 Tirza Gabrielle Ramos de Mesquita1,2
Tirza Gabrielle Ramos de Mesquita1,2 José do Espírito Santo Junior3,4
José do Espírito Santo Junior3,4 Luan Diego Oliveira da Silva4
Luan Diego Oliveira da Silva4 George Allan Villarouco Silva3Felipe Jules de Araújo1,4Suzana Kanawati Pinheiro1,2Herllon Karllos Athaydes Kerr1Lener Santos da Silva1,2Luciane Macedo de Souza1Samir Assad de Almeida4Krys Layane Guimarães Duarte Queiroz2Josué Lacerda de Souza3,4Cilana Chagas da Silva2Héctor David Graterol Sequera1,2Mara Lúcia Gomes de Souza1,2
George Allan Villarouco Silva3Felipe Jules de Araújo1,4Suzana Kanawati Pinheiro1,2Herllon Karllos Athaydes Kerr1Lener Santos da Silva1,2Luciane Macedo de Souza1Samir Assad de Almeida4Krys Layane Guimarães Duarte Queiroz2Josué Lacerda de Souza3,4Cilana Chagas da Silva2Héctor David Graterol Sequera1,2Mara Lúcia Gomes de Souza1,2 Anderson Nogueira Barbosa5
Anderson Nogueira Barbosa5 Gemilson Soares Pontes3,5,6Marcus Vinitius de Farias Guerra1,2
Gemilson Soares Pontes3,5,6Marcus Vinitius de Farias Guerra1,2 Rajendranath Ramasawmy1,2,4,6*
Rajendranath Ramasawmy1,2,4,6*- 1Programa de Pós-Graduação em Medicina Tropical, Universidade do Estado do Amazonas, Manaus, Brazil
- 2Department of Molecular Biology, Fundação de Medicina Tropical Doutor Heitor Vieira Dourado, Manaus, Brazil
- 3Programa de Pós-Graduação em Imunologia Básica e Aplicada, Instituto de Ciências Biológicas, Universidade Federal do Amazonas, Manaus, Amazonas, Brazil
- 4Faculdade de Medicina Nilton Lins, Universidade Nilton Lins, Manaus, Brazil
- 5Department of Virology, Instituto Nacional de Pesquisas da Amazônia, Manaus, Brazil
- 6Genomic Health Surveillance Network: Optimization of Assistance and Research in The State of Amazonas – REGESAM, Manaus, Amazonas, Brazil
The immunopathology associated with Leishmaniasis is a consequence of inflammation. Upon infection with Leishmania, the type of host-immune response is determinant for the clinical manifestations that can lead to either self-healing or chronic disease. Multiple pathways may determine disease severity. A comparison of systemic immune profiles in patients with cutaneous leishmaniasis caused by L. guyanensis and healthy individuals with the same socio-epidemiological characteristics coming from the same endemic areas as the patients is performed to identify particular immune profile and pathways associated with the progression of disease development. Twenty-seven plasma soluble circulating factors were evaluated between the groups by univariate and multivariate analysis. The following biomarkers pairs IL-17/IL-9 (ρ=0,829), IL-17/IL-12 (ρ=0,786), IL-6/IL-1ra (ρ=0,785), IL-6/IL-12 (ρ=0,780), IL-1β/G-CSF (ρ=0,758) and IL-17/MIP-1β (ρ=0,754) showed the highest correlation mean among the patient while only INF-γ/IL-4 (ρ=0.740), 17/MIP-1β (ρ=0,712) and IL-17/IL-9 (ρ=0,707) exhibited positive correlation among the control group. The cytokine IL-17 and IL1β presented the greater number of positive pair correlation among the patients. The linear combinations of biomarkers displayed IP-10, IL-2 and RANTES as the variables with the higher discriminatory activity in the patient group compared to PDGF, IL-1ra and eotaxin among the control subjects. IP-10, IL-2, IL-1β, RANTES and IL-17 seem to be predictive value of progression to the development of disease among the Lg-infected individuals.
Introduction
Leishmaniasis, a vector-borne infectious disease caused by Leishmania spp (L.), continues to be a public health burden in over 98 countries worldwide and still is a neglected tropical disease (1). L.-infected individuals can manifest a wide spectrum of clinical symptoms that is guided by the immunological status and the genetic background of the individual, the L. spp, and the environment. The disease outcome may range from asymptomatic, localized cutaneous lesions (cutaneous leishmaniasis (CL), severe mucosal lesions (mucosal Leishmaniasis (ML) and to life threatening visceral leishmaniasis (VL). CL and ML are also known as American Tegumentary leishmaniasis (ATL) in the American continent.
The major species that cause ATL in Brazil are L. braziliensis (Lb), L. guyanensis (Lg), L. lainsoni, L. amazonensis, L. shawi, L. naiffi and L. lindenbergi. L. braziliensis is responsible for the majority of the cases (2). However, in the Amazonas, L. guyanensis is the main etiological agent of ATL and represent 95% of the CL cases (3).
Different pathophysiological mechanisms are suggested to lead to the development of the different clinical manifestations of leishmaniasis and still, treatment with antimonials targeting the parasite continues to be the drug of choice with no headway in the development of new therapies (4). Pentavalent antimony (Sbv) is still the current first-line treatment in Brazil for CL and is administered daily by intravenous injection for 20 days. Patients often complain about the toxic side effects of the drug, and often need a second or third round of therapy when the drug fails to resolve disease (5). Treatment failure with Sbv in patients with CL can be as high as 45% (6, 7). Amphotericin B, pentamidine or miltefosine are alternative treatment options in patients with no response to Sbv therapy or relapsed.
High rate of treatment failure and the adverse effects of antimony as well as abandon of treatment require the search of new alternative treatment or therapy in combination with the current drug that may shorten the therapy in terms of days and lower toxic adverse effects. Furthermore, in endemic areas of leishmaniasis, only a proportion of individuals progress to disease development upon infection.
To understand the immunopathogenesis of CL caused by L. guyanensis (Lg-CL), this study attempted to find immunological pathways that could be involved in the progression to disease development in susceptible individuals and thus lead to the identification of targets for host-directed immunotherapies.
Materials and methods
Ethics approval and informed consent
This study was approved by the Research Ethics Committee of the Fundação de Medicina Tropical Doutor Heitor Vieira Dourado (FMT-HVD) and granted under the file number CAAE 09995212.0.0000.0005. All the participants or their responsible party for individuals less than 18 years old in the study provided written inform consent. Prior to the signing of the inform consent, explanation about the study was given to the participants and they were free to participate.
Study population
This study was conducted in the peri-rural areas of Manaus, capital of the state of Amazonas, Brazil. The patients with active CL infected with L. guyanensis (Lg-CL) were followed at the FMT-HVD, a referral hospital for treating leishmaniasis patients. The control group comprises healthy controls (HC) living in the same endemic area as the patients, sharing similar environments.
Identification of Leishmania spp
DNA was prepared from lesion biopsy specimens of all the participants with CL. The identification of the Leishmania species was performed by polymerase chain reaction (PCR) restriction fragment length polymorphism and direct nucleotide sequencing as described elsewhere (8). Only patients infected with L. guyanensis were included in the study.
Cytokine assay by Luminex
5 mL of blood from patients with Lg-CL before antimonial treatment and from healthy controls were collected. Plasma was separated and kept frozen at -80°C until plasma cytokines assay.
The levels of FGF basic, Eotaxin, G-CSF, GM-CSF, IFNγ, IL-1β, IL-1RA, IL-2, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-12 (p70), IL-13, IL-15, IL-17A, IP-10, MCP-1, MIP-1α, MIP-1β, PDGF-BB, RANTES, TNFα and VEGF were determined using the multiplex cytokine commercial kit Bio-PlexPro-Human Cytokine GrpI Panel 27-Plex (Bio-Rad) according to the manufacturer’s instructions in the Bio-Plex 200 Protein Array System (Luminex Corporation).
Statistical analysis
Plasma levels of chemokines and cytokines from patients and control groups were compared using Kruskal-Wallis test and the graph plots were constructed using R boxblot package version 3.5.1. The interrelatedness between all biomarker was assessed using Spearman correlation test with R corrplot package version 3.5.1. Values greater or equal to 0.7 were considered positive correlation, whereas values less than or equal to -0,7 were considered negative correlation. A color map matrix and a correlation network diagram (Cytoscape version 3.8.2) were built to represent the correlations between the plasma biomarkers analyzed.
Possible different biomarkers profile between patients and control groups were evaluated through Principal Component Analysis (PCA) clustering. PCA plot were generated based on the absolute abundance of biomarkers using R FactorMineR package version 3.5.1. The regions with 95% confidence levels were indicated with colored ellipses. A Linear Discriminant Analysis (LDA) were performed to determine which biomarker contributes most significantly to the discrimination of the study groups. Kruskal-Wallis (for classes) and pairwise Wilcoxon (for subclasses) tests were used to verify if the data were differentially distributed between the study groups. Logarithmic LDA score for discriminative features was built using a threshold of 2.0. The analysis was done through HutLab galaxy online platform.
Heatmap was generated through hierarchical agglomerative cluster analysis to estimate the pattern of biomarker expression in the study groups. The greatest appropriate distance metric and cluster linkage were defined based on Euclidean distance and complete linkage methods, respectively. Heatmap was created using gplots package in R version 3.5.1.
Results
Characterization of the study population
This study has a cross-sectional design and consisted of 354 patients with Lg-CL (90 males and 264 females) and 376 (107 males and 269 females) healthy individuals with the same socio-epidemiological situations coming from the same endemicity area as the patients. The average age of the male patients with Lg-CL and male healthy controls were 39.8 ± 1.57 and 45.2 ± 1.58 years old, respectively. Age of female patient with Lg-CL and female healthy controls were34.6 ± 0.80 and 43.7 ± 1.80 years old, respectively. The skin lesions of patients were mostly located in the upper and lower limbs. All patients presented recent lesions, ranging three weeks to five weeks. The healthy individuals have no history of leishmaniasis and most of them are agriculture workers as the patients. The study population was devoid of HIV and had no history of diabetes mellitus, cardiac, renal and hepatic disease. The patients had fewer or equal to six lesions and treatment-naïve at the time of enrolment. Most of the patients had only one lesion. All the patients were first time infected. The exclusion criteria were patients with Lg-CL with previous history of leishmaniasis. Pregnant women were excluded.
Leishmania infection triggers intense pro-inflammatory immune response characterized by elevated plasma levels of chemokines, cytokines and growth factors
Plasma circulating levels of 27 biomarkers were assessed in Leishmania-infected patients and healthy individuals from control group. Prior to comparison of the plasma biomarkers between patients with CL and healthy control, biomarkers were investigated if age and sex influenced the plasma levels by linear correlations using linear regressions of the R package (Supplementary Figures 1A–D). The levels of biomarkers were not influenced by sex. However, the levels of IFN-γ, IL-1β, IL12p70, IL-6, IL-17, TNFα, IL-4, IL-5, IL-13, CXCL8, CXCL10, CCL4, CCL5, GM-CSF, and PDGF were influenced by age. The P value of these biomarkers were adjusted for age using the general linear model of R package.
Patients with Lg-CL showed increased levels of IL-1β, IL-6, IL-7, TNF-α, IL-12, IFN-γ, IL-17, IL-1Ra, IL-2, IL-15, IL-4, IL-9 and IL-10 compared to the healthy individuals (Supplementary Figure 2). Likewise, higher levels of FGF-basic, PDGF, VEGF, G-CSF and GM-CSF growth factors were found in the patients group. The plasma levels of MCP-1, IL-13 and IL-5 were similar among the individuals from patient and control groups (p<0.05). These findings demonstrate that the Leishmania infection drives a broader and stronger polyclonal T cell response with the engagement of different leukocyte populations.
Leishmania induced high expression of MIP-1α, eotaxin, MIP1-β, RANTES, IP-10 and IL-8. These chemokines are crucial for the chemoattraction of natural killer cells (MIP1-β), neutrophils (MIP-1α and IL-8), eosinophils (eotaxin), T cells and dendritic cells (RANTES). The IP-10 was the biomarker with the highest level observed in the patients. The leading role of IP-10 is the activation and maintenance of immune response by chemoattraction of monocytes, T cells, NK and dendritic cells; and the promotion of T cell adhesion to endothelial cells.
Several cytokines, chemokines and growth factors are highly correlated in Leishmania-infected patients
The correlation analysis demonstrated a strong positive association among many biomarkers in patients with CL (Figures 1A, B). Total mean correlation (ρ) of 0.41 and 0.31 were observed among patients with CL and control group, respectively.
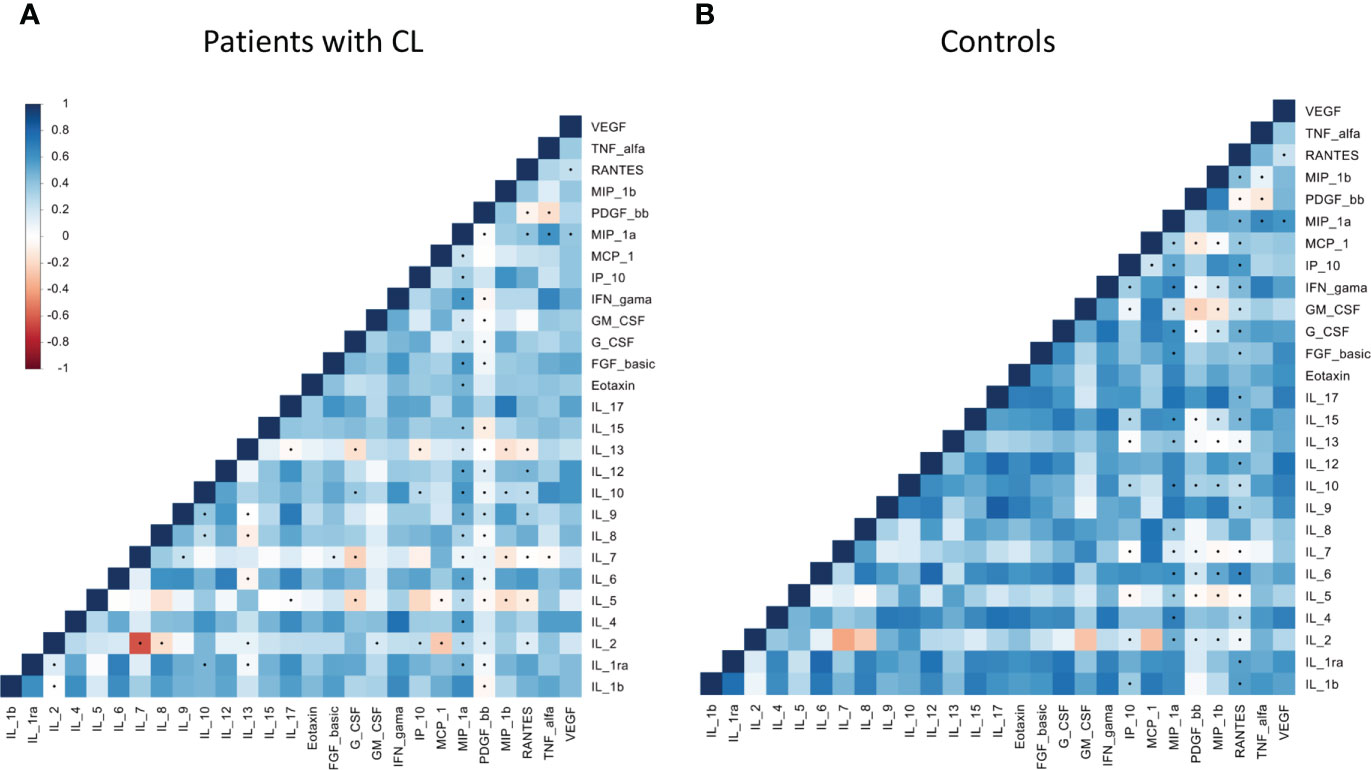
Figure 1 Correlation matrices within chemokines, cytokines and growth factors among patients with cutaneous leishmaniasis (A) and healthy controls (B) groups. Increasing values are represented by colors codes: blue for positive correlation and red for negative correlation. Significance: p< 0.05.
From a total of 340 biomarkers pairs, a mean of 30 pairs showed positive correlation in the patient group (Supplementary Table 1A). The biomarkers pairs that showed the highest correlation mean in the patient group were: Among the patients with Lg-CL, strong positive correlations of IL17/IL12, IL17/IL4, IL17/IL9, IL17/MIP1α, IL17/MIP1β, IL1RA/IL1β, IL6/IL1β, IL15/IL1β, G-CSF/IL1β, IFNγ/IL1β, MCP-1/IL1β, IL6/IL1RA, IL12/IL1RA, G-CSF/IL1RA, IL9/IL4, IFNγ/IL4, Eotaxin/IL4, IL8/IL6, IL12/IL6, G-CSF/IL6, MCP1/IL8, MIP1α/IL9, MIP1β/IL9, FGF-B/IL12, VEGF/IL12, IFNγ/IL15, IFNγ/G-GSF-F, and GM-CSF/MCP1. Only IFNγ/IL4, IL17/MIP1β and IL17/IL9 showed positive correlations in the healthy controls group (Supplementary Table 1B). The cytokine IL-17 and IL-6 presented the greater number of positive pairs correlation (Supplementary Table 1A), which may indicate their key role in the immune response against the Leishmania-infection.
Correlations network based on Spearman correlations of the 27 plasma biomarkers are shown among the patients with Lg-CL in Figure 2A and the healthy controls in Figure 2B. Positive and negative correlations are shown in red and blue, respectively. The thickness of the connections is proportional to the Spearman rank coefficient rho value. IL-17 and IL-1β were the most relevant markers revealing positive relationships among the patients with Lg-CL. IL-2 showed the highest negative correlations among the patients with Lg-CL. Among the healthy controls, strong positive correlations were observed between INF-γ and IL-4 whereas IL-7 showed strong negative relationship with G-CSF.
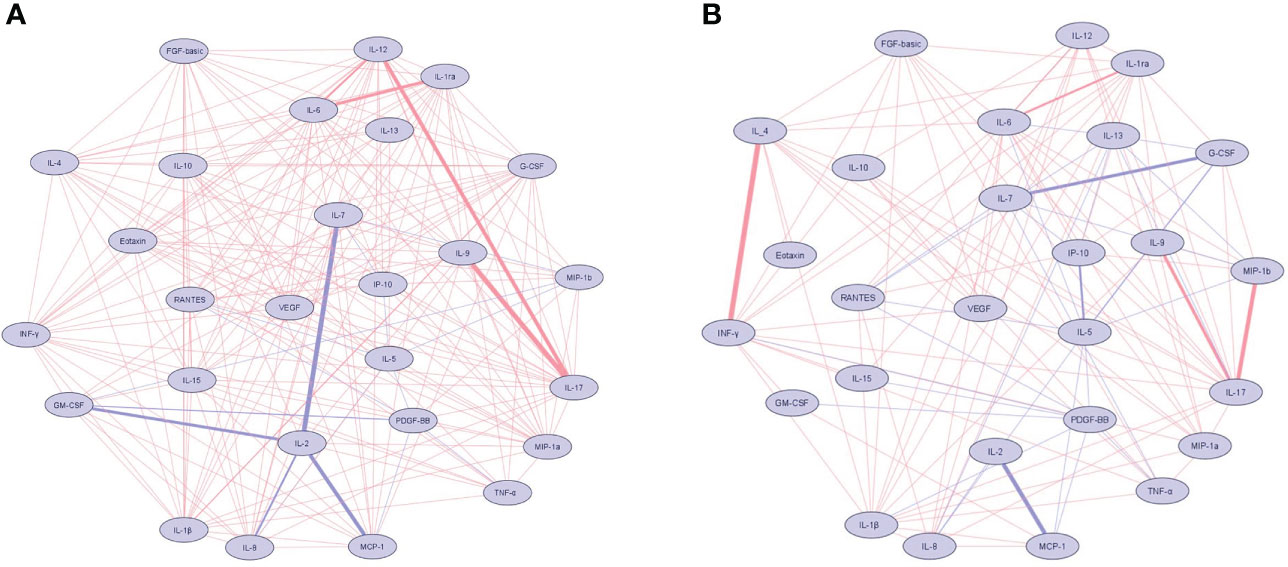
Figure 2 Network of cytokine correlations in the patients with cutaneous leishmaniasis (A) and healthy controls (B) groups. Nodes symbolize cytokines and connecting lines represent a Spearman’s correlation between two biomarkers. Positive correlations are indicated by red lines, while blue lines represent negative correlations. The absolute value of the correlation is represented by the width of the lines. The thickness and saturation of the lines were proportional to the strength of correlation. Biomarkers were arranged based on the number of connections from minimum to maximum. The network diagram was constructed using Cytoscape software version 3.8.2.
High discriminatory activity of plasma biomarkers may predict disease clinical outcome
Correlation pattern of cytokines expressions were assessed in the whole multivariate set of cytokines and chemokines in the study groups. Principal component analysis did not completely segregate the patients with Lg-CL and control groups into separate clusters (Figure 3A). However, patients with Lg-CL were spread over a wide region, which denotes a higher variance of biomarker levels in patients than in controls.
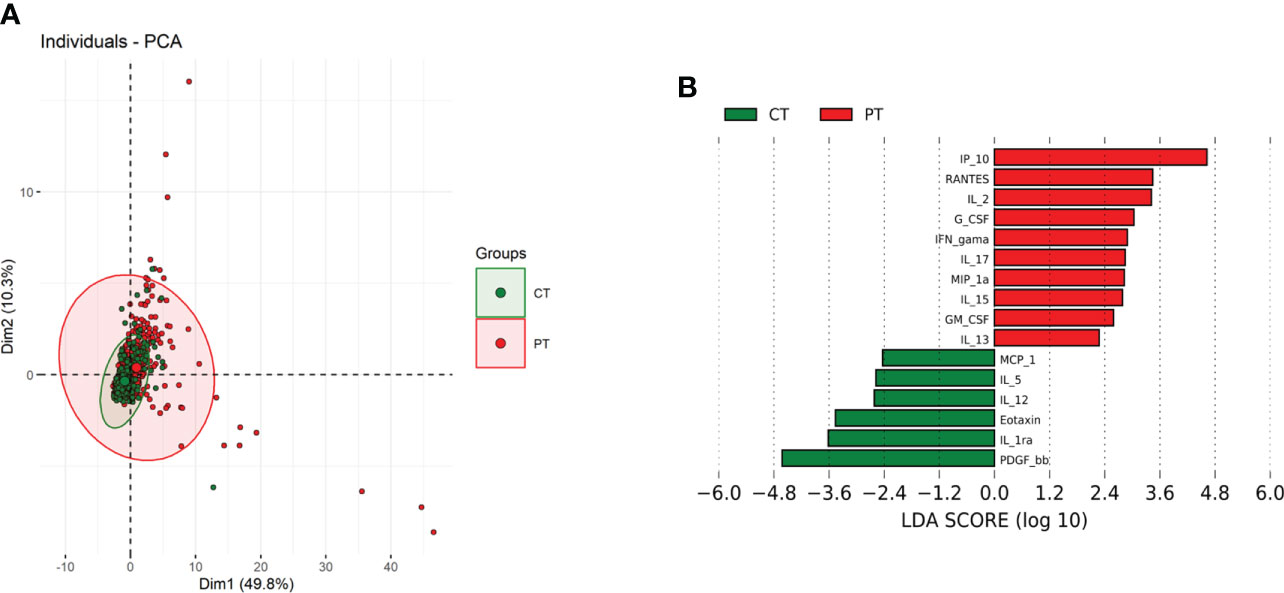
Figure 3 Principal component analysis (PCA) and Linear discriminant analysis (LDA) of reciprocally expressed biomarkers. (A) Scatterplot of the first two principal components (Dim1 and Dim2) showing two main groups, comprising 60.1% of total variance. Biomarkers from patients with cutaneous leishmaniasis (red closed circles) and of healthy control group (green closed circles) clustered in two different groups with overlapping regions. Colored ellipses indicate regions with 95% confidence levels. (B). Separation of biomarkers within two studied groups based on its discrimination power.
Additionally, the linear combinations of biomarkers demonstrated that IP-10, IL-2 and RANTES are the variables with the higher discriminatory activity in the patient group, whereas PDGF, IL-1RA and eotaxin (CCL11) seemed to be more relevant among the control subjects (Figure 3B). This correlation may indicate a functional association between these biomarkers and the immunopathology of leishmaniasis. In other words, IP-10, IL-2 and RANTES could be a predictive value of clinical parameters for progressing to disease, while PDGF, IL-1Ra and eotaxin may be protective biomarkers.
Cluster analysis of biomarkers within the patient and control groups show diffuse expression pattern
We next addressed how the global cytokines and chemokines expression would cluster in a heatmap, without considering the patient clinical manifestations. The sample dendrogram showed two main clusters. However, hierarchical clustering analysis revealed that the patients did not completely segregate from the control group based on their biomarker expression (Figure 4).
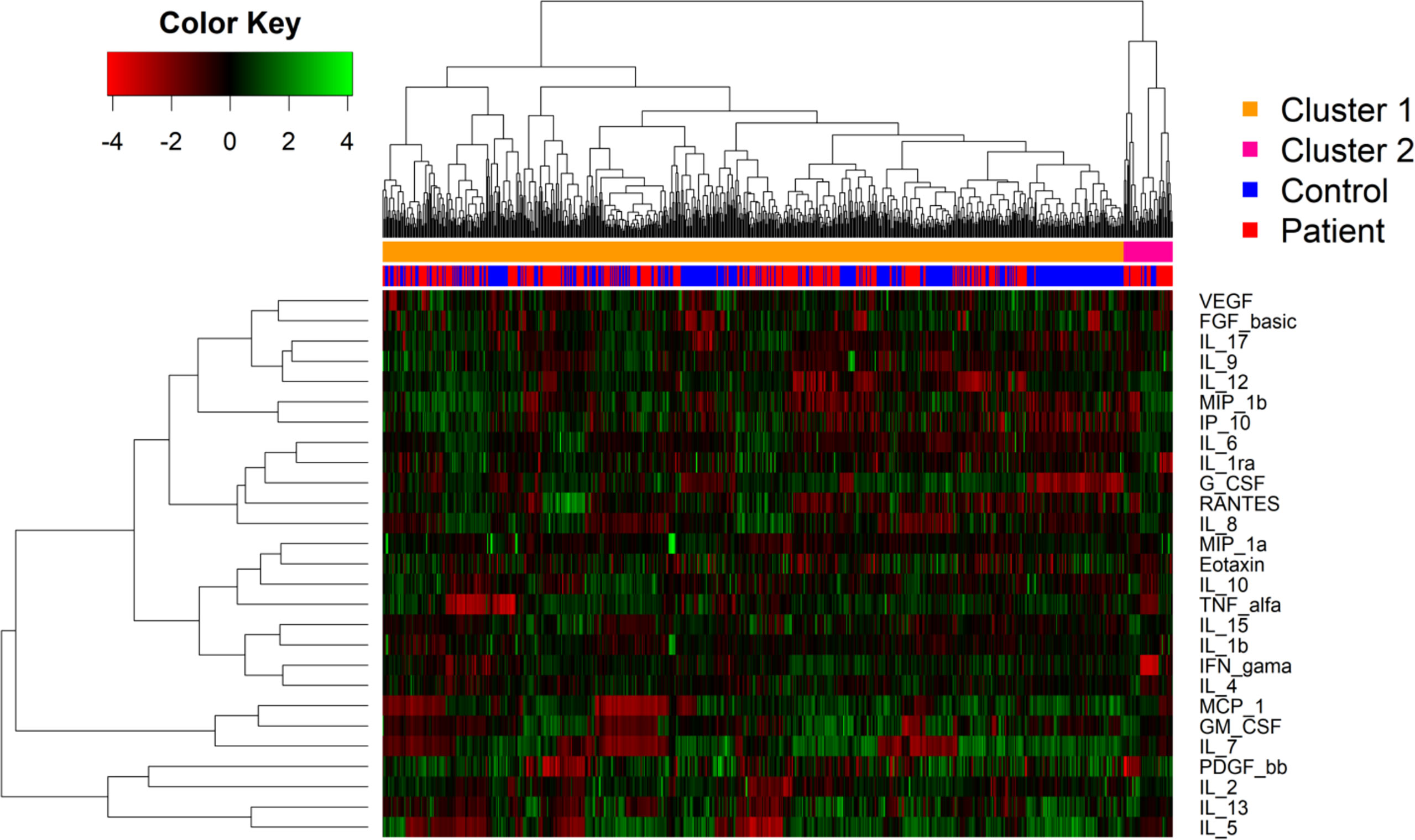
Figure 4 Heat map showing relative expression of biomarkers levels patients with cutaneous leishmaniasis and in healthy controls. Biomarker levels are expressed by color codes. Green represents the lowest and red the highest cytokine levels in the color key on the left. The rows denote the cytokines and the columns the infection status. The cluster analysis demonstrates the level of correlation among the variables represented by dendrograms. The distance between the two clusters is represented by the height where the two clusters merge.
The expression of biomarkers displayed a diffusion pattern. This may have occurred because of underlying unknown inflammatory conditions in some individuals of the control group, which also may increase the levels of the biomarkers analyzed. Moreover, the cluster 2 was predominantly composed by patients with the IFN-γ showing the highest levels. Additionally, as observed in the linear discriminant analysis, the levels of RANTES, IP-10 and IL-2 were higher among patients with Lg-CL compared to the healthy individuals, which confirms the possible role of these cytokines in the immunopathogenesis of leishmaniasis.
Discussion
Currently, there is no vaccine for leishmaniasis and pharmacotherapy with SbV is often ineffective with toxic side effects. Upon infection with the parasite, a TH-1 type immune response is required to control the pathogen and cure the disease. An immune response involves a well-coordinated interplay of antigen-recognizing cells and signaling molecules to mount a sufficient specific response against the pathogen without provoking any tissue damage. Patterns of circulating cytokines and chemokines in patients with Lg-CL may provide useful information to the understanding of the underlying mechanisms of leishmaniasis to enhance the search of novel therapies such as immunotherapy in combination to conventional therapy.
High plasma levels of inflammatory cytokines IL-1β, IL-12, IFN-γ, TNF-α, IL-6 and IL-17 were observed in patients with Lg-CL compared to healthy controls. Hierarchical analysis showed a strong link between IL-1β and IL-6. IL-12 is released from dendritic cells, CD4 and CD8 T lymphocytes. Notably, IL-1β, IL6 and TNF-α are mainly derived from innate cells. Altogether, this may suggest a strong activation of innate cells among the patients with CL culminating in the release of proinflammatory cytokines. Of note, it is well known in animal models that resistance to Leishmania- infection is driven by a TH-1 immune response (9). Exacerbated inflammatory processes and non-specific cytotoxic response participate in the pathogenesis of Lb-CL (10–13). Furthermore, high expression of cytotoxic and inflammatory-related genes was observed in biopsy specimens of skin lesion from patients with CL caused by L. braziliensis (Lb-CL) (14–17). Recently, a study reported high expression of senescence associated secretory phenotype related genes, CCL3, CCL8, CXCL1, CXCL11, CXCL13, CXCL8/IL8 (associated with the recruitment of inflammatory immune cells) and IL-6, IL-15 and IL-1β (inflammatory cytokines) in patients with Lb-CL. IFN-γ, TNF-α, CXL10, and CCL4 also were highly expressed (18). Circulating senescent T cells, with high inflammatory profile, were linked to systemic inflammation and lesion size in patients with Lb-CL (19). Transcriptome analysis from skin lesion of patients with Lb-CL showed strong signature of inflammasome activation and release of IL-1β, IFN-γ and TNF-α (11, 12). This study observed increased plasma levels of CCL3, CXCL8, IL6, IL-15, IL-1β, IFN-γ and TNF-α among the patients with Lg-CL compared to the healthy controls.
High expression levels of IL-10, TGF-β, TNF-α, IFN-γ, IL-12B, CCL2/MCP1, CCL3/MIP1α, CCL5/RANTES and CXCL10/IP10 (Inflammatory profiles) were observed in early lesions of patients with Lb-CL (20). In this study high plasma levels of inflammatory cytokines, IFN-γ, TNF-α, IL-12 and chemokines CCL2/MCP1, CCL3/MIP1α, CCL5/RANTES and CXCL10/IP10 were observed in patients with Lg-CL compared to healthy controls. All of the patients with Lg-CL participating in this study had recent lesions. Control of L.-infection is mediated by T lymphocytes upon early induction of IL-12 that leads to a TH-1 mediated immune response releasing IFN-γ to activate macrophages to keep the parasite in check (21). CXCL10 is cited to protect against L. major infection in mice by stimulating NK cell cytotoxic activity (22). IFN-γ and CXCL10 are reported to activate T lymphocytes and development of a Th1 response during active VL (23). Some studies cited that NK cells are protective against leishmaniasis while others suggested the contrary indicating a cytotoxic role contributing to tissue damage in Lb-infection (24–26). A recent study observed increased frequency of cytotoxic NK cells in cell cultures of PBMC stimulated with Lb-antigens from patients with Lb-CL before treatment, while high frequencies of exhaustion NK cells during treatment (27). Administration of Met-RANTES (an anti-CCL5) in C57/BL6 mice render them susceptible to L. major (28). Immunohistochemistry of biopsy specimens of lesions from patients with Lb-CL or mice showed the presence of T cells infiltration, macrophages, B cells, NK cells and granulocytes (29, 30).
Increased plasma levels of IL-4, IL-9 and IL-10 were observed in patients with CL while levels of IL-5 and IL-13 were similar among both groups. IL-4 and IL-10 are known to modulate TH1-mediated immune response (31). The increased levels of these cytokines in patients with Lg-CL may suggest that the patients react to counter the observed inflammatory process.
Increased levels of FGF-basic, PDGF, VEGF, G-CSF and GM-CSF growth factors were observed among the patients with CL. Growth factors modulate inflammatory process in chronic diseases and may indicate the need of tissue repair. VEGF was suggested to be important in the resolution of leishmaniasis lesions (32). GM-CSF is important during the inflammatory stage of wound healing to induce the migration of neutrophils (33). GM-CSF also induces the migration and proliferation of endothelial cells (34) and upregulate IL-6 (35). Patients with Lb-CL exhibit elevated levels of circulating Th1 lymphocytes, cytokines and chemokines (36). GM-CSF is reported to promote protection against Leishmania-infection (37).
The increased levels of chemokines among the patients with Lg-CL may suggest that the patients are reacting to induce wound healing. Chemokines induced the migration of several proinflammatory cells type to the site of lesions (38), contributing to wound healing by tissue remodeling and angiogenesis (39). IP-10 and MIP-1α attract monocytes, macrophages, and activated T cells to sites of infection to enhance wound healing and parasite elimination (40, 41). However, overexpression of IP-10 may result in delay of wound healing by enhancing a potent inflammatory response through the recruitment of lymphocytes (42). During Leishmania-infection, the chemokines MIP1-β and eotaxin are released by monocytes and can lead to an intense inflammatory response (43).
In this study, patients with Lg-CL displayed a distinct expression profile of proinflammatory cytokines compared to the healthy controls group. IL-1β displayed higher number of positive correlations among the patients with Lg-CL. Several studies have shown that tissue destruction initiated by cytolytic CD8+ T cells trigger NLRP3 inflammasome activation leading to the secretion of IL-1β (10, 11, 14, 44–47). IL-1β is a proinflammatory cytokine involved in the CL pathogenesis (48). Blocking components of NLRP3 or IL-1β decreased the severity of the disease in animal models (11). Higher frequencies of CD8+ T cells have been observed in skin lesions of patients with Lb-CL and are unrelated to parasite load, suggesting CD8+ T cells contribute to the immunopathogenesis of CL (46, 47). CD8+ T cells cocultured with Lb-infected macrophages enhanced the expression of NLRP3, AIM2, and CASP1/5 culminating to the release of IL-1β. High expression of NLRP3, AIM2, and CASP1/5 have also been observed in skin lesions of patients with Lb-CL (49).
The immune response in infectious diseases to control the pathogen depends on the type and magnitude of the response. An excess of proinflammatory cytokines may lead to tissue destruction. RNA-seq of skin biopsies from patients with Lb-CL highlights an increase of inflammatory transcript of IFNG and TNFA (16). A recent study showed that a transcriptional signature including IL-1β may predict clinical outcomes of Lb-infection (15). Notably, IL-1β and IFN-γ were suggested to predict clinical outcome in patients with Lb-CL (15). Interestingly, variants in the IL1B and IFNG genes are associated with susceptibility to L.-infection (8, 50–52).
IL-17 showed positive correlations with IL-12, IL-9, MIP1α, MIP-1β and IL-4. Th17 cells and the IL-17/IL-23 axis have crucial role in immune-mediated inflammatory diseases (53). IL-17 induces fibroblasts, macrophages, endothelial and epithelial cells to release TNF-α, IL-6, NOS and metalloproteases to increase inflammation (54, 55). Plasma IL-6, IL-1β, IFNγ, TNFα and IL-17 were higher in patients with CL. Indeed, IL6 and IL-1β were positively correlated in CL patients in this study. Interestingly, TGF-β together with IL-6 or IL-1β downregulate FOXP3 and trigger the activation of RORgt, the transcription factor of IL17, and TH17 cells differentiation (56). TNFα and IL-1β induce the upregulation of IL-6 expression (57). High levels of IL-17 were correlated with the magnitude of cellular-infiltrates in Lb-CL (58). In biopsy specimens of skin lesion of patients with CL, IL-17, RORgt and IL-23 were observed (59–62). In light of all these studies and our findings, IL-17 seems to contribute to the pathogenesis and inflammatory processes of leishmaniasis.
A recent study observed that patients with Lb-CL who failed treatment after 60 days with Sbv displayed lower plasma levels of eotaxin and IL-12p70 but increased G-CSF compared to successfully treated patients (63). Interestingly, PDGF, Eotaxin and IL1-RA showed the highest discriminatory activity among the healthy controls compared to the patients with Lg-CL in this study.
Blood transcriptional analysis of patients with active VL caused by L. infantum (chagasi), patients under remission, asymptomatic and uninfected individuals revealed molecular pathways of activation of T lymphocytes via MHC class I and type I interferon signaling besides downregulation of pathways related to myeloid cells (monocytes and neutrophils) in patients with active VL, while patients under remission showed genes correlated with activation of Notch signaling pathway and increased proportions of B cells besides T lymphocytes via MHC class I and type I interferon signaling activation as patients with active VL (64). Patients under remission also displayed a negative regulation of IL-10 signaling pathway in the transcriptional profiles. Biopsy specimens of lesions from patients with Lb-CL also displayed a transcriptional profiling of positive type I interferon signaling (14). Human myeloid derived dendritic cells infected with L. major induces a type I transcriptional signature leading to the production of IL-12 (65). All these studies point to a common response to Leishmania -infection irrespective of the species. Accordingly, our study is unable to confirm the type I Interferon signaling in Lg-CL as our study is not ample enough to answer this finding. However, high RNA expression of CXCL10 and CXCL8 as well as high plasma levels of IFN- γ, CXCL10, CXCL8 and IL-10 were observed in PBMCs and plasma from VL patients caused by L. donovani, respectively (66). Another study also observed high levels of plasma IFN- γ, TNF-α, IL-6, CXCL8, and IL-10 in active VL caused by L. infantum (chagasi) (67). It is noteworthy to highlight that we also observed high plasma levels of IFN- γ, TNF-α, IL-6 and CXCL10, CXCL8 and IL-10 among the patients with Lg-CL. Spearman correlation of IFN- γ to IL-10 among the healthy controls is similar to patients with Lg-CL (ρ = 0.62). It is well-known that IL-8 attracts neutrophils at the site of infection and IFN- γ triggers macrophage activation to keep the parasites in check. Correlation of IFN- γ to IL-4 among healthy controls was also similar among HC and patients with Lg-CL (ρ = 0.7).
This study has few limitations. Comparisons of circulating biomarkers were performed between patients with Lg-CL and healthy individuals sharing the same endemic area. The study is a transversal investigation and analyzed the patients only before treatment. It would be interesting to identify predictive markers of treatment failure or success by comparing the immune profile in infected individuals before and after completion of treatment.
Conclusion
Altogether, this study identifies plasma cytokines pathways that may lead to the development of disease in Lg-infected susceptible individuals. While common response is observed to Leishmania-infection, however, our study points that IP-10, IL-2 and RANTES could be a predictive value of clinical parameters for progressing to disease, while PDGF, IL-1Ra and eotaxin may be protective biomarkers. IL-17 and IL-1β also are potential predictive markers for progression to disease in Lg-infected individuals. Identifying potential target in these pathways may open the way for immunotherapy.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Ethics statement
The studies involving human participants were reviewed and approved by Research Ethics Committee of the Fundação de Medicina Tropical Doutor Heitor Vieira Dourado (FMT-HVD). Written informed consent to participate in this study was provided by the participants’ legal guardian/next of kin.
Author contributions
TM, MS, and RR contributed for data curation. TM, JJ, MG, and RR take responsibility for the integrity of the work as a whole, from inception to published article. TM, JJ, and RR were responsible for study design and conception and drafted the manuscript. TM, JJ, LOS, GVS, FA, SP, HK, LSS, LMS, SA, KQ, JS, CS, and HS recruited healthy controls in endemic areas. TM, JJ, MG, and RR collected and cleaned the data for formal analysis. GSP and AB were responsible for statistical analysis. TM and RR interpreted the results and drafted the manuscript. All authors revised the manuscript for important intellectual content.
Funding
This research was funded by the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), grant number 404181/2012-0 to RR, Fundação de Amparo à Pesquisa do Estado do Amazonas (FAPEAM), grant number 06201954/2015 to RR and FAPEAM RESOLUÇÃO N. 002/2008, 007/2018 E 005/2019 – PRÓ-ESTADO. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Acknowledgments
We thank all the patients and healthy individuals from the endemic area who have been willing to participate.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2022.974051/full#supplementary-material
Supplementary Figure 1 | Plasma biomarkers assessed in the total population in relation to age and sex.
Supplementary Figure 2 | Plasma cytokines, chemokines profile and growth factors in patients with cutaneous leishmaniasis and healthy controls. Plasma samples from Leishmania guyanensis-infected patients (PT) and healthy individuals(CT) were assessed by Luminex. The results are expressed in pg/mL and displayed as median and interquartile range. Comparative analysis between control vs patient was done by Kruskall-Wallis test and significant differences are expressed as p < 0.05, p < 0.01, p < 0.001 and p < 0.0001). For biomarkers influenced by age, P values were adjusted for age by general linear model of the R package.
References
1. World Health Organization. Leishmaniasis (2022). Available at: https://www.who.int/news-room/fact-sheets/detail/leishmaniasis (Accessed June 11, 2022).
2. Scorza BM, Carvalho EM, Wilson ME. Cutaneous manifestations of human and murine leishmaniasis. Int J Mol Sci (2017) 18(6):1296. doi: 10.3390/ijms18061296
3. Benício E, Cordeiro M, Monteiro H, Saboia Moura MA, Oliveira C, Nunes Gadelha EP, et al. Sustained presence of cutaneous leishmaniasis in urban manaus, the largest human settlement in the Amazon. Am J Trop Med Hygiene (2015) 93(6):1208–13. doi: 10.4269/ajtmh.14-0164
4. Uliana S, Trinconi CT, Coelho AC. Chemotherapy of leishmaniasis: present challenges. Parasitology (2018) 145(4):464–80. doi: 10.1017/S0031182016002523
5. Gadelha E, Ramasawmy R, da Costa Oliveira B, Morais Rocha N, de Oliveira Guerra JA, Allan Villa Rouco da Silva G, et al. An open label randomized clinical trial comparing the safety and effectiveness of one, two or three weekly pentamidine isethionate doses (seven milligrams per kilogram) in the treatment of cutaneous leishmaniasis in the Amazon region. PloS Negl Trop Dis (2018) 12(10):e0006850. doi: 10.1371/journal.pntd.0006850
6. Machado PR, Ampuero J, Guimarães LH, Villasboas L, Rocha AT, Schriefer A, et al. Miltefosine in the treatment of cutaneous leishmaniasis caused by Leishmania braziliensis in Brazil: a randomized and controlled trial. PloS Negl Trop Dis (2010) 4(12):e912. doi: 10.1371/journal.pntd.0000912
7. Prates FV, Dourado ME, Silva SC, Schriefer A, Guimarães LH, Brito MD, et al. Fluconazole in the treatment of cutaneous leishmaniasis caused by Leishmania braziliensis: A randomized controlled trial. Clin Infect Dis: an Off Publ Infect Dis Soc America (2017) 64(1):67–71. doi: 10.1093/cid/ciw662
8. da Silva GAV, de Mesquita TGR, de Souza Encarnação HV, do Espírito Santo JJ, da Costa Sabino K, de Aguiar Neres I, et al. A polymorphism in the IL1B gene (rs16944 T/C) is associated with cutaneous leishmaniasis caused by Leishmania guyanensis and plasma cytokine interleukin receptor antagonist. Cytokine (2019) 123:154788. doi: 10.1016/j.cyto.2019.154788
9. Awasthi A, Mathur RK, Saha B. Immune response to leishmania infection. Indian J Med Res (2004) 119(6):238–58.
10. Faria DR, Souza PE, Durães FV, Carvalho EM, Gollob KJ, Machado PR, et al. Recruitment of CD8(+) T cells expressing granzyme a is associated with lesion progression in human cutaneous leishmaniasis. Parasite Immunol (2009) 31(8):432–9. doi: 10.1111/j.1365-3024.2009.01125.x
11. Novais FO, Carvalho AM, Clark ML, Carvalho LP, Beiting DP, Brodsky IE, et al. CD8+ T cell cytotoxicity mediates pathology in the skin by inflammasome activation and IL-1β production. PloS Pathog (2017) 13(2):e1006196. doi: 10.1371/journal.ppat.1006196
12. Carvalho AM, Novais FO, Paixão CS, de Oliveira CI, Machado P, Carvalho LP, et al. Glyburide, a NLRP3 inhibitor, decreases inflammatory response and is a candidate to reduce pathology in Leishmania braziliensis infection. J Invest Dermatol (2020) 140(1):246–249.e2. doi: 10.1016/j.jid.2019.05.025
13. Santos D, Campos TM, Saldanha M, Oliveira SC, Nascimento M, Zamboni DS, et al. IL-1β production by intermediate monocytes is associated with immunopathology in cutaneous leishmaniasis. J Invest Dermatol (2018) 138(5):1107–15. doi: 10.1016/j.jid.2017.11.029
14. Novais FO, Carvalho LP, Passos S, Roos DS, Carvalho EM, Scott P, et al. Genomic profiling of human Leishmania braziliensis lesions identifies transcriptional modules associated with cutaneous immunopathology. J Invest Dermatol (2015) 135(1):94–101. doi: 10.1038/jid.2014.305
15. Amorim CF, Novais FO, Nguyen BT, Misic AM, Carvalho LP, Carvalho EM, et al. Variable gene expression and parasite load predict treatment outcome in cutaneous leishmaniasis. Sci Trans Med (2019) 11(519):eaax4204. doi: 10.1126/scitranslmed.aax4204
16. Christensen SM, Dillon L, Carvalho LP, Passos S, Novais FO, Hughitt VK, et al. Correction: Meta-transcriptome profiling of the human-Leishmania braziliensis cutaneous lesion. PloS Negl Trop Dis (2017) 11(5):e0005588. doi: 10.1371/journal.pntd.0005588
17. Campos TM, Novais FO, Saldanha M, Costa R, Lordelo M, Celestino D, et al. Granzyme b produced by natural killer cells enhances inflammatory response and contributes to the immunopathology of cutaneous leishmaniasis. J Infect Dis (2020) 221(6):973–82. doi: 10.1093/infdis/jiz538
18. Fantecelle CH, Covre LP, Garcia de Moura R, Guedes H, Amorim CF, Scott P, et al. Transcriptomic landscape of skin lesions in cutaneous leishmaniasis reveals a strong CD8+ T cell immunosenescence signature linked to immunopathology. Immunology (2021) 164(4):754–65. doi: 10.1111/imm.13410
19. Covre LP, Martins RF, Devine OP, Chambers ES, Vukmanovic-Stejic M, Silva JA, et al. Circulating senescent T cells are linked to systemic inflammation and lesion size during human cutaneous leishmaniasis. Front Immunol (2019) 9:3001. doi: 10.3389/fimmu.2018.03001
20. Costa-Silva MF, Gomes LI, Martins-Filho OA, Rodrigues-Silva R, Freire J, Quaresma PF, et al. Gene expression profile of cytokines and chemokines in skin lesions from Brazilian indians with localized cutaneous leishmaniasis. Mol Immunol (2014) 57(2):74–85. doi: 10.1016/j.molimm.2013.08.008
21. Cummings HE, Tuladhar R, Satoskar AR. Cytokines and their STATs in cutaneous and visceral leishmaniasis. J Biomed Biotechnol (2010) 2010:294389. doi: 10.1155/2010/294389
22. Vester B, Müller K, Solbach W, Laskay T. Early gene expression of NK cell-activating chemokines in mice resistant to Leishmania major. Infect Immun (1999) 67(6):3155–9. doi: 10.1128/IAI.67.6.3155-3159.1999
23. Kumar R, Singh N, Gautam S, Singh OP, Gidwani K, Rai M, et al. Leishmania specific CD4 T cells release IFNγ that limits parasite replication in patients with visceral leishmaniasis. PloS Negl Trop Dis (2014) 8(10):e3198. doi: 10.1371/journal.pntd.0003198
24. Maasho K, Sanchez F, Schurr E, Hailu A, Akuffo H. Indications of the protective role of natural killer cells in human cutaneous leishmaniasis in an area of endemicity. Infect Immun (1998) 66(6):2698–704. doi: 10.1128/IAI.66.6.2698-2704.1998
25. Pereira LI, Dorta ML, Pereira AJ, Bastos RP, Oliveira MA, Pinto SA, et al. Increase of NK cells and proinflammatory monocytes are associated with the clinical improvement of diffuse cutaneous leishmaniasis after immunochemotherapy with BCG/Leishmania antigens. Am J Trop Med Hygiene (2009) 81(3):378–83. doi: 10.4269/ajtmh.2009.81.378
26. Bogdan C. Natural killer cells in experimental and human leishmaniasis. Front Cell Infect Microbiol (2012) 2:69. doi: 10.3389/fcimb.2012.00069
27. Cunha CF, Ferraz-Nogueira R, Costa VFA, Pimentel MIF, Chometon TQ, Lyra MR, et al. Contribution of Leishmania braziliensis antigen-specific CD4+ T, CD8+ T, NK and CD3+CD56+NKT cells in the immunopathogenesis of cutaneous leishmaniasis patients: Cytotoxic, activation and exhaustion profiles. PloS One (2020) 15(3):e0229400. doi: 10.1371/journal.pone.0229400
28. Santiago HC, Oliveira CF, Santiago L, Ferraz FO, de Souza D, de-Freitas LA, et al. Involvement of the chemokine RANTES (CCL5) in resistance to experimental infection with Leishmania major. Infect Immun (2004) 72(8):4918–23. doi: 10.1128/IAI.72.8.4918-4923.2004
29. Esterre P, Dedet JP, Frenay C, Chevallier M, Grimaud JA. Cell populations in the lesion of human cutaneous leishmaniasis: a light microscopical, immunohistochemical and ultrastructural study. Virchows Archiv A Pathological Anat Histopathol (1992) 421(3):239–47. doi: 10.1007/BF01611181
30. Novais FO, Santiago RC, Báfica A, Khouri R, Afonso L, Borges VM, et al. Neutrophils and macrophages cooperate in host resistance against Leishmania braziliensis infection. J Immunol (Baltimore Md. 1950) (2009) 183(12):8088–98. doi: 10.4049/jimmunol.0803720
31. Spellberg B, Edwards J. E. C.OMMAJ.R.X.X.X. Type 1/Type 2 immunity in infectious diseases. Clin Infect Dis: an Off Publ Infect Dis Soc America (2001) 32(1):76–102. doi: 10.1086/317537
32. Weinkopff T, Konradt C, Christian DA, Discher DE, Hunter CA, Scott P. Leishmania major infection-induced VEGF-A/VEGFR-2 signaling promotes lymphangiogenesis that controls disease. J Immunol (Baltimore Md.: 1950) (2016) 197(5):1823–31. doi: 10.4049/jimmunol.1600717
33. Yönem A, Cakir B, Güler S, Azal O O, Corakçi A. Effects of granulocyte-colony stimulating factor in the treatment of diabetic foot infection. Diabetes Obes Metab (2001) 3(5):332–7. doi: 10.1046/j.1463-1326.2001.00142.x
34. Bussolino F, Wang JM, Defilippi P, Turrini F, Sanavio F, Edgell CJ, et al. Granulocyte- and granulocyte-macrophage-colony stimulating factors induce human endothelial cells to migrate and proliferate. Nature (1989) 337(6206):471–3. doi: 10.1038/337471a0
35. Mann A, Breuhahn K, Schirmacher P, Blessing M. Keratinocyte-derived granulocyte-macrophage colony stimulating factor accelerates wound healing: Stimulation of keratinocyte proliferation, granulation tissue formation, and vascularization. J Invest Dermatol (2001) 117(6):1382–90. doi: 10.1046/j.0022-202x.2001.01600.x
36. França-Costa J, Van Weyenbergh J, Boaventura VS, Luz NF, Malta-Santos H, Oliveira MC, et al. Arginase I, polyamine, and prostaglandin E2 pathways suppress the inflammatory response and contribute to diffuse cutaneous leishmaniasis. J Infect Dis (2015) 211(3):426–35. doi: 10.1093/infdis/jiu455
37. Carvalho GB, Costa LE, Lage DP, Ramos FF, Santos T, Ribeiro P, et al. High-through identification of T cell-specific phage-exposed mimotopes using PBMCs from tegumentary leishmaniasis patients and their use as vaccine candidates against Leishmania amazonensis infection. Parasitology (2019) 146(3):322–32. doi: 10.1017/S0031182018001403
38. Barrientos S, Stojadinovic O, Golinko MS, Brem H, Tomic-Canic M. Growth factors and cytokines in wound healing. Wound Repair Regeneration: Off Publ Wound Healing Soc [and] Eur Tissue Repair Soc (2008) 16(5):585–601. doi: 10.1111/j.1524-475X.2008.00410.x
39. Raja, Sivamani K, Garcia MS, Isseroff RR. Wound re-epithelialization: modulating keratinocyte migration in wound healing. Front Biosci: J Virtual Library (2007) 12:2849–68. doi: 10.2741/2277
40. Dufour JH, Dziejman M, Liu MT, Leung JH, Lane TE, Luster AD. IFN-gamma-inducible protein 10 (IP-10; CXCL10)-deficient mice reveal a role for IP-10 in effector T cell generation and trafficking. J Immunol (2002) 168(7):3195–204. doi: 10.4049/jimmunol.168.7.3195
41. Ritter U, Körner H. Divergent expression of inflammatory dermal chemokines in cutaneous leishmaniasis. Parasite Immunol (2002) 24(6):295–301. doi: 10.1046/j.1365-3024.2002.00467.x
42. Christopherson K 2nd, Hromas R. Chemokine regulation of normal and pathologic immune responses. Stem Cells (Dayton Ohio) (2001) 19(5):388–96. doi: 10.1634/stemcells.19-5-388
43. Matte C, Olivier M. Leishmania-induced cellular recruitment during the early inflammatory response: modulation of proinflammatory mediators. J Infect Dis (2002) 185(5):673–81. doi: 10.1086/339260
44. Novais FO, Carvalho LP, Graff JW, Beiting DP, Ruthel G, Roos DS, et al. Cytotoxic T cells mediate pathology and metastasis in cutaneous leishmaniasis. PloS Pathog (2013) 9(7):e1003504. doi: 10.1371/journal.ppat.1003504
45. Cardoso TM, Machado Á., Costa DL, Carvalho LP, Queiroz A, Machado P, et al. Protective and pathological functions of CD8+ T cells in Leishmania braziliensis infection. Infect Immun (2015) 83(3):898–906. doi: 10.1128/IAI.02404-14
46. Santos C, Boaventura V, Ribeiro Cardoso C, Tavares N, Lordelo MJ, Noronha A, et al. CD8(+) granzyme b(+)-mediated tissue injury vs. CD4(+)IFNγ(+)-mediated parasite killing in human cutaneous leishmaniasis. J Invest Dermatol (2013) 133(6):1533–40. doi: 10.1038/jid.2013.4
47. Crosby EJ, Clark M, Novais FO, Wherry EJ, Scott P. Lymphocytic choriomeningitis virus expands a population of NKG2D+CD8+ T cells that exacerbates disease in mice coinfected with Leishmania major. J Immunol (2015) 195(7):3301–10. doi: 10.4049/jimmunol.1500855
48. Zamboni DS, Sacks DL. Inflammasomes and leishmania: in good times or bad, in sickness or in health. Curr Opin Microbiol (2019) 52:70–6. doi: 10.1016/j.mib.2019.05.005
49. Cardoso TM, Lima JB, Bonyek-Silva Í., Nunes S, Feijó D, Almeida H, et al. Inflammasome activation by CD8+ T cells from patients with cutaneous leishmaniasis caused by Leishmania braziliensis in the immunopathogenesis of the disease. J Invest Dermatol (2021) 141(1):209–213.e2. doi: 10.1016/j.jid.2020.05.106
50. da Silva GAV, Mesquita TG, Souza VC, Junior JDES, Gomes de Souza ML, Talhari AC, et al. A single haplotype of IFNG correlating with low circulating levels of interferon-γ is associated with susceptibility to cutaneous leishmaniasis caused by Leishmania guyanensis. Clin Infect Dis (2020) 71(2):274–81. doi: 10.1093/cid/ciz810
51. Fernández-Figueroa EA, Rangel-Escareño C, Espinosa-Mateos V, Carrillo-Sánchez K, Salaiza-Suazo N, Carrada-Figueroa G, et al. Disease severity in patients infected with Leishmania mexicana relates to IL-1β. PloS Negl Trop Dis (2012) 6(5):e1533. doi: 10.1371/journal.pntd.0001533
52. Moravej A, Rasouli M, Kalani M, Asaei S, Kiany S, Najafipour S, et al. IL-1β (-511T/C) gene polymorphism not IL-1β (+3953T/C) and LT-α (+252A/G) gene variants confers susceptibility to visceral leishmaniasis. Mol Biol Rep (2012) 39(6):6907–14. doi: 10.1007/s11033-012-1517-z
53. Bunte K, Beikler T. Th17 cells and the IL-23/IL-17 axis in the pathogenesis of periodontitis and immune-mediated inflammatory diseases. Int J Mol Sci (2019) 20(14):3394. doi: 10.3390/ijms20143394
54. Kolls JK, Lindén A. Interleukin-17 family members and inflammation. Immunity (2004) 21(4):467–76. doi: 10.1016/j.immuni.2004.08.018
55. Nakae S, Saijo S, Horai R, Sudo K, Mori S, Iwakura Y. IL-17 production from activated T cells is required for the spontaneous development of destructive arthritis in mice deficient in IL-1 receptor antagonist. Proc Natl Acad Sci United States America (2003) 100(10):5986–90. doi: 10.1073/pnas.1035999100
56. Wang M, Tian T, Yu S, He N, Ma D. Th17 and treg cells in bone related diseases. Clin Dev Immunol (2013) 2013:203705. doi: 10.1155/2013/203705
57. Ruddy MJ, Wong GC, Liu XK, Yamamoto H, Kasayama S, Kirkwood KL, et al. Functional cooperation between interleukin-17 and tumor necrosis factor-alpha is mediated by CCAAT/enhancer-binding protein family members. J Biol Chem (2004) 279(4):2559–67. doi: 10.1074/jbc.M308809200
58. Bacellar O, Faria D, Nascimento M, Cardoso TM, Gollob KJ, Dutra WO, et al. Interleukin 17 production among patients with American cutaneous leishmaniasis. J Infect Dis (2009) 200(1):75–8. doi: 10.1086/599380
59. Gonzalez-Lombana C, Gimblet C, Bacellar O, Oliveira WW, Passos S, Carvalho LP, et al. IL-17 mediates immunopathology in the absence of IL-10 following leishmania major infection. PloS Pathog (2013) 9(3):e1003243. doi: 10.1371/journal.ppat.1003243
60. Araujo Flores GV, Sandoval Pacheco CM, Sosa Ochoa WH, Gomes C, Zúniga C, Corbett CP, et al. Th17 lymphocytes in atypical cutaneous leishmaniasis caused by Leishmania (L.) infantum chagasi in central America. Parasite Immunol (2020) 42(11):e12772. doi: 10.1111/pim.12772
61. Gonzalez K, Calzada JE, Corbett C, Saldaña A, Laurenti MD. Involvement of the inflammasome and Th17 cells in skin lesions of human cutaneous leishmaniasis caused by leishmania (Viannia) panamensis. Mediators Inflammation (2020) 2020:9278931. doi: 10.1155/2020/9278931
62. Katara GK, Raj A, Kumar R, Avishek K, Kaushal H, Ansari NA, et al. Analysis of localized immune responses reveals presence of Th17 and treg cells in cutaneous leishmaniasis due to Leishmania tropica. BMC Immunol (2013) 14:52. doi: 10.1186/1471-2172-14-52
63. Malta-Santos H, Fukutani KF, Sorgi CA, Queiroz A, Nardini V, Silva J, et al. Multi-omic analyses of plasma cytokines, lipidomics, and transcriptomics distinguish treatment outcomes in cutaneous leishmaniasis. iScience (2020) 23(12):101840. doi: 10.1016/j.isci.2020.101840
64. Gardinassi LG, Garcia GR, Costa CH, Costa Silva V, de Miranda Santos IK. Blood transcriptional profiling reveals immunological signatures of distinct states of infection of humans with leishmania infantum. PloS Negl Trop Dis (2016) 10(11):e0005123. doi: 10.1371/journal.pntd.000512365
65. Favila MA, Geraci NS, Zeng E, Harker B, Condon D, Cotton RN, et al. Human dendritic cells exhibit a pronounced type I IFN signature following leishmania major infection that is required for IL-12 induction. J Immunol (Baltimore Md. 1950) (2014) 192(12):5863–72. doi: 10.4049/jimmunol.1203230
66. Singh N, Sundar S. Inflammatory chemokines and their receptors in human visceral leishmaniasis: Gene expression profile in peripheral blood, splenic cellular sources and their impact on trafficking of inflammatory cells. Mol Immunol (2017) 85:111–9. doi: 10.1016/j.molimm.2017.02.008
67. Peruhype-Magalhães V, Martins-Filho OA, Prata A, Silva L, Rabello A, Teixeira-Carvalho A, et al. Mixed inflammatory/regulatory cytokine profile marked by simultaneous raise of interferon-gamma and interleukin-10 and low frequency of tumour necrosis factor-alpha(+) monocytes are hallmarks of active human visceral leishmaniasis due to Leishmania chagasi infection. Clin Exp Immunol (2006) 146(1):124–32. doi: 10.1111/j.1365-2249.2006.03171.x
Keywords: Leishmania guyanensis, cytokines, chemokines, cutaneous leishmaniasis, growth factors
Citation: Mesquita TGRd, Junior JdES, Silva LDOd, Silva GAV, Araújo FJd, Pinheiro SK, Kerr HKA, Silva LSd, Souza LMd, Almeida SAd, Queiroz KLGD, Souza JLd, Silva CCd, Sequera HDG, Souza MLGd, Barbosa AN, Pontes GS, Guerra MVdF and Ramasawmy R (2022) Distinct plasma chemokines and cytokines signatures in Leishmania guyanensis-infected patients with cutaneous leishmaniasis. Front. Immunol. 13:974051. doi: 10.3389/fimmu.2022.974051
Received: 28 June 2022; Accepted: 08 August 2022;
Published: 25 August 2022.
Edited by:
Mohd Adnan, University of Hail, Saudi ArabiaReviewed by:
Tatjana Keesen, Federal University of Paraíba, BrazilAhmad Khosravi, Kerman University of Medical Sciences, Iran
Copyright © 2022 Mesquita, Junior, Silva, Silva, Araújo, Pinheiro, Kerr, Silva, Souza, Almeida, Queiroz, Souza, Silva, Sequera, Souza, Barbosa, Pontes, Guerra and Ramasawmy. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rajendranath Ramasawmy, ramasawm@gmail.com