摘要: 谷胱甘肽转移酶(GST)是一种重要的解毒酶,在抗氧化和细胞解毒中起着重要作用。二氯异氰尿酸钠属于含氯消毒剂,常用于养殖行业、自来水、医院等杀灭细菌。为了研究菲牛蛭GST基因在应对二氯异氰尿酸钠胁迫中的作用,本研究对菲牛蛭GST基因进行了克隆和表达规律研究。结果表明,菲牛蛭GST基因编码区全长是654 bp,编辑217个氨基酸,其蛋白质序列与红鲍螺,红鲍,绿叶海蛞蝓,角突臂围轮虫,紫贻贝相应序列相似性分别为97%,97%,97%,96%,96%。实时荧光定量PCR结果显示GST基因在胁迫后表达量先下降后升高,在12 h表达量达到最低,48 h表达量最高,是12 h表达量的93倍。研究结果显示菲牛蛭GST基因参与了二氯异氰尿酸钠胁迫的解毒过程,为今后菲牛蛭人工养殖过程中含氯消毒剂的合理使用,及解析GST在生物体内排毒机制提供了参考。
Abstract:
Glutathione transferase (GST) is an important detoxification enzyme, which plays an important role in antioxidant and cellular detoxification. Sodium dichloroisocyanurate is a chlorine-containing disinfectant, which is often used to kill bacteria in aquaculture industry, tap water, hospitals, etc. In order to study the role of GST gene in coping with sodium dichloroisocyanurate stress, the cloning and espression of GST gene in Poecilobdella manillensis were studied. The results showed that the total length of the GST gene coding region of the Poecilobdella manillensis was 654 bp, and the protein sequences were edited with 217 amino acids, and the protein sequences were 97%, 97%, 97%, 96% and 96% of the corresponding sequences of Haliotis rufescens, Haliotis rubra, Plakobranchus ocellatus, Brachionus angularis and Mytilus galloprovincialis. The real-time PCR results showed that the expression of GST gene decreased first and then increased after stress, reaching the lowest expression at 12 h and the highest expression at 48 h, which was 93 times the expression at 12 h. The results showed that the Poecilobdella manillensis GST gene participated in the detoxification process of sodium dichloroisocyanurate stress, which provided a reference for the rational use of chlorine-containing disinfectants in the artificial breeding process of Fifei bull leech in the future, and the analysis of the detoxification mechanism of GST in organisms.
1. 引言
菲牛蛭(Poecilobdella manillensis),又名马尼拟医蛭,俗称金边蚂蟥,属环节动物门,蛭纲,医蛭科,牛蛭属,高等无脊椎动物,雌雄同体 [1] ;以吮吸家畜与人等脊椎动物血液为生,体形较大,在国内广泛分布于广东、广西、海南岛、福建等地区,在国外分布于越南、泰国、印度、菲律宾等国家,喜生活在沼泽、沟渠、池塘、稻田等阴暗湿热环境中 [2] 。水蛭是我国一味传统中药材,据《神农本草经》记载,其具有破血、散瘀、通经的功效;主治血瘀经闭腹痛、症瘕积聚,对治疗跌打损伤也有良好效果 [3] 。
水蛭素是存在于水蛭唾液腺中的一种生物活性成分,具有抗凝血和血栓的作用,且作用专一,相对于市面上常用的肝素、重组水蛭素等,其具有特异性强、抑制效果好、无副作用等优势,是一种安全的天然凝血酶抑制剂 [4] [5] [6] 。
二氯异氰尿酸钠是一种高效、广谱的新型杀菌剂,常用于养殖行业杀灭细菌,起到消毒作用的成分主要为水解后的次氯酸 [7] 。二氯异氰尿酸钠有强腐蚀性,高浓度对黏膜、皮肤等有灼伤危险,误服后可出现呕吐等症状 [8] 。
谷胱甘肽S-转移酶(Glutathione S-transferase, GSTs)是同或异形二聚体,含有多个亚基,属于多基因家族,是重要的解毒酶,存在于多种动植物体内。GSTs可以对内、外源性化合物进行解毒,产生水溶性代谢物,并将其排出体外,以减少有毒物质在体内造成的损害 [9] 。主要有谷胱甘肽S-烷基转移酶、谷胱甘肽S-芳基转移酶、谷胱甘肽S-芳烷基转移酶、谷胱甘肽S-环氧化物转移酶、谷胱甘肽S-烯烃转移酶五大类。淡水鱼类的GST表达量与抗微囊藻毒素能力呈现高程度的相关性,用稀释不同倍数的磺胺嘧啶刺激三疣梭子蟹,其GST表达水平在下降后又上升 [10] ,对外界压力表现出了高度的敏感性,说明GST基因与生物体的排毒代谢息息相关,未来有可能作为衡量水体污染程度的指标。然而在菲牛蛭的研究上,大多集中在养殖、营养成分及水蛭素等方面 [11] [12] [13] ,因此本研究开展了菲牛蛭GST基因的克隆与表达分析,为菲牛蛭养殖提供理论基础。
2. 材料与方法
2.1. 实验材料
实验材料为活力较高的菲牛蛭幼体,于2023年5月在广西玉林采集,由科康集团提供,选取体型大小较为一致,平均体重1.01 ± 0.1 g的菲牛蛭幼体进行实验,实验开始前7天暂养于曝气的淡水中,温度、pH和溶解氧分别保持在在25℃~28℃、7.5~8.0和6.8~7 mg/L。养殖容器为圆柱形透明塑料罐,底边直径和高为15 cm、20 cm,暂养及实验时加入1 L水。
2.2. 菲牛蛭胁迫实验
将60条菲牛蛭幼苗随机分为两组,对照组和实验组,每组30条。在实验组中加入终浓度为6 mg/L的二氯异氰尿酸钠,并于3、6、9、12、24和48 h取对照组和实验组存活小水蛭全体。
2.3. 菲牛蛭总RNA的提取和cDNA合成
采用TRIzol法提取菲牛蛭的总RNA,并利用NanoDrop2000在260 nm处测量吸光度来估计其浓度。通过电泳检查RNA完整性,并将样品保存在−80℃以备后用。逆转录反应使用带有gDNA橡皮擦(TaKaRa,产品编号:RR047A)的Prime Script™RT试剂盒进行。我们采用厂家推荐的反应条件,分为以下两步:第一步去除gDNA,第二步将RNA逆转录为cDNA。第一步反应条件为42℃反应5 min,第二步反应条件为37℃反应60 min,85℃反应5 min,然后保持4℃。
2.4. 基因克隆
利用逆转录聚合酶链反应(RT-PCR)克隆GST基因编码区序列。利用Oligo6.0软件设计GST特异性扩增引物(表1)。将扩增的cDNA片段连接到pEASY®-Blunt Simple Cloning vectors (TransGenBiotech, China)上,通过蓝/白筛选和PCR鉴定重组菌。采用含有插入的GST片段的质粒作为模板进行DNA测序。
2.5. 生物信息学分析
同源序列比对使用国家生物技术信息中心(NCBI)的BLAST程序,Expert Protein Analysis System (EXPASY)服务器(https://web.expasy.org/protparam/)用于氨基酸组成分析、分子质量和等电点预测。Softberry程序(http://linux1.softberry.com/)用于预测亚细胞定位,Signalp程序用于预测n端信号肽。通过Motif Scan (https://myhits.sib.swiss/cgi-bin/motif_scan)对蛋白功能位点进行检测和分析。用Clustalx软件比较氨基酸序列。使用MEGA 10.0进行序列比对和系统发育分析(邻居连接法,有1000个假重复),I-TASSER程序(http://zhanglab.ccmb.med.umich.edu/I-TASSER/)进行蛋白质三级结构预测,并使用PyMOL软件进行三级结构比较。使用NCBI保守结构域搜索(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)检测保守蛋白结构域。
2.6. GST基因在不同组织中的相对表达
实时定量RT-PCR (qRT-PCR)分析在实时PCR系统(美国BIO-RAD CFX Connect)上进行,使用SYBR PreMix ExTaq™ (Takara,大连,中国)。新鲜组织(菲牛蛭幼体)作为实验材料。RNA提取和cDNA获取的实验条件同2.2节。用定量引物GSTdl (表1)扩增GST片段,用Ef1 (Ef1-dl)定量引物(表1)扩增对照内参基因。
2.7. 统计分析
采用对比CT法相对定量靶基因表达水平,定量归一化至对照基因Ef1。实验组个体的靶基因相对于对照组个体的靶基因表达量为2−ΔΔct。所有数据均以均数 ± 标准差表示,采用SPSS Statistics 26进行统计分析。采用Duncan法对不同胁迫时间组间实验数据进行比较,以p ≤ 0.05为显著差异标准。
3. 结果
3.1. GST基因的克隆及序列分析
提取菲牛蛭总RNA后进行电泳,如图1A,在电泳图中18S和5S都较清晰存在,说明我们的实验得到了良好的结果,并且已经得到了能够进入下一步逆向转录的优质RNA。GST基因扩增结果如图1B所示,其长度为824 bp。
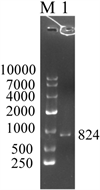
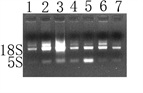 (A)(B)注:(A) 菲牛蛭总RNA电泳图;(B) M为DL10000 DNA Marker,1为样品RT-PCR GST扩增产物。
(A)(B)注:(A) 菲牛蛭总RNA电泳图;(B) M为DL10000 DNA Marker,1为样品RT-PCR GST扩增产物。
Figure 1. Total RNA extraction and GST gene amplification in Poecilobdella manillensis
图1. 菲牛蛭总RNA提取与GST基因扩增
3.2. 菲牛蛭GST基因编码蛋白质的修饰位点预测
菲牛蛭GST基因的Motif位点如图2所示,GST蛋白包含2个N-肉豆蔻酰化位点(47GLTFGQ52, 155GLLEAL160);1个糖基化位点(138NGSN141);1个ATP结合位点(76AHNLYGKT83);2个磷酸化位点(88TKVD91, 184TMSE187);1个酪蛋白激酶C磷酸化位点(198TRK200) (如图2所示)。
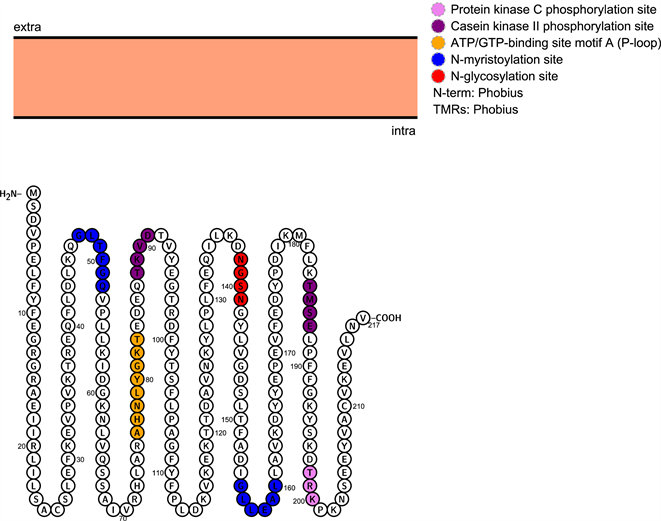
Figure 2. Map of Poecilobdella manillensis active sites
图2. 菲牛蛭活性位点图
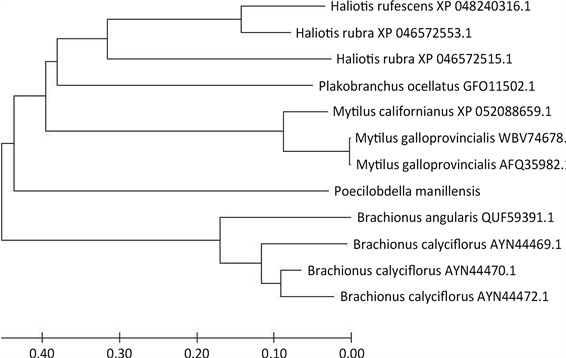
Figure 3. Poecilobdella manillensis with other species GST evolutionary tree
图3. 菲牛蛭与其他物种GST进化树
3.3. 构建菲牛蛭GST基因的系统进化树
菲牛蛭与其他物种GST基因的系统进化树如图3所示,其GST蛋白质序列与红鲍螺,红鲍,绿叶海蛞蝓,角突臂围轮虫,紫贻贝相应序列相似性分别为97%,97%,97%,96%,96%,从侧面反映了该基因在多种无脊椎动物的生活中具有重要意义且在进化的过程中高度保留。
3.4. 菲牛蛭GST基因其他生物信息学分析结果
EXPASY分析结果如图4所示,GST基因编码217个氨基酸,理论上编码区全长是654 bp,氨基酸组成的分子化学方程式为C1136H1760N278O334S5,其中,亮氨酸(Leu)含量最多,为12%,其次为赖氨酸(Lys)含量为9.2%,含量最少的是半胱氨酸(Cys),含量为0.9%,GST为弱酸性蛋白,等电点为5.15,理论分子量为24816.43 Da。

Figure 4. Amino acid sequence prediction of Poecilobdella manillensis GST gene
图4. 菲牛蛭GST基因及氨基酸序列预测
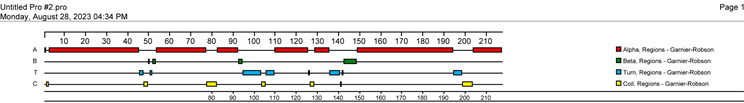
Figure 5. Secondary structure prediction of GST protein in Poecilobdella manillensis
图5. 菲牛蛭GST蛋白质二级结构预测
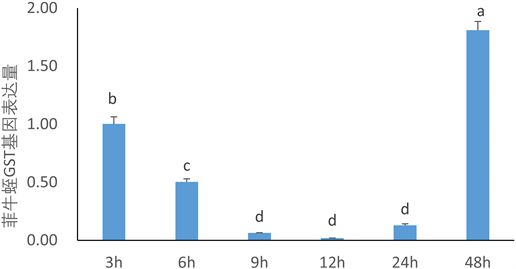
Figure 6. GST expression in Poecilobdella manillensis at different time points under sodium dichloroisocyanurate stress
图6. 二氯异氰尿酸钠胁迫下不同时间点菲牛蛭中GST表达量
利用Softberry服务器系统研究和分析了菲牛蛭GST蛋白的亚细胞定位情况,Extracellular的分值是1.3,可以进行测序进而分析其所编码的蛋白质的二级结构,通过DNA Star软件中的Protean程序,结果见图5,菲牛蛭GST蛋白质的二级结构包含了α-螺旋7个,β-折叠4个,T-转角8个与7个无规则卷曲。
3.5. 菲牛蛭不同组织中GST基因表达分析
该实验利用QRT-PCR技术以Ef1a作为内参考基因对菲牛蛭GST基因在胁迫实验中不同时间点的表达进行了全面系统的研究。GST基因表达量先下降后升高,在12 h表达量最低,48 h表达量最高,是12 h表达量的93倍(图6)。
4. 讨论
谷胱甘肽转移酶(GST)是多功能酶,参与许多毒素的解毒过程,包括除草剂,杀虫剂和药物。因此,GST的基因表达情况经常被用作环境研究中的生物标志 [14] 。目前GST基因的研究主要集中在黑腹果蝇(Drosophila melanogaster) [15] 、冈比亚疟蚊(Anopheles gambiae) [16] ,在许多水生动物中GST基因也被鉴定和表征,如近江牡蛎(Crassostrea ariakensi) [17] 、太平洋斯氏柔鱼(Ommastrephes sloani pacificus) [18] 、凡纳滨对虾(Litopenaeus vannamei) [19] 等,这些研究主要集中在致病性和毒性化学物质胁迫下的生理生化特征和表达谱。然而,对环节动物中GST基因的研究较少,尤其是水蛭。在这项研究中,我们从菲牛蛭体内克隆出GST基因。
本实验从菲牛蛭中提取总RNA后,纯化得到了优质RNA,CDS长度达到654 bp与预期效果相近。生物信息学分析结果为,GST基因编码217个氨基酸,编码区全长654 bp,氨基酸组成的分子化学方程式为C1136H1760N278O334S5,其中,亮氨酸(Leu)含量最多,为12%,其次为赖氨酸(Lys)含量为9.2%,含量最少的是半胱氨酸(Cys),含量为0.9%,GST为弱酸性蛋白,等电点为5.15,理论分子量为24816.43 Da,与孙盛明等人发现的在团头鲂中GST基因CDs长度为654 bp,编码218个蛋白,蛋白分子量为25.95 kD,等电点(pI)为5.44的结果相一致 [20] 。
GST在抗氧化和细胞解毒中起着重要作用 [21] ,这可能存在于易受感染的生物系统。本研究分析了GST基因在菲牛蛭在二氯异氰尿酸钠胁迫下不同时间点的表达量的变化模式。qRT-PCR结果表明,二氯异氰尿酸钠胁迫后GST基因表达量先下降后升高,在12小时表达量达到最低,并在48小时达到最高,可能是在免疫前期表达量较高,中期由其他免疫基因发挥拮抗作用,后期表达量又升高,再次发挥免疫作用。
消毒剂在水产养殖中有着重要作用,主要用于池塘、水体清塘消毒,苗种下塘前药浴,疾病防治等,在科学指导下,合理使用消毒剂能有效消灭养殖环境与过程中的病原微生物等致病因素,防止疾病的发生 [22] 。目前对消毒剂的研究主要集中在了氯氰菊酯 [23] 、高锰酸钾 [24] 、三唑酮 [25] ,对于二氯异氰尿酸钠的研究较少。
5. 结论
菲牛蛭作为一种药用、经济价值高的的经济物种,其应用领域涉及医用、保健、美容等多个领域,并且还在不断拓展。由于菲牛蛭天然水蛭素含量远高于其他水蛭,且天然水蛭素在药效、安全性等多个方面远远优于肝素和重组水蛭素,这使得菲牛蛭的越来越受到市场和养殖户的青睐。本研究探讨了二氯异氰尿酸钠对菲牛蛭幼体胁迫后不同时间点GST基因表达量变化,为菲牛蛭养殖过程中含氯消毒剂的合理使用提供了参考。
基金项目
玉林市科学研究与技术开发计划(玉市科20204337)。