摘要: 探究食物网的结构和复杂性,可深入了解其功能、营养动力学以及能量转化过程。本文水晶河食物网数据由北卡罗来纳州大学威明顿分校的Stuart Borrett教授整理并提供,基于11个拓扑网络指数对以上数据进行分析,通过建立水晶河的拓扑网络,研究了该地区食物网的结构和复杂性。在分析水晶河食物网时,我们得出了以下结果:该食物网的物种数为19,连接数为65,每个物种的相互作用数量为3.42,连接性为0.18。顶级物种、中间物种和基础物种的比例分别为47.4%、42.1%和10.5%。食物网的杂食性指数为36.8%,连接复杂性指数为7.22,特征路径长度为2.111,聚类系数为0.325。根据对物种间相互作用数量和连接性的研究发现,水晶河食物网的L/S的数值处于非正常范围内、L/S2的数值均处于正常范围内,由此判定水晶河食物网的复杂性处于非正常的状态。杂食性物种数量比例、特征路径长度和聚集系数等指标均表明水晶河食物网的不稳定性。对水晶河食物网结构和复杂性的研究有助于深化对该生态系统的认知,也为深入探究水晶河食物网功能和科学管理渔业资源提供理论支持。
Abstract:
Exploring the structure and complexity of food webs provides an in-depth understanding of their functionality, nutritional dynamics, and energy transformation processes. In this study, the Crystal River food web data, compiled and provided by Professor Stuart Borrett from the University of North Carolina Wilmington, were analyzed using 11 topological network indices. By establishing the topological network of Crystal River, the structure and complexity of the food web in this region were studied. When analyzing the Crystal River food web, the following results were obtained: The number of species in the food web was 19, with 65 connections, an average interaction per species of 3.42, and a connectance of 0.18. The proportions of top species, intermediate species, and basal species were 47.4%, 42.1%, and 10.5%, respectively. The omnivory index of the food web was 36.8%, the connectance complexity index was 7.22, the characteristic path length was 2.111, and the clustering coefficient was 0.325. Based on the analysis of the number of interactions and con-nectance between species, it was determined that the value of L/S in the Crystal River food web was in the abnormal range, and the value of L/S2 was in the normal range, indicating the complexity of the Crystal River food web was in the abnormal state. Indicators such as the proportion of omniv-orous species, characteristic path length, and clustering coefficient all suggest the instability of the Crystal River food web. The study of the structure and complexity of the Crystal River food web con-tributes to a deeper understanding of this ecosystem and provides theoretical support for further investigating its functionality and scientifically managing fisheries resources.
1. 引言
近年来,全球生物多样性正面临着由人类活动 [1] 、气候变异 [2] [3] 和环境污染 [4] [5] 等多方面因素引发的危机,导致众多物种面临数量减少或灭绝的风险。这种趋势对生态结构和全球生物圈有着广泛的影响。生态系统中的生物实体通过多样的相互作用形成了复杂的网络模式,被称为食物网。食物网既是物质循环的平台,也是能量流动的主要通道,因此是生态系统的基础组成元素。多年来,生态学研究一直聚焦于物种多样性、食物网的构造和稳定性。为了维护生态系统和生物多样性,需要采取一系列措施。首先,减缓污染是保护生态系统的关键。人类活动产生的污染物对生态系统造成了极大的破坏,因此需要采取有效的措施减少污染物的排放。其次,应对气候变化也是保护生态系统的重要措施。气候变化对生态系统的影响非常大,包括海平面上升、气温升高、降雨模式改变等,这些都会对生态系统造成不可逆转的影响 [6] 。因此,需要采取措施减缓气候变化的影响,例如减少温室气体排放、开展植树造林等。此外,保护自然栖息地也是保护生态系统的重要措施。自然栖息地是生物生存和繁衍的基础,因此需要采取措施保护自然栖息地,例如建立自然保护区、限制开发等。最后,加强环境监控和管理也是保护生态系统的重要措施。环境监控和管理可以帮助我们了解生态系统的状况,及时发现问题并采取措施解决。
水晶河(Crystal River)是佛罗里达州中部墨西哥湾沿岸的一个小型但高度复杂的河口生态系统(见图1) [7] 。该系统以2.43平方公里的国王湾为水源,并通过一个10公里长的一级弹簧系统与墨西哥湾相连,该系统的流量达到或超过100立方英尺每秒。作为佛罗里达州最大的泉水系统之一,其受潮汐影响的平均流量约为14立方米每秒。本研究旨在通过食物网拓扑结构指数,基于水晶河食物网中物种的摄食关系来构建食物网模型。通过与其他水体食物网的比较,本研究评估了水晶河食物网的结构复杂性,以便为其功能性研究和生态保护提供理论支持。
2. 材料和方法
2.1. 数据来源
本研究采用了一个在多项先前研究中已经使用过的数据集 [8] [9] 。该数据集是由北卡罗来纳州大学威明顿分校的Stuart Borrett教授编制的,并涵盖了50个不同的食物网。在这些食物网中,我们特别关注了位于美国佛罗里达州水晶河核电站附近的水晶河(Crystal River)食物网 [10] 。
2.2. 网络拓扑分析
在食物网络矩阵里,数字0和1被用以标识两个物种间是否有捕食关系,其中0代表无捕食关系,1则表示存在捕食关系 [11] 。网络的可视化是通过Pajek软件完成的。为了量化水晶河食物网的复杂性和结构,我们计算了11种不同的网络拓扑指标 [12] [13] [14] [15] [16] 。这些指标包括:物种总数(S)、通过捕食关系建立的连接数量(L)、每个物种的平均连接数(L/S)、网络的连接密度(L/S2)、顶级物种数量(无捕食者)、中级物种数量(有捕食者和被捕食者)、基础物种数量(仅有捕食者,即初级生产者)、杂食性物种的比例(营养级跨度大于一的物种)以及连接复杂性指数(SC),该指数用于描述食物网的稳定性。其中,C的公式如下:
特征路径长度(ChPath):所有物种对之间的平均最短路径长度,
式中,ChPatmin(i, j)表示任意物种对之间的最短路径长度,2/S(S − 1)表示节点数 [17] 。
平均聚类系数CC:描述食物网中物种节点的聚集程度;将获得的水晶河食物网拓扑指数与其他水域作对比,以此来评估水晶河食物网的结构与复杂性 [11] 。
3. 结果与分析
本研究对水晶河食物网中的19种生物进行了分析,发现连接数超过10的物种仅有1种,占总物种数的5% (表1)。这表明该食物网具有较低的连接性。在图2中,不同物种节点的大小代表每个物种的连接数(包括捕食与被捕食)。海湾鳉鱼(GULF KILLIFISH)比较特殊,可以捕杀同种,因此需要用环形箭头来表示其连接数。在海州湾食物网中,连接数最高的物种是水底无脊椎动物(13条),而该食物网连接数最低的物种仅有1条连接。其中,有5个物种的连接数仅为1条,这5个物种均为顶级物种,这也说明该食物网具有较低的连接性。总的来说,这些结果表明,水晶河食物网的结构相对简单,连接性较低。这可能会对食物网的稳定性产生影响,因为连接性较低的食物网更容易受到外部干扰的影响。因此,进一步研究水晶河食物网的稳定性和生态系统功能对于保护该生态系统的健康和可持续性至关重要。

Table 1. Crystal River food web species and number of connections
表1. 水晶河食物网物种及其连接数
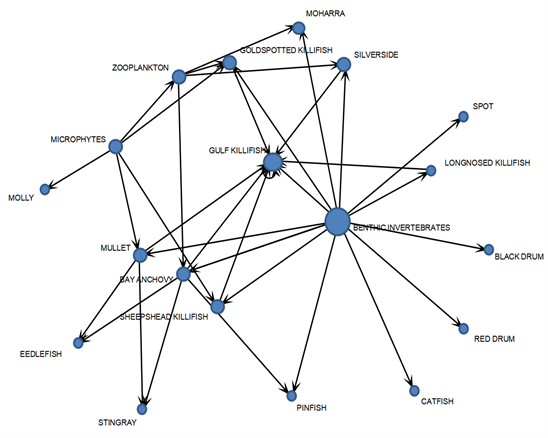
Figure 2. Topological structure diagram of Crystal River food web
图2. 水晶河食物网拓扑结构图
水晶河食物网的复杂性主要由每个物种的相互作用数量和连接性来评估(表2),通过数据分析,在19种物种之间含有64个连接数,即L = 64,该食物网的L/S较低,为3.42%,连接性L/S2为0.18 (表2)。
在水晶河食物网中,顶级物种的占比为47.4% (表2),意味着这个食物网的结构比较简单,其中存在较少的物种之间的相互作用。基础物种数的占比较小,仅占总物种数的10.5% (表2)。杂食性指标的大小对食物网稳定性有重要影响,与之相关的特征会对其产生显著效果。水晶河杂食性物种占比为36.8%,食物网的特征路径长度为2.111,平均聚类系数为0.325 (表2)。
Table 2. Crystal River food web structure and complexity index
表2. 水晶河食物网结构与复杂性指数
Table 3. Comparison of topological indices of Crystal River and other overseas aquatic food webs
表3. 水晶河与国外其他水域食物网拓扑指数的对比
4. 讨论
4.1. 水晶河食物网的复杂性
水晶河的食物网复杂性主要通过物种之间的相互作用数量(L/S)和连接性(L/S2)来评估。L/S和L/S2的数值大小影响着食物网的复杂性,根据过往的研究显示,随着L/S和L/S2的增大会导致食物网的复杂性增强 [25] 。在此次研究中测得L/S和L/S2的数值分别为3.42和0.18,相较于表3与其他食物网对比发现,本研究中的食物网的数值较低,由此可以确定水晶河食物网的连接性较低。食物网的复杂性程度会影响种群和群落的稳定性。水晶河食物网的L/S的数值处于非正常范围内、L/S2的数值均处于正常范围内,由此判定水晶河食物网的复杂性处于非正常的状态。根据表3的调查数据显示,水晶河的基础物种仅占食物网的10.5%,这一比例远低于资源型食物网的占比。此外,国际上的研究表明,随着对食物网生态系统的不断研究,S值会逐渐增大,这会导致L/S、L/S2的数值减少。这种趋势将进一步降低水晶河食物网的复杂性。因此,我们需要对水晶河食物网进行长期的研究,以更深入地了解其动态变化。这样的研究将有助于我们更好地理解水晶河食物网的生态系统,为保护和管理水晶河的生态环境提供科学依据。
4.2. 水晶河食物网结构特征
经过研究发现,水晶河食物网的结构不稳定,其中杂食性物种和中间物种的比例偏低,尤其是杂食性物种比例最低。这个比例的大小直接影响着食物网的稳定性 [26] 。研究结果表明,水晶河食物网的稳定性较低。虽然水晶河的特征路径长度高于其他食物网,但由于其物种数量较少且食物链路径较为单一,水晶河食物网的抵御外界干扰能力较弱。此外,水晶河的连接复杂性指数极低,表明其物种之间的连接不稳定,对食物网的稳定性不利。这意味着水晶河食物网中的物种之间缺乏相互作用,导致食物链的断裂和物种灭绝的风险增加。水晶河的平均聚类系数为0.325,较高的聚类系数指数意味着食物网中大部分生物之间存在较强的种间相互作用,这对于维持食物网的稳定性不利。因此,我们需要进一步研究水晶河食物网的结构和动态变化,以制定有效的保护和管理措施,维护水晶河生态系统的稳定性和健康发展。
综合分析表明,水晶河的食物网结构处于不稳定的状态。这种不稳定性主要表现在杂食性物种数量比例、特征路径长度和聚集系数等指标上。对水晶河食物网结构和复杂性的研究对于深入了解该生态系统的运作机制非常重要。同时,这些研究也为科学管理渔业资源提供了理论支持。通过深入研究水晶河食物网的功能和结构,我们可以更好地了解不同物种之间的相互作用,从而更好地保护和管理这些资源。因此,我们需要进一步加强对水晶河食物网的研究,以便更好地保护和管理这个生态系统。