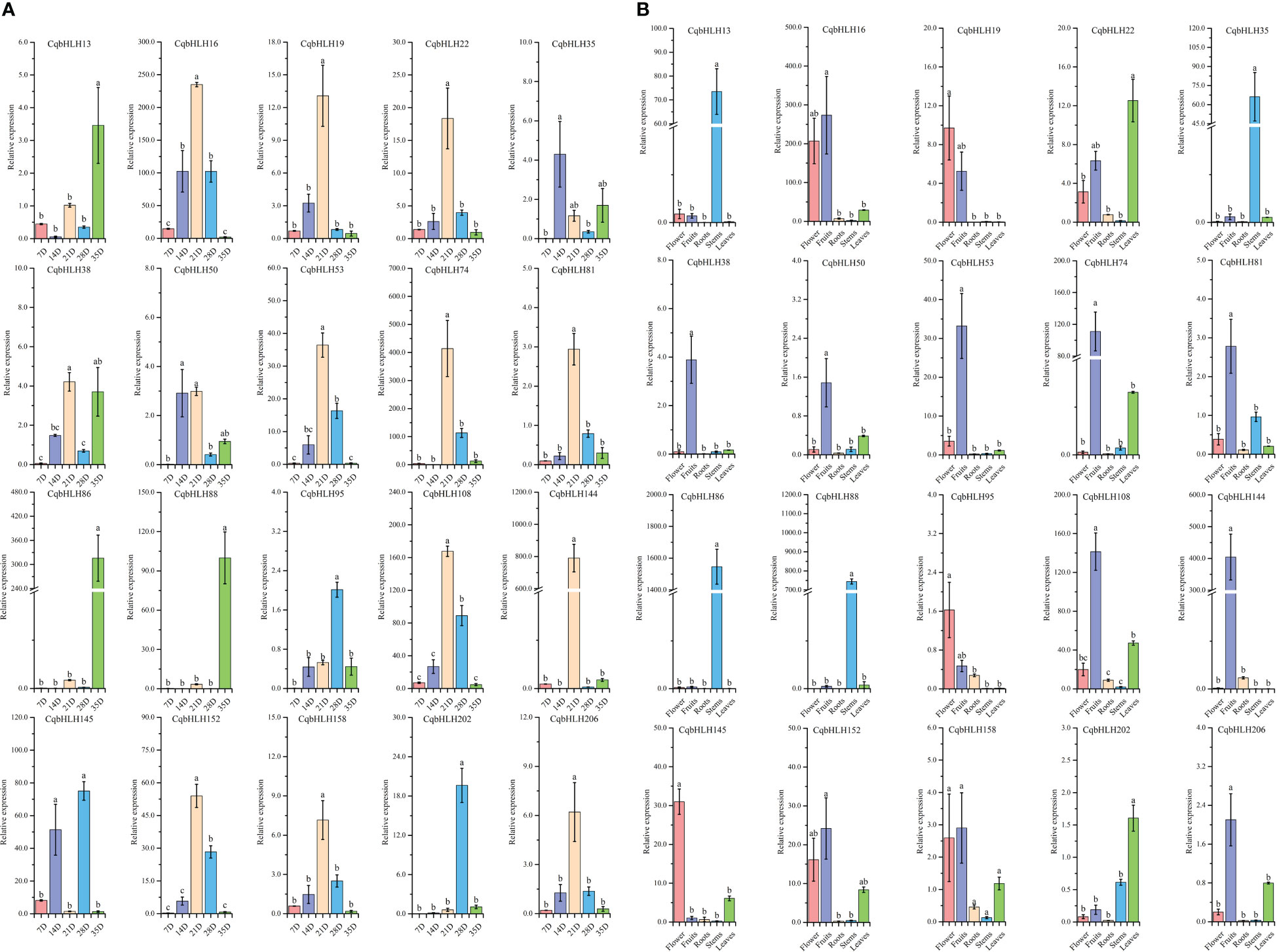
Figure 7 (A) The expression patterns of 20 CqbHLH genes at different stages after fruits appeared were detected by qRT-PCR. Among them, 7 D, 14 D, 21 D, 28 D, and 35 D represent the 7th, 14th, 21st, 28th, and 35th days after fruits appeared. (B) The tissue-specific expression patterns of 20 bHLH genes in roots, stems, leaves, flowers, and fruits were detected by qRT-PCR. The error value is obtained from the three measurements. The lowercase letters above the bar graph represent significant differences between different letters (p< 0.05, LSD).
3.8 Effects of different treatments on CqbHLH expression
To study the effects of abiotic stresses under different stresses on the CqbHLH gene, we carried out six abiotic stress treatments: strong ultraviolet radiation stress (UV), flooding stress (flooding), drought stress (drought), salt stress (NaCl), heat stress (heat) and cold stress (cold), and detected the gene expression of 20 CqbHLH genes selected under the six stresses by qRT-PCR (Figure 3A). We found that different stresses induced or inhibited the expression of some genes differently and that significant changes occurred in the early stage of stress. For example, CqbHLH145 is a high expression under six types of stress. In the roots, stems and leaves, its expression was significantly upregulated under cold stress; however, under intense UV radiation, it was significantly upregulated in roots and stems. In contrast, it was significantly upregulated and then significantly downregulated in leaves. Under drought stress, CqbHLH158 was significantly upregulated in the stems, whereas under cold stress, CqbHLH158 was downregulated in the roots, stems, and leaves. Under flooding stress, CqbHLH16 was upregulated in the roots, stems, and leaves, whereas CqbHLH108 was downregulated in the roots, stems, and leaves. Interestingly, we found that the CqbHLH202 gene was upregulated and then downregulated in the stems under cold and heat stress, possibly indicating that the CqbHLH202 gene is crucial for temperature regulation. CqbHLH13 had the opposite expression pattern: it was downregulated in cold-stressed roots and then upregulated back to normal levels. Under heat stress, however, the expression levels first increased and then decreased to normal. We studied the correlation between the expressions of the 20 CqbHLHs genes (Figure 3B). There were two obvious positive correlation regions, and each contained four genes. Area I includes CqbHLH108, CqbHLH144, CqbHLH81 and CqbHLH95. Area II included CqbHLH86, CqbHLH202, CqbHLH13 and CqbHLH158. However, there was also a negative correlation; for example, between CqbHLH16 and CqbHLH206 and CqbHLH86, CqbHLH202, CqbHLH13, and CqbHLH158.
4 Discussion
Based on the whole genome, studying the TF family of a species is conducive to the systematic exploration of the genes and their functions in the long-term development of this species. We identified 218 C. quinoa bHLH genes, whose proteins showed remarkable differences in amino acid number; the longest (CqbHLH86) was 962 amino acids, and the shortest (CqbHLH32) was 84 amino acids, indicating significant variability (Atchley et al., 1999; de Martin et al., 2021). The C. quinoa bHLH subgroup variation is mainly in the basic region, indicating its importance in the bHLH domain (Atchley et al., 1999; Toledo-Ortiz et al., 2003). The basic region determines the determining activity of the bHLH gene and forms homodimers or heterodimers with other transcription factors (Heim et al., 2003; Toledo-Ortiz et al., 2003). The basic region can be determined by whether the CqbHLH gene has DNA-binding activity and can form homodimers and heterodimers (Heim et al., 2003; Toledo-Ortiz et al., 2003). Genes with DNA activity can be further divided into E-box and Non-E-box genes, and E-box can be subdivided into G-box and Non-G-box, according to the essential amino acid residues in the basic region (Atchley et al., 1999; Toledo-Ortiz et al., 2003). In the C. quinoa bHLH family, there were 11 (5.04%) CqbHLHs without DNA-binding activity; 32 (14.68%) CqbHLHs were classified as non-E-box proteins and 175 (80.28%) CqbHLHs were classified as E-box proteins. Among the 175 E-box proteins, 174 (79.82%) were classified as G-box proteins, and one (0.46%) CqbHLH was classified as a non-G-box protein. Among the 20 CqbHLH subgroups, we found that subgroup 8 was the most conservative, while subgroup 11 was the most variable. This situation is the same as that of the subgroup 8 and 11 of the bHLH family species of the dicotyledon A. thaliana (Toledo-Ortiz et al., 2003) and monocotyledon O. sativa (Carretero-Paulet et al., 2010). Monocotyledonous S. italica and S. bicolor had conservative the subgroup 8 and 11 (Fan et al., 2021a; Fan et al., 2021b). In contrast, the monocotyledonous B. distachyon had the most conservation in subgroup 8 (Niu et al., 2017); subgroup 11 showed greater variation, indicating no specificity between dicotyledons and monocotyledons and suggesting that the subgroup 11 of CqbHLHs may have greater functional differences.
We constructed a phylogenetic tree using 218 CqbHLHs and AtbHLHs. Except for subgroup 20, we found at least one CqbHLH subgroup for each AtbHLH subgroups. This indicates that the bHLH family differentiated before Arabidopsis and C. quinoa diverged. Although the number of members and subgroups of the bHLH family in different species is quite different, they may play a similar basic role in development, adaptation, and evolution. For example, in Hordeum vulgare (Ke et al., 2020a), B. napus (Ke et al., 2020b), Raphanus sativus (Wang et al., 2022), and Nicotiana tabacum (Bai et al., 2020), it was found that the bHLH genes in these species are significantly expressed under abiotic stress, proving that the bHLH transcription factors play an essential role in plant development, adaptation, and evolution. In the NJ tree constructed using A. thaliana data (Figure 1), we divided C. quinoa into subgroup 1–21, but no subgroup 20 was found. However, in A. thaliana (Toledo-Ortiz et al., 2003), S. lycopersicum (Sun et al., 2015), and B. napus (Ke et al., 2020b), subgroup 20 was not lost. Therefore, based on the gene structure and evolutionary analysis of A. thaliana, O. sativa, and C. quinoa bHLH itself, we speculate that during the differentiation process, a certain subgroup replaced subgroup 20 and exercised its biological functions, or that subgroup 20 did not differentiate or get lost due to environmental selection. In addition, one CqbHLH had no obvious cluster, which indicates that C. quinoa may have a new evolutionary direction. In terms of gene structure, we found that CqbHLH135 was similar but different from subgroup 16. However, CqbHLH135, CqbHLH133, and CqbHLH134 were tandem repeats. Therefore, we speculate that CqbHLH135 may have differentiated from subgroup 2 earlier. It is similar to subgroup 16 in gene structure but has its unique function during differentiation. S. italica has its subgroup, but the function of these special members in forming their subgroups has not been clarified (Fan et al., 2021a).
In the gene structure analysis, we found that some members had fewer introns than others of the same subgroup; they may have been lost during long-term evolution or reduced in number due to environmental selection, such as CqbHLH7 and CqbHLH74 of subgroup 14. In plants, the number of introns is related to the expression level, and a more compact gene structure may be conducive to rapid gene expression upon stimulation (Wray et al., 2003). The expansion of this gene family occurs mainly through gene replication (Schwechheimer et al., 1998; Li et al., 2006; Wang et al., 2018c). The C. quinoa bHLH family contains 12 tandem repeats, including 26 CqbHLH genes and 95 pairs of fragment repeats, suggesting that the role of fragment repeat may be more significant than that of tandem duplication.
The bHLH TFs have many members, second only to the MYB TFs, which have essential and extensive functions (Buck and Atchley, 2003; Hao et al., 2021). However, research mainly focuses on two model plants, Arabidopsis and rice, and the bHLH family of other plants has not yet been systematically studied. We found that subgroup 18 has the largest number of members (10.55%), similar to the bHLH family of Arabidopsis (Toledo-Ortiz et al., 2003). The CqbHLH145 gene of the selected subgroup 18 members showed a significant increase in gene expression during growth, development, and biological stress and accounted for a large proportion (13.68%) of the fragment repeat in the C. quinoa bHLH family. Therefore, we speculate that, compared with other species, the subgroup 18 genes may play a critical role in the growth and development of C. quinoa, but this has not been proven. In addition, we selected 20 CqbHLH genes from the 20 subgroups (there were no C. quinoa bHLH members in subgroup 20) to study their responses to six abiotic stresses and reproductive stages. Although there were significant differences in the expression levels between the genes, some showed two or more significant differential expression levels. For example, CqbHLH202 of subgroup 21 is high expression under NaCl stress. In A. thaliana, AtbHLH92, also in subgroup 21, is a high expression under NaCl stress (Jiang et al., 2009). In C. quinoa leaves under cold stress, 11 genes were significantly upregulated, and eight were significantly downregulated, indicating that CqbHLH genes have synergistic or antagonistic effects under various stresses. Fan et al. (2021a) found that in S. italica, there was also up/downregulated expression of various SibHLH genes under abiotic stress. We found that the CqbHLH145 gene was significantly upregulated under six stresses, which was not found in A. thaliana and is worthy of further study (Hao et al., 2021).
Cis-acting elements regulate the activity of target genes by binding to trans-acting factors, thereby regulating gene expression (Liu et al., 2019). In this study, 218 cis-acting elements in the promoter region of the CqbHLH genes were predicted, and it was found that the most hormone responsive elements in the promoter region of the CqbHLH genes were the abscisic acid (ABA) responsive elements. bHLH gene is usually involved in ABA signaling pathways to regulate plant drought resistance, such as bHLH122 in A. thaliana, which enhanced drought resistance in A. thaliana by inhibiting the catabolism and metabolism of ABA (Liu et al., 2014). In C. quinoa, it has been found that under drought stress, the bHLH gene is highly expressed in both the stems and leaves of C. quinoa. This may be due to the involvement of the bHLH gene in the ABA signaling pathway, increasing ABA content, or reducing ABA degradation, to improve the drought resistance of C. quinoa. Other studies have found that bHLH is also related to the development of leaf stomata (Yang et al., 2017; Zhang et al., 2018b), while more bHLH genes are highly expressed in C. quinoa leaves. Therefore, it is speculated that the bHLH gene of C. quinoa is also involved in the development of leaf stomata, or in regulating the opening and closing of leaf stomata to improve drought resistance of C. quinoa. This is also consistent with the physiological characteristics of C. quinoa, a plant with strong drought resistance.
We found that 20 CqbHLH genes were positively regulated in the gene expression heat map, indicating that multiple CqbHLH genes may be co-expressed to perform this physiological function. In addition, it is crucial to study the role of the bHLH family in the reproductive organs of C. quinoa. We investigated the expression of 20 bHLH genes at different times in the main organs of C. quinoa at the reproductive stage. The CqbHLH genes also play important roles in the reproductive development of C. quinoa. For example, the expression level of CqbHLH144 in the fruits was significantly higher than in the roots, stems, leaves, and flowers. Gene expression reached its highest level on the 21st day after flowering. CqbHLH145, like CqbHLH144, exhibits significant changes in gene expression during reproductive development. These two genes are likely to regulate the reproductive developmental stage of C. quinoa. In the maize bHLH family, the yeast two-hybrid test showed that ZmbHLH23 and ZmbHLH180 have a synergistic expression (Zhang et al., 2018a). Moreover, according to the gene correlation heat map, we found that two genes (CqbHLH145 and CqbHLH144) showed a significant positive correlation. We speculate that these two genes interact synergistically and coordinate their physiological functions under stress and in the reproductive stages. In conclusion, these results indicated that some functions of CqbHLH may be related to the network mechanisms between genes.
5 Conclusion
In this study, we systematically studied the C. quinoa bHLH gene family at the genome level and performed a series of analyses and verifications. 218 C. quinoa bHLH genes were analyzed, these CqbHLH genes were divided into 20 groups based on their protein motifs and gene structure, and 218 CqbHLH genes were irregularly distributed on 18 chromosomes and Chr00.Gene replication events may have produced some CqbHLH genes; fragment repeat contributes more to amplifying the CqbHLH gene family than tandem duplication. In terms of homology, a phenomenon was discovered that one CqbHLH gene and multiple B. distachyon bHLH genes are homologous. Among the six representative plants, the CqbHLH gene and S. lycopersicum bHLH genes have the most homologous pairs. qRT-PCR results showed that the 20 selected CqbHLH genes were affected by abiotic stress. CqbHLH88 and CqbHLH144 showed significant effects on abiotic stress resistance; CqbHLH88 and CqbHLH144 also have important effects on the reproductive development stages of C. quinoa. We speculate that CqbHLH88 and CqbHLH144 are greatly significant in the life cycle of C. quinoa. In summary, this study provides basic information on the biological function and bioinformatics of C. quinoa.
Data availability statement
The entire Chenopodium quinoa genome sequence information was obtained from the Phytozome database (http://www.phytozome.net/). The datasets supporting the conclusions of this study are included in the article and its additional files. Further inquiries can be directed to the corresponding author.
Author contributions
GX wrote the manuscript, and GX and YF analysed the data and designed the experiment. GX, YF, CZ, HY, LF, XC, YY, XY, WW, LK, and CL edit graphics and schedules. JC supervised the study, and JC and JR reviewed and revised the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This research was supported by the National Science Foundation of China (32160669 and 32161143005), and the Guizhou Provincial Science and Technology Plan Project (Qiankehe Support [2022] key026.
Acknowledgments
We thank all our laboratory colleagues for providing useful discussions and technical assistance. We are very grateful to the editor and reviewers for critically evaluating the manuscript and providing constructive comments for its improvement.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2023.1171518/full#supplementary-material
Supplementary Table 1 | 218 CqbHLH genes identified and protein sequences.
Supplementary Table 2 | AtbHLH protein sequence for phylogeny.
Supplementary Table 3 | The cis-acting element of the promoter sequence 2,000 bp upstream of 218 bHLH genes in Chenopodium quinoa.
Supplementary Table 4 | The one pairs of tandem duplicates in CqbHLH genes.
Supplementary Table 5 | The 95 pairs of segmental duplicates in CqbHLH genes.
Supplementary Table 6 | One-to-one orthologous relationships between Chenopodium quinoa and 6 different plants.
Supplementary Table 7 | bHLH protein sequences of six species.
Supplementary Table 8 | 218 Conserved Motifs of CqbHLH Protein and 7 species bHLH protein conserved motifs.
Supplementary Table 9 | Primer sequences for qPCR.
Abbreviations
bHLH, Basic helix-loop-helix; CqbHLH, Chenopodium quinoa bHLH; qPR-PCR, Quantitative real-time polymerase chain reaction; HMM, Hidden Markov Model; pI, Isoelectric point; LG, Linkage group.
References
Altschul, S. F., Madden, T. L., Schäffer, A. A., Zhang, J., Zhang, Z., Miller, W., et al. (1997). Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25 (17), 3389–3402. doi: 10.1093/nar/25.17.3389
Atchley, W. R., Terhalle, W., Dress, A. (1999). Positional dependence, cliques, and predictive motifs in the bHLH protein domain. J. Mol. Evol. 48 (5), 501–516. doi: 10.1007/PL00006494
Bai, G., Yang, D. H., Chao, P., Yao, H., Fei, M., Zhang, Y., et al. (2020). Genome-wide identification and expression analysis of NtbHLH gene family in tobacco (Nicotiana tabacum) and the role of NtbHLH86 in drought adaptation. Plant Divers. 43 (6), 510–522. doi: 10.1016/j.pld.2020.10.004
Bailey, T. L., Boden, M., Buske, F. A., Frith, M., Grant, C. E., Clementi., L., et al. (2009). MEME SUITE: tools for motif discovery and searching. Nucleic Acids Res. 37, W202–W208. doi: 10.1093/nar/gkp335
Bateman, A., Birney, E., Durbin, R., Finn, R. D., Hollich, V., Griffiths-Jones, S. (2000). The pfam protein families database. Nucleic Acids Res. 28 (1), 263–266. doi: 10.1093/nar/gkh121
Buck, M. J., Atchley, W. R. (2003). Phylogenetic analysis of plant basic helix-loop-helix proteins. J. Mol. Evol. 56 (6), 742–750. doi: 10.1007/s00239-002-2449-3
Carretero-Paulet, L., Galstyan, A., Roig-Villanova, I., Martínez-García, J. F., Bilbao-Castro, J. R., Robertson, D. L. (2010). Genome-wide classification and evolutionary analysis of the bHLH family of transcription factors in arabidopsis, poplar, rice, moss, and algae. Plant Physiol. 153 (3), 1398–1412. doi: 10.1104/pp.110.153593
Chen, F., Hu, Y., Vannozzi, A., Wu, K. C., Cai, H. Y., Qin, Y., et al. (2017). The WRKY transcription factor family in model plants and crops. Crit. Rev. Plant Sci. 36 (5-6), 311–335. doi: 10.1080/07352689.2018.1441103
Crews, S. T. (1998). Control of cell lineage-specific development and transcription by bHLH-PAS proteins. Gene Dev. 12 (5), 607–620. doi: 10.1101/gad.12.5.607
Crozatier, M., Valle, D., Dubois, L., Ibnsouda, S., Vincent, A. (1996). Collier, a novel regulator of drosophila head development, is expressed in a single mitotic domain. Curr. Biol. 6 (6), 707–718. doi: 10.1016/S0960-9822(09)00452-7
Dang, C. V., Dolde, C., Gillison, M. L., Kato, G. L. (1992). Discrimination between related DNA sites by a single amino acid residue of myc-related basic-helix-loop-helix proteins. P Natl. Acad. Sci. U.S.A. 89 (2), 599–602. doi: 10.1073/pnas.89.2.599
Dang, C. W., Wang, Y., Chen, K. P., Yao, Q., Zhang, D. B., Guo, M. (2011). The basic helix-Loop-Helix transcription factor family in the pea aphid, Acyrthosiphon pisum. J. Insect Sci. 11 (84), 84. doi: 10.1673/031.011.8401
de Martin, X., Sodaei, R., Santpere, G. (2021). Mechanisms of binding specificity among bHLH transcription factors. Int. J. Mol. Sci. 22 (17), 9150. doi: 10.3390/ijms22179150
Duek, P. D., Fankhauser, C. (2005). bHLH class transcription factors take center stage in phytochrome signaling. Trends Plant Sci. 10 (2), 51–54. doi: 10.1016/j.tplants.2004.12.005
Fan, Y., Lai, D., Yang, H., Xue, G., He, A., Chen, L., et al. (2021a). Genome-wide identification and expression analysis of the bHLH transcription factor family and its response to abiotic stress in foxtail millet (Setaria italica l.). BMC Genomics 22 (1), 1–18. doi: 10.1186/s12864-021-08095-y
Fan, Y., Yang, H., Lai, D., He, A., Xue, G., Feng, L., et al. (2021b). Genome-wide identification and expression analysis of the bHLH transcription factor family and its response to abiotic stress in sorghum [Sorghum bicolor (L.) moench]. BMC Genomics 22 (1), 1–17. doi: 10.1186/s12864-021-07652-9
Feller, A., Machemer, K., Braun, E. L., Grotewold, E. (2011). Evolutionary and comparative analysis of MYB and bHLH plant transcription factors. Plant J. 66 (1), 94–116. doi: 10.1111/j.1365-313X.2010.04459.x
Finn, R. D., Clements, J., Eddy, S. R. (2011). HMMER web server: interactive sequence similarity searching. Nucleic Acids Res. 39, W29–W37. doi: 10.1093/nar/gkr367
Fisher, A., Caudy, M. (1998). The function of hairy-related bHLH repressor proteins in cell fate decisions. BioEssays: News Rev. mol. Cell. Dev. Biol. 20 (4), 298–306. doi: 10.1002/(SICI)1521-1878(199804)20:4<298::AID-BIES6>3.0.CO;2-M
Geng, J., Liu, J. H. (2018). The transcription factor CsbHLH18 of sweet orange functions in the modulation of cold tolerance and homeostasis of reactive oxygen species by regulating the antioxidant gene. J. Exp. Bot. 69 (10), 2677–2692. doi: 10.1093/jxb/ery065
Guo, A. Y., Zhu, Q. H., Chen, X., Luo, J. C. (2007). [GSDS: a gene structure display server]. Yi chuan= Hereditas. 29 (8), 1023–1026. doi: 10.16288/j.yczz.2007.08.004
Hao, Y., Zong, X., Ren, P., Qian, Y., Fu, A. (2021). Basic helix-Loop-Helix (bHLH) transcription factors regulate a wide range of functions in arabidopsis. Int. J. Mol. Sci. 22 (13), 7152. doi: 10.3390/ijms22137152
Hardin, P. E. (2004). Transcription regulation within the circadian clock: the e-box and beyond. J. Biol. Rhythm. 19 (5), 348–360. doi: 10.1177/0748730404268052
Heim, M. A., Jakoby, M., Werber, M., Martin, C., Weisshaar, B., Bailey, P. C. (2003). The basic helix-loop-helix transcription factor family in plants: a genome-wide study of protein structure and functional diversity. Mol. Biol. Evol. 20 (5), 735–747. doi: 10.1093/molbev/msg088
Honkanen, S., Thamm, A., Arteaga-Vazquez, M. A., Dolan, L. (2018). Negative regulation of conserved RSL class I bHLH transcription factors evolved independently among land plants. eLife 7, e38529. doi: 10.7554/eLife.38529
Ikeda, M., Mitsuda, N., Ohme-Takagi, M. (2013). ATBS1 INTERACTING FACTORs negatively regulate arabidopsis cell elongation in the triantagonistic bHLH system. Plant Signal Behav. 8 (3), e23448. doi: 10.4161/psb.23448
Jarvis, D. E., Ho, Y. S., Lightfoot, D. J., Schmöckel, S. M., Li, B., Borm, T. J. A., et al. (2017). The genome of chenopodium quinoa. Nature 542 (7641), 307–312. doi: 10.1038/nature21370
Jiang, Y., Yang, B., Deyholos, M. K. (2009). Functional characterization of the arabidopsis bHLH92 transcription factor in abiotic stress. Mol. Genet. Genomics 282 (5), 503–516. doi: 10.1007/s00438-009-0481-3
Ke, Q., Tao, W., Li, T., Pan, W., Chen, X., Wu, X., et al. (2020a). Genome-wide identification, evolution and expression analysis of basic helix-loop-helix (bHLH) gene family in barley (Hordeum vulgare l.). Curr. Genomics 21 (8), 621–644. doi: 10.2174/1389202921999201102165537
Ke, Y. Z., Wu, Y. W., Zhou, H. J., Chen, P., Wang, M. M., Liu, M. M., et al. (2020b). Genome-wide survey of the bHLH super gene family in Brassica napus. BMC Plant Biol. 20 (1), 115. doi: 10.1186/s12870-020-2315-8
Krishnamurthy, P., Vishal, B., Khoo, K., Rajappa, S., Loh, C. S., Kumar, P. P. (2019). Expression of AoNHX1 increases salt tolerance of rice and arabidopsis, and bHLH transcription factors regulate AtNHX1 and AtNHX6 in arabidopsis. Plant Cell Rep. 38 (10), 1299–1315. doi: 10.1007/s00299-019-02450-w
Krzywinski, M., Schein, J., Birol, I., Connors, J., Gascoyne, R., Horsman, D., et al. (2009). Circos: an information aesthetic for comparative genomics. Genome Res. 19 (9), 1639–1645. doi: 10.1101/gr.092759.109
Ledent, V., Vervoort, M. (2001). The basic helix-Loop-Helix protein family: comparative genomics and phylogenetic analysis. Genome Res. 11 (5), 754–770. doi: 10.1101/gr.177001
Letunic, I., Bork, P. (2018). 0 years of the SMART protein domain annotation resource. Nucleic Acids Res. 46 (D1), D493–D496. doi: 10.1093/nar/gkx922
Li, X., Duan, X., Jiang, H., Sun, Y., Tang, Y., Yuan, Z., et al. (2006). Genome-wide analysis of basic/helix-loop-helix transcription factor family in rice and arabidopsis. Plant Physiol. 141 (4), 1167–1184. doi: 10.1104/pp.106.080580
Li, F., Guo, S., Zhao, Y., Chen, D., Chong, K., Xu, Y. (2010). Overexpression of a homopeptide repeat-containing bHLH protein gene (OrbHLH001) from dongxiang wild rice confers freezing and salt tolerance in transgenic arabidopsis. Plant Cell Rep. 29 (9), 977–986. doi: 10.1007/s00299-010-0883-z
Li, C., Qiu, J., Ding, L., Huang, M., Yang, G., Yin, J., et al. (2017). Anthocyanin biosynthesis regulation of DhMYB2 and DhbHLH1 in dendrobium hybrids petals. Plant Physiol. Biochem. 112, 335–345. doi: 10.1016/j.plaphy.2017.01.019
Li, J., Wang, T., Han, J., Ren, Z. (2020). Genome-wide identification and characterization of cucumber bHLH family genes and the functional characterization of CsbHLH041 in NaCl and ABA tolerance in arabidopsis and cucumber. BMC Plant Biol. 20 (1), 272. doi: 10.1186/s12870-020-02440-1
Li, C., Yan, C., Sun, Q., Wang, J., Yuan, C., Mou, Y., et al. (2021). The bHLH transcription factor AhbHLH112 improves the drought tolerance of peanuts. BMC Plant Biol. 21 (1), 540. doi: 10.1186/s12870-021-03318-6
Liu, M., Ma, Z., Sun, W., Huang, L., Wu, Q., Tang, Z., et al. (2019). Genome-wide analysis of the NAC transcription factor family in tartary buckwheat (Fagopyrum tataricum). BMC Genomics 20 (1), 113. doi: 10.1186/s12864-019-5500-0
Liu, M., Ma, Z., Wang, A., Zheng, T., Huang, L., Sun, W., et al. (2018). Genome-wide investigation of the auxin response factor gene family in tartary buckwheat (Fagopyrum tataricum). Int. J. Mol. Sci. 19 (11), 3526. doi: 10.3390/ijms19113526
Liu, Y., Patra, B., Pattanaik, S., Wang, Y., Yuan, L. (2019). GATA and phytochrome interacting factor transcription factors regulate light-induced vindoline biosynthesis in Catharanthus roseus. Plant Physiol. 180 (3), 1336–1350. doi: 10.1104/pp.19.00489
Liu, J., Shen, Y., Cao, H., He, K., Chu, Z., Li, N. (2022). OsbHLH057 targets the AATCA cis-element to regulate disease resistance and drought tolerance in rice. Plant Cell Rep. 41 (5), 1285–1299. doi: 10.1007/s00299-022-02859-w
Liu, W., Tai, H., Li, S., Gao, W., Zhao, M., Xie, C., et al. (2014). bHLH122 is important for drought and osmotic stress resistance in arabidopsis and in the repression of ABA catabolism. New Phytol. 201 (4), 1192–1204. doi: 10.1111/nph.12607
Liu, X. F., Yin, X. R., Allan, A. C., Wang, L. W., Shi, Y. N., Huang, Y. J., et al. (2013). The role of MrbHLH1 and MrMYB1 in regulating anthocyanin biosynthetic genes in tobacco and Chinese bayberry (Myrica rubra) during anthocyanin biosynthesis. Plant Cell Tiss Org. 115, 285–298. doi: 10.1007/s11240-013-0361-8
Livak, K. J., Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2(-delta delta C(T)) method. Methods (San Diego Calif.). 25 (4), 402–408. doi: 10.1006/meth.2001.1262
Luo, B. X., Deng, L. H., Chen, F., Chen, S. Y., Li, W. B., Xiao, G. Y. (2009). Construction of over-expression vector of OsbHLH1 gene and transformation of rice. Biotechnol. Bull. 7, 76–81. doi: 10.13560/j.cnki.biotech.bull.1985.2009.07.023
Murre, C., McCaw, P. S., Baltimore, D. (1989). A new DNA binding and dimerization motif in immunoglobulin enhancer binding, daughterless, MyoD, and myc proteins. Cell 56 (5), 777–783. doi: 10.1016/0092-8674(89)90682-X
Niu, X., Guan, Y., Chen, S., Li, H. (2017). Genome-wide analysis of basic helix-loop-helix (bHLH) transcription factors in Brachypodium distachyon. BMC Genomics 18 (1), 619. doi: 10.1186/s12864-017-4044-4
Phillips, S. E. (1994). Built by association: structure and function of helix-loop-helix DNA- binding proteins. Structure 2 (1), 1–4. doi: 10.1016/s0969-2126(00)00002-2
Pires, N., Dolan, L. (2010a). Early evolution of bHLH proteins in plants. Plant Signal Behav. 5 (7), 911–912. doi: 10.4161/psb.5.7.12100
Pires, N., Dolan, L. (2010b). Origin and diversification of basic-helix-loop-helix proteins in plants. Mol. Biol. Evol. 27 (4), 862–874. doi: 10.1093/molbev/msp288
Qiu, J. R., Huang, Z., Xiang, X. Y., Xu, W. X., Wang, J. T., Chen, J., et al. (2020). MfbHLH38, a Myrothamnus flabellifolia bHLH transcription factor, confers tolerance to drought and salinity stresses in arabidopsis. BMC Plant Biol. 20 (1), 542. doi: 10.1186/s12870-020-02732-6
Schwechheimer, C., Zourelidou, M., Bevan, M. W. (1998). PLANT TRANSCRIPTION FACTOR STUDIES. Annu. Rev. Plant Physiol. Plant Mol. Biol. 49, 127–150. doi: 10.1146/annurev.arplant.49.1.127
Singh, V. K., Mangalam, A. K., Dwivedi, S., Naik, S. (1998). Primer premier: program for design of degesnerate primers from a protein sequence. BioTechniques 24 (2), 318–319. doi: 10.2144/98242pf02
Song, X. M., Huang, Z. N., Duan, W. K., Ren, J., Liu, T. K., Li, Y., et al. (2014). Genome-wide analysis of the bHLH transcription factor family in Chinese cabbage (Brassica rapa ssp. pekinensis). Mol. Genet. Genomics 289 (1), 77–91. doi: 10.1007/s00438-013-0791-3
Sudhakar Reddy, P., Srinivas Reddy, D., Sivasakthi, K., Bhatnagar-Mathur, P., Vadez, V., Sharma, K. K. (2016). Evaluation of sorghum [Sorghum bicolor (L.)] reference genes in various tissues and under abiotic stress conditions for quantitative real-time PCR data normalization. Front. Plant Sci. 7. doi: 10.3389/fpls.2016.00529
Sun, H., Fan, H. J., Ling, H. Q. (2015). Genome-wide identification and characterization of the bHLH gene family in tomato. BMC Genomics 16 (1), 9. doi: 10.1186/s12864-014-1209-2
Sun, W., Jin, X., Ma, Z., Chen, H., Liu, M. (2020). Basic helix-loop-helix (bHLH) gene family in tartary buckwheat (Fagopyrum tataricum): genome-wide identification, phylogeny, evolutionary expansion and expression analyses. Int. J. Biol. Macromol. 155, 1478–1490. doi: 10.1016/j.ijbiomac.2019.11.126
Thompson, J. D., Gibson, T. J., Higgins, D. G. (2003). Multiple sequence alignment using ClustalW and ClustalX. Curr. Protoc. Bioinf. (1), 2–3. doi: 10.1002/0471250953.bi0203s00
Toledo-Ortiz, G., Huq, E., Quail, P. H. (2003). The arabidopsis basic/helix-loop-helix transcription factor family. Plant Cell 15 (8), 1749–1770. doi: 10.1105/tpc.013839
Wang, R., Li, Y., Gao, M., Han, M., Liu, H. (2022). Genome-wide identification and characterization of the bHLH gene family and analysis of their potential relevance to chlorophyll metabolism in Raphanus sativus l. BMC Genomics 23 (1), 548. doi: 10.1186/s12864-022-08782-4
Wang, P., Su, L., Gao, H., Jiang, X., Wu, X., Li, Y., et al. (2018a). Genome-wide characterization of bHLH genes in grape and analysis of their potential relevance to abiotic stress tolerance and secondary metabolite biosynthesis. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.00064
Wang, Y., Tang, H., Debarry, J. D., Tan, X., Li, J., Wang, X., et al. (2012). MCScanX: a toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 40 (7), e49. doi: 10.1093/nar/gkr1293
Wang, Y., Wang, S., Tian, Y., Wang, Q., Chen, S., Li, H., et al. (2021). Functional characterization of a sugar beet BvbHLH93 transcription factor in salt stress tolerance. Int. J. Mol. Sci. 22 (7), 3669. doi: 10.3390/ijms22073669
Wang, P., Wang, H., Wang, Y., Ren, F., Liu, W. (2018b). Analysis of bHLH genes from foxtail millet (Setaria italica) and their potential relevance to drought stress. PloS One 13 (11), e0207344. doi: 10.1371/journal.pone.0207344
Wang, D., Zhang, Y., Zhang, Z., Zhu, J., Yu, J. (2010). KaKs_Calculator 2.0: a toolkit incorporating gamma-series methods and sliding window strategies. Genom Proteom Bioinf. 8 (1), 77–80. doi: 10.1016/S1672-0229(10)60008-3
Wang, R., Zhao, P., Kong, N., Lu, R., Pei, Y., Huang, C., et al. (2018c). Genome-wide identification and characterization of the potato bHLH transcription factor family. Genes (Basel) 9 (1), 54. doi: 10.3390/genes9010054
Wei, K., Chen, H. (2018). Comparative functional genomics analysis of bHLH gene family in rice, maize, and wheat. BMC Plant Biol. 18 (1), 309. doi: 10.1186/s12870-018-1529-5
Wray, G. A., Hahn, M. W., Abouheif, E., Balhoff, J. P., Pizer, M., Rockman, M. V., et al. (2003). The evolution of transcriptional regulation in eukaryotes. Mol. Biol. Evol. 20 (9), 1377–1419. doi: 10.1093/molbev/msg140
Xie, T., Chen, C., Li, C., Liu, J., Liu, C., He, Y. (2018). Genome-wide investigation of WRKY gene family in pineapple: evolution and expression profiles during development and stress. BMC Genomics 19 (1), 490. doi: 10.1186/s12864-018-4880-x
Yang, J., Gao, M., Huang, L., Wang, Y., van Nocker, S., Wan, R., et al. (2017). Identification and expression analysis of the apple (Malus × domestica) basic helix-loop-helix transcription factor family. Sci. Rep. 7 (1), 28. doi: 10.1038/s41598-017-00040-y
Zhai, Y., Zhang, L., Xia, C., Fu, S., Zhao, G., Jia, J., et al. (2016). The wheat transcription factor, TabHLH39, improves tolerance to multiple abiotic stressors in transgenic plants. Biochem. Biophys. Res. Commun. 473 (4), 1321–1327. doi: 10.1016/j.bbrc.2016.04.071
Zhang, Z., Chen, J., Liang, C., Liu, F., Hou, X., Zou, X. (2020). Genome-wide identification and characterization of the bHLH transcription factor family in pepper (Capsicum annuum l.). Front. Genet. 11. doi: 10.3389/fgene.2020.570156
Zhang, C., Feng, R., Ma, R., Shen, Z., Cai, Z., Song, Z., et al. (2018a). Genome-wide analysis of basic helix-loop-helix superfamily members in peach. PloS One 13 (4), e0195974. doi: 10.1371/journal.pone.0195974
Zhang, T., Lv, W., Zhang, H., Ma, L., Li, P., Ge, L., et al. (2018b). Genome-wide analysis of the basic helix-Loop-Helix (bHLH) transcription factor family in maize. BMC Plant Biol. 16, 18(1):235. doi: 10.1186/s12870-018-1441-z
Keywords: abiotic stress, whole genome analysis, transcription factor, bHLH gene family, Chenopodium quinoa
Citation: Xue G, Fan Y, Zheng C, Yang H, Feng L, Chen X, Yang Y, Yao X, Weng W, Kong L, Liu C, Cheng J and Ruan J (2023) bHLH transcription factor family identification, phylogeny, and its response to abiotic stress in Chenopodium quinoa. Front. Plant Sci. 14:1171518. doi: 10.3389/fpls.2023.1171518
Received: 22 February 2023; Accepted: 21 April 2023;
Published: 05 July 2023.
Edited by:
Himabindu Kudapa, International Crops Research Institute for the Semi-Arid Tropics (ICRISAT), IndiaReviewed by:
Shengjun Feng, Zhejiang Agriculture and Forestry University, ChinaMuhammad Waseem, Hainan University, China
Qi Chen, Anhui Agricultural University, China
Copyright © 2023 Xue, Fan, Zheng, Yang, Feng, Chen, Yang, Yao, Weng, Kong, Liu, Cheng and Ruan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jingjun Ruan, jjruan@gzu.edu.cn