
Co-Expression of JcNAC1- and JcZFP8-Improved Agronomic Traits of Tobacco and Enhanced Drought Resistance through NbbHLH1 and NbbHLH2
 , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Co-Expression of JcNAC1/JcZFP8 Not Only Improves PEG-Drought Resistance, but Also Overcomes the Adverse Agronomic Traits Brought by Single Gene Transformation
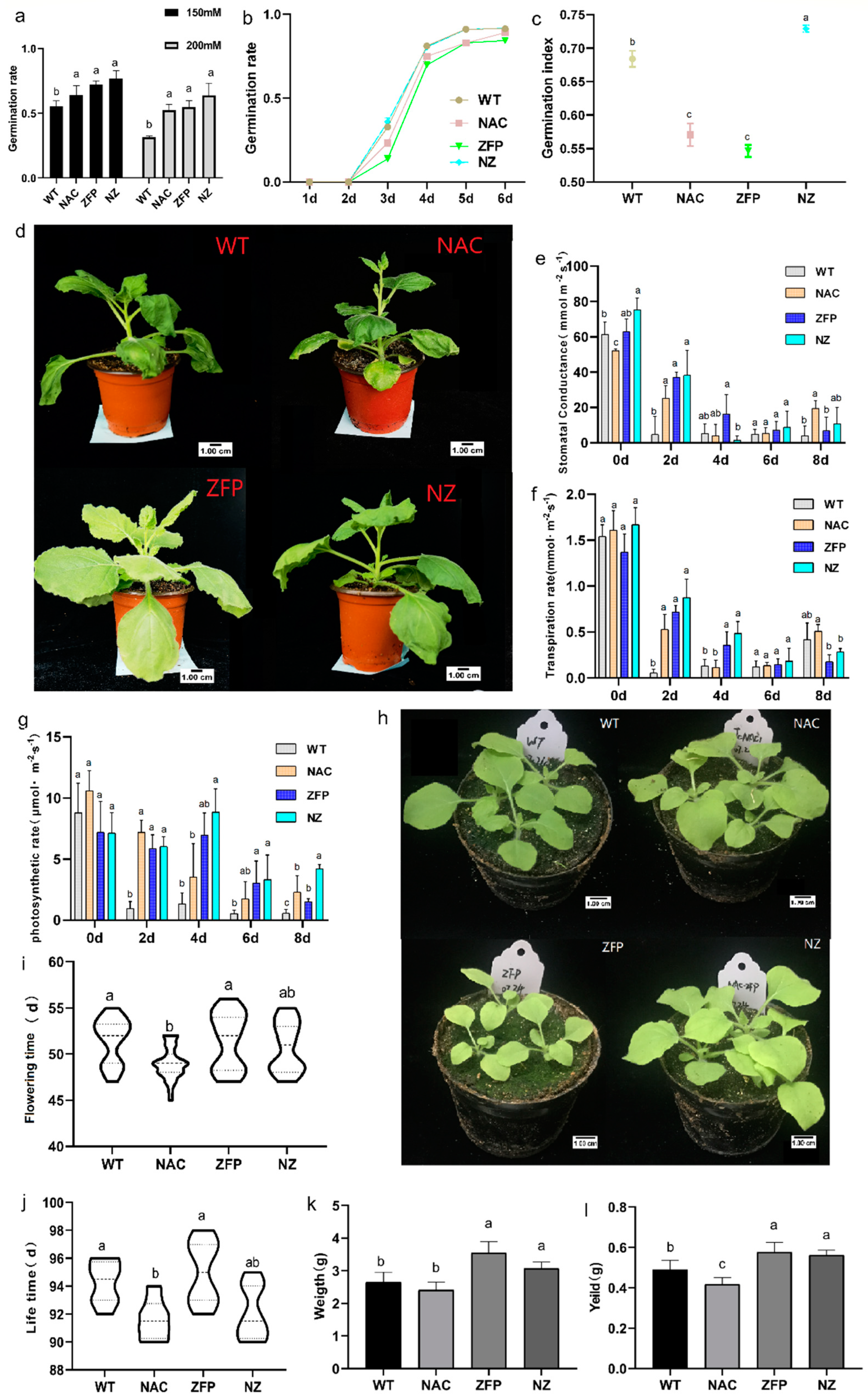
2.1.1. Co-Expression of JcNAC1 and JcZFP8 Can Increase the Germination Rate of Plants under Mannitol-Drought
2.1.2. NAC, ZFP, and NZ Transgenic Tobacco Performed Better in PEG-Drought (Low Water Potential) Compared to WT
2.1.3. Double Gene Co-Expressing Plants Overcome the Adverse Agronomic Traits Brought by Single Gene Transformation
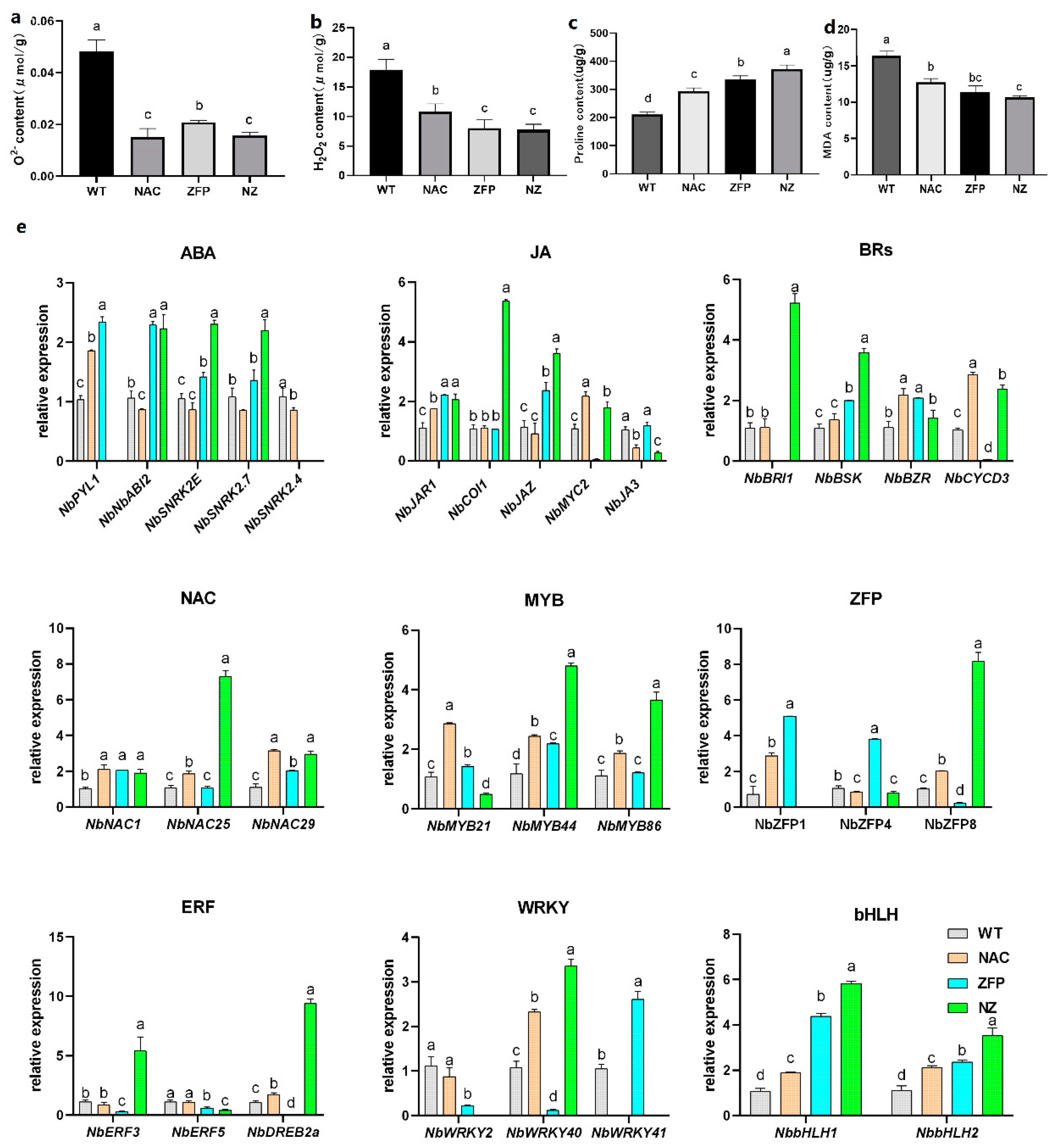
2.2. More Proline and Less Reactive Oxygen Species (ROS) Level Endows the Double Gene Overexpression Lines with Stronger PEG-Drought Resistance
2.2.1. Under 10%PEG6000 Treatment, the Accumulation of ROS in Transgenic Plants Was Less
2.2.2. NbbHLH1 and NbbHLH2 Genes Increased Linearly from WT to Single-Overexpression Lines to NZ under PEG-Drought
2.3. The NbbHLH1 and NbbHLH2 Were Involved in Drought Resistance of Tobacco
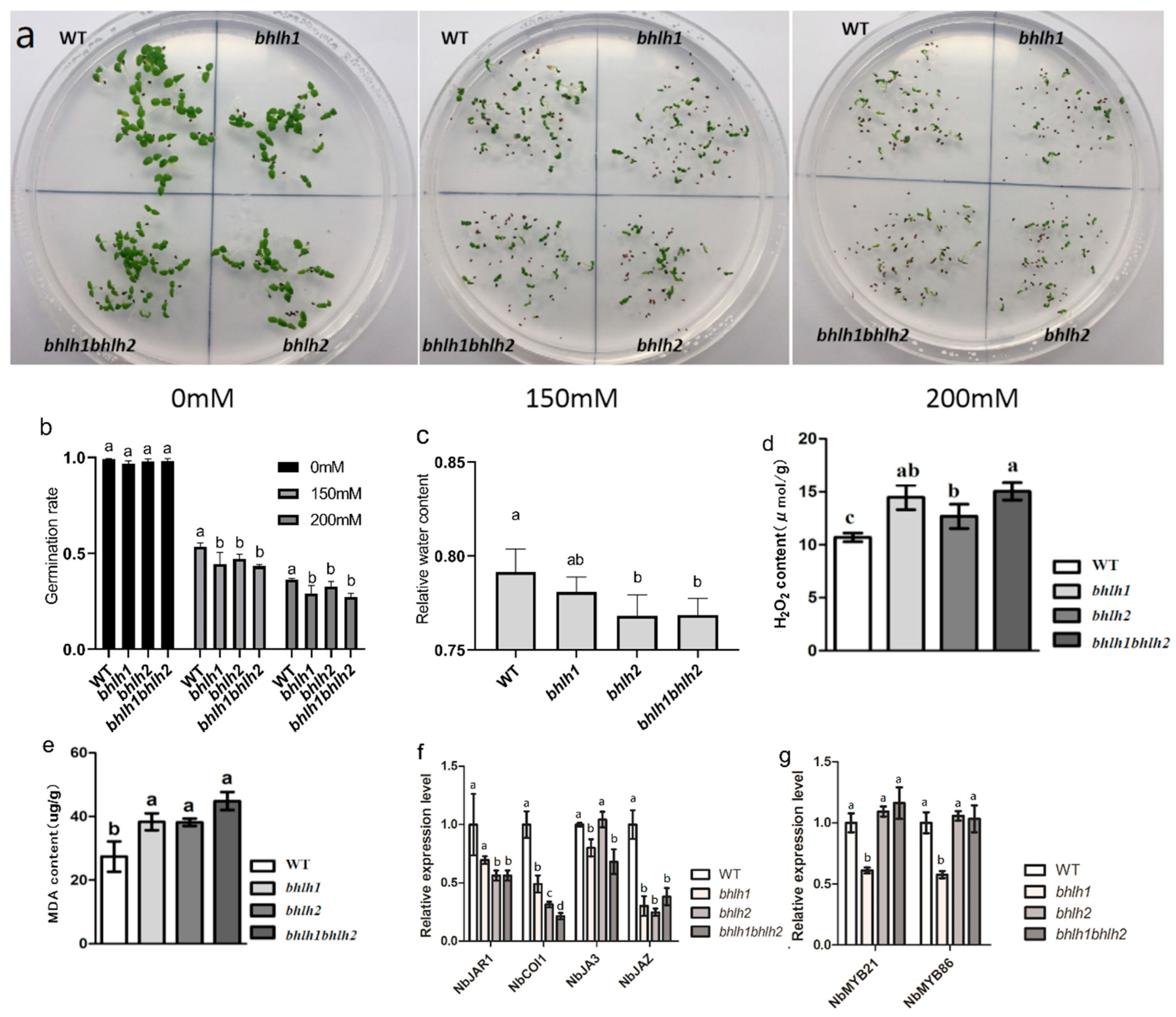
2.3.1. The Germination Rate of bhlh1, bhlh2, and bhlh1bhlh2 Decreased under Mannitol Treatment
2.3.2. H2O2 and MDA Was Increased in bhlh1, bhlh2, and bhlh1bhlh2
2.3.3. NbbHLH1 and NbbHLH2 May Not Affect Drought Resistance by Regulating the Expression of NbMYB21 and NbMYB86
3. Discussion
4. Materials and Methods
4.1. Construction and Transformation of JcNAC1, JcZFP8, and JcNAC1-JcZFP8
4.2. Construction and Transformation of NbbHLH1, NbbHLH2, and NbbHLH1–NbbHLH2
4.3. Plant Materials and Drought Stress Treatments
4.4. Germination Rates and Other Growth Trait
4.5. Flowering Period, Life Cycle, Weight, and Yield
4.6. Photosynthesis, Stomatal Conductance, and Transpiration Rate
4.7. Expression Analysis of Important Genes in Drought Treatments
4.8. Measurements of Physiological Parameters
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Diatta, A.A.; Min, D.; Jagadish, S.V.K. Chapter Two—Drought stress responses in non-transgenic and transgenic alfalfa—Current status and future research directions. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Cambridge, MA, USA, 2021; Volume 170, pp. 35–100. [Google Scholar]
- Shinwari, Z.K.; Jan, S.A.; Nakashima, K.; Yamaguchi-Shinozaki, K. Genetic engineering approaches to understanding drought tolerance in plants. Plant Biotechnol. Rep. 2020, 14, 151–162. [Google Scholar] [CrossRef]
- Selvaraj, M.G.; Jan, A.; Ishizaki, T.; Valencia, M.; Dedicova, B.; Maruyama, K.; Ogata, T.; Todaka, D.; Yamaguchi-Shinozaki, K.; Nakashima, K.; et al. Expression of the CCCH-tandem zinc finger protein gene OsTZF5 under a stress-inducible promoter mitigates the effect of drought stress on rice grain yield under field conditions. Plant Biotechnol. J. 2020, 18, 1711–1721. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Annu. Rev. Plant Biol. 2006, 57, 781–803. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Yamaguchi-Shinozaki, K. Metabolic engineering: Towards water deficiency adapted crop plants. J. Plant Physiol. 2021, 258–259, 153375. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, R.; Kobayashi, Y.; Koyama, H.; Sahoo, L. Cowpea NAC1/NAC2 transcription factors improve growth and tolerance to drought and heat in transgenic cowpea through combined activation of photosynthetic and antioxidant mechanisms. J. Integr. Plant Biol. 2022, 65, 25–44. [Google Scholar] [CrossRef]
- Alam, I.; Batool, K.; Cui, D.L.; Yang, Y.Q.; Lu, Y.H. Comprehensive genomic survey, structural classification and expression analysis of C2H2 zinc finger protein gene family in Brassica rapa L. PLoS ONE 2019, 14, e0216071. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Fang, L.; Zhu, Z.; Zhang, L.; Sun, X.; Wang, Y.; Wang, Q.; Li, S.; Xin, H. The transcription factor VaNAC17 from grapevine (Vitis amurensis) enhances drought tolerance by modulating jasmonic acid biosynthesis in transgenic Arabidopsis. Plant Cell Rep. 2020, 39, 621–634. [Google Scholar] [CrossRef]
- Wang, Y.; Liao, Y.; Quan, C.; Li, Y.; Yang, S.; Ma, C.; Mo, Y.; Zheng, S.; Wang, W.; Xu, Z.; et al. C2H2-type zinc finger OsZFP15 accelerates seed germination and confers salinity and drought tolerance of rice seedling through ABA catabolism. Environ. Exp. Bot. 2022, 199, 104873. [Google Scholar] [CrossRef]
- Yuan, X.; Huang, P.; Wang, R.; Li, H.; Lv, X.; Duan, M.; Tang, H.; Zhang, H.; Huang, J. A Zinc Finger Transcriptional Repressor Confers Pleiotropic Effects on Rice Growth and Drought Tolerance by Down-Regulating Stress-Responsive Genes. Plant Cell Physiol. 2018, 59, 2129–2142. [Google Scholar] [CrossRef]
- Liang, Y.; Ma, F.; Li, B.; Guo, C.; Hu, T.; Zhang, M.; Liang, Y.; Zhu, J.; Zhan, X. A bHLH transcription factor, SlbHLH96, promotes drought tolerance in tomato. Hortic. Res. 2022, 9, uhac198. [Google Scholar] [CrossRef]
- Todd, A.T.; Liu, E.; Polvi, S.L.; Pammett, R.T.; Page, J.E. A functional genomics screen identifies diverse transcription factors that regulate alkaloid biosynthesis in Nicotiana benthamiana. Plant J. 2010, 62, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Yan, C.; Sun, Q.; Wang, J.; Yuan, C.; Mou, Y.; Shan, S.; Zhao, X. The bHLH transcription factor AhbHLH112 improves the drought tolerance of peanut. BMC Plant Biol. 2021, 21, 540. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.; Wan, S.; Ma, Q.; Yang, L.; Hu, W.; Kuang, L.; Xie, J.; Huang, Y.; Liu, D.; Liu, Y. A Novel bHLH Transcription Factor PtrbHLH66 from Trifoliate Orange Positively Regulates Plant Drought Tolerance by Mediating Root Growth and ROS Scavenging. Int. J. Mol. Sci. 2022, 23, 15053. [Google Scholar] [CrossRef] [PubMed]
- Shailani, A.; Joshi, R.; Singla-Pareek, S.L.; Pareek, A. Stacking for future: Pyramiding genes to improve drought and salinity tolerance in rice. Physiol. Plant. 2021, 172, 1352–1362. [Google Scholar] [CrossRef]
- Kudo, M.; Kidokoro, S.; Yoshida, T.; Mizoi, J.; Todaka, D.; Fernie, A.R.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Double overexpression of DREB and PIF transcription factors improves drought stress tolerance and cell elongation in transgenic plants. Plant Biotechnol. J. 2017, 15, 458–471. [Google Scholar] [CrossRef]
- Kudo, M.; Kidokoro, S.; Yoshida, T.; Mizoi, J.; Kojima, M.; Takebayashi, Y.; Sakakibara, H.; Fernie, A.R.; Shinozaki, K.; Yamaguchi-Shinozaki, K. A gene-stacking approach to overcome the trade-off between drought stress tolerance and growth in Arabidopsis. Plant J. 2019, 97, 240–256. [Google Scholar] [CrossRef]
- Zhang, Y.; Deng, G.; Fan, W.; Yuan, L.; Wang, H.; Zhang, P. NHX1 and eIF4A1-stacked transgenic sweetpotato shows enhanced tolerance to drought stress. Plant Cell Rep. 2019, 38, 1427–1438. [Google Scholar] [CrossRef]
- Srivastava, R.; Kobayashi, Y.; Koyama, H.; Sahoo, L. Overexpression of cowpea NAC transcription factors promoted growth and stress tolerance by boosting photosynthetic activity in Arabidopsis. Plant Sci. 2022, 319, 111251. [Google Scholar] [CrossRef]
- Qin, X.; Zheng, X.; Huang, X.; Lii, Y.; Shao, C.; Xu, Y.; Chen, F. A novel transcription factor JcNAC1 response to stress in new model woody plant Jatropha curcas. Planta 2014, 239, 511–520. [Google Scholar] [CrossRef]
- Shi, X.; Wu, Y.; Dai, T.; Gu, Y.; Wang, L.; Qin, X.; Xu, Y.; Chen, F. JcZFP8, a C2H2 zinc finger protein gene from Jatropha curcas, influences plant development in transgenic tobacco. Electron. J. Biotechnol. 2018, 34, 76–82. [Google Scholar] [CrossRef]
- Manna, M.; Thakur, T.; Chirom, O.; Mandlik, R.; Deshmukh, R.; Salvi, P. Transcription factors as key molecular target to strengthen the drought stress tolerance in plants. Physiol. Plant. 2020, 172, 847–868. [Google Scholar] [CrossRef]
- Yoon, Y.; Seo, D.H.; Shin, H.; Kim, H.J.; Kim, C.M.; Jang, G. The Role of Stress-Responsive Transcription Factors in Modulating Abiotic Stress Tolerance in Plants. Agronomy 2020, 10, 788. [Google Scholar] [CrossRef]
- Sengupta, S.; Pehlivan, N.; Mangu, V.; Rajasekaran, K.; Baisakh, N. Characterization of a Stress-Enhanced Promoter from the Grass Halophyte, Spartina alterniflora L. Biology 2022, 11, 1828. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, F.; Shah, S.H.; Jan, A. Overexpression of the DREB1A gene under stress-inducible promoter delays leaf senescence and improves drought tolerance in rice. Cereal Res. Commun. 2023, 11, 1828. [Google Scholar] [CrossRef]
- Xiao, S.; Wu, Y.; Xu, S.; Jiang, H.; Hu, Q.; Yao, W.; Zhang, M. Field evaluation of TaDREB2B-ectopic expression sugarcane (Saccharum spp. hybrid) for drought tolerance. Front. Plant Sci. 2022, 13, 963377. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Montesino, R.; Bouza-Morcillo, L.; Marquez, J.; Ghita, M.; Duran-Nebreda, S.; Gomez, L.; Holdsworth, M.J.; Bassel, G.; Onate-Sanchez, L. A Regulatory Module Controlling GA-Mediated Endosperm Cell Expansion Is Critical for Seed Germination in Arabidopsis. Mol. Plant 2019, 12, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.E.; Bang, S.W.; Kim, S.H.; Seo, J.S.; Yoon, H.-B.; Kim, Y.S.; Kim, J.-K. Overexpression of OsERF83, a Vascular Tissue-Specific Transcription Factor Gene, Confers Drought Tolerance in Rice. Int. J. Mol. Sci. 2021, 22, 7656. [Google Scholar] [CrossRef]
- Tang, K.; Dong, B.; Wen, X.; Yin, Y.; Xue, M.; Su, Z.; Wang, M. Ectopic expression of the AmDREB1F gene from Ammopiptanthus mongolicus enhances stress tolerance of transgenic Arabidopsis. Sheng Wu Gong Cheng Xue Bao = Chin. J. Biotechnol. 2021, 37, 4329–4341. [Google Scholar] [CrossRef]
- Nguyen, K.H.; Mostofa, M.G.; Watanabe, Y.; Tran, C.D.; Rahman, M.M.; Tran, L.-S.P. Overexpression of GmNAC085 enhances drought tolerance in Arabidopsis by regulating glutathione biosynthesis, redox balance and glutathione-dependent detoxification of reactive oxygen species and methylglyoxal. Environ. Exp. Bot. 2019, 161, 242–254. [Google Scholar] [CrossRef]
- Kobayashi, M.; Horiuchi, H.; Fujita, K.; Takuhara, Y.; Suzuki, S. Characterization of grape C-repeat-binding factor 2 and B-box-type zinc finger protein in transgenic Arabidopsis plants under stress conditions. Mol. Biol. Rep. 2012, 39, 7933–7939. [Google Scholar] [CrossRef]
- Srivastava, R.; Sahoo, L. Balancing yield trade-off in legumes during multiple stress tolerance via strategic crosstalk by native NAC transcription factors. J. Plant Biochem. Biotechnol. 2021, 30, 708–729. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, W.; Ran, L.; Chen, Z.; Wang, C.; Dou, Y.; Qin, Y.; Suo, Q.; Li, Y.; Zeng, J.; et al. DELLA-NAC Interactions Mediate GA Signaling to Promote Secondary Cell Wall Formation in Cotton Stem. Front. Plant Sci. 2021, 12, 655127. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Chai, Z.; Yao, W.; Chen, B.; Zhang, M. Interactions between ScNAC23 and ScGAI regulate GA-mediated flowering and senescence in sugarcane. Plant Sci. 2021, 304, 110806. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Koyama, H.; Bhati, K.K.; Alok, A. The biotechnological importance of the plant-specific NAC transcription factor family in crop improvement. J. Plant Res. 2021, 134, 475–495. [Google Scholar] [CrossRef]
- Du, X.; Su, M.; Jiao, Y.; Xu, S.; Song, J.; Wang, H.; Li, Q. A Transcription Factor SlNAC10 Gene of Suaeda liaotungensis Regulates Proline Synthesis and Enhances Salt and Drought Tolerance. Int. J. Mol. Sci. 2022, 23, 9625. [Google Scholar] [CrossRef]
- Liu, S.; Liu, Y.; Liu, C.; Yu, X.; Ma, H. Chickpea C2H2-Type Zinc Finger Protein ZF2 is a Positive Regulator in Drought Response in Arabidopsis. Phyton 2023, 92, 577–590. [Google Scholar] [CrossRef]
- Pascual, M.B.; Canovas, F.M.; Avila, C. The NAC transcription factor family in maritime pine (Pinus Pinaster): Molecular regulation of two genes involved in stress responses. BMC Plant Biol. 2015, 15, 254. [Google Scholar] [CrossRef]
- Han, G.; Wei, X.; Dong, X.; Wang, C.; Sui, N.; Guo, J.; Yuan, F.; Gong, Z.; Li, X.; Zhang, Y.; et al. Arabidopsis ZINC FINGER PROTEIN1 Acts Downstream of GL2 to Repress Root Hair Initiation and Elongation by Directly Suppressing bHLH Genes. Plant Cell 2020, 32, 206–225. [Google Scholar] [CrossRef]
- Guo, J.; Sun, B.; He, H.; Zhang, Y.; Tian, H.; Wang, B. Current Understanding of bHLH Transcription Factors in Plant Abiotic Stress Tolerance. Int. J. Mol. Sci. 2021, 22, 4921. [Google Scholar] [CrossRef]
- Gao, S.; Li, C.; Chen, X.; Li, S.; Liang, N.; Wang, H.; Zhan, Y.; Zeng, F. Basic helix-loop-helix transcription factor PxbHLH02 enhances drought tolerance in Populus (Populus simonii x P. nigra). Tree Physiol. 2022, 43, 185–202. [Google Scholar] [CrossRef]
- Ren, Y.-R.; Yang, Y.-Y.; Zhao, Q.; Zhang, T.-E.; Wang, C.-K.; Hao, Y.-J.; You, C.-X. MdCIB1, an apple bHLH transcription factor, plays a positive regulator in response to drought stress. Environ. Exp. Bot. 2021, 188, 104523. [Google Scholar] [CrossRef]
- Wang, J.; Song, L.; Gong, X.; Xu, J.; Li, M. Functions of Jasmonic Acid in Plant Regulation and Response to Abiotic Stress. Int. J. Mol. Sci. 2020, 21, 1446. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wu, Y.; Tian, Y.; Dai, T.; Xie, G.; Xu, Y.; Chen, F. Overexpressing Jatropha curcas CBF2 in Nicotiana benthamiana improved plant tolerance to drought stress. Gene 2020, 742, 144588. [Google Scholar] [CrossRef] [PubMed]
- Xing, H.-L.; Dong, L.; Wang, Z.-P.; Zhang, H.-Y.; Han, C.-Y.; Liu, B.; Wang, X.-C.; Chen, Q.-J. A CRISPR/Cas9 toolkit for multiplex genome editing in plants. BMC Plant Biol. 2014, 14, 327. [Google Scholar] [CrossRef] [PubMed]
- de Souza, R.R.; Moraes, M.P.; Paraginski, J.A.; Moreira, T.F.; Bittencourt, K.C.; Toebe, M. Effects of Trichoderma asperellum on Germination Indexes and Seedling Parameters of Lettuce Cultivars. Curr. Microbiol. 2021, 79, 5. [Google Scholar] [CrossRef] [PubMed]
- Lee Yun, D.; Jung Song, S.; Jin Kim, Y. Plant Maturity and Vernalization Affect Flowering in Dianthus japonicus Thunb. Hortic. J. 2020, 89, 37–44. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; Berni, J.A.J.; Suárez, L.; Sepulcre-Cantó, G.; Morales, F.; Miller, J.R. Imaging chlorophyll fluorescence with an airborne narrow-band multispectral camera for vegetation stress detection. Remote Sens. Environ. 2009, 113, 1262–1275. [Google Scholar] [CrossRef]
- Yan, H.; Jia, H.; Chen, X.; Hao, L.; An, H.; Guo, X. The cotton WRKY transcription factor GhWRKY17 functions in drought and salt stress in transgenic Nicotiana benthamiana through ABA signaling and the modulation of reactive oxygen species production. Plant Cell Physiol. 2014, 55, 2060–2076. [Google Scholar] [CrossRef]
- Tian, H.; Zhou, Q.; Liu, W.; Zhang, J.; Chen, Y.; Jia, Z.; Shao, Y.; Wang, H. Responses of photosynthetic characteristics of oat flag leaf and spike to drought stress. Front. Plant Sci. 2022, 13, 917528. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, X.; Lai, Z.; Wang, L.; Ma, R.; Ren, Y.; Wang, X.; Cheng, C.; Wang, T.; Chen, F.; Xu, Y. Co-Expression of JcNAC1- and JcZFP8-Improved Agronomic Traits of Tobacco and Enhanced Drought Resistance through NbbHLH1 and NbbHLH2. Plants 2023, 12, 3029. https://doi.org/10.3390/plants12173029
Niu X, Lai Z, Wang L, Ma R, Ren Y, Wang X, Cheng C, Wang T, Chen F, Xu Y. Co-Expression of JcNAC1- and JcZFP8-Improved Agronomic Traits of Tobacco and Enhanced Drought Resistance through NbbHLH1 and NbbHLH2. Plants. 2023; 12(17):3029. https://doi.org/10.3390/plants12173029
Chicago/Turabian StyleNiu, Xianfei, Zhiping Lai, Linghui Wang, Rui Ma, Yingying Ren, Xueying Wang, Cheng Cheng, Ting Wang, Fang Chen, and Ying Xu. 2023. "Co-Expression of JcNAC1- and JcZFP8-Improved Agronomic Traits of Tobacco and Enhanced Drought Resistance through NbbHLH1 and NbbHLH2" Plants 12, no. 17: 3029. https://doi.org/10.3390/plants12173029