
Plant Hormone and Inorganic Ion Concentrations in the Xylem Exudate of Grafted Plants Depend on the Scion–Rootstock Combination
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
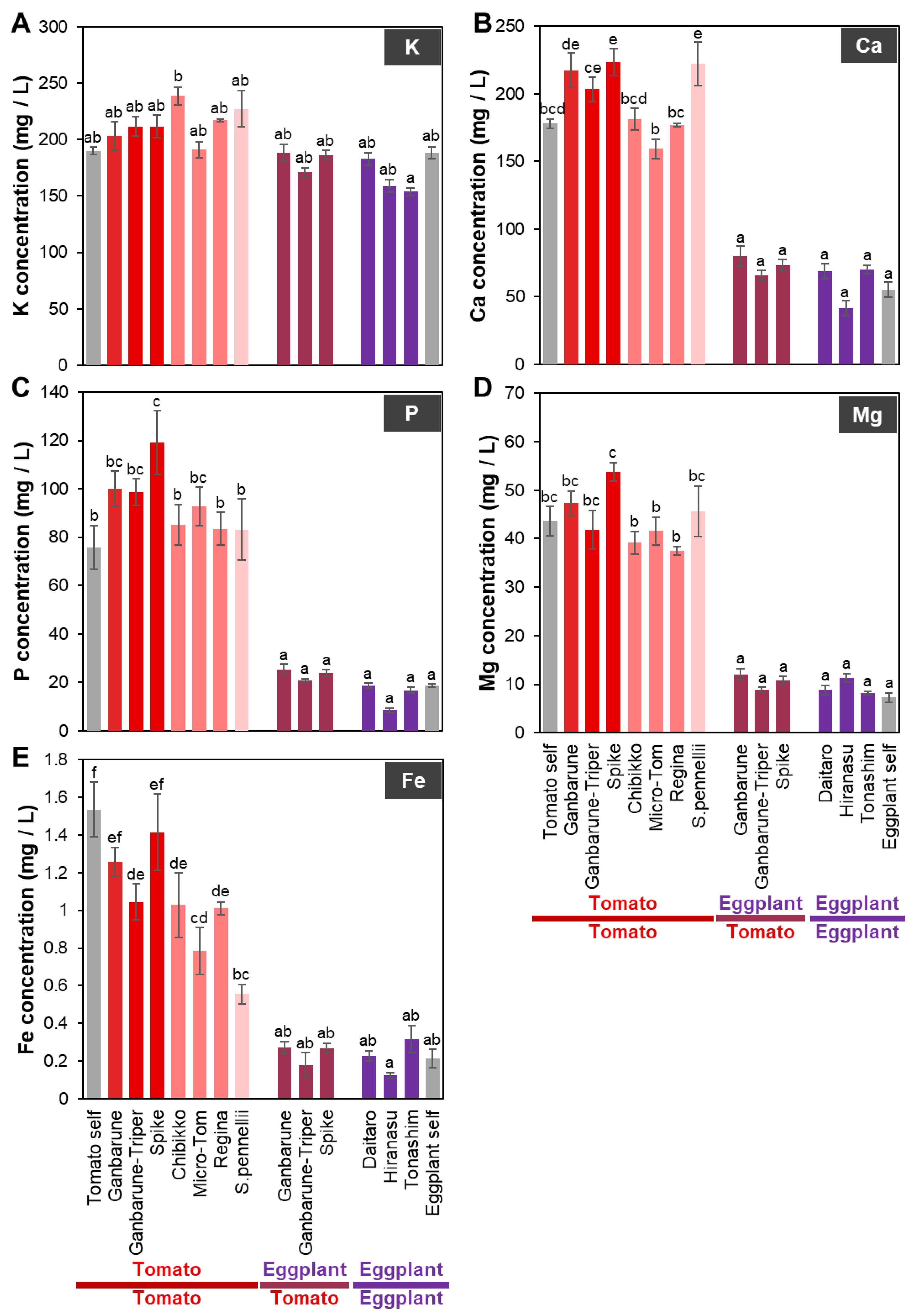
2.1. Inorganic Ion Concentrations in the Xylem Exudate of Tomato and Eggplant Grafted onto Various Rootstocks
2.1.1. Ca Concentration in the Xylem Exudates of Plants Grafted onto Various Rootstocks
2.1.2. P Concentration in the Xylem Exudates of Plants Grafted onto Various Rootstocks
2.1.3. Fe Concentration in the Xylem Exudates of Plants Grafted onto Various Rootstocks
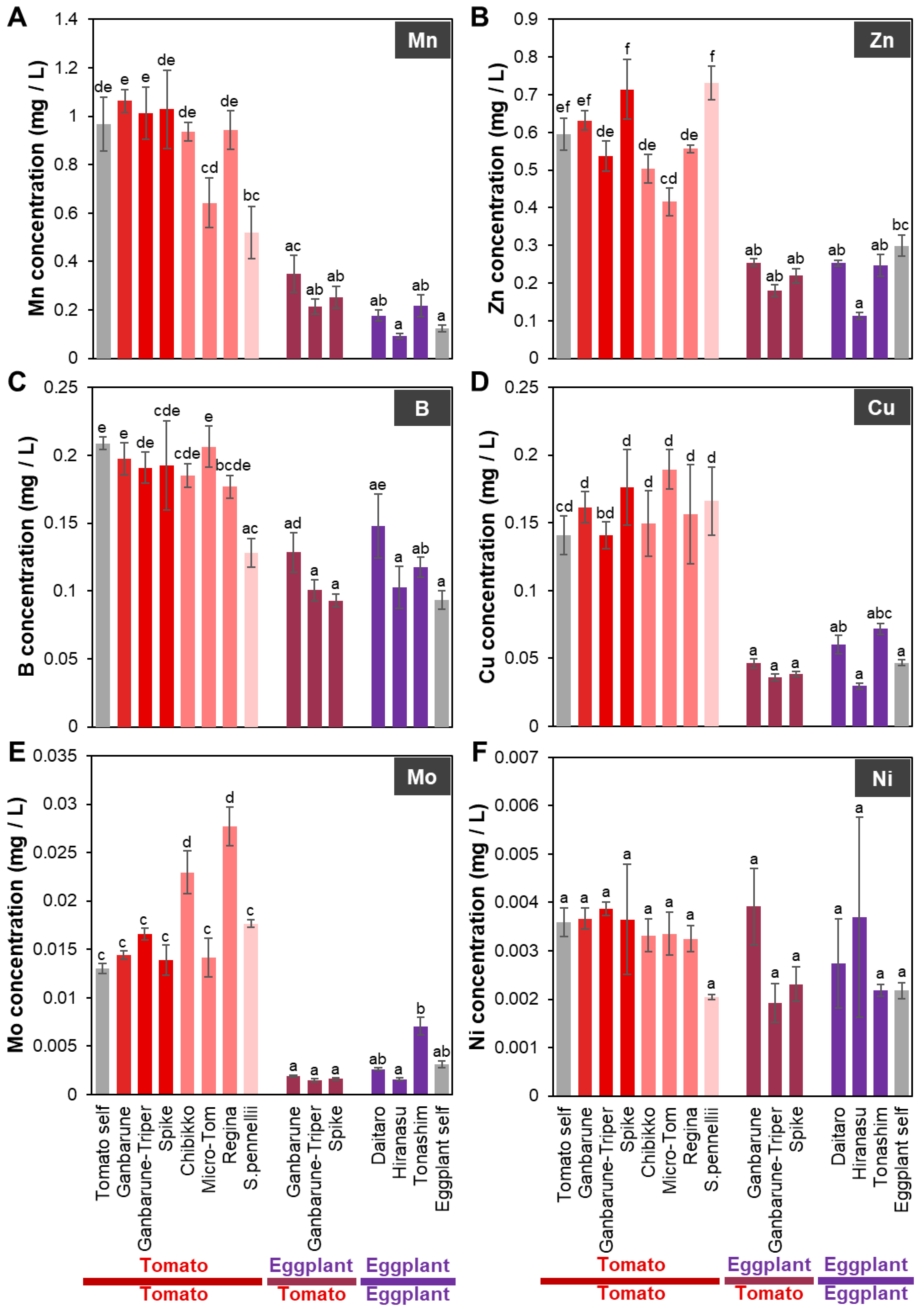
2.1.4. Mn Concentration in the Xylem Exudates of Plants Grafted onto Various Rootstocks
2.1.5. Zn Concentration in the Xylem Exudates of Plants Grafted onto Various Rootstocks
2.1.6. B Concentration in the Xylem Exudates of Plants Grafted onto Various Rootstocks
2.1.7. Mo Concentration in the Xylem Exudates of Plants Grafted onto Various Rootstocks
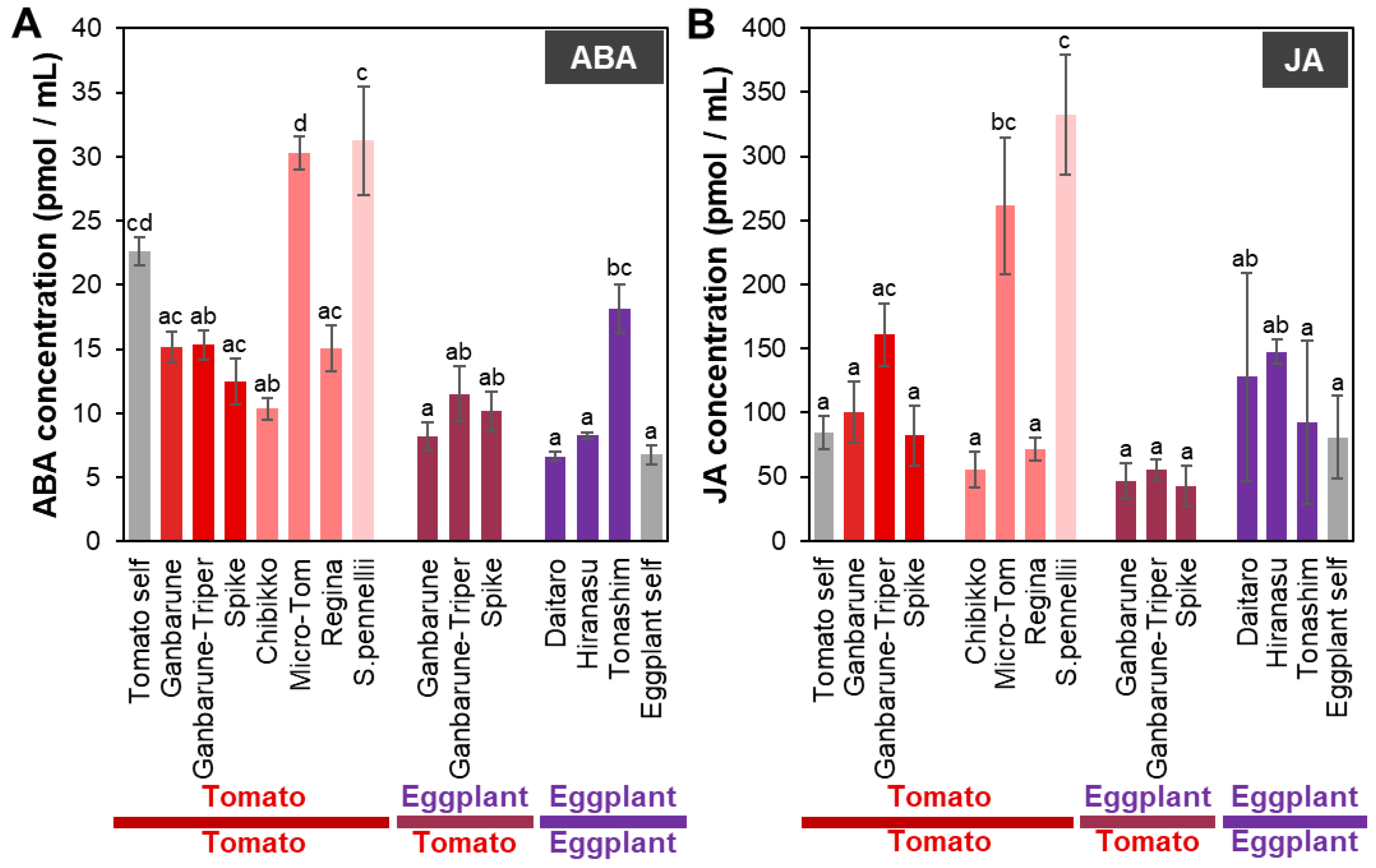
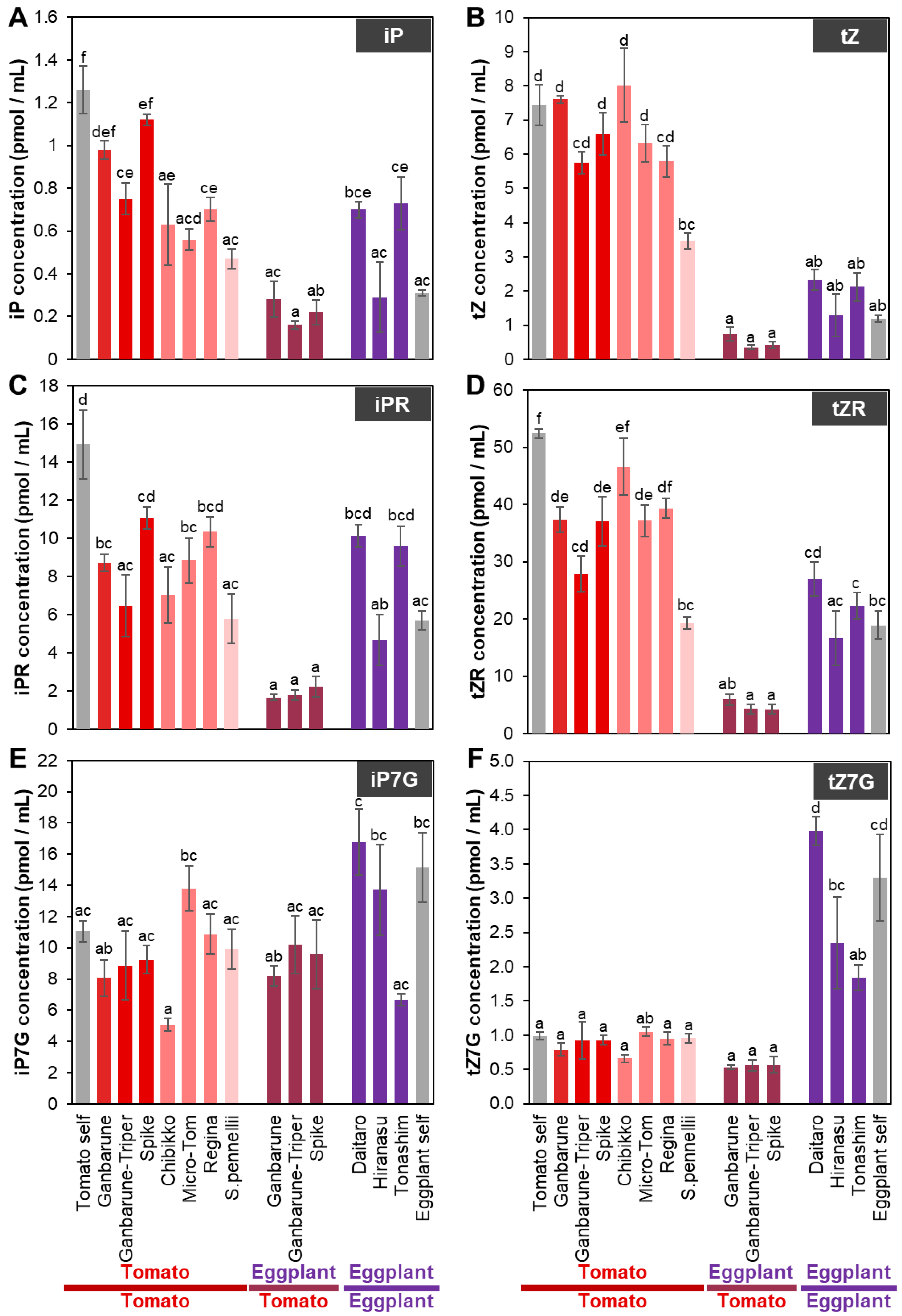
2.2. Plant Hormone Concentrations in the Xylem Exudates of Tomato and Eggplant Plants Grafted on Various Rootstocks
2.2.1. ABA Concentration in the Xylem Exudates of Plants Grafted onto Various Rootstocks
2.2.2. JA Concentration in the Xylem Exudates of Plants Grafted onto Various Rootstocks
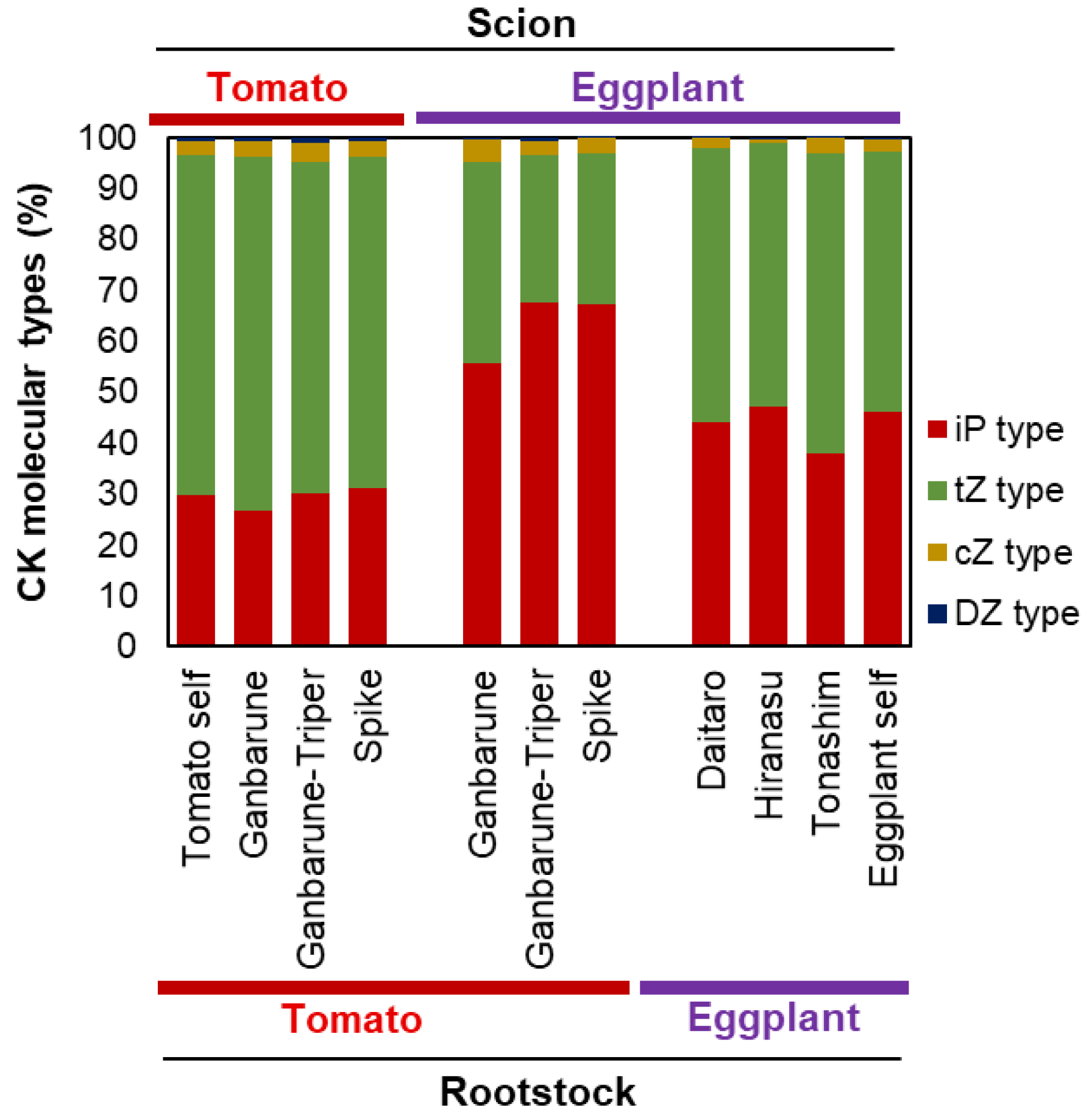
2.2.3. CK Concentrations in the Xylem Exudates of Plants Grafted onto Various Rootstocks
2.3. Scions Alter Inorganic Ion Concentrations and CK Molecular Types Transported from the Rootstock to the Scion
3. Materials and Methods
3.1. Plant Materials and Growth Condition
3.2. Collection of Xylem Exudates from the Main Stem
3.3. Ionome Analysis
3.4. Hormonome Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sattelmacher, B. Tansley review no. 22 The apoplast and its significance for plant mineral nutrition. New Phytol. 2001, 149, 167–192. [Google Scholar] [CrossRef] [PubMed]
- Satoh, S. Organic substances in xylem sap delivered to above-ground organs by the roots. J. Plant Res. 2006, 119, 179–187. [Google Scholar] [CrossRef]
- Yin, Y.G.; Kobayashi, Y.; Sanuki, A.; Kondo, S.; Fukuda, N.; Ezura, H.; Sugaya, S.; Matsukura, C. Salinity induces carbohydrate accumulation and sugar-regulated starch biosynthetic genes in tomato (Solanum lycopersicum L. cv. ‘Micro-Tom’) fruits in an ABA- and osmotic stress- independent manner. J. Exp. Bot. 2010, 61, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Ho, L.C.; Grange, R.I.; Picken, A.J. An analysis of the accumulation of water and dry-matter in tomato fruit. Plant Cell Environ. 1987, 10, 157–162. [Google Scholar] [CrossRef]
- Boawn, L.C.; Rasmussen, P.E. Crop response to excessive zinc fertilization of alkaline soil 1. Agron. J. 1971, 63, 874–876. [Google Scholar] [CrossRef]
- Assuncao, A.G.L.; Herrero, E.; Lin, Y.F.; Huettel, B.; Talukdar, S.; Smaczniak, C.; Immink, R.G.H.; van Eldik, M.; Fiers, M.; Schat, H.; et al. Arabidopsis thaliana transcription factors bZIP19 and bZIP23 regulate the adaptation to zinc deficiency. Proc. Natl. Acad. Sci. USA 2010, 107, 10296–10301. [Google Scholar] [CrossRef] [Green Version]
- Cakmak, I.; Ozkan, H.; Braun, H.J.; Welch, R.M.; Romheld, V. Zinc and iron concentrations in seeds of wild, primitive, and modern wheats. Food Nutr. Bull. 2000, 21, 401–403. [Google Scholar] [CrossRef] [Green Version]
- Yamaji, N.; Xia, J.X.; Mitani-Ueno, N.; Yokosho, K.; Ma, J.F. Preferential delivery of zinc to developing tissues in rice is mediated by P-type heavy metal ATPase OsHMA2. Plant Physiol. 2013, 162, 927–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carroll, M.D.; Loneraga, J.F. Response of plant species to concentrations of zinc in solution. I. growth and zinc content of plants. Aust. J. Agric. Res. 1968, 19, 859–868. [Google Scholar] [CrossRef]
- Foy, C.D.; Chaney, R.L.; White, M.C. The physiology of metal toxicity in plants. Annu. Rev. Plant Physiol. 1978, 29, 511–566. [Google Scholar] [CrossRef]
- Chaney, R.L. Zinc phytotoxicity. Zinc Soils Plants 1993, 55, 135–150. [Google Scholar]
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Elsevier: London, UK, 2012; pp. 1–651. [Google Scholar]
- Russell, E.W. Soil Conditions and Plant Growth, 9th ed.; Longman: London, UK, 1961; pp. 1–849. [Google Scholar]
- Shear, C.B. Calcium-related disorders of fruits and vegetables. Hortscience 1975, 10, 361–365. [Google Scholar] [CrossRef]
- Yoshida, Y.; Irie, N.; Vinh, T.D.; Ooyama, M.; Tanaka, Y.; Yasuba, K.; Goto, T. Incidence of blossom-end rot in relation to the water-soluble calcium concentration in tomato fruits as affected by calcium nutrition and cropping season. J. Jpn. Soc. Hortic. Sci. 2014, 83, 282–289. [Google Scholar] [CrossRef] [Green Version]
- Riefler, M.; Novak, O.; Strnad, M.; Schmulling, T. Arabidopsis cytokinin receptor mutants reveal functions in shoot growth, leaf senescence, seed size, germination, root development, and cytokinin metabolism. Plant Cell 2006, 18, 40–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiba, T.; Takei, K.; Kojima, M.; Sakakibara, H. Side-chain modification of cytokinins controls shoot growth in Arabidopsis. Dev. Cell 2013, 27, 452–461. [Google Scholar] [CrossRef] [Green Version]
- Matsubayashi, Y. Posttranslationally modified small-peptide signals in plants. Annu. Rev. Plant Biol. 2014, 65, 385–413. [Google Scholar] [CrossRef]
- Okamoto, S.; Tabata, R.; Matsubayashi, Y. Long-distance peptide signaling essential for nutrient homeostasis in plants. Curr. Opin. Plant Biol. 2016, 34, 35–40. [Google Scholar] [CrossRef]
- Tabata, R.; Sumida, K.; Yoshii, T.; Ohyama, K.; Shinohara, H.; Matsubayashi, Y. Perception of root-derived peptides by shoot LRR-RKs mediates systemic N-demand signaling. Science 2014, 346, 343–346. [Google Scholar] [CrossRef]
- Ohkubo, Y.; Tanaka, M.; Tabata, R.; Ogawa-Ohnishi, M.; Matsubayashi, Y. Shoot-to-root mobile polypeptides involved in systemic regulation of nitrogen acquisition. Nat. Plants 2017, 3, 17029. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.M.; Oda, M. Grafting of herbaceous vegetable and ornamental crops. Hortic. Rev. 2003, 28, 61–124. [Google Scholar] [CrossRef]
- Lee, J.M.; Kubota, C.; Tsao, S.J.; Bie, Z.; Echevarria, P.H.; Morra, L.; Oda, M. Current status of vegetable grafting: Diffusion, grafting techniques, automation. Sci. Hortic. 2010, 127, 93–105. [Google Scholar] [CrossRef]
- Schwarz, D.; Rouphael, Y.; Colla, G.; Venema, J.H. Grafting as a tool to improve tolerance of vegetables to abiotic stresses: Thermal stress, water stress and organic pollutants. Sci. Hortic. 2010, 127, 162–171. [Google Scholar] [CrossRef]
- Goldschmidt, E.E. Plant grafting: New mechanisms, evolutionary implications. Front. Plant Sci. 2014, 5, 727. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Lucini, L.; RouphaeI, Y.; Cardarelli, M.; Kalunke, R.M.; Colla, G. Insight into the role of grafting and arbuscular mycorrhiza on cadmium stress tolerance in tomato. Front. Plant Sci. 2015, 6, 477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouphael, Y.; Rea, E.; Cardarelli, M.; Bitterlich, M.; Schwarz, D.; Colla, G. Can adverse effects of acidity and aluminum toxicity be alleviated by appropriate rootstock selection in cucumber? Front. Plant Sci. 2016, 7, 1283. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Cardarelli, M.; Colla, G.; Rea, E. Yield, mineral composition, water relations, and water use efficiency of grafted mini-watermelon plants under deficit irrigation. Hortscience 2008, 43, 730–736. [Google Scholar] [CrossRef] [Green Version]
- Edelstein, M.; Plaut, Z.; Ben-Hur, M. Sodium and chloride exclusion and retention by non-grafted and grafted melon and Cucurbita plants. J. Exp. Bot. 2011, 62, 177–184. [Google Scholar] [CrossRef] [Green Version]
- Gioia, D.F.; Aprile, A.; Sabella, E.; Santamaria, P.; Pardossi, A.; Miceli, A.; De Bellis, L.; Nutricati, E. Grafting response to excess boron and expression analysis of genes coding boron transporters in tomato. Plant Biol. 2017, 19, 728–735. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Hayashi, M.; Kanamaru, T.; Watanabe, T.; Mametsuka, S.; Tanaka, Y. Studies on bloom on the surface of cucumber fruits, relation between the degree of bloom occurrence and contents of mineral elements. Bull. Fukuoka Agric. Res. Cent. B9 1989, 9, 1–6. [Google Scholar]
- Mitani, N.; Yamaji, N.; Ago, Y.; Iwasaki, K.; Ma, J.F. Isolation and functional characterization of an influx silicon transporter in two pumpkin cultivars contrasting in silicon accumulation. Plant J. 2011, 66, 231–240. [Google Scholar] [CrossRef]
- Somers, T.C. A connection between potassium levels in the harvest and relative quality in Australian red wines. Aust. Wine Brew. Spirit Rev. 1977, 24, 32–34. [Google Scholar]
- Kodur, S.; Tisdall, J.M.; Tang, C.; Walker, R.R. Accumulation of potassium in grapevine rootstocks (Vitis) grafted to ‘Shiraz’ as affected by growth, root-traits and transpiration. Aust. J. Grape Wine Res. 2010, 49, 7–13. [Google Scholar]
- Niu, M.L.; Sun, S.T.; Nawaz, M.A.; Sun, J.Y.; Cao, H.S.; Lu, J.Y.; Huang, Y.; Bie, Z.L. Grafting cucumber onto pumpkin induced early stomatal closure by increasing aba sensitivity under salinity conditions. Front. Plant Sci. 2019, 10, 1290. [Google Scholar] [CrossRef]
- Tworkoski, T.; Fazio, G. Hormone and growth interactions of scions and size-controlling rootstocks of young apple trees. Plant Growth Regul. 2016, 78, 105–119. [Google Scholar] [CrossRef]
- Van Hooijdonk, B.; Woolley, D.; Warrington, I.; Tustin, S. Rootstocks modify scion architecture, endogenous hormones, and root growth of newly grafted ‘Royal Gala’ apple trees. J. Am. Soc. Hortic. Sci. 2011, 136, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Stevens, G.A.; Westwood, M.N. Fruit-set and cytokinin-like activity in the xylem sap of sweet cherry (Prunus avium) as affected by rootstock. Physiol. Plant. 1984, 61, 464–468. [Google Scholar] [CrossRef]
- Yamasaki, A.; Yamashita, M.; Furuya, S. Mineral concentrations and cytokinin activity in the xylem exudate of grafted watermelons as affected by rootstocks and crop load. J. Jpn. Soc. Hortic. Sci. 1994, 62, 817–826. [Google Scholar] [CrossRef] [Green Version]
- Sorce, C.; Massai, R.; Picciarelli, P.; Lorenzi, R. Hormonal relationships in xylem sap of grafted and ungrafted Prunus rootstocks. Sci. Hortic. 2002, 93, 333–342. [Google Scholar] [CrossRef]
- De Freitas, S.T.; Padda, M.; Wu, Q.Y.; Park, S.; Mitcham, E.J. Dynamic alternations in cellular and molecular components during blossom-end rot development in tomatoes expressing sCAX1, a constitutively active Ca2+/H+ antiporter from Arabidopsis. Plant Physiol. 2011, 156, 844–855. [Google Scholar] [CrossRef] [Green Version]
- Ho, L.C.; White, P.J. A cellular hypothesis for the induction of blossom-end rot in tomato fruit. Ann. Bot. 2005, 95, 571–581. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Arredondo, D.L.; Leyva-Gonzalez, M.A.; Gonzalez-Morales, S.I.; Lopez-Bucio, J.; Herrera-Estrella, L. Phosphate nutrition: Improving low-phosphate tolerance in crops. Annu. Rev. Plant Biol. 2014, 65, 95–123. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.C.; Song, L.; Zhang, Y.; Zheng, Z.; Liu, D. Arabidopsis PHL2 and PHR1 act redundantly as the key components of the central regulatory system controlling transcriptional responses to phosphate starvation. Plant Physiol. 2016, 170, 499–514. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.W.; Qiu, Q.Y.; Chen, Y.H.; Lin, D.B.; Huang, J.Z.; Huang, T.B. Metabolite alteration in response to low phosphorus stress in developing tomato fruits. Plant Physiol. Biochem. 2021, 159, 234–243. [Google Scholar] [CrossRef] [PubMed]
- Cordell, D.; Drangert, J.O.; White, S. The story of phosphorus: Global food security and food for thought. Glob. Environ. Chang.-Hum. Policy Dimens. 2009, 19, 292–305. [Google Scholar] [CrossRef]
- Briat, J.F.; Dubos, C.; Gaymard, F. Iron nutrition, biomass production, and plant product quality. Trends Plant Sci. 2015, 20, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Connorton, J.M.; Balk, J. Iron biofortification of staple crops: Lessons and challenges in plant genetics. Plant Cell Physiol. 2019, 60, 1447–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alejandro, S.; Holler, S.; Meier, B.; Peiter, E. Manganese in plants: From acquisition to subcellular allocation. Front. Plant Sci. 2020, 11, 300. [Google Scholar] [CrossRef] [Green Version]
- Henriques, F.S. Reduction in chloroplast number accounts for the decrease in the photosynthetic capacity of Mn-deficient pecan leaves. Plant Sci. 2004, 166, 1051–1055. [Google Scholar] [CrossRef]
- Hebbern, C.A.; Laursen, K.H.; Ladegaard, A.H.; Schmidt, S.B.; Pedas, P.; Bruhn, D.; Schjoerring, J.K.; Wulfsohn, D.; Husted, S. Latent manganese deficiency increases transpiration in barley (Hordeum vulgare). Physiol. Plant. 2009, 135, 307–316. [Google Scholar] [CrossRef]
- Alejandro, S.; Cailliatte, R.; Alcon, C.; Dirick, L.; Domergue, F.; Correia, D.; Castaings, L.; Briat, J.F.; Mari, S.; Curie, C. Intracellular distribution of manganese by the trans-golgi network transporter NRAMP2 is critical for photosynthesis and cellular redox homeostasis. Plant Cell 2017, 29, 3068–3084. [Google Scholar] [CrossRef] [Green Version]
- Longnecker, N.E.; Graham, R.D.; Card, G. Effects of manganese deficiency on the pattern of tillering and development of barley (Hordeum vulgare c.v. galleon). Field Crop. Res. 1991, 28, 85–102. [Google Scholar] [CrossRef]
- Hebbern, C.A.; Pedas, P.; Schjoerring, J.K.; Knudsen, L.; Husted, S. Genotypic differences in manganese efficiency: Field experiments with winter barley (Hordeum vulgare L.). Plant Soil 2005, 272, 233–244. [Google Scholar] [CrossRef]
- Pedas, P.; Hebbern, C.A.; Schjoerring, J.K.; Holm, P.E.; Husted, S. Differential capacity for high-affinity manganese uptake contributes to differences between barley genotypes in tolerance to low manganese availability. Plant Physiol. 2005, 139, 1411–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millaleo, R.; Reyes-Diaz, M.; Ivanov, A.G.; Mora, M.L.; Alberdi, M. Manganese as essential and toxic element for plants: Transport, accumulation and resistance mechanisms. J. Soil Sci. Plant Nutr. 2010, 10, 476–494. [Google Scholar] [CrossRef] [Green Version]
- Clairmont, K.B.; Hagar, W.G.; Davis, E.A. Manganese toxicity to chlorophyll synthesis in tobacco callus. Plant Physiol. 1986, 80, 291–293. [Google Scholar] [CrossRef] [PubMed]
- Subrahmanyam, D.; Rathore, V.S. Influence of manganese toxicity on photosynthesis in ricebean (Vigna umbellata) seedlings. Photosynthetica 2000, 38, 449–453. [Google Scholar] [CrossRef]
- Nable, R.O.; Houtz, R.L.; Cheniae, G.M. Early inhibition of photosynthesis during development of Mn toxicity in tobacco. Plant Physiol. 1988, 86, 1136–1142. [Google Scholar] [CrossRef] [Green Version]
- Amao, Y.; Ohashi, A. Effect of Mn ion on the visible light induced water oxidation activity of photosynthetic organ grana from spinach. Catal. Commun. 2008, 10, 217–220. [Google Scholar] [CrossRef]
- Zhao, J.J.; Wang, W.Y.; Zhou, H.K.; Wang, R.L.; Zhang, P.; Wang, H.C.; Pan, X.L.; Xu, J. Manganese toxicity inhibited root growth by disrupting auxin biosynthesis and transport in Arabidopsis. Front. Plant Sci. 2017, 8, 272. [Google Scholar] [CrossRef] [Green Version]
- Alam, S.; Akiha, F.; Kamei, S.; Huq, S.M.I.; Kawai, S. Mechanism of potassium alleviation of manganese phytotoxicity in barley. J. Plant Nutr. 2005, 28, 889–901. [Google Scholar] [CrossRef]
- St Clair, S.B.; Lynch, J.P. Element accumulation patterns of deciduous and evergreen tree seedlings on acid soils: Implications for sensitivity to manganese toxicity. Tree Physiol. 2005, 25, 85–92. [Google Scholar] [CrossRef] [Green Version]
- Blamey, F.P.C.; Hernandez-Soriano, M.C.; Cheng, M.M.; Tang, C.X.; Paterson, D.J.; Lombi, E.; Wang, W.H.; Scheckel, K.G.; Kopittke, P.M. Synchrotron-based techniques shed light on mechanisms of plant sensitivity and tolerance to high manganese in the root environment. Plant Physiol. 2015, 169, 2006–2020. [Google Scholar] [CrossRef] [Green Version]
- Leskova, A.; Giehl, R.F.H.; Hartmann, A.; Fargasova, A.; von Wiren, N. Heavy metals induce iron deficiency responses at different hierarchic and regulatory levels. Plant Physiol. 2017, 174, 1648–1668. [Google Scholar] [CrossRef] [Green Version]
- Funakawa, H.; Miwa, K. Synthesis of borate cross-linked rhamnogalacturonan II. Front. Plant Sci. 2015, 6, 223. [Google Scholar] [CrossRef] [Green Version]
- Dell, B.; Huang, L.B. Physiological response of plants to low boron. Plant Soil 1997, 193, 103–120. [Google Scholar] [CrossRef]
- Reid, R. Update on boron toxicity and tolerance in plants. Adv. Plant Anim. Boron Nutr. 2007, 1, 83–90. [Google Scholar] [CrossRef]
- Nable, R.O.; Cartwright, B.; Lance, R.C.M. Genotypic differences in boron accumulation in barley—Relative susceptibilities to boron deficiency and toxicity. Genet. Asp. Plant Miner. Nutr. 1990, 42, 243–251. [Google Scholar]
- Takano, J.; Wada, M.; Ludewig, U.; Schaaf, G.; von Wiren, N.; Fujiwara, T. The Arabidopsis major intrinsic protein NIP5;1 is essential for efficient boron uptake and plant development under boron limitation. Plant Cell 2006, 18, 1498–1509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takano, J.; Noguchi, K.; Yasumori, M.; Kobayashi, M.; Gajdos, Z.; Miwa, K.; Hayashi, H.; Yoneyama, T.; Fujiwara, T. Arabidopsis boron transporter for xylem loading. Nature 2002, 420, 337–340. [Google Scholar] [CrossRef]
- Tejada-Jimenez, M.; Chamizo-Ampudia, A.; Galvan, A.; Fernandez, E.; Llamas, A. Molybdenum metabolism in plants. Metallomics 2013, 5, 1191–1203. [Google Scholar] [CrossRef]
- Mendel, R.R.; Leimkuhler, S. The biosynthesis of the molybdenum cofactors. J. Biol. Inorg. Chem. 2015, 20, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Colla, G.; Suarez, C.M.C.; Cardarelli, M.; Rouphael, Y. Improving nitrogen use efficiency in melon by grafting. Hortscience 2010, 45, 559–565. [Google Scholar] [CrossRef] [Green Version]
- Edelstein, M.; Ben-Hur, M.; Cohen, R.; Burger, Y.; Ravina, I. Boron and salinity effects on grafted and non-grafted melon plants. Plant Soil 2005, 269, 273–284. [Google Scholar] [CrossRef]
- Arao, T.; Takeda, H.; Nishihara, E. Reduction of cadmium translocation from roots to shoots in eggplant (Solanum melongena) by grafting onto Solanum torvum rootstock. Soil Sci. Plant Nutr. 2008, 54, 555–559. [Google Scholar] [CrossRef] [Green Version]
- Savvas, D.; Papastavrou, D.; Ntatsi, G.; Ropokis, A.; Olympios, C.; Hartmann, H.; Schwarz, D. Interactive effects of grafting and manganese supply on growth, yield, and nutrient uptake by tomato. Hortscience 2009, 44, 1978–1982. [Google Scholar] [CrossRef] [Green Version]
- Kodur, S.; Tisdall, J.M.; Tang, C.; Walker, R.R. Accumulation of potassium in grapevine rootstocks (Vitis) as affected by dry matter partitioning, root traits and transpiration. Aust. J. Grape Wine Res. 2010, 16, 273–282. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1755-0238.2009.00088.x (accessed on 8 September 2022).
- Ng, L.M.; Melcher, K.; Teh, B.T.; Xu, H.E. Abscisic acid perception and signaling: Structural mechanisms and applications. Acta Pharmacol. Sin. 2014, 35, 567–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puertolas, J.; Alcobendas, R.; Alarcon, J.J.; Dodd, I.C. Long-distance abscisic acid signalling under different vertical soil moisture gradients depends on bulk root water potential and average soil water content in the root zone. Plant Cell Environ. 2013, 36, 1465–1475. [Google Scholar] [CrossRef]
- Correia, B.; Pinto-Marijuan, M.; Castro, B.B.; Brossa, R.; Lopez-Carbonell, M.; Pinto, G. Hormonal dynamics during recovery from drought in two Eucalyptus globulus genotypes: From root to leaf. Plant Physiol. Biochem. 2014, 82, 151–160. [Google Scholar] [CrossRef]
- Dodd, I.C.; Theobald, J.C.; Richer, S.K.; Davies, W.J. Partial phenotypic reversion of ABA-deficient flacca tomato (Solanum lycopersicum) scions by a wild-type rootstock: Normalizing shoot ethylene relations promotes leaf area but does not diminish whole plant transpiration rate. J. Exp. Bot. 2009, 60, 4029–4039. [Google Scholar] [CrossRef]
- Li, B.B.; Feng, Z.G.; Xie, M.; Sun, M.Z.; Zhao, Y.X.; Liang, L.Y.; Liu, G.J.; Zhang, J.H.; Jia, W.S. Modulation of the root-sourced ABA signal along its way to the shoot in Vitis riparia x Vitis labrusca under water deficit. J. Exp. Bot. 2011, 62, 1731–1741. [Google Scholar] [CrossRef]
- Wang, Y.S.; Liu, F.L.; Jensen, C.R. Comparative effects of deficit irrigation and alternate partial root-zone irrigation on xylem pH, ABA and ionic concentrations in tomatoes. J. Exp. Bot. 2012, 63, 1907–1917. [Google Scholar] [CrossRef] [PubMed]
- Patane, C.; Cosentino, S.L. Effects of soil water deficit on yield and quality of processing tomato under a Mediterranean climate. Agric. Water Manag. 2010, 97, 131–138. [Google Scholar] [CrossRef]
- Favati, F.; Lovelli, S.; Galgano, F.; Miccolis, V.; Di Tommaso, T.; Candido, V. Processing tomato quality as affected by irrigation scheduling. Sci. Hortic. 2009, 122, 562–571. [Google Scholar] [CrossRef]
- Gundlach, H.; Muller, M.J.; Kutchan, T.M.; Zenk, M.H. Jasmonic acid is a signal transducer in elicitor-induced plant-cell cultures. Proc. Natl. Acad. Sci. USA 1992, 89, 2389–2393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomma, B.; Eggermont, K.; Penninckx, I.; Mauch-Mani, B.; Vogelsang, R.; Cammue, B.P.A.; Broekaert, W.F. Separate jasmonate-dependent and salicylate-dependent defense-response pathways in Arabidopsis are essential for resistance to distinct microbial pathogens. Proc. Natl. Acad. Sci. USA 1998, 95, 15107–15111. [Google Scholar] [CrossRef] [Green Version]
- Wasternack, C.; Hause, B. Jasmonates: Biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 review in Annals of Botany. Ann. Bot. 2013, 111, 1021–1058. [Google Scholar] [CrossRef]
- Anderson, J.P.; Badruzsaufari, E.; Schenk, P.M.; Manners, J.M.; Desmond, O.J.; Ehlert, C.; Maclean, D.J.; Ebert, P.R.; Kazan, K. Antagonistic interaction between abscisic acid and jasmonate-ethylene signaling pathways modulates defense gene expression and disease resistance in Arabidopsis. Plant Cell 2004, 16, 3460–3479. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Davis, L.C.; Verpoorte, R. Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv. 2005, 23, 283–333. [Google Scholar] [CrossRef]
- Bandurska, H.; Stroinski, A.; Kubis, J. The effect of jasmonic acid on the accumulation of ABA, proline and spermidine and its influence on membrane injury under water deficit in two barley genotypes. Acta Physiol. Plant. 2003, 25, 279–285. [Google Scholar] [CrossRef]
- Kim, S.K.; Sohn, E.Y.; Joo, G.J.; Lee, I.J. Influence of jasmonic acid on endogenous gibberellin and abscisic acid in salt-stressed chard plant. J. Environ. Biol. 2009, 30, 333–338. [Google Scholar] [PubMed]
- De Ollas, C.; Hernando, B.; Arbona, V.; Gomez-Cadenas, A. Jasmonic acid transient accumulation is needed for abscisic acid increase in citrus roots under drought stress conditions. Physiol. Plant. 2013, 147, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Fragoso, V.; Rothe, E.; Baldwin, I.T.; Kim, S.G. Root jasmonic acid synthesis and perception regulate folivore-induced shoot metabolites and increase Nicotiana attenuata resistance. New Phytol. 2014, 202, 1335–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harb, A.; Krishnan, A.; Ambavaram, M.M.R.; Pereira, A. Molecular and physiological analysis of drought stress in Arabidopsis reveals early responses leading to acclimation in plant growth. Plant Physiol. 2010, 154, 1254–1271. [Google Scholar] [CrossRef] [Green Version]
- Grebner, W.; Stingl, N.E.; Oenel, A.; Mueller, M.J.; Berger, S. Lipoxygenase6-dependent oxylipin synthesis in roots is required for abiotic and biotic stress resistance of Arabidopsis. Plant Physiol. 2013, 161, 2159–2170. [Google Scholar] [CrossRef] [Green Version]
- Riemann, M.; Dhakarey, R.; Hazman, M.; Miro, B.; Kohli, A.; Nick, P. Exploring jasmonates in the hormonal network of drought and salinity responses. Front. Plant Sci. 2015, 6, 1077. [Google Scholar] [CrossRef] [Green Version]
- Sakakibara, H. Cytokinins: Activity, biosynthesis, and translocation. Annu. Rev. Plant Biol. 2006, 57, 431–449. [Google Scholar] [CrossRef] [Green Version]
- Beveridge, C.A.; Murfet, I.C.; Kerhoas, L.; Sotta, B.; Miginiac, E.; Rameau, C. The shoot controls zeatin riboside export from pea roots. Evidence from the branching mutant rms4. Plant J. 1997, 11, 339–345. [Google Scholar] [CrossRef]
- Hirose, N.; Takei, K.; Kuroha, T.; Kamada-Nobusada, T.; Hayashi, H.; Sakakibara, H. Regulation of cytokinin biosynthesis, compartmentalization and translocation. J. Exp. Bot. 2008, 59, 75–83. [Google Scholar] [CrossRef] [Green Version]
- Bishopp, A.; Lehesranta, S.; Vaten, A.; Help, H.; El-Showk, S.; Scheres, B.; Helariutta, K.; Mahonen, A.P.; Sakakibara, H.; Helariutta, Y. Phloem-transported cytokinin regulates polar auxin transport and maintains vascular pattern in the root meristem. Curr. Biol. 2011, 21, 927–932. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, T.; Suzaki, T.; Soyano, T.; Kojima, M.; Sakakibara, H.; Kawaguchi, M. Shoot-derived cytokinins systemically regulate root nodulation. Nat. Commun. 2014, 5, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mok, D.W.S.; Mok, M.C. Cytokinin metabolism and action. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 89–118. [Google Scholar] [CrossRef]
- Werner, T.; Schmulling, T. Cytokinin action in plant development. Curr. Opin. Plant Biol. 2009, 12, 527–538. [Google Scholar] [CrossRef] [PubMed]
- Kieber, J.J.; Schaller, G.E. Cytokinin signaling in plant development. Development 2018, 145, dev149344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurakawa, T.; Ueda, N.; Maekawa, M.; Kobayashi, K.; Kojima, M.; Nagato, Y.; Sakakibara, H.; Kyozuka, J. Direct control of shoot meristem activity by a cytokinin-activating enzyme. Nature 2007, 445, 652–655. [Google Scholar] [CrossRef]
- Tokunaga, H.; Kojima, M.; Kuroha, T.; Ishida, T.; Sugimoto, K.; Kiba, T.; Sakakibara, H. Arabidopsis lonely guy (LOG) multiple mutants reveal a central role of the LOG-dependent pathway in cytokinin activation. Plant J. 2012, 69, 355–365. [Google Scholar] [CrossRef]
- Ko, D.; Kang, J.; Kiba, T.; Park, J.; Kojima, M.; Do, J.; Kim, K.Y.; Kwon, M.; Endler, A.; Song, W.Y.; et al. Arabidopsis ABCG14 is essential for the root-to-shoot translocation of cytokinin. Proc. Natl. Acad. Sci. USA 2014, 111, 7150–7155. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.W.; Novak, O.R.; Wei, Z.Y.; Gou, M.Y.; Zhang, X.B.; Yu, Y.; Yang, H.J.; Cai, Y.H.; Strnad, M.; Liu, C.J. Arabidopsis ABCG14 protein controls the acropetal translocation of root-synthesized cytokinins. Nat. Commun. 2014, 5, 3274. [Google Scholar] [CrossRef] [Green Version]
- Beveridge, C.A.; Symons, G.M.; Murfet, I.C.; Ross, J.J.; Rameau, C. The rms1 mutant of pea has elevated indole-3-acetic acid levels and reduced root-sap zeatin riboside content but increased branching controlled by graft-transmissible signal(s). Plant Physiol. 1997, 115, 1251–1258. [Google Scholar] [CrossRef] [Green Version]
- Che, J.; Yokosho, K.; Yamaji, N.; Ma, J.F. A vacuolar phytosiderophore transporter alters iron and zinc accumulation in polished rice grains. Plant Physiol. 2019, 181, 276–288. [Google Scholar] [CrossRef] [Green Version]
- Kojima, M.; Kamada-Nobusada, T.; Komatsu, H.; Takei, K.; Kuroha, T.; Mizutani, M.; Ashikari, M.; Ueguchi-Tanaka, M.; Matsuoka, M.; Suzuki, K.; et al. Highly sensitive and high-throughput analysis of plant hormones using MS-probe modification and liquid chromatography-tandem mass spectrometry: An application for hormone profiling in Oryza sativa. Plant Cell Physiol. 2009, 50, 1201–1214. [Google Scholar] [CrossRef] [PubMed]
- Kojima, M.; Sakakibara, H. Highly sensitive high-throughput profiling of six phytohormones using MS-probe modification and liquid chromatography-tandem mass spectrometry. Methods Mol. Biol. 2012, 918, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Shinozaki, Y.; Hao, S.; Kojima, M.; Sakakibara, H.; Ozeki-Iida, Y.; Zheng, Y.; Fei, Z.; Zhong, S.; Giovannoni, J.J.; Rose, J.K.; et al. Ethylene suppresses tomato (Solanum lycopersicum) fruit set through modification of gibberellin metabolism. Plant J. 2015, 83, 237–251. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kawaguchi, K.; Nakaune, M.; Ma, J.F.; Kojima, M.; Takebayashi, Y.; Sakakibara, H.; Otagaki, S.; Matsumoto, S.; Shiratake, K. Plant Hormone and Inorganic Ion Concentrations in the Xylem Exudate of Grafted Plants Depend on the Scion–Rootstock Combination. Plants 2022, 11, 2594. https://doi.org/10.3390/plants11192594
Kawaguchi K, Nakaune M, Ma JF, Kojima M, Takebayashi Y, Sakakibara H, Otagaki S, Matsumoto S, Shiratake K. Plant Hormone and Inorganic Ion Concentrations in the Xylem Exudate of Grafted Plants Depend on the Scion–Rootstock Combination. Plants. 2022; 11(19):2594. https://doi.org/10.3390/plants11192594
Chicago/Turabian StyleKawaguchi, Kohei, Makoto Nakaune, Jian Feng Ma, Mikiko Kojima, Yumiko Takebayashi, Hitoshi Sakakibara, Shungo Otagaki, Shogo Matsumoto, and Katsuhiro Shiratake. 2022. "Plant Hormone and Inorganic Ion Concentrations in the Xylem Exudate of Grafted Plants Depend on the Scion–Rootstock Combination" Plants 11, no. 19: 2594. https://doi.org/10.3390/plants11192594