 Carolina P. Pacini
Carolina P. Pacini Maria V. D. Soares
Maria V. D. Soares João F. Lacerda
João F. Lacerda- 1Instituto de Medicina Molecular João Lobo Antunes, Faculdade de Medicina, Universidade de Lisboa, Lisbon, Portugal
- 2Serviço de Hematologia e Transplantação de Medula, Hospital de Santa Maria, ULS Santa Maria, Lisbon, Portugal
Allogeneic Hematopoietic Stem Cell Transplantation (allo-HSCT) is the only curative therapy for many hematologic malignancies, whereby the Graft-versus-Leukemia (GVL) effect plays a pivotal role in controlling relapse. However, the success of GVL is hindered by Graft-versus-Host Disease (GVHD), where donor T cells attack healthy tissues in the recipient. The ability of natural regulatory T cells (Treg) to suppress immune responses has been exploited as a therapeutical option against GVHD. Still, it is crucial to evaluate if the ability of Treg to suppress GVHD does not compromise the benefits of GVL. Initial studies in animal models suggest that Treg can attenuate GVHD while preserving GVL, but results vary according to tumor type. Human trials using Treg as GVHD prophylaxis or treatment show promising results, emphasizing the importance of infusion timing and Treg/Tcon ratios. In this review, we discuss strategies that can be used aiming to enhance GVL post-Treg infusion and the proposed mechanisms for the maintenance of the GVL effect upon the adoptive Treg transfer. In order to optimize the therapeutic outcomes of Treg administration in allo-HSCT, future efforts should focus on refining Treg sources for infusion and evaluating their specificity for antigens mediating GVHD while preserving GVL responses.
Introduction
Graft-versus-Leukemia (GVL) is a crucial aspect of the success of Allogeneic Hematopoietic Stem Cell Transplantation (allo-HSCT) in the treatment of hematologic malignancies. This effect relies on the ability of donor T cells to recognize and eliminate tumor cells in the recipient’s body. However, this potent immune response can also lead to Graft-versus-Host Disease (GVHD), where donor-derived T cell clones react to antigens within healthy tissues of the recipient. The fine balance between GVHD and GVL responses is key for long-term leukemia-free survival.
Regulatory T cells (Treg) are a specialized subpopulation of CD4+ T cells, characterized by high and constitutive expression of CD25 (interleukin-2 receptor alfa-chain) and the transcription factor Foxp3. Treg cells play an essential role in controlling the immune response and maintaining homeostasis. This is achieved through suppression mechanisms that limit the proliferation and function of other cell types (1–3). Indeed, several studies have shown that both the occurrence and the severity of GVHD are strictly and significantly related to reduced absolute numbers and frequencies of Foxp3+CD4+CD25+ Tregs (4–8). Furthermore, patients receiving grafts with higher numbers of donor Foxp3+ Treg display a lower risk of developing GVHD (9–11). Accordingly, therapeutic strategies to reconstitute this population have emerged as a potential alternative treatment for GVHD. Therefore, in order for Treg cells to be used as a safe and effective therapeutical approach to treat GVHD, the suppressive effects of Treg in GVL must be considered, as they would likely lead to disease relapse and treatment failure.
Considerations on the Treg influence on GVL: lessons from animal models
Initial evidence from animal models suggested that Treg may suppress GVHD while preserving GVL. In 2003, using a mismatched mouse model, Edinger et al. (12) demonstrated that Treg prevent GVHD induction by inhibiting the expansion of alloreactive donor T cells, but not their activation or cytolytic capacity, allowing for the preservation of alloreactive T cells’ ability to eradicate established tumors. In the same year, Jones et al. (13) verified the relevance of controlling the timing of Treg administration, showing that only the very early infusion of expanded Treg regulates severe acute GVHD in an haploidentical mice model. The drawback of this strategy is that it may reduce the chance of sufficient GVL effect to occur. However, a delayed infusion of Treg after GVHD onset would give appropriate time for unrestricted alloreactive responses to be established. To confirm their hypothesis, they tested a less aggressive system, in which mice were matched for the major histocompatibility complex (MHC) but mismatched for minor histocompatibility antigens (mHA). In this model, donor-derived Treg, infused as late as ten days after the transplant/tumor challenge, increased animal survival without compromising the GVL response. In summary, the use of Treg in GHVD prophylaxis or treatment must consider the suppressive mechanisms and the time frame of Treg infusion, so that crucial GVL responses are not hampered.
Although animal studies provide valuable information, the complexity of the GVHD/GVL balance requires studies that explore these mechanisms in humans. In addition, the high diversity of protocols used in mouse models leads to contradictory results. For instance, differences in tumor cells injected in mice can lead to distinctive data that must be examined carefully, as some tumor lines can be easier to eradicate than others. The lack of GVL inhibition reported by Edinger and colleagues when using fresh Treg was tested on A20 and BCL1 cell lines (12). This was confirmed in another study, where the infusion of ex-vivo expanded recipient-specific Tregs preserved GVL following the injection of A20 lymphoma cells. However, GVL was not preserved when the P815 mastocytoma cell line was tested using the same GVHD mouse model (14), and also in a different study (15).
To bring mouse models closer to the reality of human tumors, Zhang et al. (16) tested a tumor cell line that is more aggressive and representative of human myeloid leukemia. In this setting, the Treg infusion was found to impair GVL responses by suppressing both responder T cell proliferation and proinflammatory cytokine production. In another study, however, the co-infusion of human Treg and Tcon in mice that had leukemia engraftment by primary human AML cells, SUP-B15 or Burkitt´s cells line showed that Treg did not compromise the anti-tumor effect of Tcon and still avoided GVHD (17). Such discrepancies highlight the importance of considering the nature of the tumors involved, as their biological features, such as aggressiveness and location, will likely influence Treg-mediated GVL suppression, especially in cases where both GVHD and GVL responses are driven by the same mechanisms. This stresses the importance of addressing this issue during pre-clinical Treg tests for GVHD therapy in humans.
Treg as a prophylactic or therapeutical approach for GVHD and their impact on GVL in human trials
The possibility that Treg impair anti-tumor specific T cell activity is a matter of concern when their clinical usage in GVHD is addressed. While studying immune reconstitution post-HSCT, Nadal et al. (18) have shown that disease-relapsed patients undergoing HSCT for myeloid leukemia had twice the number of peripheral blood (PB) Treg cells in circulation than patients in remission. Indeed, Tregs were the only predictive variable of disease relapse, by logistic regression analysis, in this study. On the other hand, Wolf et al. (19) did not find an association between Treg numbers in the graft and the risk of malignancy relapse in 58 patients in an HLA-matched context. It is therefore important to clarify the impact of Treg-based therapies in the GVHD/GVL balance in the clinical setting.
The promising potential of Treg manipulation to control GVHD has led to clinical trials evaluating Treg infusions before and after GVHD onset, where careful assessment of the impact on GVL was considered. Although Treg infusions result in increased Treg numbers that are essential to prevent or treat GVHD, the preservation of the GVL effect requires the presence of conventional T cells (Tcon) in circulation in the patient. In order to control the Treg : Tcon ratio to sustain the balance between GVHD and GVL responses, some studies tested the co-infusion of donor-Treg and donor-Tcon cells, either in 1:1 (20) or 2:1 Treg to Tcon ratio (17, 21). In one of the first trials, 28 patients undergoing haploidentical HSCT received an early infusion of Tregs four days before the transplant and the Tcon infusion. This regimen prevented GVHD in the absence of any post-transplantation immunosuppression, promoted lymphoid reconstitution and improved immunity to opportunistic pathogens. Furthermore, there was no apparent compromise of the GVL effect, as only one relapse occurred in their high relapse-risk cohort (21). Subsequent studies from the same group expanded the trial to a longer 45-month follow-up, demonstrating once more that infused donor Treg used prophylactically resulted in GVHD suppression without loss of GVL activity, as seen by the very low cumulative incidence of relapse compared to the historical controls (17). The authors’ subsequent trial in 50 patients in a myeloablative conditioning regimen, adapted to the patient’s age, tested the same Treg and Tcon co-infusion immunotherapy showing once again, a low rate (4%) of leukemia relapse (22).
Importantly, the timing of either Treg or Tcon donor cell infusion is relevant (Table 1). For instance, in a prophylactic approach, the administration of Treg two to four days before the transplant/Tcon infusion was especially advantageous for GVHD prevention, as it allowed time for a robust expansion of Treg in vivo while preserving GVL (17, 21, 26). In that way, this strategy reduces the need to transfer high numbers of Treg cells, which can be difficult to obtain without in vivo or ex vivo expansion protocols (23, 27). Besides the infusion timing, the clinical trials that explore the use of Treg in GVHD also vary in the source of Treg, in their purification for direct infusion or expansion, and in the level of Foxp3 expression of the product. Such variability in experimental design impacts differently on GVL responses and affects the comparisons across studies, as summarized in Tables 1 and 2. It is worth mentioning that fresh polyclonal Treg, ex vivo expanded Treg and in vitro induced Treg (iTreg) are all different products that should be investigated in specific clinical trials assessing safety and efficacy in each setting.
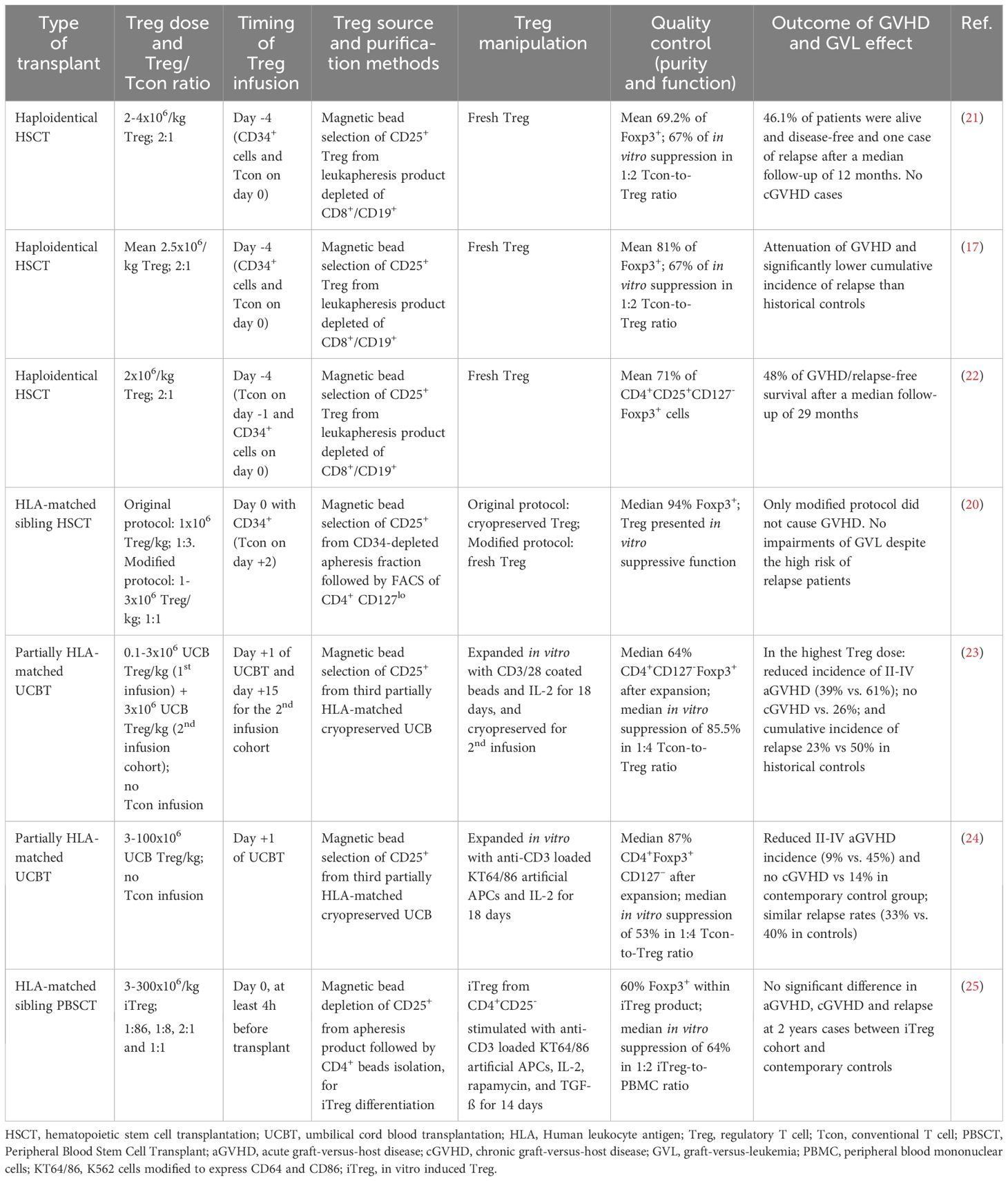
Table 1 The impact of the heterogeneity of adoptive Treg infusions for GVHD prophylaxis on the GVL effect.
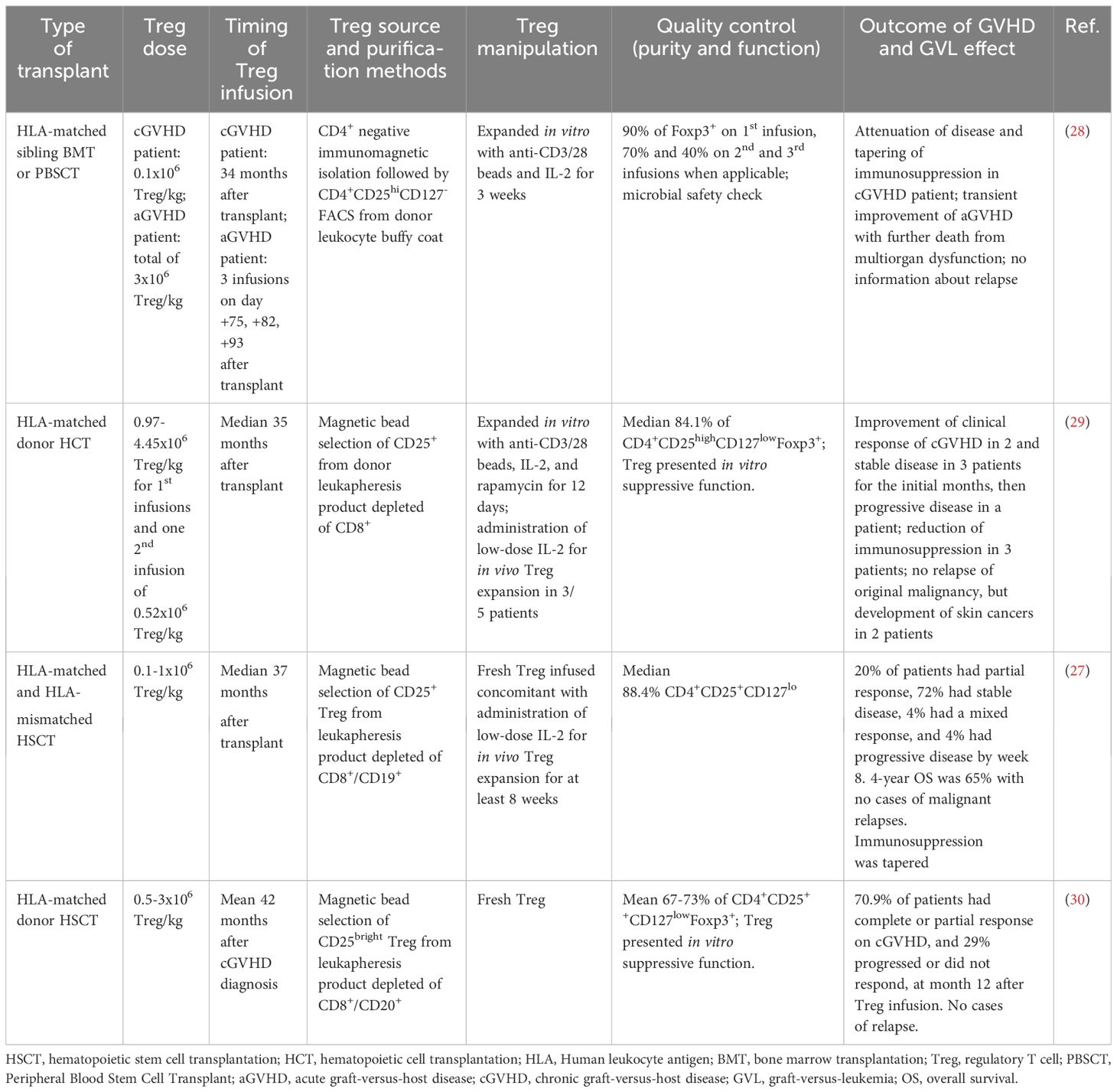
Table 2 The impact of the heterogeneity of adoptive Treg infusions for GVHD treatment on the GVL effect.
In the umbilical cord blood (UCB) transplant context, the transfer of the highest dose (3x106) of UCB expanded Treg/kg correlated with reduced aGVHD, no cGVHD cases, and a lower relapse rate than historical controls (23). In a subsequent study, these authors were also able to show that there was no increase in disease relapse as compared to the control group even when high doses of Treg were used (100x106 UCB Treg/kg) (24). However, in this UCB transplant model, Treg were collected from a third-party donor, which likely limited their survival and activity in the patients. Other clinical trials also did not find impairments in the GVL in HLA-matched sibling transplant, either when using an iTreg product, obtained from CD4+CD25- precursors, that were further expanded in vitro (25), or when using freshly isolated, highly purified sorted Treg followed by a Tcon infusion (20).
While most of the studies focus on the prophylactic administration of Treg to prevent GVHD, some trials have instead investigated the ability of late Treg infusions for GVHD treatment (27–30) (Table 2). The rationale behind this approach is based on the knowledge that leukemia is likely to have been eradicated at that point and therefore such studies do not generally prioritize the evaluation of the GVL response. Accordingly, relapse of the original tumor was not observed, although other skin cancers arose in some patients after Treg infusion in one of the studies (29). Thus, in trials using Treg therapeutically in the treatment of severe GVHD, other aspects besides leukemia relapse must be carefully evaluated, such as responses to infections or the emergence of new tumors (29, 31), that may also be hindered by the immunosuppressive activity of Treg.
Enhancing the GVL effect after Treg infusion for GVHD suppression
In order to decrease the aforementioned risks, the selective expansion of alloantigen-specific Treg (allo-Treg) for GVHD therapy has been pursued, as it generates more suppressive, specialized, and overall efficient Tregs than those that are polyclonally expanded (14, 32–36). Despite the greater specificity of allo-Treg, the possibility remains that some mHA can mediate GVHD but also be involved in GVL responses. For instance, due to H-Y mHA, which is only expressed in males, there is an increased chance of GVHD in male recipients from female HLA-matched donors, but also a reduced risk of relapse in those recipients (37). In mice, H-Y-specific iTreg preserved GVL when infused in mice with pre-established leukemia (36). The same group was able to expand human H-Y-specific Treg ex vivo, with the premise that they would prevent GVHD while sparing GVL responses against other mHA such as HA-1 and HA-2 (35, 38). However, the authors have not published to date the results relating to the use of their HY-specific iTreg in a clinical setting (15). Thus, it remains to be clarified whether monoclonal mHA-specific Treg would preserve GVL in humans or not. Likely, the enrichment of donor Treg cells specific to an array of mHA in the healthy tissues of the recipient would provide a more efficient and less broad suppression of alloreactive responses, possibly sparing the GVL effect.
More recently, the use of chimeric antigen receptors (CARs) has been exploited for the generation of antigen-specific Treg to promote transplantation tolerance. This approach likely allows the use of smaller cell doses and reduces the risk of broad and unspecific immune suppression. MacDonald et al. (39) created a CAR Treg specific to one of the most common mismatched antigens, the HLA-A2. Its forced expression on human Tregs promoted stronger proliferation compared to endogenous TCR stimulation, thus allowing the required doses for adoptive therapy to be obtained more easily. Importantly, the HLA-A2-specific CAR-Treg retained their phenotype and stability after in vitro expansion and suppressed allo-responses in a xenogeneic mouse model in vitro more effectively than polyclonal Treg. Of note, CAR-modified CD4+ Tcon can promote antigen-specific lysis as efficiently as CD8+ cells and produce high levels of pro-inflammatory cytokines, that can lead to cytokine release syndrome (CRS). Nevertheless, several studies have shown that CAR-Treg does not present a risk for CRS due to the reduced production of inflammatory cytokines after activation (39–41). Regarding cytolysis, although the authors could not find great levels of cytolytic activity in their CAR-modified Tregs in immunodeficient xenogeneic mice (39), such risks still need to be evaluated in humans as HLA-A2 expression is ubiquitous, thus increasing the likelihood of direct tissue damage in HLA-A2+ recipients. In fact, another study indicated that CAR-Tregs are able to employ some level of antigen-specific cytotoxicity despite CAR-specificity (42). Therefore, caution should be employed when considering broadly expressed antigens, such as HLA class I molecules, for CAR-Treg constructs, as this may induce a robust CAR stimulation, resulting in a generalized immunosuppressive state that may ultimately impair GVL responses.
With such limitations in mind, other targets started to be investigated for CAR-Treg creation. CD19-directed CAR-T cells have been approved and successfully used for the treatment of B-cell malignancies. Therefore, some studies generated and examined the potential of CD19-targeted CAR-Tregs to suppress B cells while delaying GVHD. Bolivar-Wagers et al. (40) used a fully MHC-mismatched allo-HSCT mouse model to test murine Treg containing a human CD19 CAR construct, that caused B cell aplasia without systemic toxicity. In a mouse model for aGVHD, they observed that in the presence of the human CD19 target, such CAR-Treg suppressed aGVHD efficiently by reducing proinflammatory cytokine-producing by Tcon and increasing Treg in Treg/Tcon ratio especially in the colon, a key target organ in aGVHD. Moreover, such CAR-Treg cells showed antigen-specific killing capacity that depended on perforin but not granzyme B (GZB) production, thus providing direct targeting of CD19+ tumor cells and reducing lymphoma cell growth in vivo (40). On the other hand, another study engineered CD19 CAR-Treg from human CD45RA+ purified Treg and used the CD28 costimulatory domain instead of 4-1BB (41), as in the previous study. In this case, CD45RA+ CD19 CAR-Treg did not present cytolytic activity, although they were able to suppress human B cell Ig production and differentiation into plasma cells in a xenogeneic mouse model of GVHD, also leading to attenuation of the disease.
These studies indicate that CAR-Treg administration has the potential to suppress GVHD while maintaining sufficient antitumor response. More recently, a clinical trial using a CD6-CAR Treg in GVHD was launched, aiming to take advantage of CD6-targeted anti-inflammatory response (clinicaltrials.gov identifier: NCT05993611). Noteworthy, it is important to acknowledge the differences in targets and costimulatory domains used in CAR construct, as they will dramatically impact the final product and therefore the effect and safety of the therapy (41, 42). Overall, more preclinical studies and the development of novel molecular on/off switches on CAR-Treg are important for the advancement of this strategy for GVHD therapy while preserving GVL (43).
Besides genetic modifications of the classical Foxp3+ Treg population, some authors are exploring the clinical potential of distinct regulatory subsets, such as type 1 regulatory (Tr1) cells. Tr1 are peripherally generated cells with suppressor ability associated with high IL-10 production, in the absence of Foxp3 expression. It has been shown that IL-10-engineered human CD4+ resemble natural Tr1s and present direct GZB-mediated cytotoxicity against myeloid leukemic cell lines, in an HLA class I-dependent manner, regardless of TCR specificity (44). In different humanized mouse models, these cells suppressed xenogeneic GVHD while preventing leukemia development by mediating directly anti-tumor effects (44). In a subsequent study, the same authors observed that engineered Tr1 efficiently killed pediatric AML cell samples in vitro, suggesting that the adoptive transfer of such Tr1 could be performed alongside allo-HSCT, to prevent GVHD (45). In a Phase I clinical trial, the authors infused their IL-10-engineered donor T cell product in 12 patients. Despite achieving only partial control of GVHD, likely because of the small percentage of Tr1 cells in the product, 4 patients who attained immune reconstitution remained relapse-free for a median follow-up of 7.2 years (46). Currently, optimizations in the protocols for in vitro generation of Tr1 cells are being performed in order to increase Tr1 number and efficacy of the infusion product (47).
Other strategies are further being adopted that aim to enhance GVL after classical Foxp3+ Treg infusion. For instance, the co-administration of Treg with different substances, such as ruxolitinib, low-dose IL-2, IL-33, and rapamycin, are being tested. In fact, the effect of low-dose IL-2 after HSCT has been studied for decades, demonstrating the ability of this cytokine to modulate GVHD without impairing GVL (48–51). In recent clinical trials, the use of low-dose IL-2 prophylactically (52) or therapeutically in steroid-refractory cGVHD patients (53) did not impair GVL. Of note, in the latter case, low-dose IL-2 induced a preferential increase in Treg cell counts (53). Given such results, another trial combined low-dose IL-2 with donor-derived Treg cell therapy seeking to induce greater Treg expansion in vivo. Again, GVL was not abrogated since no relapse cases were reported. However, clinical improvement was only observed in 20% of patients, likely due to the advanced phase of cGVHD and to the low number of donor-Treg infused (27).
Using a mouse model, Meguri et al. (54) have shown that IL-2 therapy affects Treg and effector T cells (Teff) responses differently depending on the immune environment in the host. In mild inflammatory conditions, the IL-2 therapy controlled GVHD without affecting GVL. Moreover, their results indicate that in an immune-tolerant state after HSCT, IL-2 therapy may even enhance the GVL effect without exacerbating GVHD (54). More recently, adaptations to the previous approaches using IL-2 therapy have been performed aiming to specifically promote Treg expansion and not alloreactive effector T cells. For instance, orthogonal IL-2 specifically binds the ortho IL-2 receptor β-chain, which in turn was forced expressed on mouse Treg. These engineered Treg cells selectively expanded in vitro and in vivo in the presence of ortho IL-2 in an MHC-mismatched mouse model and were capable of suppressing aGVHD while maintaining GVL responses against A20 and MLL-AF9 cells, even when low cell numbers were infused (55). The authors suggest combining early Treg infusion (i.e. before Tcon in HSCT (20, 26)) with their orthogonal system to support Treg expansion in vivo (55). Another alternative improvement to the IL-2-based therapy is through the short administration of IL-2/anti-IL-2 complexes, which can be modulated to preferentially induce Treg over Teff cells. Thiolat et al. (56) found different benefits in this approach, such as prevention of GVHD development and reduction in leukemia-related death in mice, which is likely associated with the significant reduction of exhausted CD8+ T cell levels, in a mechanism that is partially mediated by CTLA-4.
Rapamycin (RAPA) selectively inhibits the mammalian target of rapamycin (mTOR), which affects T cell activation, proliferation, and differentiation, while preserving Treg function in which the mTOR pathway is constitutively inhibited (57). In NSG mice transplanted with human peripheral blood mononuclear cells (PBMC) and receiving RAPA daily for three weeks, GVHD was alleviated, and survival rates increased in comparison to the control group, that did not receive the inhibitor (58). RAPA decreased CD4+ and CD8+ T cell proliferation and apoptosis. Importantly, RAPA administration preserved GVL against the AML THP-1 line in primary and secondary transplants, where an increase in Treg proliferation was observed. In GVHD patients, RAPA treatment inhibits T cell proliferation, mainly in CD8+ T cells. RAPA further augments the anti-apoptotic protein BCL-2, Treg counts and CD25 expression in all T cell subsets. Such general increase of CD25 expression induced by RAPA poses a limitation in this approach when low-dose IL-2 therapy is considered, as it relies on the selectively high expression levels of CD25 Treg to expand Treg in vivo (58). Noteworthy, RAPA may also directly inhibit leukemia growth through mTOR inhibition. This is shown in a mouse model, where Zhang and colleagues observed that the concurrent administration of RAPA and IL-2 delayed the onset of leukemia (16).
Ruxolitinib is a Janus kinase (JAK) inhibitor with anti-inflammatory properties currently used in steroid-refractory acute and chronic GVHD. Studies in mice and humans suggested that ruxolitinib treatment per se is not associated with a higher risk of relapse compared to other immunosuppressors (59–62). Moreover, the combined treatment of ruxolitinib and human Treg administration in a mouse model suppressed GVHD without hampering GVL, despite the preferential activity of ruxolitinib in Treg over Tcon in vitro (63). Based on these results, the same group launched a clinical trial using donor Treg infusion in the treatment of cGVHD in patients with no improvement or partial responses to ruxolitinib (30).
Posttransplantation cyclophosphamide (PTCy) is frequently used as GVHD prophylaxis to induce tolerance after HLA-mismatched and HLA-matched allo-HSCT, minimizing the need for additional immunosuppression (64). PTCy mechanism of action was first associated with clonal deletion of early stimulated alloreactive T cells soon after the engraftment in mouse models. More recently, it was proposed that alloreactive T cells are not eliminated, but their expansion is constrained immediately after PTCy administration, while GVL responses occur afterwards (65). Moreover, the PTCy-mediated tolerance induction process seems to be dependent on donor Treg cells, as mice depleted of Foxp3+ Treg either before or after PTCy treatment exhibited accelerated acute GVHD development (66). However, studies in humanized mouse models did not find the presence of Treg mandatory for PTCy-induced GVHD inhibition, as PTCy significantly mitigated the xenogeneic GVHD even when human PBMC depleted of CD25+ cells were infused (67). On the other hand, mice receiving untouched PBMC showed improved survival rates than those in which Treg were depleted. Moreover, when untouched were used, GVL was decreased, but not abrogated (67). Overall, those studies point to an indispensable and nonredundant role for Treg in PTCy activity, which in turn does not disturb the GVL effect.
Noteworthy, studies in human allo-HSCT demonstrated that Treg cells recovered faster than Tcon after PTCy therapy, due to a higher expression of aldehyde dehydrogenase (ALDH) in Treg than in Tcon that seems to provide the former increased resistance to Cy, since ALDH is a major mechanism of Cy inactivation in vivo (68). In fact, analysis of patients undergoing allo-HSCT and PTCy have shown higher frequency of Treg cells 30 days after transplant compared to patients without PTCy. CD8+ T cells in five out of 23 patients who relapsed expressed less GZB and perforin than CD8+ T cells from patients in remission for a median follow-up of 13.1 months, suggesting a positive correlation between these markers and GVL (69). Similar to PTCy, azacytidine has been used to reduce GVHD without impairing GVL. Azacytidine also suppresses effector T cell proliferation but not Treg. Post-transplant azacytidine induced an increase in Treg cell numbers while promoting a CD8+ T-cell cytotoxic response in AML patients receiving three treatment cycles (70). In mice, azacytidine was able to convert Teff into T cells with a regulatory phenotype through hypomethylation of the Foxp3 promoter, resulting in augmented Foxp3 expression (71). As the correlation between Treg, GVL, PTCy or azacytidine has been explored more recently, additional studies are needed to unravel such complex associations.
To clarify the issue of Treg to Teff ratios in the GVHD/GVL balance, a recent study used a computational model to explore the bidirectional molecular interactions between Tregs and Teffs in allo-HSCT, as a core regulatory network that may be used as a strategy to enhance the GVL effect (72). The model predicted shifts in Tregs and Teffs numbers upon simultaneous blockade of CD25, TNFR2 and CTLA-4, suggesting this would favor GVL after allo-HSCT without causing GVHD. However, this model requires clinical testing (72). In fact, mouse studies in HLA-mismatched and matched settings have shown that the modulation of another member of the TNF receptor superfamily the TNFRSF25, alongside CD25 pathway, promotes the expansion of Treg in vivo. Treg up-regulated activation markers and showed enhanced suppressive function, that ameliorated GVHD while preserving GVL (73, 74). A subsequent study from the same group tested a mouse model of transplant that combines mobilized PB and in vivo Treg expansion by the TNFRSF25/CD25 pathway, in which the expanded Treg were essential for mediating GVHD suppression and did not interfere with GVL (75).
Alongside the use of therapeutic drugs that promote Treg expansion in vivo while preserving GVL, alternative cell-based approaches that use other suppressor cell populations are under investigation. In this setting, myeloid-derived suppressor cells (MDSC) have been shown to induce Treg cells in vivo and increase their suppressive function, while avoiding GVHD and maintaining GVL in mice (76, 77). An increasing body of evidence further shows that CD8+ Treg cells are able to attenuate GVHD and possess tumor-killing features (78, 79), raising the possibility of an approach consisting of the combined infusion of CD4+ and CD8+ Tregs in the future, as suggested by some authors (15, 80).
IL-33 is an IL-1 family member that is released upon tissue damage, inducing the activation of MDSC and Treg cells (81, 82). Indeed, in a mouse model of HSCT, Treg expanded ex vivo with IL-33 protected mice from GVHD in an amphiregulin (AREG) mediated way (83). In another study, in vivo administration of IL-33 from day -10 to 4 days after HSCT induced the expansion of ST2+ Treg, that were resistant to total body irradiation. In turn, those cells reduced effector T cell levels and controlled IL-33-driven aGVHD (84). Importantly, the blockade of the decoy IL-33 receptor, the soluble form of ST2, allowed free IL-33 to interact with membrane-bound ST2 expressed on Treg. This led to an increase in Treg frequency to the detriment of pathogenic Th17 cells, thus controlling GVHD in a mouse model. Nevertheless, the in vitro anti-tumoral cytotoxicity and the in vivo GVL activity were preserved, presumably because other CD4+ T cell subsets were less affected by the blockade in ST2 (85). IL-33 also promotes the expansion of a subset of IL-9-producing T cells that contribute to GVHD prevention. This is achieved through AREG production, thus maintaining effector T cell function, and preserving GVL (86).
Possible mechanisms by which GVL may be maintained upon adoptive Treg transfer for GVHD therapy
The reasons behind the ability of Treg to suppress allo-specific responses causing GVHD, while maintaining GVL, have been increasingly investigated and pointed out to homing discrepancies, the suppression mechanisms involved and their targets (Figure 1). Regarding the former, human PB Tregs preferentially activate at lymph nodes and migrate to GVHD target organs, such as skin, gut, liver, and lungs, but not to the blood marrow (BM) (26, 87). Thus, one possible theory postulates that GVHD suppression by Treg and the alloreactive T cell responses that control leukemia occurs in distinct tissues. In fact, most of the Treg present in human PB express CD45RO and low levels of CXCR4. This implies that once infused, such Treg are likely unable to home to the BM, restricting their suppressive activity to the periphery. This likely results in decreased GVHD while allowing Teff alloreactivity in the BM. This was tested in NSG mice receiving human leukemia, Treg and Tcon cells. These mice survived without cancer or GVHD, while this did not happen when the injected Treg expressed CD45RA and CXCR4. In that case, functional infused Tregs were found in the BM and mice died from the tumor progression, even though large doses of Tcon were given. When CXCR4 was blocked in this setting, however, Treg migration to the BM did not occur and GVL was preserved (88). Thus, these results suggest that infused Treg in the aforementioned clinical trials might not interfere with GVL due to their inability to migrate to the BM while suppressing GVHD responses that occur in the periphery.
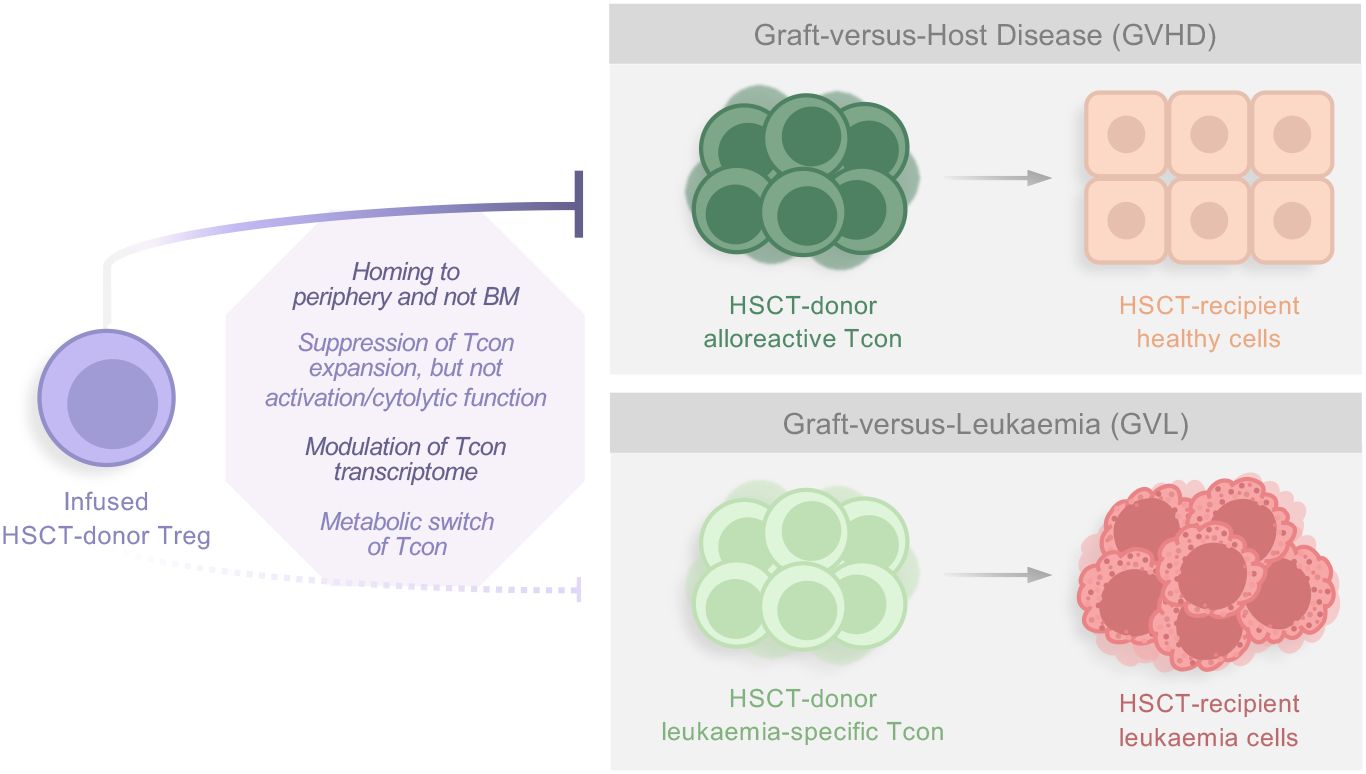
Figure 1 Factors influencing Treg suppressive activity against GVHD while preserving the GVL effect. Studies have shown that several parameters seem to be responsible for the ability of infused donor-Treg to suppress alloreactive Tcon responses causing GVHD, while not interfering with leukemia-specific Tcon, and thus maintaining GVL. In general, reasons include homing discrepancies, which suppression mechanisms are involved, and their targets. HSCT, Hematopoietic Stem Cell Transplantation; Treg, regulatory T cell; BM, blood marrow; Tcon, conventional T cell.
Recently, the same group analyzed human PB and BM samples at different time points after Treg/Tcon immunotherapy in haploidentical transplantation. Tolerogenic dendritic cells were found in PB, while pro-inflammatory dendritic cells and CD161+ Treg were detected in the BM. In vitro studies using induced CD161+ Treg isolated from healthy donors indicated that these cells favor Tcon-mediated killing capacity (89). As CD161+ Treg cells have been described to produce pro-inflammatory cytokines while still maintaining suppressor phenotype, it is possible that this cell population can even play a permissive role in GVL in vivo while actively participating in the anti-tumor effect. Such observations demand further investigation. Finally, the authors suggest that GVL is maintained due to the preferential migration of infused Treg to the periphery during the initial post-transplantation period, while the development of CD161+ Treg in the BM may contribute to the GVL effect afterwards (89).
Other studies have further shown that Tregs markedly suppress the expansion of alloreactive Tcon clones, but do not interfere with their activation, cytolytic function (12), differentiation or TCR repertoire (90). It was recently shown in a mouse model of MHC-mismatched allo-HCT, that infused Tregs are capable of modulating both pro- and anti-inflammatory gene transcription in both CD4+ and CD8+ Tcon, leaving the induction of GVL-related genes unaffected. Furthermore, this transcriptomics study indicated that Treg promotes a switch of Tcon metabolic activity from glycolysis to oxidative phosphorylation (90). Importantly, metabolic reprogramming has been studied as a strategy to maintain GVL (91, 92). Overall, these studies suggest possible mechanisms that may explain the lack of interference of infused Treg in GVL in the aforementioned human clinical trials. However, future trials should conduct more analyses in relapsed patients to rule out an effect of the infused Treg in this outcome.
Conclusions and future perspectives
In the past decade, several studies in humans and animal models have attempted to determine the impact of Treg infusions to treat GVHD on tumor relapse. The evidence gathered appears to suggest that the infusion of donor Treg does not interfere with the ability of donor T cells to clear tumor cells, thus preserving GVL. More studies are required to determine the therapeutic efficacy of such infusions. Noteworthy, the development of more potent Treg must consider the ability of these cells to spare leukemia-specific immune responses. Therefore, it is fundamental that future work in this field aims to develop improved sources of Treg for infusion while concomitantly assessing their suppressive activity against GVHD responses while preserving GVL.
Author contributions
CP: Writing – original draft. MS: Writing – review & editing. JL: Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by Fundação para a Ciência e Tecnologia, Portugal under the project PTDC/MEC-HEM/5281/2020 (DOI: 10.54499/PTDC/MEC-HEM/5281/2020). CP has been funded by a fellowship from Fundação para a Ciência e Tecnologia (2022.12709.BD) and has received the support of a fellowship from “la Caixa” Foundation (ID 100010434). This fellowship code is “LCF/BQ/DI19/11730053”.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Wing K, Sakaguchi S. Regulatory T cells exert checks and balances on self tolerance and autoimmunity. Nat Immunol. (2010) 11:7–13. doi: 10.1038/ni.1818
2. Sakaguchi S, Yamaguchi T, Nomura T, Ono M. Regulatory T cells and immune tolerance. Cell. (2008) 133:775–87. doi: 10.1016/j.cell.2008.05.009
3. Vignali DAA, Collison LW, Workman CJ. How regulatory T cells work. Nat Rev Immunol. (2008) 8:523–32. doi: 10.1038/nri2343
4. Zorn E, Kim HT, Lee SJ, Floyd BH, Litsa D, Arumugarajah S, et al. Reduced frequency of FOXP3+ CD4+CD25+ regulatory T cells in patients with chronic graft-versus-host disease. Blood. (2005) 106:2903–11. doi: 10.1182/blood-2005-03-1257
5. Zhai Z, Sun Z, Li Q, Zhang A, Liu H, Xu J, et al. Correlation of the CD4+CD25high T-regulatory cells in recipients and their corresponding donors to acute GVHD. Transplant Int. (2007) 20:440–6. doi: 10.1111/j.1432-2277.2007.00462.x
6. Matsuoka KI, Kim HT, McDonough S, Bascug G, Warshauer B, Koreth J, et al. Altered regulatory T cell homeostasis in patients with CD4+ lymphopenia following allogeneic hematopoietic stem cell transplantation. J Clin Invest. (2010) 120:1479–93. doi: 10.1172/JCI41072
7. Alho AC, Kim HT, Chammas MJ, Reynolds CG, Matos TR, Forcade E, et al. Unbalanced recovery of regulatory and effector T cells after allogeneic stem cell transplantation contributes to chronic GVHD. Blood. (2016) 127:646–57. doi: 10.1182/blood-2015-10-672345
8. Soares MV, Azevedo RI, Ferreira IA, Bucar S, Ribeiro AC. Naive and stem cell memory T cell subset recovery reveals opposing reconstitution patterns in CD4 and CD8 T cells in chronic graft vs. Host disease. Front Immunol. (2019) 10:334. doi: 10.3389/fimmu.2019.00334
9. Miura Y, Thoburn CJ, Bright EC, Phelps ML, Shin T, Matsui EC, et al. Association of Foxp3 regulatory gene expression with graft-versus-host disease. Blood. (2004) 104:2187–93. doi: 10.1182/blood-2004-03-1040
10. Rezvani K, Mielke S, Ahmadzadeh M, Kilical Y, Savani BN, Zeilah J, et al. High donor FOXP3-positive regulatory T-cell (Treg) content is associated with a low risk of GVHD following HLA-matched allogeneic SCT. Blood. (2006) 108:1291–7. doi: 10.1182/blood-2006-02-003996
11. Magenau JM, Qin X, Tawara I, Rogers CE, Kitko C, Schlough M, et al. Frequency of CD4+CD25hiFOXP3+ Regulatory T cells has diagnostic and prognostic value as a biomarker for acute graft-versus-host-disease. Biol Blood Marrow Transplant. (2010) 16:907–14. doi: 10.1016/j.bbmt.2010.02.026
12. Edinger M, Hoffmann P, Ermann J, Drago K, Fathman CG, Strober S, et al. CD4+CD25+ regulatory T cells preserve graft-versus-tumor activity while inhibiting graft-versus-host disease after bone marrow transplantation. Nat Med. (2003) 9:1144–50. doi: 10.1038/nm915
13. Jones SC, Murphy GF, Korngold R. Post-hematopoietic cell transplantation control of graft-versus-host disease by donor CD4+25+ T cells to allow an effective graft-versus-leukemia response. Biol Blood Marrow Transplant. (2003) 9:243–56. doi: 10.1053/bbmt.2003.50027
14. Trenado A, Charlotte F, Fisson S, Yagello M, Klatzmann D, Salomon BL, et al. Recipient-type specific CD4+CD25+ regulatory T cells favor immune reconstitution and control graft-versus-host disease while maintaining graft-versus-leukemia. J Clin Invest. (2003) 112:1688–96. doi: 10.1172/JCI17702
15. Heinrichs J, Li J, Nguyen H, Wu Y, Bastian D, Daethanasanmak A, et al. CD8+ Tregs promote GVHD prevention and overcome the impaired GVL effect mediated by CD4+ Tregs in mice. Oncoimmunology. (2016) 5:1–11. doi: 10.1080/2162402X.2016.1146842
16. Zhang P, Tey S-K, Koyama M, Kuns RD, Olver SD, Lineburg KE, et al. Induced regulatory T cells promote tolerance when stabilized by rapamycin and IL-2 in vivo. J Immunol. (2013) 191:5291–303. doi: 10.4049/jimmunol.1301181
17. Martelli MF, Di Ianni M, Ruggeri L, Falzetti F, Carotti A, Terenzi A, et al. HLA-haploidentical transplantation with regulatory and conventional T-cell adoptive immunotherapy prevents acute leukemia relapse. Blood. (2014) 124:638–44. doi: 10.1182/blood-2014-03-564401
18. Nadal E, Garin M, Kaeda J, Apperley J, Lechler R, Dazzi F. Increased frequencies of CD4+CD25high Tregs correlate with disease relapse after allogeneic stem cell transplantation for chronic myeloid leukemia. Leukemia. (2007) 21:472–9. doi: 10.1038/sj.leu.2404522
19. Wolf D, Wolf AM, Fong D, Rumpold H, Strasak A, Clausen J, et al. Regulatory T-cells in the graft and the risk of acute graft-versus-host disease after allogeneic stem cell transplantation. Transplantation. (2007) 83:1107–13. doi: 10.1097/01.tp.0000260140.04815.77
20. Meyer EH, Laport G, Xie BJ, MacDonald K, Heydari K, Sahaf B, et al. Transplantation of donor grafts with defined ratio of conventional and regulatory T cells in HLA-matched recipients. JCI Insight. (2019) 4:e127244. doi: 10.1172/jci.insight.127244
21. Di Ianni M, Falzetti F, Carotti A, Terenzi A, Castellino F, Bonifacio E, et al. Tregs prevent GVHD and promote immune reconstitution in HLA-haploidentical transplantation. Blood. (2011) 117:3921–8. doi: 10.1182/blood-2010-10-311894
22. Pierini A, Ruggeri L, Carotti A, Falzetti F, Saldi S, Terenzi A, et al. Haploidentical age-adapted myeloablative transplant and regulatory and effector T cells for acute myeloid leukemia. Blood Adv. (2021) 5:1199–208. doi: 10.1182/bloodadvances.2020003739
23. Brunstein CG, Miller JS, Cao Q, McKenna DH, Hippen KL, Curtsinger J, et al. Infusion of ex vivo expanded T regulatory cells in adults transplanted with umbilical cord blood: Safety profile and detection kinetics. Blood. (2011) 117:1061–70. doi: 10.1182/blood-2010-07-293795
24. Brunstein CG, Miller JS, McKenna DH, Hippen KL, DeFor TE, Sumstad D, et al. Umbilical cord blood-derived T regulatory cells to prevent GVHD: Kinetics, toxicity profile, and clinical effect. Blood. (2016) 127:1044–51. doi: 10.1182/blood-2015-06-653667
25. MacMillan ML, Hippen KL, McKenna DH, Kadidlo D, Sumstad D, Defor TE, et al. First-in-human phase 1 trial of induced regulatory T cells for graft-versus-host disease prophylaxis in HLA-matched siblings. Blood Adv. (2021) 5:1425–36. doi: 10.1182/bloodadvances.2020003219
26. Nguyen VH, Zeiser R, DaSilva DL, Chang DS, Beilhack A, Contag CH, et al. In vivo dynamics of regulatory T-cell trafficking and survival predict effective strategies to control graft-versus-host disease following allogeneic transplantation. Blood. (2007) 109:2649–56. doi: 10.1182/blood-2006-08-044529
27. Whangbo JS, Nikiforow S, Kim HT, Wahl J, Reynolds CG, Rai SC, et al. A phase 1 study of donor regulatory T-cell infusion plus low-dose interleukin-2 for steroid-refractory chronic graft-vs-host disease. Blood Adv. (2022) 6:5786–96. doi: 10.1182/bloodadvances.2021006625
28. Trzonkowski P, Bieniaszewska M, Juścińska J, Dobyszuk A, Krzystyniak A, Marek N, et al. First-in-man clinical results of the treatment of patients with graft versus host disease with human ex vivo expanded CD4+CD25+CD127- T regulatory cells. Clin Immunol. (2009) 133:22–6. doi: 10.1016/j.clim.2009.06.001
29. Theil A, Tuve S, Oelschlägel U, Maiwald A, Döhler D, Oßmann D, et al. Adoptive transfer of allogeneic regulatory T cells into patients with chronic graft-versus-host disease. Cytotherapy. (2015) 17:473–86. doi: 10.1016/j.jcyt.2014.11.005
30. Soares MVD, Escamilla Gomez V, Azevedo RI, Pereira PNG, Velázquez TC, Garcia-Calderón CB, et al. Phase I/II clinical trials of donor-derived purified regulatory T cells for the treatment of steroid-refractory chronic graft versus host disease. Blood. (2022) 140:880–2. doi: 10.1182/blood-2022-163394
31. Landwehr-Kenzel S, Müller-Jensen L, Kuehl JS, Abou-el-Enein M, Hoffmann H, Muench S, et al. Adoptive transfer of ex vivo expanded regulatory T cells improves immune cell engraftment and therapy-refractory chronic GvHD. Mol Ther. (2022) 30:2298–314. doi: 10.1016/j.ymthe.2022.02.025
32. Trenado A, Sudres M, Tang Q, Maury S, Charlotte F, Grégoire S, et al. Ex vivo-expanded CD4+CD25+ Immunoregulatory T cells prevent graft-versus-host-disease by inhibiting activation/differentiation of pathogenic T cells. J Immunol. (2006) 176:1266–73. doi: 10.4049/jimmunol.176.2.1266
33. Sagoo P, Ali N, Garg G, Nestle FO, Lechler RI, Lombardi G. Human regulatory T cells with alloantigen specificity are more potent inhibitors of alloimmune skin graft damage than polyclonal regulatory T cells. Sci Transl Med. (2011) 3:83ra42. doi: 10.1126/scitranslmed.3002076
34. Sela U, Olds P, Park A, Schlesinger SJ, Steinman RM. Dendritic cells induce antigen-specific regulatory T cells that prevent graft versus host disease and persist in mice. J Exp Med. (2011) 208:2489–96. doi: 10.1084/jem.20110466
35. Veerapathran A, Pidala J, Beato F, Betts B, Kim J, Turner JG, et al. Human regulatory T cells against minor histocompatibility antigens: Ex vivo expansion for prevention of graft-versus-host disease. Blood. (2013) 122:2251–61. doi: 10.1182/blood-2013-03-492397
36. Li J, Heinrichs J, Haarberg K, Semple K, Veerapathran A, Liu C, et al. HY-specific induced regulatory T cells display high specificity and efficacy in the prevention of acute graft-versus-host disease. J Immunol. (2015) 195:717–25. doi: 10.4049/jimmunol.1401250
37. Randolph SSB, Gooley TA, Warren EH, Appelbaum FR, Riddell SR. Female donors contribute to a selective graft-versus-leukemia effect in male recipients of HLA-matched, related hematopoietic stem cell transplants. Blood. (2004) 103:347–52. doi: 10.1182/blood-2003-07-2603
38. Veerapathran A, Pidala J, Beato F, Yu X-Z, Anasetti C. Ex vivo expansion of human Tregs specific for alloantigens presented directly or indirectly. Blood. (2011) 118:5671–80. doi: 10.1182/blood-2011-02
39. MacDonald KG, Hoeppli RE, Huang Q, Gillies J, Luciani DS, Orban PC, et al. Alloantigen-specific regulatory T cells generated with a chimeric antigen receptor. J Clin Invest. (2016) 126:1413–24. doi: 10.1172/JCI82771
40. Bolivar-Wagers S, Loschi ML, Jin S, Thangavelu G, Larson JH, McDonald-Hyman CS, et al. Murine CAR19 Tregs suppress acute graft-versus-host disease and maintain graft-versus-tumor responses. JCI Insight. (2022) 7:1–19. doi: 10.1172/jci.insight.160674
41. Imura Y, Ando M, Kondo T, Ito M, Yoshimura A. CD19-targeted CAR regulatory T cells suppress B cell pathology without GvHD. JCI Insight. (2020) 5:e136185. doi: 10.1172/JCI.INSIGHT.136185
42. Boroughs AC, Larson RC, Choi BD, Bouffard AA, Riley LS, Schiferle E, et al. Chimeric antigen receptor costimulation domains modulate human regulatory T cell function. JCI Insight. (2019) 4:e126194. doi: 10.1172/jci.insight.126194
43. Rodgers DT, Mazagova M, Hampton EN, Cao Y, Ramadoss NS, Hardy IR, et al. Switch-mediated activation and retargeting of CAR-T cells for B-cell Malignancies. Proc Natl Acad Sci U.S.A. (2016) 113:E459–68. doi: 10.1073/pnas.1524155113
44. Locafaro G, Andolfi G, Russo F, Cesana L, Spinelli A, Camisa B, et al. IL-10-engineered human CD4+ Tr1 cells eliminate myeloid leukemia in an HLA class I-dependent mechanism. Mol Ther. (2017) 25:2254–69. doi: 10.1016/j.ymthe.2017.06.029
45. Cieniewicz B, Uyeda MJ, Chen P, Sayitoglu EC, Liu JMH, Andolfi G, et al. Engineered type 1 regulatory T cells designed for clinical use kill primary pediatric acute myeloid leukemia cells. Haematologica. (2021) 106:2588–97. doi: 10.3324/haematol.2020.263129
46. Bacchetta R, Lucarelli B, Sartirana C, Gregori S, Lupo Stanghellini MT, Miqueu P, et al. Immunological outcome in haploidentical-HSC transplanted patients treated with IL-10-anergized donor T Cells. Front Immunol. (2014) 5:16. doi: 10.3389/fimmu.2014.00016
47. Chen PP, Cepika AM, Agarwal-Hashmi R, Saini G, Uyeda MJ, Louis DM, et al. Alloantigen-specific type 1 regulatory T cells suppress through CTLA-4 and PD-1 pathways and persist long-term in patients. Sci Transl Med. (2021) 13:eabf5264. doi: 10.1126/scitranslmed.abf5264
48. Sykes M, Romick ML, Sachs DH. Interleukin 2 prevents graft-versus-host disease while preserving the graft-versus-leukemia effect of allogeneic T cells. Proc Natl Acad Sci U.S.A. (1990) 87:5633–7. doi: 10.1073/pnas.87.15.5633
49. Matsuoka KI, Koreth J, Kim HT, Bascug G, McDonough S, Kawano Y, et al. Low-dose interleukin-2 therapy restores regulatory T cell homeostasis in patients with chronic graft-versus-host disease. Sci Transl Med. (2013) 5:1–12. doi: 10.1126/scitranslmed.3005265
50. Hirakawa M, Matos TR, Liu H, Koreth J, Kim HT, Paul NE, et al. Low-dose IL-2 selectively activates subsets of CD4+ Tregs and NK cells. JCI Insight. (2016) 1:e89278. doi: 10.1172/jci.insight.89278
51. Soiffer RJ, Murray C, Gonin R, Ritz J. Effect of low-dose interleukin-2 on disease relapse after T-cell- depleted allogeneic bone marrow transplantation. Blood. (1994) 84:964–71. doi: 10.1182/blood.V84.3.964.964
52. Kennedy-Nasser AA, Ku S, Castillo-Caro P, Hazrat Y, Wu MF, Liu H, et al. Ultra low-dose IL-2 for GVHD prophylaxis after allogeneic hematopoietic stem cell transplantation mediates expansion of regulatory t cells without diminishing antiviral and antileukemic activity. Clin Cancer Res. (2014) 20:2215–25. doi: 10.1158/1078-0432.CCR-13-3205
53. Koreth J, Kim HT, Jones KT, Lange PB, Reynolds CG, Chammas MJ, et al. Efficacy, durability, and response predictors of low-dose interleukin-2 therapy for chronic graft-versus-host disease. Blood. (2016) 128:130–7. doi: 10.1182/blood-2016-02
54. Meguri Y, Asano T, Yoshioka T, Iwamoto M, Ikegawa S, Sugiura H, et al. Responses of regulatory and effector T-cells to low-dose interleukin-2 differ depending on the immune environment after allogeneic stem cell transplantation. Front Immunol. (2022) 13:891925. doi: 10.3389/fimmu.2022.891925
55. Ramos TL, Bolivar-Wagers S, Jin S, Thangavelu G, Simonetta F, Lin PY, et al. Prevention of acute GVHD using an orthogonal IL-2/IL-2Rβ system to selectively expand regulatory T cells. vivo. Blood. (2023) 141:1337–52. doi: 10.1182/blood.2022018440
56. Thiolat A, Pilon C, Caudana P, Moatti A, To NH, Sedlik C, et al. Treg-targeted IL-2/anti-IL-2 complex controls graft-versus-host disease and supports anti-tumor effect in allogeneic hematopoietic stem cell transplantation. Haematologica. (2024) 109:129–42. doi: 10.3324/haematol.2022.282653
57. Zeiser R, Leveson-Gower DB, Zambricki EA, Kambham N, Beilhack A, Loh J, et al. Differential impact of mammalian target of rapamycin inhibition on CD4 CD25 Foxp3 regulatory T cells compared with conventional CD4 T cells. Blood. (2008) 111:453–62. doi: 10.1182/blood-2007-06-094482
58. Ehx G, Ritacco C, Hannon M, Dubois S, Delens L, Willems E, et al. Comprehensive analysis of the immunomodulatory effects of rapamycin on human T cells in graft-versus-host disease prophylaxis. Am J Transplant. (2021) 21:2662–74. doi: 10.1111/ajt.16505
59. Choi J, Cooper ML, Alahmari B, Ritchey J, Collins L, Holt M, et al. Pharmacologic blockade of JAK1/JAK2 reduces GvHD and preserves the graft-versus-leukemia effect. PloS One. (2014) 9:e109799. doi: 10.1371/journal.pone.0109799
60. Carniti C, Gimondi S, Vendramin A, Recordati C, Confalonieri D, Bermema A, et al. Pharmacologic inhibition of JAK1/JAK2 signaling reduces Experimental murine acute GVHD while preserving GVT effects. Clin Cancer Res. (2015) 21:3740–9. doi: 10.1158/1078-0432.CCR-14-2758
61. Zeiser R, Burchert A, Lengerke C, Verbeek M, Maas-Bauer K, Metzelder SK, et al. Ruxolitinib in corticosteroid-refractory graft-versus-host disease after allogeneic stem cell transplantation: a multicenter survey. Leukemia. (2015) 29:2062–8. doi: 10.1038/leu.2015.212
62. Zeiser R, von Bubnoff N, Butler J, Mohty M, Niederwieser D, Or R, et al. Ruxolitinib for glucocorticoid-refractory acute graft-versus-host disease. New Engl J Med. (2020) 382:1800–10. doi: 10.1056/nejmoa1917635
63. Rodríguez-Gil A, Escamilla-Gómez V, Nufer M, Andújar-Sánchez F, Lopes-Ramos T, Bejarano-García JA, et al. Combined treatment of graft versus host disease using donor regulatory T cells and ruxolitinib. Sci Rep. (2022) 12:1–13. doi: 10.1038/s41598-022-12407-x
64. Kanakry CG, Bolaños-Meade J, Kasamon YL, Zahurak M, Durakovic N, Furlong T, et al. Low immunosuppressive burden after HLA-matched related or unrelated BMT using posttransplantation cyclophosphamide. Blood. (2017) 129:1389–93. doi: 10.1182/blood-2016-09-737825
65. Wachsmuth LP, Patterson MT, Eckhaus MA, Venzon DJ, Gress RE, Kanakry CG. Posttransplantation cyclophosphamide prevents graft-versus-host disease by inducing alloreactive T cell dysfunction and suppression. J Clin Invest. (2019) 129:2357–73. doi: 10.1172/JCI124218
66. Ganguly S, Ross DB, Panoskaltsis-Mortari A, Kanakry CG, Blazar BR, Levy RB, et al. Donor CD4+Foxp3+regulatory T cells are necessary for posttransplantation cyclophosphamide-mediated protection against GVHD in mice. Blood. (2014) 124:2131–41. doi: 10.1182/blood-2013-10-525873
67. Ritacco C, Köse MC, Courtois J, Canti L, Beguin C, Dubois S, et al. Post-transplant cyclophosphamide prevents xenogeneic graft-versus-host disease while depleting proliferating regulatory T cells. iScience. (2023) 26:106085. doi: 10.1016/j.isci.2023.106085
68. Kanakry CG, Ganguly S, Zahurak M, Bolaños-Meade J, Thoburn C, Perkins B, et al. Aldehyde dehydrogenase expression drives human regulatory T cell resistance to posttransplantation cyclophosphamide. Sci Transl Med. (2013) 5:211ra157. doi: 10.1126/scitranslmed.3006960
69. Zhao C, Bartock M, Jia B, Shah N, Claxton DF, Wirk B, et al. Post-transplant cyclophosphamide alters immune signatures and leads to impaired T cell reconstitution in allogeneic hematopoietic stem cell transplant. J Hematol Oncol. (2022) 15:1–7. doi: 10.1186/s13045-022-01287-3
70. Goodyear OC, Dennis M, Jilani NY, Loke J, Siddique S, Ryan G, et al. Azacitidine augments expansion of regulatory T cells after allogeneic stem cell transplantation in patients with acute myeloid leukemia (AML). Blood. (2012) 119:3361–9. doi: 10.1182/blood-2011-09-377044
71. Cooper ML, Choi J, Karpova D, Vij K, Ritchey J, Schroeder MA, et al. Azacitidine mitigates graft-versus-host disease via differential effects on the proliferation of T effectors and natural regulatory T cells In Vivo. J Immunol. (2017) 198:3746–54. doi: 10.4049/jimmunol.1502399
72. Nishiyama N, Ruoff P, Jimenez JC, Miwakeichi F, Nishiyama Y, Yata T. Modeling the interaction between donor-derived regulatory T cells and effector T cells early after allogeneic hematopoietic stem cell transplantation. BioSystems. (2023) 227–228:104889. doi: 10.1016/j.biosystems.2023.104889
73. Copsel S, Wolf D, Kale B, Barreras H, Lightbourn CO, Bader CS, et al. Very low numbers of CD4+ FoxP3+ Tregs expanded in donors via TL1A-ig and low-dose IL-2 exhibit a distinct activation/functional profile and suppress GVHD in a preclinical model. Biol Blood Marrow Transplant. (2018) 24:1788–94. doi: 10.1016/j.bbmt.2018.04.026
74. Wolf D, Barreras H, Bader CS, Copsel S, Lightbourn CO, Pfeiffer BJ, et al. Marked in Vivo Donor Regulatory T Cell Expansion via Interleukin-2 and TL1A-Ig Stimulation Ameliorates Graft-versus-Host Disease but Preserves Graft-versus-Leukemia in Recipients after Hematopoietic Stem Cell Transplantation. Biol Blood Marrow Transplant. (2017) 23:757–66. doi: 10.1016/j.bbmt.2017.02.013
75. Barreras H, Copsel SN, Bader CS, Ding Y, Wolf D, Cash C, et al. Regulatory T cell amelioration of graft-versus-host disease following allogeneic/xenogeneic hematopoietic stem cell transplantation using mobilized mouse and human peripheral blood donors. Transplant Cell Therapy Off Publ Am Soc Transplant Cell Ther. (2023) 29:341.e1–9. doi: 10.1016/j.jtct.2023.02.015
76. Janikashvili N, Gérard C, Thébault M, Brazdova A, Boibessot C, Cladière C, et al. Efficiency of human monocyte-derived suppressor cell-based treatment in graft-versus-host disease prevention while preserving graft-versus-leukemia effect. Oncoimmunology. (2021) 10:1880046. doi: 10.1080/2162402X.2021.1880046
77. Zhang J, Chen HM, Ma G, Zhou Z, Raulet D, Rivera AL, et al. The mechanistic study behind suppression of GVHD while retaining GVL activities by myeloid-derived suppressor cells. Leukemia. (2019) 33:2078–89. doi: 10.1038/s41375-019-0394-z
78. Beres AJ, Haribhai D, Chadwick AC, Gonyo PJ, Williams CB, Drobyski WR. CD8+ Foxp3+ Regulatory T cells are induced during graft-versus-host disease and mitigate disease severity. J Immunol. (2012) 189:464–74. doi: 10.4049/jimmunol.1200886
79. Iamsawat S, Daenthanasanmak A, Voss JH, Nguyen H, Bastian D, Liu C, et al. Stabilization of foxp3 by targeting JAK2 enhances efficacy of CD8 induced regulatory T cells in the prevention of graft-versus-host disease. J Immunol. (2018) 201:2812–23. doi: 10.4049/jimmunol.1800793
80. Bolivar-Wagers S, Larson JH, Jin S, Blazar BR. Cytolytic CD4+ and CD8+ Regulatory T-cells and implications for developing immunotherapies to combat graft-versus-host disease. Front Immunol. (2022) 13:864748. doi: 10.3389/fimmu.2022.864748
81. Matta BM, Lott JM, Mathews LR, Liu Q, Rosborough BR, Blazar BR, et al. IL-33 is an unconventional alarmin that stimulates IL-2 secretion by dendritic cells to selectively expand IL-33R/ST2+ Regulatory T cells. J Immunol. (2014) 193:4010–20. doi: 10.4049/jimmunol.1400481
82. Turnquist HR, Zhao Z, Rosborough BR, Liu Q, Castellaneta A, Isse K, et al. IL-33 expands suppressive CD11b+ Gr-1int and regulatory T cells, including ST2L+ Foxp3+ Cells, and mediates regulatory T cell-dependent promotion of cardiac allograft survival. J Immunol. (2011) 187:4598–610. doi: 10.4049/jimmunol.1100519
83. Yang J, Ramadan A, Reichenbach DK, Loschi M, Zhang J, Griesenauer B, et al. Rorc restrains the potency of ST2+ regulatory T cells in ameliorating intestinal graft-versus-host disease. JCI Insight. (2019) 4:1–14. doi: 10.1172/jci.insight.122014
84. Matta BM, Reichenbach DK, Zhang X, Mathews L, Koehn BH, Dwyer GK, et al. Peri-alloHCT IL-33 administration expands recipient T-regulatory cells that protect mice against acute GVHD. Blood. (2016) 128:427–39. doi: 10.1182/blood-2015-12-684142
85. Zhang J, Ramadan AM, Griesenauer B, Li W, Turner MJ, Liu C, et al. ST2 blockade reduces sST2-producing T cells while maintaining protective mST2-expressing T cells during graft-versus-host disease. Sci Transl Med. (2015) 7:1–13. doi: 10.1126/scitranslmed.aab0166
86. Ramadan A, Griesenauer B, Adom D, Kapur R, Hanenberg H, Liu C, et al. Specifically differentiated T cell subset promotes tumor immunity over fatal immunity. J Exp Med. (2017) 214:3577–96. doi: 10.1084/jem.20170041
87. Nguyen VH, Shashidhar S, Chang DS, Ho L, Kambham N, Bachmann M, et al. The impact of regulatory T cells on T-cell immunity following hematopoietic cell transplantation. Blood. (2008) 111:945–53. doi: 10.1182/blood-2007-07-103895
88. Ruggeri L, Carotti A, Pierini A, Falzetti F, Terenzi A, Urbani E, et al. How Adoptive Immunotherapy with Conventional T and Regulatory T Cells Exerts a Gvl Effect without GvHD, after Haploidentical Hematopoietic Transplantation. Blood. (2018) 132:3333–3. doi: 10.1182/blood-2018-99-112512
89. Guardalupi F, Sorrentino C, Corradi G, Giancola R, Baldoni S, Ulbar F, et al. A pro-inflammatory environment in bone marrow of Treg transplanted patients matches with graft-versus-leukemia effect. Leukemia. (2023) 37:1572–5. doi: 10.1038/s41375-023-01932-x
90. Lohmeyer JK, Hirai T, Turkoz M, Buhler S, Lopes Ramos T, Köhler N, et al. Analysis of the T-cell repertoire and transcriptome identifies mechanisms of regulatory T-cell suppression of GVHD. Blood. (2023) 141:1755–67. doi: 10.1182/blood.2022017982
91. Huang Y, Zou Y, Jiao Y, Shi P, Nie X, Huang W, et al. Targeting glycolysis in alloreactive T cells to prevent acute graft-versus- host disease while preserving graft- versus-leukemia effect. Front Immunol. (2022) 13:751296. doi: 10.3389/fimmu.2022.751296
Keywords: graft-versus-leukemia, regulatory T cell, immunomodulation, GvHD, cell therapy, hematopoietic stem cell transplantation
Citation: Pacini CP, Soares MVD and Lacerda JF (2024) The impact of regulatory T cells on the graft-versus-leukemia effect. Front. Immunol. 15:1339318. doi: 10.3389/fimmu.2024.1339318
Received: 15 November 2023; Accepted: 08 April 2024;
Published: 22 April 2024.
Edited by:
Mahasweta Gooptu, Dana–Farber Cancer Institute, United StatesReviewed by:
Govindarajan Thangavelu, University of Minnesota Twin Cities, United StatesSawa Ito, University of Pittsburgh, United States
Copyright © 2024 Pacini, Soares and Lacerda. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: João F. Lacerda, jlacerda@medicina.ulisboa.pt