
Changes in Antioxidative Compounds and Enzymes in Small-Leaved Linden (Tilia cordata Mill.) in Response to Mistletoe (Viscum album L.) Infestation





Abstract
:1. Introduction
2. Results
2.1. Effect of Mistletoe Infestation on the Relative Water Content and the Content of Chlorophylls in Linden Leaves
2.2. Effect of Mistletoe Infestation on Oxidative Stress Parameters in Linden Leaves
2.3. Effect of Mistletoe Infestation on Non-Enzymatic Antioxidants in Linden Leaves
2.4. Effect of Mistletoe Infestation on Antioxidative Enzyme Activity in Linden Leaves
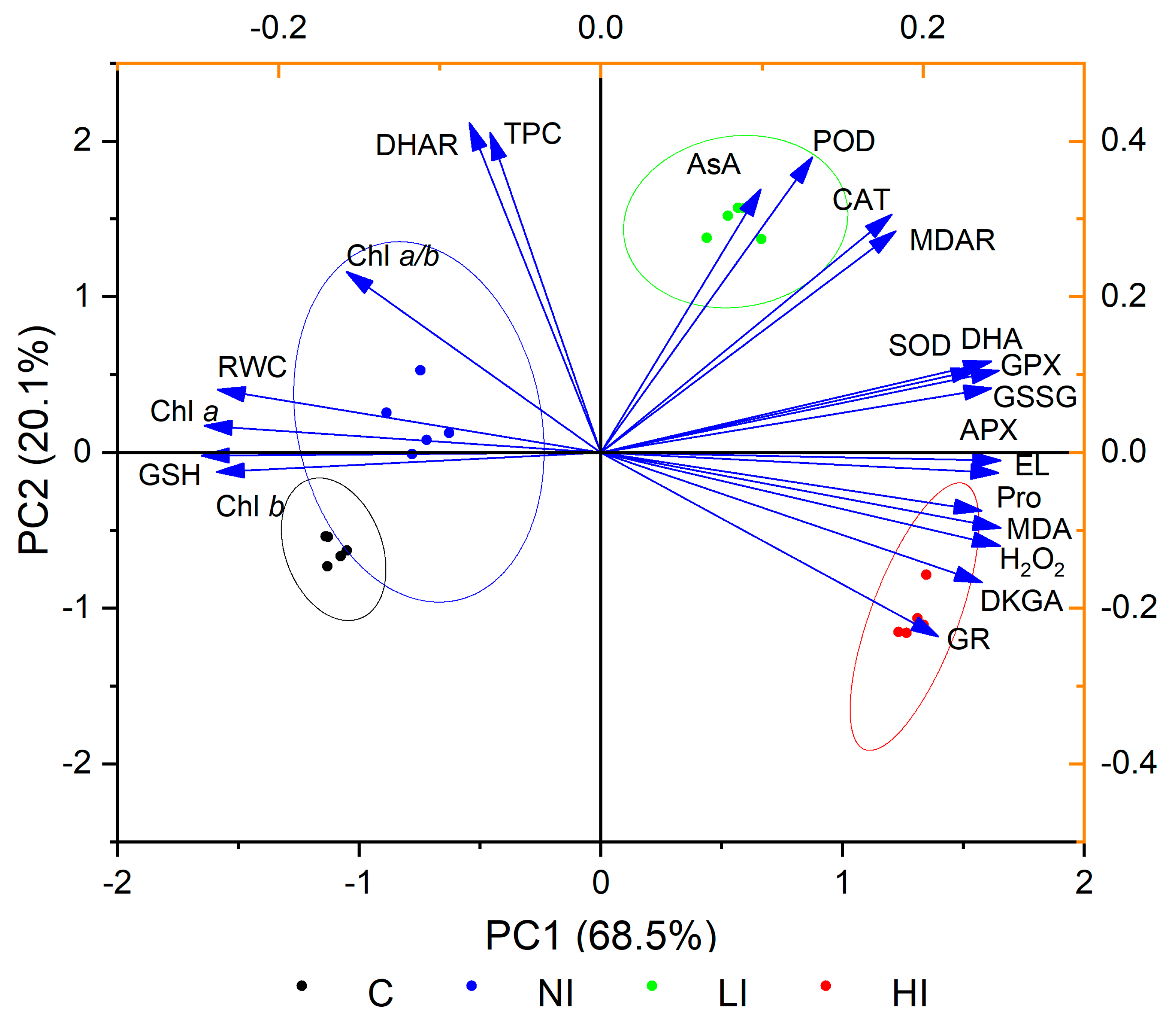
2.5. Interaction between Relative Water Content, Chlorophyll Content, Oxidative Stress, and Antioxidative Response of Linden Trees to Mistletoe Infestation
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Determination of Relative Water Content
4.3. Determination of Chlorophyll Content
4.4. Determination of Oxidative Stress Parameters
4.5. Determination of Non-Enzymatic Antioxidant Content
4.6. Determination of Antioxidative Enzyme Activity
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Varga, I.; Poczai, P.; Tiborcz, V.; Aranyi, N.R.; Baltazár, T.; Bartha, D.; Pejchal, M.; Hyvönen, J. Changes in the distribution of European mistletoe (Viscum album) in Hungary during the last hundred years. Folia Geobot. Phytotaxon. 2014, 49, 559–577. [Google Scholar] [CrossRef]
- Rigling, A.; Eilmann, B.; Koechli, R.; Dobbertin, M. Mistletoe-induced crown degradation in scots pine in a xeric environment. Tree Physiol. 2010, 30, 845–852. [Google Scholar] [CrossRef] [Green Version]
- Mutlu, S.; Ilhan, V.; Turkoglu, H.I. Mistletoe (Viscum album) infestation in the scots pine stimulates drought-dependent oxidative damage in summer. Tree Physiol. 2016, 36, 479–489. [Google Scholar] [CrossRef] [Green Version]
- Sangüesa-Barreda, G.; Linares, J.C.; Camarero, J. Mistletoe effects on scots pine decline following drought events: Insights from within-tree spatial patterns, growth and carbohydrates. Tree Physiol. 2012, 32, 585–598. [Google Scholar] [CrossRef] [Green Version]
- Scalon, M.C.; Rossatto, D.R.; Franco, A.C. How does mistletoe infection affect seasonal physiological responses of hosts with different leaf phenology? Flora Morphol. Distrib. Funct. Ecol. Plants 2021, 281, 151871. [Google Scholar] [CrossRef]
- Díaz-Limón, M.P.; Cano-Santana, Z.; Queijeiro-Bolaños, M.E. Mistletoe infection in an urban forest in Mexico City. Urban For. Urban Gree. 2016, 17, 126–134. [Google Scholar] [CrossRef]
- Dmuchowski, W.; Brągoszewska, P.; Gozdowski, D.; Baczewska-Dąbrowska, A.H.; Chojnacki, T.; Jozwiak, A.; Swiezewska, E.; Suwara, I.; Gworek, B. Strategies of urban trees for mitigating salt stress: A case study of eight plant species. Trees 2020, 1–16. [Google Scholar] [CrossRef]
- Skrypnik, L.; Maslennikov, P.; Feduraev, P.; Pungin, A.; Belov, N. Ecological and landscape factors affecting the spread of European mistletoe (Viscum album L.) in urban areas (A Case Study of the Kaliningrad City, Russia). Plants 2020, 9, 394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Jaegere, T.; Hein, S.; Claessens, H. A review of the characteristics of small-leaved lime (Tilia cordata Mill.) and their implications for silviculture in a changing climate. Forests 2016, 7, 56. [Google Scholar] [CrossRef] [Green Version]
- Urošević, M.A.; Jovanović, G.; Stević, N.; Deljanin, I.; Nikolić, M.; Tomašević, M.; Samson, R. Leaves of common urban tree species (Aesculus hippocastanum, Acer platanoides, Betula pendula and Tilia cordata) as a measure of particle and particle-bound pollution: A 4-year study. Air Qual. Atmos. Health 2019, 12, 1081–1090. [Google Scholar] [CrossRef]
- Šilinskas, B.; Povilaitienė, A.; Urbaitis, G.; Aleinikovas, M.; Varnagirytė-Kabašinskienė, I. The wood quality of small-leaved lime (Tilia cordata Mill.) trees in an urban area: A pilot study. Forests 2021, 12, 420. [Google Scholar] [CrossRef]
- Martynova, M.; Sultanova, R.; Odintsov, G.; Sazgutdinova, R.; Khanova, E. Growth of Tilia cordata Mill. in urban forests. South East Eur. For. 2020, 11, 51–59. [Google Scholar] [CrossRef]
- Fishman, M.R.; Shirasu, K. How to resist parasitic plants: Pre- and post-attachment strategies. Curr. Opin. Plant Biol. 2021, 62, 102004. [Google Scholar] [CrossRef] [PubMed]
- Albert, M.; Axtell, M.J.; Timko, M.P. Mechanisms of resistance and virulence in parasitic plant–host interactions. Plant Physiol. 2021, 185, 1282–1291. [Google Scholar] [CrossRef]
- Hegenauer, V.; Körner, M.; Albert, M. Plants under stress by parasitic plants. Curr. Opin. Plant Biol. 2017, 38, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Cui, S.; Ichihashi, Y.; Shirasu, K. The haustorium, a specialized invasive organ in parasitic plants. Annu. Rev. Plant Biol. 2016, 67, 643–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czarnocka, W.; Karpiński, S. Friend or foe? Reactive oxygen species production, scavenging and signaling in plant response to environmental stresses. Free Radic. Biol. Med. 2018, 122, 4–20. [Google Scholar] [CrossRef]
- Rajput, V.D.; Harish; Singh, R.; Verma, K.; Sharma, L.; Quiroz-Figueroa, F.; Meena, M.; Gour, V.; Minkina, T.; Sushkova, S.; et al. Recent developments in enzymatic antioxidant defence mechanism in plants with special reference to abiotic stress. Biology 2021, 10, 267. [Google Scholar] [CrossRef]
- Biswas, K.; Adhikari, S.; Tarafdar, A.; Kumar, R.; Saha, S.; Ghosh, P. Reactive oxygen species and antioxidant defence systems in plants: Role and crosstalk under biotic stress. In Sustainable Agriculture in the Era of Climate Change; Roychowdhury, R., Choudhury, S., Hasanuzzaman, M., Srivastava, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2020; pp. 265–292. [Google Scholar]
- Per, T.S.; Khan, N.A.; Reddy, P.S.; Masood, A.; Hasanuzzaman, M.; Khan, M.I.R.; Anjum, N.A. Approaches in modulating proline metabolism in plants for salt and drought stress tolerance: Phytohormones, mineral nutrients and transgenics. Plant Physiol. Biochem. 2017, 115, 126–140. [Google Scholar] [CrossRef]
- El Moukhtari, A.; Cabassa-Hourton, C.; Farissi, M.; Savouré, A. How does proline treatment promote salt stress tolerance during crop plant development? Front. Plant Sci. 2020, 11, 1127. [Google Scholar] [CrossRef]
- Ben Rejeb, K.; Abdelly, C.; Savouré, A. How reactive oxygen species and proline face stress together. Plant Physiol. Biochem. 2014, 80, 278–284. [Google Scholar] [CrossRef]
- Anselmo-Moreira, F.; Teixeira-Costa, L.; Ceccantini, G.; Furlan, C.M. Mistletoe effects on the host tree Tapirira guianensis: Insights from primary and secondary metabolites. Chemoecology 2019, 29, 11–24. [Google Scholar] [CrossRef]
- Patykowski, J.; Kołodziejek, J. Comparative analysis of antioxidant activity in leaves of different hosts infected by mistletoe (Viscum album L. subsp. album). Arch. Biol. Sci. 2013, 65, 851–861. [Google Scholar] [CrossRef]
- Hu, B.; Sakakibara, H.; Takebayashi, Y.; Peters, F.S.; Schumacher, J.; Eiblmeier, M.; Arab, L.; Kreuzwieser, J.; Polle, A.; Ren-nenberg, H. Mistletoe infestation mediates alteration of the phytohormone profile and anti-oxidative metabolism in bark and wood of its host Pinus sylvestris. Tree Physiol. 2017, 37, 676–691. [Google Scholar] [CrossRef]
- Skrypnik, L.; Maslennikov, P.; Feduraev, P.; Pungin, A.; Belov, N. Specific features of the response of the antioxidant system of urban trees to mistletoe infection. In E3S Web of Conferences, IV International Scientific and Practical Conference “Sustainable Development and Green Growth on the Innovation Management Platform” (SDGG 2021), Kaliningrad, Russia, 27–28 May 2021; EDP Sciences: Les Ulis, France, 2021; Volume 291, p. 02013. [Google Scholar]
- Kubov, M.; Fleischer, P., Jr.; Rozkošný, J.; Kurjak, D.; Konôpková, A.; Galko, J.; Húdoková, H.; Lalík, M.; Rell, S.; Pittner, J.; et al. Drought or severe drought? Hemiparasitic yellow mistletoe (Loranthus europaeus) amplifies drought stress in sessile oak trees (Quercus petraea) by altering water status and physiological responses. Water 2020, 12, 2985. [Google Scholar] [CrossRef]
- Escher, P.; Peuke, A.D.; Bannister, P.; Fink, S.; Hartung, W.; Jiang, F.; Rennenberg, H. Transpiration, CO2 assimilation, WUE, and stomatal aperture in leaves of Viscum album (L.): Effect of abscisic acid (ABA) in the xylem sap of its host (Populus x euamericana). Plant Physiol. Biochem. 2008, 46, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Nemeskéri, E.; Helyes, L. Physiological responses of selected vegetable crop species to water stress. Agronomy 2019, 9, 447. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, B.; Vogg, G.; Fürst, U.B.; Albert, M. Parasitic plants of the genus Cuscuta and their interaction with susceptible and resistant host plants. Front. Plant Sci. 2015, 6, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, G.; Asthir, B. Molecular responses to drought stress in plants. Biol. Plant. 2017, 61, 201–209. [Google Scholar] [CrossRef]
- Nieves-Cordones, M.; De la Calle, M.L.; Ródenas, R.; Martínez, V.; Rubio, F.; Rivero, R.M. Critical responses to nutrient deprivation: A comprehensive review on the role of ROS and RNS. Environ. Exp. Bot. 2019, 161, 74–85. [Google Scholar] [CrossRef]
- Zechmann, B. Subcellular roles of glutathione in mediating plant defense during biotic stress. Plants 2020, 9, 1067. [Google Scholar] [CrossRef]
- Boubakri, H. The role of ascorbic acid in plant–pathogen interactions. In Ascorbic Acid in Plant Growth, Development and Stress Tolerance; Hossain, M.A., Munné-Bosch, S., Burritt, D.J., Diaz-Vivancos, P., Fujita, M., Lorence, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 255–271. [Google Scholar]
- Kumar, S.; Abedin, M.M.; Singh, A.K.; Das, S. Role of phenolic compounds in plant-defensive mechanisms. In Plant Phenolics in Sustainable Agriculture; Lone, R., Shuab, R., Kamili, A.N., Eds.; Springer: Singapore, 2020; Volume 1, pp. 517–532. [Google Scholar]
- Furlan, C.M.; Moreira, F.A.; Teixeira-Costa, L.; Ceccantini, G.; Salminen, J.-P. Does Phoradendron perrottetii (mistletoe) alter polyphenols levels of Tapirira guianensis (host plant)? Plant Physiol. Biochem. 2019, 136, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Maslennikov, P.V.; Chupakhina, G.N.; Skrypnik, L.N.; Feduraev, P.V.; Melnik, A.S. The contribution of polyphenols to plant resistance to Pb soil pollution. Int. J. Environ. Stud. 2018, 75, 719–731. [Google Scholar] [CrossRef]
- Naderi, S.; Fakheri, B.-A.; Amiri, R.M.; Mahdinezhad, N. Tolerance responses in wheat landrace Bolani are related to enhanced metabolic adjustments under drought stress. Plant Physiol. Biochem. 2020, 150, 244–253. [Google Scholar] [CrossRef] [PubMed]
- André, C.M.; Schafleitner, R.; Legay, S.; Lefèvre, I.; Aliaga, C.A.A.; Nomberto, G.; Hoffmann, L.; Hausman, J.-F.; Larondelle, Y.; Evers, D. Gene expression changes related to the production of phenolic compounds in potato tubers grown under drought stress. Phytochemistry 2009, 70, 1107–1116. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Dumanović, J.; Nepovimova, E.; Natić, M.; Kuča, K.; Jaćević, V. The significance of reactive oxygen species and antioxidant defense system in plants: A concise overview. Front. Plant Sci. 2021, 11, 552969. [Google Scholar] [CrossRef] [PubMed]
- Shawon, R.A.; Kang, B.S.; Lee, S.G.; Kim, S.K.; Lee, H.J.; Katrich, E.; Gorinstein, S.; Ku, Y.G. Influence of drought stress on bioactive compounds, antioxidant enzymes and glucosinolate contents of Chinese cabbage (Brassica rapa). Food Chem. 2020, 308, 125657. [Google Scholar] [CrossRef]
- Nadgórska-Socha, A.; Kandziora-Ciupa, M.; Ciepał, R.; Barczyk, G. Robinia pseudoacacia and Melandrium album in trace elements biomonitoring and air pollution tolerance index study. Int. J. Environ. Sci. Technol. 2016, 13, 1741–1752. [Google Scholar] [CrossRef] [Green Version]
- Sumanta, N.; Haque, C.I.; Nishika, J.; Suprakash, R. Spectrophotometric analysis of chlorophylls and carotenoids from com-monly grown fern species by using various extracting solvents. Res. J. Chem. Sci. 2014, 4, 63–69. [Google Scholar]
- Ahmad, P.; Ahanger, M.A.; Alyemeni, M.N.; Wijaya, L.; Alam, P. Exogenous application of nitric oxide modulates osmolyte metabolism, antioxidants, enzymes of ascorbate-glutathione cycle and promotes growth under cadmium stress in tomato. Protoplasma 2018, 255, 79–93. [Google Scholar] [CrossRef]
- Wang, Y.; Ding, M.; Gu, X.; Wang, J.; Pang, Y.; Gao, L.; Xia, T. Retracted: Analysis of interfering substances in the measurement of malondialdehyde content in plant leaves. Am. J. Biochem. Biotechnol. 2013, 9, 235–242. [Google Scholar] [CrossRef]
- Junglee, S.; Urban, L.; Sallanon, H.; Lopez-Lauri, F. Optimized assay for hydrogen peroxide determination in plant tissue using potassium iodide. Am. J. Anal. Chem. 2014, 5, 730–736. [Google Scholar] [CrossRef] [Green Version]
- Dash, S.P.; Dixit, S.; Sahoo, S. Phytochemical and biochemical characterizations from leaf extracts from Azadirachta Indica: An important medicinal plant. Biochem. Anal. Biochem. 2017, 6. [Google Scholar] [CrossRef]
- Lwalaba, J.L.W.; Louis, L.T.; Zvobgo, G.; Richmond, M.E.A.; Fu, L.; Naz, S.; Mwamba, M.; Mundende, R.P.M.; Zhang, G. Physiological and molecular mechanisms of cobalt and copper interaction in causing phyto-toxicity to two barley genotypes differing in Co tolerance. Ecotoxicol. Environ. Saf. 2020, 187, 109866. [Google Scholar] [CrossRef] [PubMed]
- Maslennikov, P.; Golovina, E.; Artemenko, A. Ecological and geochemical conditions for the accumulation of antioxidants in the leaves of Lathyrus maritimus (L.) Bigel. Plants 2020, 9, 746. [Google Scholar] [CrossRef] [PubMed]
- Usman, K.; Abu-Dieyeh, M.H.; Zouari, N.; Al-Ghouti, M.A. Lead (Pb) bioaccumulation and antioxidative responses in Tetraena qataranse. Sci. Rep. 2020, 10, 17070. [Google Scholar] [CrossRef]
- Wang, C.-Q.; Xu, H.-J.; Liu, T. Effect of selenium on ascorbate–glutathione metabolism during peg-induced water deficit in Trifolium repens L. J. Plant Growth Regul. 2011, 30, 436–444. [Google Scholar] [CrossRef]
- Sairam, R.K.; Rao, K.; Srivastava, G. Differential response of wheat genotypes to long term salinity stress in relation to oxidative stress, antioxidant activity and osmolyte concentration. Plant Sci. 2002, 163, 1037–1046. [Google Scholar] [CrossRef]
- Aebi, H. Catalase In Vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Hartikainen, H.; Xue, T.; Piironen, V. Selenium as an anti-oxidant and pro-oxidant in ryegrass. Plant Soil 2000, 225, 193–200. [Google Scholar] [CrossRef]
- Kandziora-Ciupa, M.; Nadgórska-Socha, A.; Barczyk, G.; Ciepał, R. Bioaccumulation of heavy metals and ecophysiological responses to heavy metal stress in selected populations of Vaccinium myrtillus L. and Vaccinium vitis-idaea L. Ecotoxicology 2017, 26, 966–980. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Peres-Neto, P.R.; Jackson, D.A.; Somers, K.M. Giving meaningful interpretation to ordination axes: Assessing loading significance in principal component analysis. Ecology 2003, 84, 2347–2363. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Treatment | RWC, % | Chlorophyll a, mg g−1 DW | Chlorophyll b, mg g−1 DW | Total Chlorophyll, mg g−1 DW | Chlorophyll a/b Ratio |
|---|---|---|---|---|---|
| C | 89.2 ± 0.7 a | 2.25 ± 0.03 a | 1.56 ± 0.02 a | 3.80 ± 0.04 a | 1.44 ± 0.01 a |
| NI | 89.1 ± 1.0 a | 2.52 ± 0.04 a | 1.51 ± 0.05 a | 3.75 ± 0.05 a | 1.51 ± 0.03 a |
| LI | 83.6 ± 0.5 b | 1.79 ± 0.06 b | 1.23 ± 0.03 b | 3.03 ± 0.07 b | 1.45 ± 0.02 a |
| HI | 77.8 ± 0.9 c | 1.52 ± 0.05 c | 1.14 ± 0.03 b | 2.66 ± 0.06 c | 1.33 ± 0.03 b |
| Treatment | Electrolyte Leakage, % | MDA, nmol g−1 DW | H2O2, nmol g−1 DW |
|---|---|---|---|
| C | 12.4 ± 0.7 c | 93.6 ± 2.0 c | 2.24 ± 0.03 a |
| NI | 15.8 ± 0.6 c | 99.4 ± 3.1 c | 2.25 ± 0.03 a |
| LI | 23.8 ± 1.3 b | 137.8 ± 4.4 b | 1.79 ± 0.05 b |
| HI | 30.8 ± 1.4 a | 183.8 ± 3.6 a | 1.52 ± 0.6 c |
| Treatment | Proline, µmol g−1 DW | GSH, µmol g−1 DW | GSSG, µmol g−1 DW | AsA, µg g−1 DW | DHA, µg g−1 DW | DKGA, µg g−1 DW | TPC, mg g−1 DW |
|---|---|---|---|---|---|---|---|
| C | 155.2 ± 5.1 c | 53.8 ± 1.6 a | 4.10 ± 0.11 b | 111.6 ± 3.0 c | 44.2 ± 1.4 c | 36.8 ± 2.0 c | 9.48 ± 0.39 b |
| NI | 219.0 ± 5.5 b | 45.6 ± 1.2 b | 4.12 ± 0.14 b | 218.6 ± 5.8 a | 63.8 ± 2.7 b | 38.4 ± 1.5 c | 9.20 ± 0.41 b |
| LI | 239.8 ±5.4 b | 32.8 ± 1.4 c | 5.70 ± 0.10 a | 230.6 ± 3.7 a | 79.4 ± 1.5 a | 61.4 ± 2.3 b | 12.92 ± 0.86 a |
| HI | 321.4 ± 4.7 a | 23.0 ± 1.5 d | 5.84 ± 0.12 a | 178.2 ± 5.2 b | 84.6 ± 1.5 a | 110.1 ± 4.2 a | 5.44 ± 0.29 c |
| Treatment | SOD, U mg−1 Protein | CAT, nmol H2O2 mg−1 Protein min−1 | APX, µmol AsA mg−1 Protein min−1 | MDAR, µmol NADPH mg−1 Protein min−1 | DHAR, µmol AsA mg−1 Protein min−1 | GPX, µmol GSH mg−1 Protein min−1 | GR, µmol NADPH mg−1 Protein min−1 | POD, µmol Guaiacol mg−1 Protein min−1 |
|---|---|---|---|---|---|---|---|---|
| C | 4.46 ± 0.05 c | 131.1 ± 2.9 c | 14.1 ± 1.0 d | 3.47 ± 0.06 c | 1.96 ± 0.03 b | 1.33 ± 0.03 d | 2.22 ± 0.04 b | 0.58 ± 0.04 c |
| NI | 4.44 ± 0.04 c | 134.2 ± 2.8 c | 24.1 ± 1.3 c | 3.49 ± 0.04 c | 2.77 ± 0.06 a | 2.18 ± 0.03 c | 2.21 ± 0.03 b | 0.68 ± 0.01 bc |
| LI | 7.57 ± 0.06 a | 227.8 ± 3.9 a | 36.6 ± 1.4 b | 4.50 ± 0.05 a | 2.87 ± 0.04 a | 3.30 ± 0.05 b | 2.29 ± 0.02 b | 1.48 ± 0.05 a |
| HI | 7.46 ± 0.11 b | 174.0 ± 2.7 b | 48.5 ± 1.8 a | 3.97 ± 0.04 b | 1.49 ± 0.05 c | 3.53 ±0.04 a | 2.68 ± 0.04 a | 0.80 ± 0.02 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skrypnik, L.; Maslennikov, P.; Feduraev, P.; Pungin, A.; Belov, N. Changes in Antioxidative Compounds and Enzymes in Small-Leaved Linden (Tilia cordata Mill.) in Response to Mistletoe (Viscum album L.) Infestation. Plants 2021, 10, 1871. https://doi.org/10.3390/plants10091871
Skrypnik L, Maslennikov P, Feduraev P, Pungin A, Belov N. Changes in Antioxidative Compounds and Enzymes in Small-Leaved Linden (Tilia cordata Mill.) in Response to Mistletoe (Viscum album L.) Infestation. Plants. 2021; 10(9):1871. https://doi.org/10.3390/plants10091871
Chicago/Turabian StyleSkrypnik, Liubov, Pavel Maslennikov, Pavel Feduraev, Artem Pungin, and Nikolay Belov. 2021. "Changes in Antioxidative Compounds and Enzymes in Small-Leaved Linden (Tilia cordata Mill.) in Response to Mistletoe (Viscum album L.) Infestation" Plants 10, no. 9: 1871. https://doi.org/10.3390/plants10091871