Assessment of the interactions between cetaceans and fisheries at the south of the Cetacean Migration Corridor and neighboring waters (Western Mediterranean)
 Mar Izquierdo-Serrano
Mar Izquierdo-Serrano  Ohiana Revuelta
Ohiana Revuelta  Raúl Míguez-Lozano
Raúl Míguez-Lozano  Patricia Gozalbes
Patricia Gozalbes  David Ruiz-García
David Ruiz-García  Juan Antonio Raga
Juan Antonio Raga  Jesús Tomás*
Jesús Tomás*- Marine Zoology Unit, Institute Cavanilles of Biodiversity and Evolutionary Biology, University of Valencia, Valencia, Spain
The Cetacean Migration Corridor is an important marine protected area for cetacean species in the Western Mediterranean, and part of its waters constitute the main fishing grounds for the Valencia region (East Spain) fishing fleet. Here we aim to assess the interactions between all active fisheries operating in the waters of the Valencia region and the cetacean species inhabiting this area. A total of 282 face-to-face interviews (51.37% of the total fleet) to fishers were held on 20 ports at the study area to gather information about cetacean by-catch and all types of interactions between fisheries and cetaceans. The interviewed bottom trawlers (n = 148 boats) reported a monthly by-catch rate (C) of 0.01 dolphins per vessel. Bottom trawling vessels operating at neritic zones only reported by-catch of common bottlenose dolphin (C = 0.009), while those fishing at oceanic zones by-caught striped dolphin (C = 0.006) and common bottlenose dolphin (C = 0.003). The interviewed artisanal fishers (n = 114 boats), the second most important fishery in the study area, only reported one dolphin by-catch event, but 90 of these boats communicated continued negative interactions with the common bottlenose dolphin, causing gear damage and catch take. The interviewed vessels gave an estimation of their annual economic loss produced by this interaction (mean ± SD: 2,998.10 ± 2095.02 € per boat). Other fisheries operating in the Valencia region were purse-seiners (n = 15 boats) and pelagic longlines, a fishery that has almost disappeared in the study area (n = 5 boats). The first one had the highest estimated dolphin by-catch rate (C= 0.04) and mainly interacted with common bottlenose dolphin, but also with striped dolphin. The information gathered from interviews was complimented with long-term stranding data (1990–2020). Despite the fact that only 7.80% of all recorded stranded cetaceans showed fisheries interaction signs, 26.72% of the common bottlenose dolphins recorded showed evidence of this interaction, thus confirming that it is the cetacean species most affected by fisheries in the area. Records of stranded striped dolphin (6.45%) also showed evidence of fisheries interaction. Based on both dolphin by-catch and gear damage, management plans are needed in the area to ensure cetacean conservation and also fisheries sustainability in waters inside and around marine protected areas at the Spain’s Mediterranean.
1. Introduction
The Western Mediterranean basin hosts up to eight resident cetacean species. The striped dolphin (Stenella coeruleoalba) is the most abundant species in the area, but common bottlenose dolphin (Tursiops truncatus), short-beaked common dolphin (Delphinus delphis), Risso’s dolphin (Grampus griseus), long-finned pilot whale (Globicephala melas), fin whale (Balaenoptera physalus), sperm whale (Physeter macrocephalus), and Cuviers’ beaked whale (Ziphius cavirostris) also have stable populations in the area (Gómez de Segura et al., 2006; Boisseau et al., 2010). Moreover, other vagrant species have been reported in this area, such as the minke whale (Balaenoptera acutorostrata; Fraija-Fernández et al., 2015, and references therein) or the humpback whale (Megaptera novaengliae; Violi et al., 2021) among others. The area between East Iberian Peninsula and Balearic Islands harbors the Cetacean Migration Corridor, a Specially Protected Area of Mediterranean Importance (SPAMI). This marine protected area (MPA) covers 46.385 km2 and constitutes a key area for cetacean conservation in the Western Mediterranean. Active hydrocarbon prospections and extractive activity are banned at the demarcation of this MPA, except those related to research under permit (OceanCare, 2021). Despite the existence of this MPA, cetaceans still face several threats within the area, such as fisheries interaction. Thus, it is necessary to identify and quantify them to improve the effectiveness of this MPA for helping cetacean species conservation.
Anthropogenic threats affecting cetaceans worldwide include habitat degradation, many types of pollution, acoustic disturbances, marine traffic, and the incidental interaction with fisheries, among others (Nortarbartolo di Sciara, 2016; Nelms et al., 2021, and references therein). Fisheries by-catch is a persistent threat for cetaceans in many seas, with severe impacts on the health and viability of many populations. Cetaceans and fishing fleets usually share niche, and their distribution and fishing activities overlap geographically, entailing negative consequences for cetaceans (Avila et al., 2018; Carpentieri et al., 2021; Moore et al., 2021).
Interactions between cetaceans and different types of fisheries have been described in many places (Lewison et al., 2014). Fishery discards constitute an important food source for cetaceans, attracting them to fishing grounds (Bonizzoni et al., 2022). In areas such as the Atlantic Ocean, bottom trawling entails a high number of cetacean accidental catches (López et al., 2003; Fernández-Contreras et al., 2010). However, absent or low by-catch rates have been reported in the Mediterranean Sea (Gonzalvo et al., 2008; Fortuna et al., 2010). On the other hand, depredation events on artisanal fisheries by dolphins occur frequently, causing damage to nets and to commercial captures (Snape et al., 2018), although low by-catch rates on this fishery have also been reported (Lauriano et al., 2004; Díaz López, 2006; Brotons et al., 2008). Conversely, pelagic longline and purse-seine are fisheries with high levels of cetacean by-catch reported, particularly of small delphinid species, in the Western Mediterranean (Aguilar, 1991; Zahri et al., 2007; Macías-López et al., 2012).
The Valencia region (East Spain, Western Mediterranean) has a big fishing fleet that operates in waters inside and neighboring the Cetacean Migration Corridor SPAMI. This fishing fleet includes bottom trawling, artisanal (using several fishing gears), purse-seine, and pelagic longline vessels (Generalitat Valenciana, 2020). Despite the existing overlapping between fishing grounds and cetacean distribution and habitat use in these waters, there is scarce information about their interactions. To date, only one study described fisheries interactions between one species of cetaceans and artisanal vessels in the area (Revuelta et al., 2018).
Face-to-face interviews to fishers are considered a useful and cost-effective tool for identifying specific problematic interactions between fisheries and threatened marine species, although the results obtained with this methodology are highly dependent on the fishers’ reliability (Moore et al., 2010; Goetz et al., 2014). Data from interviews allow to estimate the minimum cetacean by-catch rates in a specific area (López et al., 2003). Prior studies across the Western Spanish Mediterranean denote the importance of such sources of information to analyze marine megafauna by-catch rates and the associated economic loss relative to such interactions (Carreras et al., 2004; Domènech et al., 2015; Revuelta et al., 2018).
Stranding data, although subject to several legal restrictions that constrain access to the animals and also to other caveats, when recorded over long periods and large areas, provide valuable information and have been, for long, a valuable source to determine the species’ population status. Considering that working with marine protected species with ocean-wide distribution is often costly and logistically difficult, stranding networks are crucial to evidence the threats to which marine megafauna is exposed, including fisheries interactions (e.g., Leeney et al., 2008; Tomás et al., 2008; Casale et al., 2010). The postmortem examination of stranded animals can unveil the relative mortality consequence of by-catch interactions for both resident and migratory cetacean species (Puig-Lozano et al., 2020; Duras et al., 2021; Peltier et al., 2021). Combining different sources of information, such as face-to-face interviews to fishers and long-term stranding data, can contribute to the obtainment of an accurate assessment of the magnitude at which cetaceans are exposed to fisheries interactions in a specific area.
Here we combine detailed interviews conducted in the fishing ports of the Valencia region with long-term stranding data to gain knowledge about the problem between local fisheries and cetaceans. The specific objectives of the present study are (1) to provide an assessment of the interaction between fisheries operating throughout the Valencia region (East Spain) and the inhabiting cetacean species, and (2) to determine specific areas of cetacean–fisheries interaction in waters inside and neighboring the Cetacean Migration Corridor MPA.
2. Materials and methods
2.1 Study area
The Valencia region coastline extends over 419 km (37°51′ N, 0°45′ W; 40°31′ N, 0°31′ E) across East Spain (Western Mediterranean). The coastal waters are not homogeneous, with a narrower continental shelf at the southernmost province (Alicante) that expands over the central province (Valencia) and is widest at the northernmost province (Castellón, Figure 1). The waters of the Valencia region are included within the Geographical Subarea n°6 from the General Fisheries Commission for the Mediterranean (GFCM GSA06) and comprises part of the Cetacean Migration Corridor MPA. The present study covered 20 ports distributed throughout the three provinces of the Valencia region to provide a heterogeneous sample and analyze potential differences in cetacean–fisheries interactions between provinces.
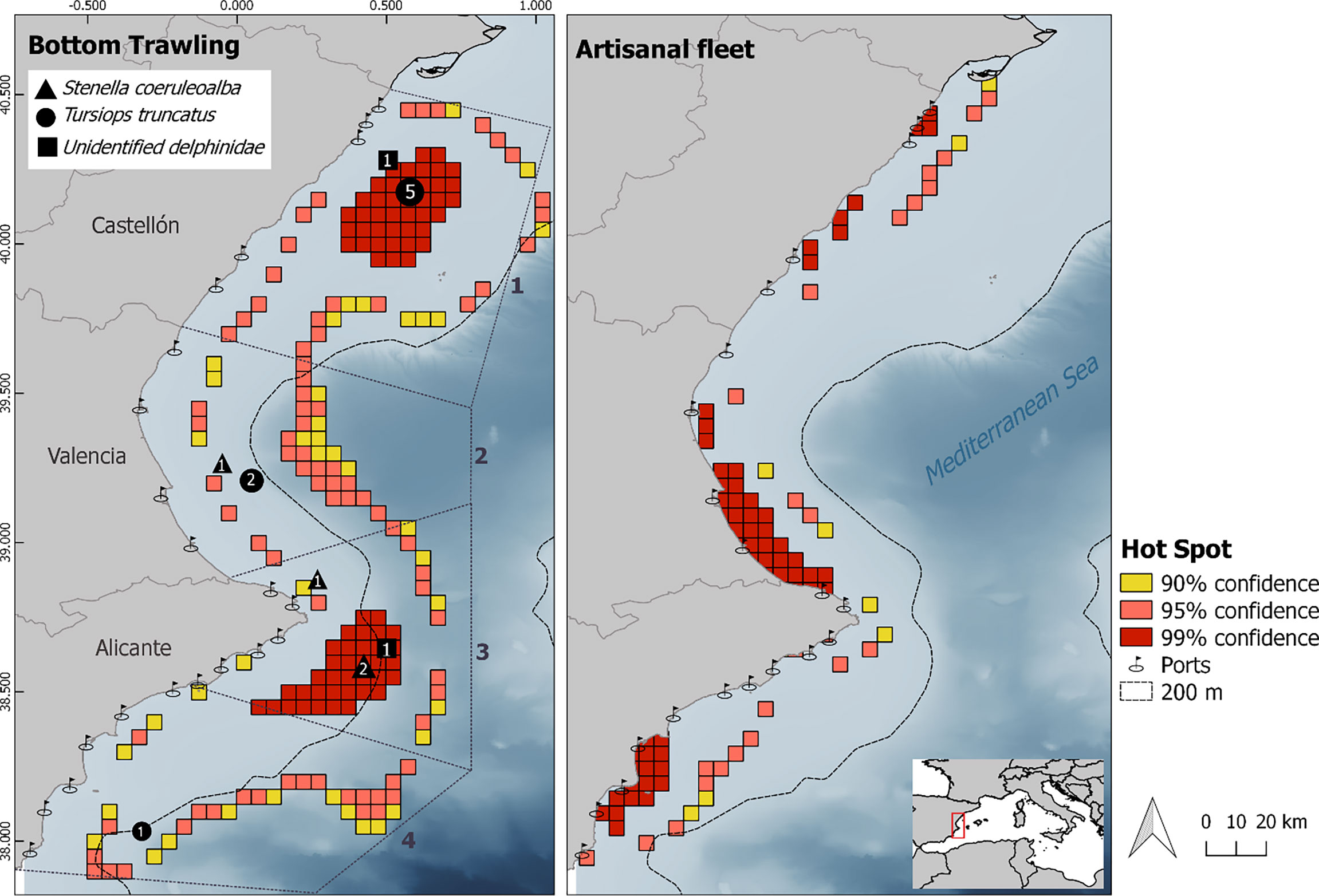
Figure 1 Map of the study area (Valencia region, Western Mediterranean) showing the spatial clusters of fishing activity (hot spots) of bottom trawling (left) and artisanal fleets (right) during the interview survey period. Cold spots are not represented. The black symbols show the number of delphinid by-catch events reported by the interviewed bottom trawl vessels. The map includes the distribution of 20 surveyed ports in the three provinces. The bathymetry is depicted with darker shades of blue representing deeper waters and dashed light gray line of the 200-m bathymetric contour. The stratification of the study area in the subzones stablished for bottom trawling fishery is also shown in the map on the left.
2.2 Fishing fleet description
The bottom trawling vessels operating in the study area have a mean vessel length of 21.25 ± 3.41 m (range: 16.00–29.00 m) (Ministry of Agriculture, Fisheries and Food, 2020). This fishery has many demersal target species, such as European hake (Merluccius merluccius), red mullet (Mullus barbatus), monkfish (Lophius spp.), and Norway lobster (Nephrops norvegicus) (Sala et al., 2019). The Valencian bottom trawling fishing fleet is constituted by 209 vessels (Generalitat Valenciana, 2020), differenciating those fishing in coastal areas all year round from those targeting red shrimp, Aristeus antennatus, at deeper waters (400-800 m) at some periods of the year, similarly as described in the adjacent areas of Spain’s Mediterranean (Carbonell et al., 1999). This fleet discards a long list of species [see Carbonell et al. (1998)] that can potentially be used as a food resource by marine vertebrates (e.g., Tomás et al., 2001).
Artisanal vessels fish at less than 12 nautical miles from the coast, have a mean vessel length of 10.10 ± 2.00 m (range: 6.00–15.00 m) (Ministry of Agriculture, Fisheries and Food, 2020), and use different gears, mainly trammel nets and also gillnets, pots, and demersal longlines. A total of 294 artisanal vessels of this fleet are active (Generalitat Valenciana, 2020) and are distributed along all fishing ports of the study area. Only vessels using trammel nets and gillnets were included in the present study since they are the most commonly used fishing gears in this fleet and the ones previously described as interacting with cetaceans. The principal target species are common cuttlefish (Sepia officinalis), common sole (Solea solea), and red mullet (Mullus spp.) for trammel nets and Atlantic bonito (Sarda sarda), gilt-head bream (Sparus aurata), and white seabream (Diplodus sargus) for gillnets (Revuelta et al., 2018).
Purse-seine vessels are at least 11 m in length, operate at night in the continental shelf, and target basically European pilchard (Sardina pilchardus) and European anchovy (Engraulis encrasicolus) (Boubekri et al., 2019). This fishery only has 38 active vessels according to the census of the Generalitat Valenciana (2020). In the Valencia region, pelagic longline is also a minoritarian fishery (n = 5 boats) (Generalitat Valenciana, 2020). The pelagic longline boats have variable ranges of vessel length (12–27 m), and its fishing grounds, as purse-seines, cover a big area depending on fish school locations. Pelagic longliners in the Western Mediterranean target swordfish (Xiphias gladius), bluefin tuna (Thunnus thynnus), and albacore (Thunnus alalunga) (Macías-López et al., 2012).
2.3 Interviews to fishers
Interviews were carried out during October–December 2020 at 20 out of 21 ports with fishing vessels at the study area by a trained team of the University of Valencia. Despite the fact that the interviews were conducted after the COVID-19 lockdown situation, the Spanish government considered fishing as an essential activity; hence, fishing activity practically did not stop the year before the interviews. Then, we can assume that the COVID-19 situation did not affect the interview campaign and the results of the present study in the Valencia region. Fishers’ associations and port authorities of every port were contacted before conducting the interviews. The interview campaign days were maximized as well as adjusted to the routines of each fishery (Goetz et al., 2014). The interviews were made by large to more than 20% of the active vessels of the two main fisheries (bottom trawling and artisanal) in the study area, following the precedence of other studies in the area [Revuelta et al. (2018) and references therein]. However, in a few ports, the required 20% was narrowly missed: artisanals in Vinaroz and Santa Pola ports and bottom trawlers in Sagunto and Torrevieja ports (Table 1). One questionnaire session was held per boat, with the skipper being preferably the person interviewee, and the interview duration lasted approximately 20 min.
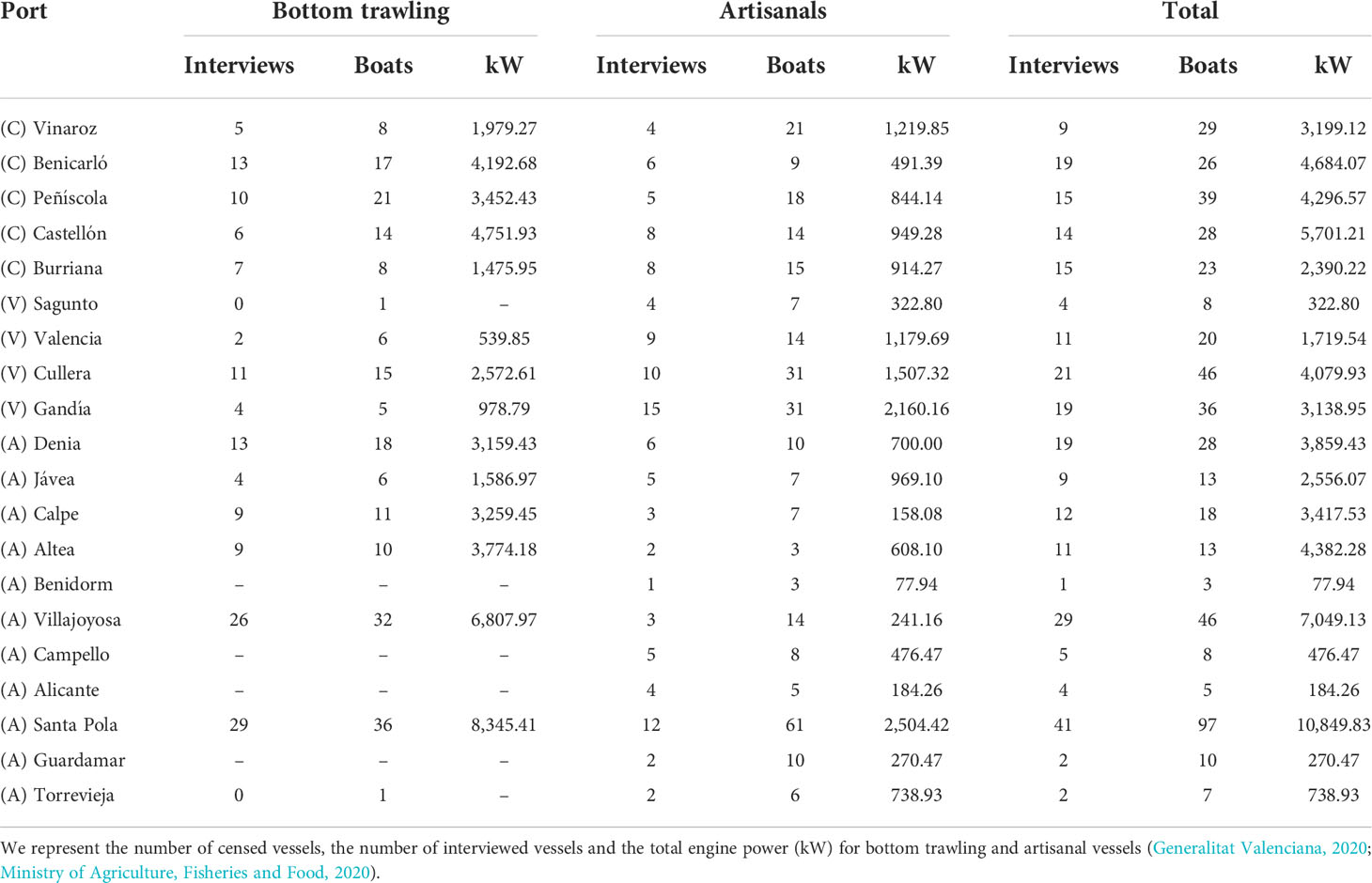
Table 1 We represent the number of censed vessels, the number of interviewed vessels and the total engine power (kW) for bottom trawling and artisanal vessels (Generalitat Valenciana, 2020; Ministry of Agriculture, Fisheries and Food, 2020). We present the number of censed vessels, the number of interviewed vessels, and the total engine power (kW) for bottom trawling and artisanal vessels (Generalitat Valenciana, 2020; Ministry of Agriculture, Fisheries and Food, 2020).
Before the interview was started, the fishers were informed about the confidentiality of their answers and that the personal and professional information asked would be used only for this study. The fishers were asked about their hierarchical level in the crew, number of crew members, fishing gear used (in case of boats using different gears in different periods), and fishing characteristics (vessel length, minimum and maximum depth of fishing operations, five most common target species, spatial information about their fishing activities, and number of months of fishing activity in the previous year to the interview). We also gathered information about gear damage caused by dolphins, estimation of their respective annual economic loss due to this interaction, and cetacean by-catch events in that period. Finally, the interviewees were asked about their perception of cetacean population trends at their fishing grounds. The interviews were conducted with the support of photographs of cetacean species with a higher probability of interacting with fisheries in the area (common bottlenose dolphin, striped dolphin, common short-beaked dolphin, Risso’s dolphin, and long-finned pilot whale) to gather accurate information of interacting species (Moore et al., 2010; Revuelta et al., 2018). Only fully completed interviews were considered for further analysis.
2.4 Data analysis
2.4.1 By-catch
By-catch, defined as the unintended capture of marine biota in a fishery targeting different species (Gray and Kennelly, 2018), was related with bottom trawlers’ mean fishing depth (m). Mean fishing depth (m) per vessel was calculated using the maximum and minimum depth (m) reported by each interviewee. The waters of the study area where bottom trawlers operate were divided into two marine zones based on bathymetry, neritic (≤200-m depth) and oceanic (>200-m depth) (Hedgpeth, 1957). To analyze differences in the operations of bottom trawlers, we followed Domènech et al. (2015), stratifying information gathered by bottom trawlers in four fishing subareas according to reported fishing zones, the previously stated bathymetrical parameters, trawl fishing depth, and port distribution in these areas (see Figure 1).
Cetacean by-catch rate (C) was determined by dividing the reported annual captures of cetaceans (Cobs) by the reported fishing effort of a fishery (Fobs), which is the total number of reported months of fishing activity in the previous year to the interview. Annual cetacean by-catch (Cest) was estimated by multiplying C by the estimated fishing effort for a fishery (Fest).
Estimated fishing effort (Fest) was extrapolated by multiplying the number of total active censed vessels of a fishery by Fobs, which was respectively divided by the number of interviewed vessels.
Maximum and minimum 95% confidence intervals (95% CI) were obtained for annual cetacean by-catch estimation (Cest) following Greenwood (1996). Cetacean by-catch rates and annual cetacean by-catch estimations were also obtained for bottom trawlers and purse-seiners for those reported species, common bottlenose dolphin and striped dolphin. Additional cetacean by-catch rates were also given for bottom trawlers according to fishing bathymetric zones (Neritic/Oceanic) and fishing subareas (1-4). As artisanal and pelagic longline vessels only reported one capture of cetaceans each, descriptive information was given for both fisheries.
2.4.2 Gear damage: Economic loss in artisanal vessels
In order to evaluate the economic loss produced by cetaceans reported by interviewed artisanal vessels, an ANOVA test (Mangiafico, 2015) was performed to analyze differences between the three provinces. A Spearman correlation test was used to study the correlation between the reported economic loss and the number of months that the artisanal vessels used nets on the previous year to the interviews campaign. A chi-square test was used to compare the artisanal fishers’ perception on possible variation in dolphin abundance at their with the one of fishers from other fisheries. In order to establish areas of higher dolphin–fisheries interaction, we considered engine power (kW) of the active artisanal vessels at the 20 surveyed ports, gathered from the Ministry of Agriculture, Fisheries and Food (2020), as a proxy of fishing capacity (Crosti et al., 2017). Then, we related engine power with annual economic loss reported by fishers in each port. Significance for these tests was stablished at α= 0.05. Statistical analysis was performed using R 4.0.5.
2.4.3 Spatial analysis of bottom trawling and artisanal fleet
The use of fishers’ knowledge through geographical information systems (GISs) allows describing spatially accumulation of fishing activities and fishing grounds (Léopold et al., 2014; Aylesworth et al., 2017). Spatial information about the location of fishing activities was collected during interviews using a base map with a grid of 5 × 5 km cells, including bathymetry, relevant points of reference and localities names. Fishers were asked to draw on the map where they normally fish and each fishing polygon was digitized into GIS vector polygons. Aggregate fishing effort density per cell was expressed as the number of vessels identifying the cell as a fishing ground.
Hot Spot Analysis tool (Getis-Ord Gi*) was used to describe and to map statistically significant spatial clusters for the two main fisheries in the region (bottom trawling and artisanal fleets).
The Getis-Ord Gi∗ statistic (Getis and Ord, 1992) determines the spatial clustering of grid cell values that are higher (hot spot) than expected by random distribution. It performs significant tests between nearby cells in the surrounding neighborhood area using a z-score (Getis and Ord, 1992). The resultant z-scores and p-values indicate where features with either high or low values cluster spatially. We considered as hot spot the areas where cells with high z-score and low p-value were spatially clustered.
Distributional maps were created at three levels of confidence (99, 95, and 90%), and all clusters that were within the 99% confidence level were considered for displaying the more intensely hot clusters. All data analysis and a georeferenced distribution map of each fishery fleet were generated using the free open-source Geographic Information System program QGIS, version 3.4.6 (QGIS, 2021).
2.5 Analysis of cetacean strandings
2.5.1 Strandings with evidence of fisheries interaction
We compiled the cases of stranded cetaceans found along the Valencia region coast between 1990 and 2020 displaying fisheries interaction signs. Such stranding events were registered by the cetacean and marine turtle stranding network of the Valencian community that follows the protocol explained in Gozalbes et al. (2010). This network records dead or injured cetaceans and sea turtles stranded on beaches or found floating dead or in a weakened condition. It is coordinated via a 24-h telephone hotline by the Marine Zoology Unit of the University of Valencia (MZU-UV). Since a diagnostic necropsy was not performed in all recorded stranded cetaceans, only stranding cases with available photographs for visual external examination and/or those necropsied or directly examined by the personnel of the MZU-UV were considered to establish fisheries interactions.
The external signs of fisheries interactions in stranded cetaceans considered here were (1) entanglement in fishing gear still attached to the animal in the stranding or by-catch events directly reported by fishers, (2) net marks and superficial skin lesions clearly caused by fishing gear, (3) jaw/skull fracture and broken teeth (these kind of injuries are produced when fishers, particularly trawlers, drop the captures from the net over the ship deck), (4) amputations of different parts of the body with a human cause (e.g., strangulation of flippers by nets), (5) incisions/cuts into the body cavity or over the skin of clear anthropogenic origin, (6) long-term tail entanglement, and (7) remains of fishing gear inside the body cavity and/or the digestive tracts (Duras et al., 2021) (see the examples in Supplementary Figure S1A).
We analyzed trends of common bottlenose dolphin and striped dolphin strandings with fisheries interaction signs (the only two cetacean species with a sufficient sample size) over time through regression analysis. We used a linear regression t-test to determine whether the slope of the regression line differs significantly from zero. We explored these trends in two periods—a three-decade study period (1990–2020) and in the last decade (2009–2020)—to gather long-term and recent trends for these two species. We excluded data from the years 1990 and 2007 for striped dolphin since massive stranding of the species occurred in these years due to a Morbillivirus epizootic (Duignan et al., 1992; Raga et al., 2008).
2.5.2 Spatial analysis of stranding data
Kernel density maps were produced to describe the spatial distribution of strandings of common bottlenose dolphins and striped dolphins throughout the historical records in the Valencia region coasts between 1990 and 2020. This was only performed for these two species because they are the most abundant cetaceans in the area (Gómez de Segura et al., 2004; Gómez de Segura et al., 2006) and because both account for the vast majority of cetacean interactions reported during the interviews.
The kernel density tool calculates the magnitude per unit of area from point features using the kernel function to produce a more generalized density raster. This allows a visual representation of the density of strandings in an area by creating “hot spots”. This required the creation of point shapefiles from geographic coordinates of strandings as an input layer to generate a kernel density estimation (KDE) representing strandings per square kilometer.
Core-stranding areas were identified using fixed kernel density estimations (Worton, 1989) in QGIS version 3.4.6 Geographic Information System Software (QGIS Development Team, 2021). A rule-based ad hoc method was applied to estimate the appropriate smoothing parameter (h) for delineating kernel contours (Kie, 2013). We used 90% KDE to estimate the overall stranding range and 50% KDE to represent the core area of dolphin species’ strandings.
Since strandings have been previously related to fishing capacity (Byrd et al., 2014; Crosti et al., 2017), we mapped the engine power (kW) of the 21 fishing ports of the Valencia region to visually determine overlapping and assess the spatial relationships of common bottlenose dolphin and striped dolphin stranding (50% KDE) distributions and engine power as a measure of fishing capacity. We represented the proportion of engine power at which each port contributes to the total engine power of the fishing fleet from the study area in Figure 2.
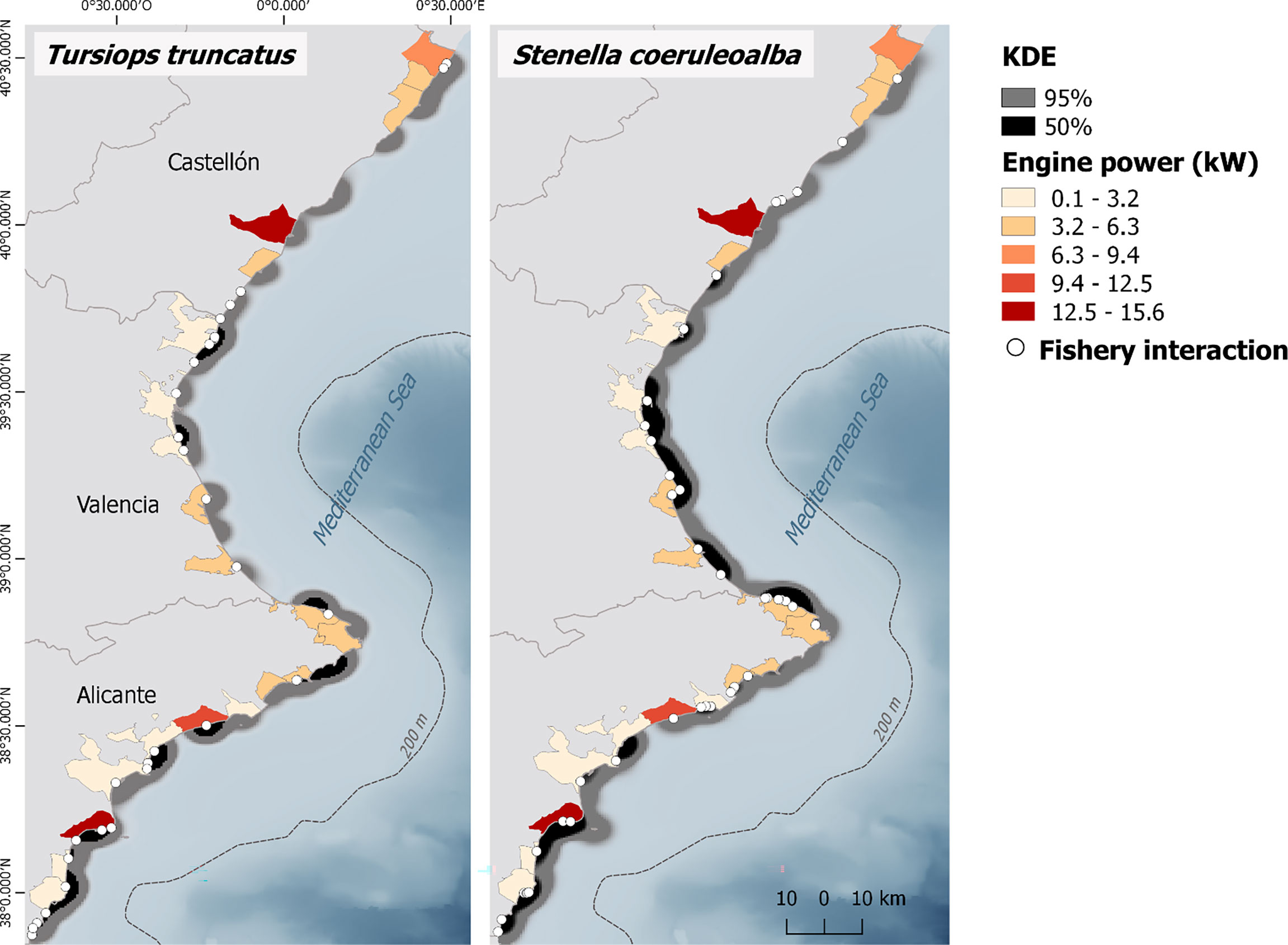
Figure 2 Kernel density estimations (KDE) of Tursiops truncatus (left) and Stenella coeruleoalba (right) stranding events from records of the Valencia region stranding network for the period 1990-2020, according to 5 × 5-km grid cells. The core stranding areas of both species (50% KDE) are represented. The legend inside the figure also shows the ports in five strata representing the proportions of engine power (kW) of each fishing port in relation to the total engine power of the Valencia region fishing fleet. Colored areas represent the municipality of the port. White circles show stranded individuals recorded with fishery interaction signs. Additional details of the study area are shown in Figure 1.
3. Results
3.1 Interview survey
A total of 282 interviews (one per boat) were held during the campaign on 20 out of the 21 ports of the studied area; thus, according to the 2020 census (Generalitat Valenciana, 2020), we surveyed 51.37% of the active censed fleet. The fishers reported 24 accidental catches of small cetaceans, all from the family Delphinidae. There were significant differences regarding by-catch and gear damage events reported among the different gears; therefore, the results per fishery are given below.
3.1.1 Bottom trawling: Fishing ground distribution and cetacean by-catch
The distribution map of the bottom trawling fishing grounds created from the information provided by fishers during interviews (Figure 1) was consistent with the map derived from the VMS data provided by the Spanish Government and North East Atlantic Fisheries Commission (ICES, 2008). The data show that bottom trawlers from the Valencia region ports operated at a mean ± SD depth of 168.30 ± 187. 25 m (range = 20.21–1,203.50 m). For the surveyed bottom trawlers (n = 148), there were significant differences between the mean fishing depth (m) among the four stratified subareas (ANOVA, F = 6.241, p< 0.001). Trawlers operating at subarea 3 (mean ± SD: 286.90 ± 223.30 m) and subarea 4 (mean ± SD = 298.70 ± 185.70 m) fished deeper than the ones in subarea 1 (mean ± SD= 141.74 ± 169.25 m). Trawlers operating at subarea 2 fished at a mean ± SD fishing depth of 249.58± 145.03 m, which did not show significant differences with any of the other subareas.
Based on the Getis-Ord Gi* statistic, significant clusters of fishing activity were identified in the waters of the Valencia region. Those higher z-score values defined as hot spots are shown in Figure 1. A hot spot area was found at the northern part of the study area located in the waters over the continental shelf near the 200-m isobaths. A second large hot spot area was detected at the south of the study area, showing that bottom trawling fishing grounds concentrate at both neritic and oceanic zones depending on the area.
Overall, the bottom trawl interviewees reported a by-catch of 16 dolphins for the period October 2019–November 2020, which meant a by-catch rate of 0.01 dolphins captured monthly per vessel. Extrapolating to the entire active fleet of this fishery, this meant an annual cetacean by-catch estimation in the Valencia region of 23 delphinids (95% CI = 7–39). The reported species composition was nine common bottlenose dolphins, four striped dolphins, and three unidentified dolphins by-caught, with differences in the captured species between the neritic and oceanic zones (Table 2). The common bottlenose dolphin by-catch rate was higher in vessels working in the neritic zone (C = 0.009) than in the oceanic zone (C = 0.003). The striped dolphin by-catch events were only reported by fishers operating in the oceanic zone (C = 0.006). There were also differences in by-catch rates between the bottom trawl fishing subareas as the common bottlenose dolphin by-catch rate was higher in subarea 1 (C = 0.012) and subarea 2 (C = 0.018), while for the striped dolphin by-catch rate, it was higher in subareas 2 and 3 (C = 0.006) (Table 2). On this fishery, no substantial gear damage due to dolphin feeding interactions was informed by the interviewees and did not represent a substantial economic loss for this fishery.

Table 2 By-catch of Tursiops truncatus and Stenella coeruleoalba reported by bottom trawling vessels according to depth zones (neritic/oceanic), fishing subareas (1–4), and totally. The summary of total reported by-catch individuals, by-catch rates, and annual cetacean by-catch estimations with its respective 95% confidence intervals (95% CI) is shown per species. We also present the bottom trawling interviewed vessels, censed vessels, and the estimated fishing effort (Fest) for each stratum and totally.
3.1.2 Artisanal fleet fishing ground distribution and interactions with cetaceans
A total of 114 interviews were made to artisanal fishers (one per vessel) and were used to map the individual areas of artisanal fishing grounds. The spatial distribution of the interviewed artisanal vessels shows a prevalence of exploitation of the neritic zone. In fact, this fishery showed a mean fishing depth of 34.86 ± 18.80 m (8.52–140.00 m). Figure 1 shows the spatial cluster distribution using the Getis-Ord Gi* and those fishing grounds showing a higher density of artisanal vessels at the study. The larger areas identified by statistically significant Gi* showed the importance of a variety of fishing zones along the coast. The higher z-score values, in red, correspond to the area between Alicante and Santa Pola ports, the area between Denia and Cullera ports, and finally the area around the three northernmost ports (Peñíscola, Vinaróz, and Benicarló). Other hot spots were also evident around the port of Castellón.
Only one by-catch event of a dolphin that the interviewed fisher did not identify at a species level was reported. However, a total of 103 (90.35%) of the interviewed artisanal fishers reported negative interactions with dolphins. In all cases, common bottlenose dolphin was identified as the species causing this interaction. The fishers reported that dolphins cause economic loss by taking or damaging the target species in nets and/or captures and damaging their fishing nets. From all artisanal fishers who reported a negative interaction with dolphins, 90 (87.38%) gave an estimation of their annual economic loss. The mean annual economic loss reported per artisanal vessel was 2,998.10 ± 2,095.02 € (range: 100–9,000 €). Economic loss was significantly different between the three provinces (ANOVA, F = 3.205, p< 0.05), the lowest mean ± SD being reported at Castellón province (1,716.67 ± 1,617.17 € per vessel) and the highest at Valencia province (3,050.00 ± 2,246.89 € per vessel). This province has ports with high artisanal engine power, such as Gandía, Cullera, and Valencia city (total for Valencia province: 5,169.97 kW) (Table 1). The reported economic loss was also found to be positively correlated (r2 = 0.24) with the number of months that the artisanal fishers used nets (Spearman, S = 187,987, p< 0.05).
3.1.3 Interactions between cetaceans and other fisheries operating in the region
The purse-seine fishers interviewed (n = 15 boats) reported four common bottlenose dolphins and two striped dolphins in by-catch events, which supposed a by-catch rate of 0.04 dolphins accidentally caught monthly per vessel. The estimated annual delphinid by-catch for this fishery was 15 dolphin individuals (95% CI: 3–26) for the entire purse-seine fleet. The common bottlenose dolphin by-catch rate was 0.026, and the estimation of the annual by-catch was 10 individuals. The striped dolphin by-catch rate was 0.013, and the estimation of annual by-catch for this species was five individuals. We were able to gather scarce information about mortality due to by-catch for bottom trawling and artisanal and pelagic longline fisheries, although inconsistently. However, all purse-seine fishers interviewed did provide this information. The fishers reported that 33% of cetaceans accidentally caught by this fishery were released alive, while the rest were discharged already dead from nets. Regarding the purse-seiners’ gear damage produced by dolphins, four out of 15 interviewees (26.67%) reported sporadic damage on their fishing gear produced by striped dolphin and common bottlenose dolphin, with an associated economic loss (mean ± SD: 9700.00 ± 7596.05 € per vessel). In addition, negative interactions due to depredation by other species, such as tuna species, were reported during the interview campaign.
The pelagic longline vessels interviewed (n = 5) operate throughout the study area waters in a mean fishing depth of 201.20 ± 62.70 m (range: 101.86–283.36 m). One pelagic longline vessel reported one accidental capture of a Risso’s dolphin, with no gear damage produced by cetaceans at this fishery according to interviewees.
3.1.4 Cetacean population trends according to fishers
In the context of all fisheries, 42.91% of fishers perceived that the cetacean population abundance remains stable at their fishing grounds, while 44.70% assured that the presence of dolphin species has increased. The proportion of artisanal fishers perceiving an increase of dolphin population at their fishing grounds (63.55%) was significantly higher than the one for bottom trawling fishers (33.78%) (chi-square test, χ2 = 32.66, p< 0.001), purse-seines (13.33%) (chi-square test, χ2 = 74. 71, p< 0. 001), and pelagic longlines (20.00%) (chi-square test, χ2 = 41. 40, p< 0.001).
3.2 Cetacean stranding data in the Valencia region
Regarding the information gathered from the stranding network of the Valencia region, a sum of 1,411 cetaceans were stranded at the Valencia region coast between 1990 and 2020. The percentage of stranded cetaceans showing fisheries interactions evidence on the study area and period reaches 7.80% of the total number of cetacean stranding records (Table 3).
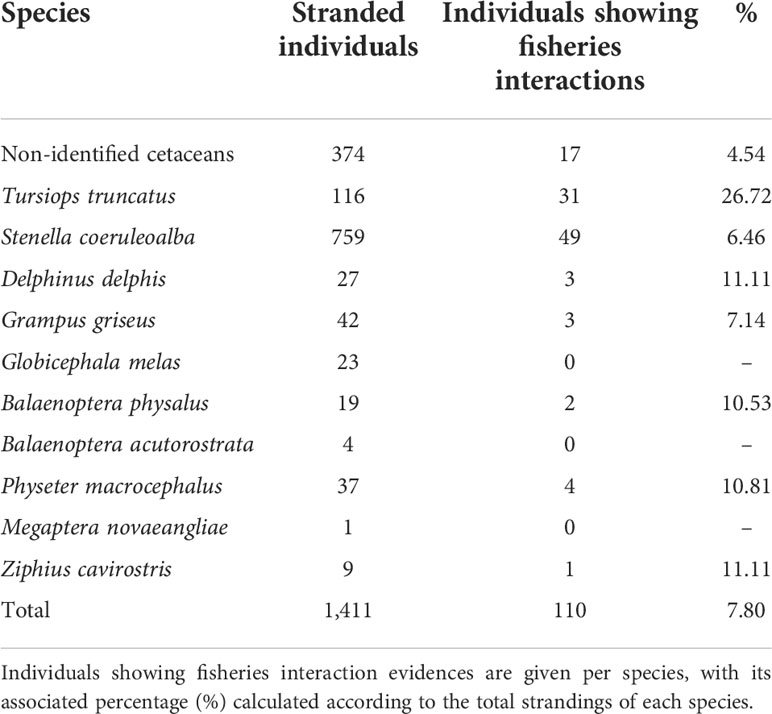
Table 3 Number of cetacean individuals stranded on the Valencian Community coast between 1990 and 2020. Individuals showing fisheries’ interaction evidence are given per species, with its associated percentage (%) calculated according to the total strandings of each species.
3.2.1 Evidences of fisheries interactions in stranded cetaceans
Between 1990 and 2020, a sum of 110 cetaceans recorded by the stranding network of the Valencia region, either stranded or bycaught, showed different evidence of fisheries interactions. Fourteen of them were recorded entangled in fishing gears. One common bottlenose dolphin, one striped dolphin, and two unidentified cetaceans were reported to the network after being having been captured by bottom trawlers. One common bottlenose dolphin, two striped dolphins, and one Risso’s dolphin were reported after being entangled in the nets of artisanal vessels. One fin whale and one Cuviers’ beaked whale were found entangled in drift nets, and one common short-beaked common dolphin was recorded after being entangled in a longline fishing gear. Three unidentified delphinids were also recorded as by-caught in fishing gears not specified in the records of the network.
3.2.2 Common bottlenose dolphin and striped dolphin strandings’ density distribution
Striped dolphin and common bottlenose dolphin species both show a higher number of stranding records at the study area. From 1990 to 2020, the study period, stranding reports were filed for 166 common bottlenose dolphins and 759 striped dolphins throughout the Valencia region coastline. The mean annual common bottlenose dolphin stranding records during the study period was 5.35 ± 3.20, ranging from 0 in 1999 to 14 in 2003. A positive trend in strandings with fisheries interactions signs for the species was detected for the 1990–2020 period (t-test for regression slope, t = 4.295, p< 0.001) and also for the 2009–2020 period (t = 3.883, p< 0.01). Common bottlenose dolphin was the species that showed the highest percentage of strandings with fisheries interactions signs (Table 3).
For the striped dolphin, the mean annual stranding records during the study period was 24.50 ± 28.13, ranging from 6 in 1996 to 154 in 1990. For this species, a positive trend in strandings with fisheries interaction signs was found during the whole studied period (t = 3.355, p< 0.01), but not for the 2009–2020 period (t = 0.968, p> 0.05). Figure 2 shows the locations of the stranding records of individuals of these two species showing evidences of fisheries interactions from 1990 to 2020.
The spatial distribution of common bottlenose dolphin strandings in the region estimated by KDE indicated that the highest stranding densities occurred along the Alicante coastline (Figure 2). The estimates also showed two isolated clusters in the Valencia province, around Sagunto and at the south of Valencia city. There was no substantial stranding density for common bottlenose dolphins in the Castellón province (Figure 2).
The spatial distribution of striped dolphins stranded in the Valencia region coasts also reveals the lowest stranding density along the Castellón province coastline for this species (Figure 2). The highest stranding densities occurred between the ports of Valencia, Cullera, and Gandia. The estimates also showed a small concentration of strandings at the north of the Valencia province and at the southernmost port of the Castellón province. Another area of concentration of strandings was located at the south of the Alicante province, between the ports of Alicante and Torrevieja, and an isolated cluster around El Campello port (Figure 2).
On the other hand, the distribution of engine power contribution of the active censed fleet shows a concentration of ports with higher contribution at the south of the study area (Figure 2), as only Santa Pola and Villajoyosa ports accumulate 28.23% of the engine power of the Valencia region fishing fleet (Ministry of Agriculture, Fisheries and Food, 2020) (Table 1, Figure 2). This concentration of engine power at the south of the region seems to be associated with common bottlenose dolphin and striped dolphin stranding distribution there. On the contrary, some ports at Castellón province highly contributing to the total engine power of the study area seem not to be associated with the strandings of both the common bottlenose dolphin and the striped dolphin due to the absence of KDE core-stranding areas for these species. The lack of association between fishing capacity and stranding distribution can also be seen at ports found at the Valencia province, the province with the lowest fishing capacity contribution value, but with KDE core-stranding areas, especially for striped dolphin.
4. Discussion
4. 1 Problem of cetacean–fisheries interactions according to fishers
Despite the diversity of cetaceans in the Western Mediterranean waters and particularly in the Valencia region (Gozalbes et al., 2010), according to fishers’ perception, only two species exhibit an important interaction with fisheries in this area, the common bottlenose dolphins and the striped dolphins. Both species were defined as “Vulnerable” in the Mediterranean Sea, and fisheries interactions were already defined as a threat for them, especially for the common bottlenose dolphin (IUCN, 2021). This interaction seems to be negative in a twofold way. Firstly, fisheries may cause the accidental capture of these species, either because both share the same area or because dolphins may be attracted to fishing grounds by captures in nets or fishing discards, although the by-catch rates seem to be lower in the Valencia region waters compared to other areas (López et al., 2003). Secondly, when predating on captures, dolphins can produce economical loss by reducing the captures of target species and by damaging fishing gears during depredation events, thus increasing the fishers’ negative perception of these threatened species. Recent protection figures have been created to protect cetacean species in Spain’s Mediterranean waters, such as the Cetacean Migration Corridor (OceanCare, 2021). However, this large marine protected area mainly comprises open waters; thus, little protection may be offered to dolphins inhabiting or migrating throughout the coastal waters. Despite the existence of several marine protected areas in waters over the continental shelf at the Valencia region, even Sites of Community Importance were designated to specifically protect common bottlenose dolphin (Revuelta et al., 2018); those are much smaller than the common bottlenose dolphin distribution in the study area [see Gómez de Segura et al. (2004); Gómez de Segura et al. (2008)]. Marine protected areas also function as nurseries for many target species of regional fisheries, and fishing activity is intense around these areas. All these imply a frequent overlapping between dolphin distribution range and fishing grounds.
Bottom trawling is one of the two main fisheries operating in the area regarding the number of censed vessels, engine power, and fishing effort. Several cetacean species distributions (Gómez de Segura et al., 2004; Gómez de Segura et al., 2008) overlap with the estimated bottom trawling fishing hot spot areas. By-catch events of common bottlenose dolphins and striped dolphins by bottom trawls were reported also in low numbers in other areas of the Mediterranean Sea (Bearzi, 2002; Fortuna et al., 2010). The observed differences in the reported by-catch of cetacean species among subareas could be related to the mean fishing depth of bottom trawlers operating on each subarea. Subarea 1 has a wider continental shelf, which implies that the fishing activities of bottom trawlers concentrate at shallower waters. In fact, bottom trawlers fishing in subarea 1 and in the neritic zone of other subareas reported higher numbers of common bottlenose dolphin accidentally caught, supporting that depth is one factor ruling common bottlenose dolphin interaction with this fishery (Gonzalvo et al., 2008).
On the other hand, bottom trawlers working in the oceanic zone and in waters off subareas with a narrower continental shelf (subareas 2, 3, and 4) work at a higher mean fishing depth (m) and, therefore, reported striped dolphin accidental catches. This result may support the idea about how prey abundance and distribution influence the preferred habitats for cetaceans (Cañadas et al., 2002; Giannoulaki et al., 2016) and the consequent threats associated with the habitat of the species.
Striped dolphin has oceanic distribution in the region (Gómez de Segura et al., 2008, and references therein); however, in the last decade, this species seems to have changed its feeding habits in the Western Mediterranean, increasing the proportion of demersal preys (Aznar et al., 2017), also targeted by bottom trawlers. Moreover, long-term boat‐based surveys seemed to indicate a switching trend in the habitat preference of striped dolphin since 2008, expanding its range to the continental shelf (Fraija-Fernández et al., 2015). Despite the fact that more detailed studies on the current distribution and habitat use by striped dolphins in the area are needed, these evidence may explain the interaction of this species with bottom trawling in the present study. Further monitoring of both the striped dolphin habitat use and by-catch by bottom-trawling is necessary to avoid future conservation problems for the species related to its interaction with this fishery.
The artisanal fleet using trammel nets and gillnets also showed hot spot areas of fishing activity throughout the Valencia region coastal waters. In the present study, artisanal fishers reported little dolphin by-catch numbers as in a previous study held in the area (Revuelta et al., 2018) and in other areas of the Mediterranean Sea (Díaz López, 2006; Brotons et al., 2008; Gonzalvo et al., 2015; Pennino et al., 2015). This may be indicative that dolphins are able to detect and avoid the nets in most of the cases. This ability would let the dolphins approach the nets and take captures, therefore causing fishing gear damage and economic loss (Snape et al., 2018). Despite that there are no accurate common bottlenose dolphin population estimations in the studied area since almost two decades (Gómez de Segura et al., 2006), this species seems abundant and has been seen frequently in coastal waters (MZU-UV, studies in progress), often in hot spot areas of artisanal fishing grounds. Moreover, the opportunistic feeding strategy of the species includes demersal target species of this fishing gear (Blanco et al., 2001; Giménez et al., 2017). Overall, the high percentage of artisanal fishers that reported negative interactions with common bottlenose dolphin in our study (see also Revuelta et al., 2018) is not unexpected. The nearshore distribution of this fishery overlaps with common bottlenose dolphin distribution typically found at the continental shelf (Gannier, 2005; Gnone et al., 2011). However, as was described above, changes in striped dolphin distribution in the area could include this species in the conflict with local artisanal fishers, particularly because this fishing hot spot area is found at the south of Valencia province (Figure 1), located west to the most important area for this species in the waters off the Valencia region (Gómez de Segura et al., 2008).
Our results also show that there were differences between the three provinces. Ports at Alicante, as reported by Revuelta et al. (2018), but specially at Valencia province, seemed to have higher economic loss in contrast to those found at Castellón, the northernmost province. There was a generalized fisher’s perception that the interaction with dolphins is increasing throughout all the study areas in the last 5 years, which also has been recorded in other regions of Spain and in other areas of the Mediterranean Sea (Mónaco et al., 2020). In fact, the highest proportion of fishers perceiving an increase of dolphin presence in their fishing grounds belonged to artisanal fisheries (63.55%). However, the economic loss reported by artisanal fishers in our study was similar to the values registered by Revuelta et al. (2018) in interviews made in 2015; hence, despite that fishers have become more active in clamming for this issue with dolphins, the problem seems not to have economically worsened in the last years.
In order to have a complete assessment of the interaction between cetaceans and fisheries, we also took into consideration the two other minoritarian fisheries regarding the number of censed vessels: the purse-seines and pelagic longlines. According to interviews, purse-seiners had the highest by-catch rate compared with other fishing gears. The by-catch events of dolphins, more specifically of common bottlenose dolphin and striped dolphin, were reported. These species have also been captured by purse-seines elsewhere (Tudela, 2004; Zahri et al., 2007; Marçalo et al., 2015). The coastal distribution of purse-seiners targeting small pelagic fish schools again explains the interaction with common bottlenose dolphin. Striped dolphin by-catch events in this fishery could be explained because purse-seiners target species are part of the diet of this dolphin (Gómez-Campos et al., 2011; Aznar et al., 2017), and as explained before, by the potential habitat switch of striped dolphin in the area (Aznar et al., 2017).
The pelagic longline fleet at the study area currently has a small representation despite being bigger in the past (Generalitat Valenciana, 2020), and its involvement in by-catch events has decreased due to laws forcing changes in the fishing gear (Tomás et al., 2008). In the present study, one accidental capture of one Risso’s dolphin was reported by this pelagic fishery. It is possible that the interaction between this fishery and this species could have been underestimated or undetected in the past since the Risso’s dolphin distribution overlaps with the fishing grounds of this fishery in waters over the slope and in open waters, and the species has already been reported being by-caught by pelagic longlines in the Western Mediterranean (Macías-López et al., 2012).
4.2 Assessment of fisheries interactions according to strandings
Cetacean long-term stranding data (1990–2020) show a small percentage of cetaceans stranded with evidence of fisheries interactions. Only in a few of these cases could the specific fishing gear be identified. Common bottlenose dolphin and striped dolphin were the two species showing more strandings with fisheries interaction signs; however, our data gathering revealed that other species may interact with fisheries in the study area, although sporadically. This reduced number of records may be a consequence of the oceanic distribution of most cetacean species inhabiting Spain’s Mediterranean waters [see Gómez de Segura et al. (2004)], which makes it difficult for the carcasses of dead animals to reach the coast. Nonetheless, the stranding data support the information provided by fishers during interviews, particularly concerning the interaction with common bottlenose dolphin and also the striped dolphin. The increase in stranding events with fisheries interaction signs of individuals of these two dolphin species over the whole study period may be conditioned by the improvement of the network, particularly during the first 10 years, since no significant trend was found for striped dolphins in the latter years. However, this did happen for the common bottlenose dolphin.
The common bottlenose dolphin has been also reported as the species with a higher percentage of individuals stranded with fisheries interactions signs in a recent study carried out in the Catalonian coast, north to the Valencia region (Cuvertoret-Sanz et al., 2020). In that study, fishery interaction was the most frequent cause of death for the analyzed common bottlenose dolphins, thus confirming the magnitude of this threat in the Western Mediterranean [as informed by Hammond et al. (2012)]. As said before, in the present study, we detected a recent increasing trend in common bottlenose dolphin strandings with signs of fisheries interactions, which may support the perception of increase in abundance of common bottlenose dolphin stocks as reported by fishers during the interviews. This increasing trend in strandings could be linked to an increase in dolphin population size as reported elsewhere (e.g., Leeney et al., 2008) as well as to the increase in associated negative fishery interactions [see also Powell and Wells (2011)]. Moreover, Cuvertoret-Sanz et al. (2020) also reported striped dolphin as a species dying because of fishery interaction. In our study, this species has the highest number of individuals with fisheries interaction signs, even more than the common bottlenose dolphin, but it has a lower percentage probably because it is the most abundant cetacean in the area and, thus, in the stranding network. It has also suffered several episodes of massive strandings caused by a Morbillivirus epizootic (Van Bressem et al., 2014). Nonetheless, although for this species a recently increasing trend on stranding individuals with fisheries interaction signs was not detected, the total number of records could confirm the interaction level at which the species is exposed at the study area.
The high stranding density observed for the two species of delphinids at the south of the study area could be related with the location of the ports Santa Pola and Villajoyosa, both contributing to a high percentage of the engine power of the regional fishing fleet, a factor that has been correlated with strandings in other places (Byrd et al., 2014; Crosti et al., 2017). Areas with higher engine power are supposed to contain a larger fleet. Santa Pola and Villajoyosa are the ports with the highest bottom trawling fleet in the Valencia region. However, in other parts of the study area engine power does not match with strandings distribution. The high contribution of fishing capacity found at the northern ports is not associated with the strandings of striped dolphin, probably because this is not the preferred area of distribution for the species in the Valencia region waters (Gómez de Segura et al., 2006) and also because the continental shelf is much wider in this area, and this species is preferably distributed in waters over the slope. The fishing fleet engine power in Castellón is also not related with common bottlenose dolphin stranding distribution, probably because the local stocks can be found at waters further from the coast, over the wide continental shelf near Columbretes Islands (Revuelta et al., 2018). Carcasses of animals dead in open waters are less likely to reach the coast and be recorded (Leeney et al., 2008). The opposite situation seems to happen at Valencia province, where low engine power contribution values do not explain the clusters of striped dolphin strandings, probably associated with the proximity to the preferred area of distribution for striped dolphin in the region. Therefore, fishing capacity alone does not seem to explain the stranding distribution throughout the region The effect of other variables such as species distribution (Leeney et al., 2008), abundance of cetacean species and/or of their prey, or drifting conditions and migration of carcasses due to sea currents and winds (Peltier et al., 2014; Saavedra et al., 2017; Jog et al., 2022) should be also considered to explain cetacean distribution. In fact, at the study area, the stranding clusters of both dolphin species found at the south of the Valencia province could be explained by the existence of a main north–south surface coastal current, already described as a potential factor explaining loggerhead sea turtle stranding distribution in the area (Tomás et al., 2008).
4.3 Combination of methodologies and future management actions
As described in the previous section, stranding-based studies are subject to several gaps, and several factors could explain stranding aggregations at certain locations. As said before, interview-based studies depend on reliability of the interviewed fishers. However, working with marine protected megafauna is often costly and logistically difficult. Hence, combining data obtained from these relatively low-cost methodologies over a long period and large area, despite not fully accurate, may help in gaining knowledge on cetacean species and assessing threats that affect them in a certain area so as to help in their conservation. Through a combination of interviews to fishers, stranding records, and spatial analysis, we could assess cetacean–fisheries interactions happening in waters inside and neighboring the Cetacean Migration Corridor MPA, a key area for cetacean protection. Our results, especially for common bottlenose dolphin and striped dolphin, reflect the need of monitoring and applying management and conservation measures on waters inside and near this protected area.
The current situation of local fisheries is critical due to several factors, including the reduction of target species’ stocks, the increase of fuel cost caused by the current global situation, the recent COVID-19 pandemic situation, and the lack of generational handover. The economical loss caused by dolphins is already seen as another problem added to this situation and may result in illegal actions against this protected species. Valencia fishers are still very collaborative with conservation workers and aware of conservation issues; however, management actions are needed for both keeping the fishing activity sustainable and preserving the dolphin populations in the area. Future work must focus on updating extant knowledge in cetacean species abundance, distribution, behavior, and habitat use in order to assess spatial risk (Jog et al., 2022), but it is also necessary to implement management actions for reducing cetacean–fisheries interactions, such as the use of visual deterrent devices, acoustic deterrent devices, and gear modifications, also contributing with compensation funds to fishers in order to reduce these interactions (Jog et al., 2022).
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors without undue reservation.
Author contributions
MI-S, JT, JAR, and OR contributed to the conception and design of the study. MI-S, RM-L, and DR-G undertook most of the interviews. MI-S, PG, OR, RM-L, and DR-G created the databases. MI-S, OR, and RM-L performed the data analyses and statistics. MI-S, OR, RM-L, and JT wrote the manuscript. JT and JAR searched for funding for the study. All authors contributed to the article and approved the submitted version.
Funding
This work is supported by the Biodiversity Foundation of the Ministry for the Ecological Transition and the Demographic Challenge, the Generalitat Valenciana (Spain), the EU project LIFE MEDTURTLES (LIFE18 NAT/IT/000103), and project AICO/2021/022 granted by Conselleria d’Innovació, Universitats, Ciència i Societat Digital, Generalitat Valenciana.
Acknowledgments
We would like to thank the team working on face-to-face interview campaigns held on the Valencia region ports, particularly to Francesc Domènech Rosell and Ángela Casas Quirante. We also thank Juan Eymar and the Department of Agriculture, Rural Development, Climate Emergency and Ecological Transition of the regional governement (Generalitat Valenciana) for the contacts made with fishers’ associations previously to the interview campaign. We strongly acknowledge all fisher associations and fishers who participated in the interviews. Thanks for the support of the Asociación Chelonia and the US Fish and Wildlife Service Division of International Conservation. We are also grateful to the past and present members of the staff of the Marine Zoology Unit of the Cavanilles Research Institute of Biodiversity and Evolutionary Biology (University of Valencia) for helping in the long-term stranding data collection.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.981638/full#supplementary-material
References
Aguilar A. (1991). Calving and early mortality in the western Mediterranean striped dolphin, Stenella coeruleoalba. Can. J. Zool. 69 (5), 1408–1412. doi: 10.1139/z91-199
Avila I. C., Kaschner K., Dormann C. F. (2018). Current global risks to marine mammals: Taking stock of the threats. Biol. Conserv. 221, 44–58. doi: 10.1016/j.biocon.2018.02.021
Aylesworth L., Phoonsawat R., Suvanachai P., Vincent A. C. J. (2017). Generating spatial data for marine conservation and management. Biodivers. Conserv. 26 (2), 383–399. doi: 10.1007/s10531-016-1248-x
Aznar F. J., Míguez-Lozano R., Ruiz B., de Castro A. B., Raga J. A., Blanco C. (2017). Long-term changes, (1990-2012) in the diet of striped dolphins Stenella coeruleoalba from the Western Mediterranean. Mar. Ecol. Prog. Ser. 568, 231–247. doi: 10.3354/meps12063
Bearzi G. (2002). Interactions between cetacean and fisheries in the Mediterranean sea. Cetaceans of the Mediterranean and black seas: state of knowledge and conservation strategies, vol. 9. Ed. di Sciara G.N. (Monaco: ACCOBAMS), 78–97.
Blanco C., Salomón O., Raga J. A. (2001). Diet of the bottlenose dolphin (Tursiops truncatus) in the western Mediterranean Sea. J. Mar. Biol. Assoc. U. K. 81 (6), 1053–1058. doi: 10.1017/S0025315401005057
Boisseau O., Lacey C., Lewis T., Moscrop A., Danbolt M., McLanaghan R. (2010). Encounter rates of cetaceans in the Mediterranean Sea and contiguous Atlantic area. J. Mar. Biol. Assoc. U. K. 90 (8), 1589–11599. doi: 10.1017/s0025315410000342
Bonizzoni S., Hamilton S., Reeves R. R., Genov T., Bearzi G. (2022). Odontocete cetaceans foraging behind trawlers, worldwide. Rev. Fish. Biol. Fish. 32, 827–877. doi: 10.1007/s11160-022-09712-z
Boubekri I., Amara R., Mazurek H. (2019). Characterization of purse seine fishery of Ziama Bay (Gulf of Béjaia, SW mediterranean): Implications for sustainability management. Ocean Coast. Manage. 175, 79–89. doi: 10.1016/j.ocecoaman.2019.03.024
Brotons J. M., Grau A. M., Rendell L. (2008). Estimating the impact of interactions between bottlenose dolphins and artisanal fisheries around the Balearic islands. Mar. Mamm. Sci. 24 (1), 112–127. doi: 10.1111/j.1748-7692.2007.00164.x
Byrd B. L., Hohn A. A., Lovewell G. N., Altman K. M., Barco S. G., Friedlaender A., et al. (2014). Strandings as indicators of marine mammal biodiversity and human interactions off the coast of north Carolina. Fish. Bull. 112 (1), 1–23. doi: 10.7755/FB.112.1.1
Cañadas A., Sagarminaga R., Garcıa-Tiscar S. (2002). Cetacean distribution related with depth and slope in the Mediterranean waters off southern Spain. Deep-Sea Res. 49 (11), 2053–2073. doi: 10.1016/s0967-0637(02)00123-1
Carbonell A., Martin P., de Ranieri S., WEDIS team (1998). Discards of the western Mediterranean trawl fleets. Commun. Int. Mer. Médit. 35, 392–393.
Carbonell A., Carbonell M., Demestre M., Grau A., Monserrat S. (1999). The red shrimp Aristeus antennatus (Risso 1816) fishery and biology in the Balearic islands, Western Mediterranean. Fish. Res. 44 (1), 1–13. doi: 10.1016/s0165-7836(99)00079-x
Carpentieri P., Nastasi A., Sessa M., Srour A. (2021). Incidental catch of vulnerable species in Mediterranean and black Sea fisheries – a review. Rome: FAO. doi: 10.4060/cb5405en
Carreras C., Cardona L., Aguilar A. (2004). Incidental catch of the loggerhead turtle Caretta caretta off the Balearic islands (western Mediterranean). Biol. Conserv. 117 (3), 321–329. doi: 10.1016/j.biocon.2003.12.010
Casale P., Affronte M., Insacco G., Freggi D., Vallini C., Pino d’Astore P., et al. (2010). Sea Turtle strandings reveal high anthropogenic mortality in Italian waters. Aquat. Conserv.: Mar. Freshw. Ecosyst. 20 (6), 611–620. doi: 10.1002/aqc.1133
Crosti R., Arcangeli A., Romeo T., Andaloro F. (2017). Assessing the relationship between cetacean strandings (Tursiops truncatus and Stenella coeruleoalba) and fishery pressure indicators in Sicily (Mediterranean Sea) within the framework of the EU habitats directive. Eur. J. Wildl Res. 63 (3), 1–13. doi: 10.1007/s10344-017-1111-8
Cuvertoret-Sanz M., López-Figueroa C., O’Byrne A., Canturri A., Martí-Garcia B., Pintado E., et al. (2020). Causes of cetacean stranding and death on the catalonian coast (western Mediterranean Sea), 2012-2019. Dis. Aquat. Organ. 142, 239–253. doi: 10.3354/dao03550
Díaz López B. (2006). Interactions between Mediterranean bottlenose dolphins (Tursiops truncatus) and gillnets off Sardinia, Italy. ICES J. Mar. Sci. 63, 946–951. doi: 10.1016/j.icesjms.2005.06.012
Domènech F., Álvarez de Quevedo I., Merchán M., Revuelta O., Vélez-Rubio G., Bitón S., et al. (2015). Incidental catch of marine turtles by Spanish bottom trawlers in the western Mediterranean. Aquat. Conserv.: Mar. Freshw. Ecosyst. 25 (5), 678–689. doi: 10.1002/aqc.2463
Duignan P. J., Geraci J. R., Raga J. A., Calzada N. (1992). Pathology of morbillivirus infection in striped dolphins (Stenella coeruleoalba) from Valencia and Murcia, Spain. Can. J. Vet. Res. 56 (3), 242–248.
Duras M., Galov A., Korpes K., Kolenc M., Baburić M., Gudan Kurilj A., et al. (2021). Cetacean mortality due to interactions with fisheries and marine litter ingestion in the Croatian part of the Adriatic Sea from 1990 to 2019. Vet. Arh. 91 (2), 189–206. doi: 10.24099/vet.arhiv.1254
Fernández-Contreras M. M., Cardona L., Lockyer C. H., Aguilar A. (2010). Incidental bycatch of short-beaked common dolphins (Delphinus delphis) by pairtrawlers off northwestern Spain. ICES J. Mar. Sci. 67 (8), 1732–1738. doi: 10.1093/icesjms/fsq077
Fortuna C. M., Vallini C., Filidei E., Ruffino M., Consalvo I., Di Muccio S., et al. (2010). By-catch of cetaceans and other species of conservation concern during pair trawl fishing operations in the Adriatic Sea (Italy). Chem. Ecol. 26 (sup 1), 65–76. doi: 10.1080/02757541003627662
Fraija-Fernández N., Gozalbes P., Tomás J., Balbuena J. A., Domènech F., Eymar J., et al. (2015). Long-term boat-based surveys in the central Spanish Mediterranean, (2003–2013): cetacean diversity and distribution. In: Book of abstracts of the 29th. Malta: European Cetacean Society conference, P 141.
Gannier A. (2005). Summer distribution and relative abundance of delphinids in the Mediterranean Sea. Rev. Ecol. (Terre Vie). 60 (3), 223–238.
Generalitat Valenciana. (2020). Available at: https://agroambient.gva.es/va/web/pesca/flota-pesquera (Accessed July 15, 2021).
Getis A., Ord J. K. (1992). The analysis of spatial association by use of distance statistics. in Geographical analysis, vol. 24. (Ohio State University Press), 189–206.
Giannoulaki M., Markoglou E., Valavanis V. D., Alexiadou P., Cucknell A., Frantzis A. (2016). Linking small pelagic fish and cetacean distribution to model suitable habitat for coastal dolphin species, Delphinus delphis and Tursiops truncatus, in the Greek seas (Eastern Mediterranean). Aquat. Conserv.: Mar. Freshw. Ecosyst. 27 (2), 436–451. doi: 10.1002/aqc.2669
Giménez J., Marçalo A., Ramírez F., Verborgh P., Gauffier P., Esteban R., et al. (2017). Diet of bottlenose dolphins (Tursiops truncatus) from the Gulf of Cádiz: Insights from stomach content and stable isotope analyses. PloS One 12 (9), e0184673. doi: 10.1371/journal.pone.0184673
Gnone G., Bellingeri M., Dhermain F., Dupraz F., Nuti S., Bedocchi D., et al. (2011). Distribution, abundance, and movements of the bottlenose dolphin (Tursiops truncatus) in the pelagos sanctuary MPA (north-west Mediterranean Sea). Aquat. Conserv.: Mar. Freshw. Ecosyst. 21 (4), 372–388. doi: 10.1002/aqc.1191
Goetz S., Read F. L., Santos M. B., Pita C., Pierce G. J. (2014). Cetacean occurrence, habitat preferences and potential for cetacean-fishery interactions in Iberian Atlantic waters: results from cooperative research involving local stakeholders. Aquat. Conserv.: Mar. Freshw. Ecosyst. 25 (1), 138–154. doi: 10.1002/aqc.2481
Gómez-Campos E., Borrell A., Cardona L., Forcada J., Aguilar A. (2011). Overfishing of small pelagic fishes increases trophic overlap between immature and mature striped dolphins in the Mediterranean Sea. PloS One 6 (9), e24554. doi: 10.1371/journal.pone.0024554
Gómez de Segura A., Tomás J., Raga J. A. (2004) Sector centro (Comunidad Valenciana y región de Murcia). In Proyecto Mediterráneo. Zonas de especial interés para la conservación de los cetáceos en el Mediterráneo español. Naturaleza y Parques Nacionales Serie Técnica, vol. 3 . Eds. Raga A., Pantoja J.. (Madrid: Ministerio de Medio Ambiente), 67–131.
Gómez de Segura A., Crespo E. A., Pedraza S. N., Hammond P. S., Raga J. A. (2006). Abundance of small cetaceans in the waters of the central Spanish Mediterranean. Mar. Biol. 150 (1), 149–160. doi: 10.1007/s00227-006-0334-0
Gómez de Segura A., Hammond P. S., Raga J. A. (2008). Influence of environmental factors on small cetacean distribution in the Spanish Mediterranean. J. Mar. Biol. Assoc. U. K. 88 (6), 1185–1192. doi: 10.1017/S0025315408000386
Gonzalvo J., Valls M., Cardona L., Aguilar A. (2008). Factors determining the interaction between common bottlenose dolphins and bottom trawlers off the Balearic archipelago (western Mediterranean Sea). J. Exp. Mar. Biol. Ecol. 367 (1), 47–52. doi: 10.1016/j.jembe.2008.08.013
Gonzalvo J., Giovos I., Moutopoulos D. K. (2015). Fishermen's perception on the sustainability of small-scale fisheries and dolphin–fisheries interactions in two increasingly fragile coastal ecosystems in western Greece. Aquat. Conserv.: Mar. Freshw. Ecosyst. 25 (1), 91–106. doi: 10.1002/aqc.2444
Gozalbes P., Jiménez J., Raga J. A., Esteban J. A., Tomás J., Gómez J. A., et al. (2010). Cetáceos y tortugas marinas en la comunitat valenciana. 20 años de seguimiento. In Collecció treballs tècnics de biodiversitat, 3. ed. consellería de medio ambiente, agua, urbanismo y vivienda (Valencia: Generalitat Valenciana), 61 pp.
Gray C. A., Kennelly S. J. (2018). Bycatches of endangered, threatened and protected species in marine fisheries. Rev. Fish. Biol. Fish. 28 (3), 521–541. doi: 10.1007/s11160-018-9520-7
Greenwood J. J. (1996). Basic techniques. In: Ecological Census Techniques. Ed. Sutherland W. J. (Cambridge: Cambridge University Press), 11–110.
Hammond P. S., Bearzi G., Bjørge A., Forney K. A., Karkzmarski L., Kasuya T., et al. (2012). Tursiops truncatus. In The IUCN red list of threatened species 2012. IUCN. doi: 10.2305/IUCN.UK.2012.RLTS.T22563A17347397.en
Hedgpeth J. W. (1957). Classification of marine environments. In Treatise on marine ecology and paleoecology (USA: Geological Society of America Memoirs), vol. 67, 17–28. doi: 10.1130/mem67v1-p17
International Council for the Exploration of the Sea (ICES) (2008). Report of the ICES advisory committee 2008. ICES advice 2008 (Copenhagen: International Council for the Exploration of the Sea), 842 pp.
IUCN (2021) The IUCN red list of threatened species. version 2021-3. Available at: https://www.iucnredlist.org (Accessed July 18, 2022).
Jog K., Sutaria P., Diedrich A., Grech A., Marsh H. (2022). Marine mammal interactions with fisheries: Review of research and management trends across commercial and small-scale fisheries. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.758013
Kie J. G. (2013). A rule-based ad hoc method for selecting a bandwidth in kernel home-range analyses. Anim. Biotelemetry. 1 (1), 13. doi: 10.1186/2050-3385-1-13
Lauriano G., Fortuna C. M., Moltedo G., Notarbartolo di Sciara G. (2004). Interaction between common bottlenose dolphin (Tursiops truncatus) and the artisanal fishery in asinara island national park (Sardinia): Assessment of catch damage and economic loss. J. Cetacean Res. Manage. 6, 165–173.
Leeney R. H., Amies R., Broderick A. C., Witt M. J., Loveridge J., Doyle J., et al. (2008). Spatio-temporal analysis of cetacean strandings and bycatch in a UK fisheries hotspot. Biodiver. Conserv. 17, 2323–2338. doi: 10.1007/s105331-008-9377-5
Léopold M., Guillemot N., Rocklin D., Chen C. (2014). A framework for mapping small-scale coastal fisheries using fishers’ knowledge. ICES J. Mar. Sci. 71 (7), 1781–1792. doi: 10.1093/icesjms/fst204
Lewison R. L., Crowder L. B., Wallace B. P., Moore J. E., Cox T., Zydelis R., et al. (2014). Global patterns of marine mammal, seabird, and sea turtle bycatch reveal taxa-specific and cumulative megafauna hotspots. PNAS 111 (14), 5271–5276. doi: 10.1073/pnas.1318960111
López A., Pierce G. J., Santos M. B., Gracia J., Guerra A. (2003). Fishery by-catches of marine mammals in Galician waters: Results from on-board observations and an interview survey of fishermen. Biol. Conserv. 111 (1), 25–40. doi: 10.1016/S0006-3207(02)00244-6
Macı́as-López D., Barcelona S. G., Báez J. C., de la Serna J. M., and Ortiz de Urbina J. M. (2012). Marine mammal bycatch in Spanish Mediterranean large pelagic longline fisheries, with a focus on Risso’s dolphin (Grampus griseus). Aquat. Living Resour. 25 (4), 321–331. doi: 10.1051/alr/2012038
Mangiafico S. S. (2015). An R companion for the handbook of biological statistics, version 1.3.2 (New Brunswick: Rutgers Cooperative Extension).
Marçalo A., Katara I., Feijo D., Araujo H., Oliveira I., Santos J., et al. (2015). Quantification of interactions between the Portuguese sardine purse-seine fishery and cetaceans. ICES J. Mar. Sci. 72 (8), 2438–2449. doi: 10.1093/icesjms/fsv076
Ministry of Agriculture, Fisheries and Food (2020) Fishing fleet census. Available at: https://servicio.pesca.mapama.es/censo/ConsultaBuqueRegistro/Buques/Search (Accessed August 8, 2021).
Mónaco C., Aguilera R., Camiñas J. A., Laspina M., Molina M., Said A., et al. (2020). Interactions between cetaceans and small-scale fisheries inthe Mediterranean. Conclusive Report. Published by Low Impact Fishers of Europe. 24 pp.
Moore J. E., Cox T. M., Lewison R. L., Read A. J., Bjorkland R., McDonald S. L., et al. (2010). An interview-based approach to assess marine mammal and sea turtle captures in artisanal fisheries. Biol. Conserv. 143 (3), 795–805. doi: 10.1016/j.biocon.2009.12.023
Moore J. E., Heinemann D., Francis T. B., Hammond P. S., Long K. J., Punt A., et al. (2021). Estimating bycatch mortality for marine mammals: Concepts and best practices. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.752356
Nelms S. E., Alfaro-Shigueto J., Arnould J. P. Y., Avila I. C., Nash S. B., Campbell E., et al. (2021). Marine mammal conservation: over the horizon. Endanger. Species Res. 44, 291–325. doi: 10.3354/esr01115
Nortarbartolo di Sciara G. (2016). Marine mammals in the Mediterranean Sea: An overview. Medit. Mar. Mam. Ecol. Cons. 75, 1–36. doi: 10.1016/bs.amb.2016.08.005
OceanCare (2021) Quiet waters for whales and dolphins: the one-time opportunity to avoid, reduce and mitigate noise-generating activities in the Mediterranean Cetacean Migration Corridor. Available at: https://www.oceancare.org/wp-content/uploads/2021/11/Quiet-Waters-en-web.pdf (Accessed May 1, 2022).
Peltier H., Jepson P. D., Dabin W., Deaville R., Daniel P., Van Canneyt O., et al. (2014). The contribution of stranding data to monitoring and conservation strategies for cetaceans: developing spatially explicit mortality indicators for common dolphins (Delphinus delphis) in the eastern north-Atlantic. Ecol. Ind. 39, 203–214. doi: 10.1016/j.ecolind.2013.12.019
Peltier H., Authier M., Caurant F., Dabin W., Daniel P., Dars C., et al. (2021). In the wrong place at the wrong time: Identifying spatiotemporal co-occurrence of bycaught common dolphins and fisheries in the Bay of Biscay (NE Atlantic) from 2010 to 2019. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.704005
Pennino M. G., Rotta A., Pierce G. J., Bellido J. M. (2015). Interaction between bottlenose dolphin (Tursiops truncatus) and trammel nets in the archipelago de la maddalena, Italy. Hydrobiologia. 747 (1), 69–82. doi: 10.1007/s10750-014-2127-7
Powell J. R., Wells R. S. (2011). Recreational fishing depredation and associated behaviors involving common bottlenose dolphins (Tursiops truncatus) in Sarasota bay, Florida. Mar. Mamm. Sci. 27 (1), 111–129. doi: 10.1111/j.1748-7692.2010.00401.x
Puig-Lozano R., Fernández A., Sierra E., Saavedra P., Suárez-Santana C. M., de la Fuente J., et al. (2020). Retrospective study of fishery interactions in stranded cetaceans, Canary Islands. Front. Vet. Sci. 7. doi: 10.3389/fvets.2020.567258
QGIS Development Team (2021) QGIS geographic information system. open-source geospatial foundation project. Available at: http://qgis.osgeo.org (Accessed January 10, 2022).
Raga J. A., Banyard A., Domingo M., Corteyn M., Van Bressem M. F., Fernández M., et al. (2008). Dolphin Morbillivirus epizootic resurgence, Mediterranean Sea. Emerg. Infect. Dis. 14 (3), 471–473. doi: 10.3201/eid1403.071230
Revuelta O., Domènech F., Fraija-Fernández N., Gozalbes P., Novillo O., Penadés-Suay J., et al. (2018). Interaction between bottlenose dolphins (Tursiops truncatus) and artisanal fisheries in the Valencia region (Spanish Mediterranean Sea). Ocean Coast. Manage. 165, 117–125. doi: 10.1016/j.ocecoaman.2018.08.001
Saavedra C., Pierce G. J., Gago J., Jusufovski D., Cabrero A., Cerviño S., et al. (2017). Factors driving patterns and trends in strandings of small cetaceans. Mar. Biol. 164, 165. doi: 10.1007/s00227-017-3200-3
Sala A., Notti E., Bonanomi S., Pulcinella J., Colombelli A. (2019). Trawling in the Mediterranean: An exploration of empirical relations connecting fishing gears, otterboards and propulsive characteristics of fishing vessels. Front. Mar. Sci. 6. doi: 10.3389/fmars.2019.00534
Snape R. T. E., Broderick A. C., Ç̧iç̧ek B. A., Fuller W. J., Tregenza N., Witt M. J., et al. (2018). Conflict between dolphins and a data-scarce fishery of the European union. Hum. Ecol. 46 (3), 423–433. doi: 10.1007/s10745-018-9989-7
Tomás J., Aznar F. J., Raga J. A. (2001). Feeding ecology of the loggerhead turtle Caretta caretta in the western Mediterranean. J. Zool. 255, 525–532. doi: 10.1017/S0952836901001613
Tomás J., Gozalbes P., Raga J. A., Godley B. J. (2008). Bycatch of loggerhead sea turtles: insights from 14 years of stranding data. Endanger. Species Res. 5 (2-3), 161–169. doi: 10.3354/esr00116
Tudela S. (2004). Ecosystem effects of fishing in the Mediterranean: an analysis of the major threats of fishing gear and practices to biodiversity and marine habitats. stud. rev. General Fisheries Commission for the Mediterranean Vol. 74 (Rome: FAO), 44p.
Van Bressem M. F., Duignan P., Banyard A., Barbieri M., Colegrove K., De Guise S., et al. (2014). Cetacean Morbillivirus: Current knowledge and future directions. Viruses. 6 (12), 5145–5181. doi: 10.3390/v6125145
Violi B., Verga A., Jones L., Calogero G., Soldano G., Cheeseman T., et al. (2021). A wanderer in the Mediterranean Sea: the case of a humpback whale (Megaptera novaeangliae) from the West indies. Aquat. Mamm. 47 (6), 599–611. doi: 10.1578/AM.47.6.2021.599
Worton B. J. (1989). Kernel methods for estimating the utilization distribution in home-range studies. Ecology. 70 (1), 164–168. doi: 10.2307/1938423
Keywords: Cetacean, fisheries interactions, common bottlenose dolphin, striped dolphin, interviews, strandings, Valencia region
Citation: Izquierdo-Serrano M, Revuelta O, Míguez-Lozano R, Gozalbes P, Ruiz-García D, Raga JA and Tomás J (2022) Assessment of the interactions between cetaceans and fisheries at the south of the Cetacean Migration Corridor and neighboring waters (Western Mediterranean). Front. Mar. Sci. 9:981638. doi: 10.3389/fmars.2022.981638
Received: 29 June 2022; Accepted: 01 August 2022;
Published: 24 August 2022.
Edited by:
Morgana Vighi, Consultant, Barcelona, SpainReviewed by:
Natascha Wosnick, Federal University of Paraná, BrazilArianna Azzellino, Politecnico di Milano, Italy
Copyright © 2022 Izquierdo-Serrano, Revuelta, Míguez-Lozano, Gozalbes, Ruiz-García, Raga and Tomás. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jesús Tomás, jesus.tomas@uv.es