Physiological Responses of Selected Vegetable Crop Species to Water Stress


Institute of Horticulture, Szent István University, H-2100 Gödöllő, Hungary
*
Author to whom correspondence should be addressed.
Agronomy 2019, 9(8), 447; https://doi.org/10.3390/agronomy9080447
Submission received: 21 July 2019
/
Revised: 6 August 2019
/
Accepted: 11 August 2019
/
Published: 13 August 2019
(This article belongs to the Special Issue Physiological and Molecular Characterization of Crop Resistance to Abiotic Stresses)
Abstract
:The frequency of drought periods influences the yield potential of crops under field conditions. The change in morphology and anatomy of plants has been tested during drought stress under controlled conditions but the change in physiological processes has not been adequately studied in separate studies but needs to be reviewed collectively. This review presents the responses of green peas, snap beans, tomatoes and sweet corn to water stress based on their stomatal behaviour, canopy temperature, chlorophyll fluorescence and the chlorophyll content of leaves. These stress markers can be used for screening the drought tolerance of genotypes, the irrigation schedules or prediction of yield.
1. Introduction
As a result of climate change, the increasing atmospheric CO2 enhances the photosynthesis capacity and improves water use efficiency therefore the amount of yield will increase in most of vegetable crops, however its advantage cannot be shown under limited water and nitrogen deficiency. The high temperature during reproductive growth is harmful for many important vegetable crops, such as tomatoes, peppers, beans and sweet corn, and yield reduction will probably occur [1]. The frequency of drought periods decreases vegetable yield and quality, however soluble solid content of produce may be increased by water deficiency in some crops [2,3]. Nevertheless, the occurrence of excess precipitation causes waterlogging in soils, the symptoms of which are similar to water deficit. Soil waterlogging impedes the oxygen supply and respiration of roots, water uptake and hydraulic conductance which results in stomatal closure [4,5]. Under these conditions the stomatal closure results in a reduction of net photosynthesis which is due to the decrease in stomatal conductance, chlorophyll fluorescence and chlorophyll content of leaves [6]. Excess water causes a decline in grain filling and grain weight of corn leading to decreased yield [7]. However, water stress commonly refers to water deficits not excess water.
The selection of the vegetable crops grown under field conditions for the investigation was based on their production in the world and Europe. During the last twenty years from 1997 to 2017 the growing area of tomatoes increased intensively, that of green peas increased moderately while the growing area of snap beans and sweet corn increased slowly in the world. During this time in Europe the growing area of tomatoes and snap beans decreased from 650.4 to 496.2 thousand ha while that of green peas increased slightly from 208 to 212 thousand ha and sweet corn’s increased intensively (from 50.5 to 110.1 thousand ha) (FAOSTAT 2017 [8]. In Hungary, the production of green peas and sweet corn is performed in large field growing areas (19.5 thousand hectares and 34.5 thousand hectares, respectively) while snap beans are grown in smaller ones (1.6 thousand hectares) (FAOSTAT 2017) [8]. The other aspect of the selection was the sensitivity of plant species to water stress.
Corn, soybeans, beans and peas are considered to be moderately water stress sensitive while tomatoes belong to the extremely drought sensitive group [9,10]. The responses of plant species significantly depend on the intensity and duration of stress and their stages of development. The spring-sown green pea utilizes the precipitation well (if there is any) and requires a low temperature during vegetative growth but during the flowering and seed development periods it is sensitive to water deficiency. The crops require a warm temperature, even though they have different ripening times, snap beans have short (60 days), sweet corn has medium (75–90 days) and tomatoes have long ripening times (110–130 days), their generative stages of development coincided with dry June and July, thus they require irrigation. Irrigation scheduling and the amount of irrigation water are determined by the water stress tolerance and water use of the plant varieties. The evaluation of drought tolerance in field conditions is difficult because low soil moisture and high air temperature stress generally occur together, and it is difficult to evaluate the responses separately. Drought under field conditions promotes the evapotranspiration and affects the photosynthesis, which leads to reduced yield [11]. Use of remote-sensing methods makes the measurement of physiological responses of varieties to various strong water stresses easy. These non-destructive methods help the breeder to select drought tolerant genotypes and the growers to measure the water deficit of plants and decide the time of irrigation.
The selection for water stress tolerance in traditional breeding is based on the suitability of performance under a series of environmental conditions using extensive statistical methods. This progress could be improved by the introduction of traits which contribute to the prediction of yield in the drought-prone environments. In this study, the effect of water stress on the plants and those physiological traits which influenced the yield are mainly demonstrated. Information was gathered on the physiological responses of selected vegetable species to drought to analyse their use in breeding for high and stable yield.
2. Water Stress during Growth of Vegetable Crops
Sensitivity of plants to water stress such as snap beans and green peas differs with the stages of development. During the early stages of vegetative growth most crops are less sensitive to water scarcity [2,3,12,13], but during the generative stage the water deficiency results in changes of many physiological traits [2,14,15,16], causing the disturbance of fertility and reduction of yield. During flowering of legume plants water stress increases the ratio of flower drop [17], decreases the pod numbers and seed abortion in the pods [18,19] and increases the ratio of curved pods [20]. Under water deficiency, bean plants produce shorter shoots and smaller leaves and decrease the length of pods [21]. Semi-leafless pea varieties have reduced leaf area that is presumed to have a low water use and they have higher water use efficiency (WUE) than traditional varieties with normal leaves [22]. In sweet corn, ear differentiation begins at the six- or eight-leaf stage growth when the water deficiency decreases the length of ears and the numbers of ear rows [23], but during tasselling the water deficiency causes significant yield reduction [24,25]. Tomatoes are most sensitive to water deficiency at fruit setting and intensive fruit development periods [3], when the increasing water stress resulted in a 25 to 50% decrease in the yield [10,26,27,28,29]. During early flowering of tomatoes, water scarcity causes flower shedding and lack of fertilization [30], and during fruit setting, plants with small sized fruits are produced [10,31].
The effect of water stress on morphology and anatomy of plants has been studied by several researchers under controlled conditions [6,32,33,34,35], however, the changes in physiological responses have been less investigated under field conditions. The physiological characteristics that have an important role in the defence against drought can be measured by remote sensing techniques using non-destructive methods in open field conditions. The leaf photosynthetic activity of plants can be monitored with measurement of chlorophyll content using a portable chlorophyll meter and chlorophyll fluorescence while the measurement of stomatal conductance indicates the severity of water stress [2,3,14]. Spectral vegetation indices such as the green normalized vegetation index (GNDVI) and the normalized differential vegetation index (NDVI) were used for monitoring the growth of the plant to detect the water stress and for yield prediction [36,37,38,39]. Crop water stress index is determined by an infrared thermometry technique to indicate the change in canopy temperature of plants under water stress conditions. More physiological indices such as leaf water potential, relative water content, turgor potential, osmotic adjustment, difference between canopy and air temperature can also be used as a screening tools for testing the water stress tolerance of genotypes [40]. Studies have focused on the identification of drought tolerance-related traits using Quantitative Trait Locus (QTL)s and Marker Assisted Selection (MAS) techniques [41,42,43,44], however, the identification of the most relevant loci controlling drought tolerance and drought-related traits could be achieved by the integration of molecular genetics with physiology [45].
3. Drought Tolerance
Adaptive mechanisms promoting the survival of plants have been grouped into three categories; drought escape, drought avoidance and drought tolerance [46]. Drought escape is the ability of plants to accomplish their life cycle before the development of soil and plant water deficit. The varieties with early flowering and short maturity are able to escape drought [47], however, they are not drought tolerant in every case. The varieties with moderate drought sensitivity developed different defence strategies to avoid short- and long-term water stress which prevents the water loss in their cells and tissues. The essential defence mechanism against drought operating in the plants is the maintenance of the water status and the reduction of tissue water loss (Figure 1).
A well-developed deeply penetrating root system provides the water uptake and maintenance of water circulation inside the plant despite the low soil-moisture content. Nevertheless, in dry soil the lives of microorganisms are retarded when the activity of mycorrhiza living symbiotically with root nodules of legumes is low, which results in a decrease in the nitrogen uptake [48,49], therefore the growth of the plant is retarded. Long-term drought of soil accelerates the senescence of root nodules and production of reactive oxygen species (ROS) [50,51], therefore the nodule weight, root and shoot weight are decreased [52]. Water stress results in a change in the proportion of root weight as the ratio of root to shoot increases [53]. Under permanent low moisture content of soil, a 27–42% decrease in leaf weight and 12–27% decrease in specific leaf area of snap bean varieties were found [54]. Tomatoes are able to survive prolonged periods of low soil water content by the development of a deep root system [28,55]. In dry years, tomatoes inoculated with mycorrhiza easily endured the water scarcity, for example larger weight fruits and higher yield were produced by deficit irrigation than under non-irrigated conditions [56].
4. Reduction of Water Loss
Drought avoidance is the ability of plants to maintain high tissue water potential despite the deficiency of soil moisture. The mechanisms developed for the reduction of water loss are related to the duration of water stress.
During short-term water deficiency the leaf movement, deep penetrating roots with strong suction force and partial or total stomatal closure provide a decrease in the water loss. Leaf movement of plants not only protects from the photodamage caused by high irradiation but reduces the effective leaf area for transpiration [57]. Paraheliotropic movement of leaves occurs mainly in beans while leaf rolling is typical for maize. Fernandez and Castrillo [58] found that the extent of leaf rolling is linearly correlated with the water potential. During leaf rolling of maize the transpiration, stomatal conductance, intracellular CO2 concentration and net photosynthetic rate decreased [59]. Pastenes et al. [60] found that the degree of paraheliotropic leaf movement was larger in the water stressed plants because of lower water potential, however, it also occurred in the water supplied crops. Deep, thick and dense roots intensively promote the use of available water and the optimal development of aboveground parts. During short-term water stress (<7 nap), abscisic acid (ABA) is produced in the roots then it is transported into the leaves where ABA induces the stomatal closure and thus decreases the water loss [61,62]. Partial or total stomatal closure restricts the transpiration therefore the water and nutrient uptake is decreased, which results in a decrease in photosynthesis and growth of plants [63]. Stomatal responses of legume species are different; under water deficiency, beans have a rapid and complete stomatal closure generating the stomatal conductivity and photosynthesis decreases significantly, whereas in cowpeas (Vigna unguiculata), the stomata remain partially open and have a lower decrease in their net photosynthetic rate under the same conditions [64]. Under moderate water deficit conditions, the growth of snap beans was already retarded, and their leaf area decreased while the leaf area index (LAI) of sweet corn did not change [3,15]. Nevertheless, water scarcity did not influence the leaf area of tomatoes [65] but heat and water stress up to 6 days already significantly decreased the weight of shoots and roots of tomato seedlings under a controlled environment [66].
During long-term water deficiency, plants try to prevent the dehydration of cells of vegetative and generative organs with some morphological and physiological changes. Trichome density (leaf hairs) on the leaf protects the tissues from sunlight injury, decreases the water loss by evaporation and enhances the transpiration resistance [67]. Under water stress conditions, a lower number of trichomes was found only on the basal zone of leaves on both surfaces in comparison with irrigated plants [68]. However, according to Nobel [69], the length of trichomes can be more important than their frequency. The epicuticular wax layer of the leaf controls the water flow across the cuticle and protects from high radiation and prevents damage caused by UV light. Water stress induced the increase in the wax layer on the leaf surface of peas and the wax-rich varieties had significantly lower canopy temperature [70].
Drought tolerance is the ability of the plants to endure the long-term moisture deficit and survive the water loss. When the morphological changes seem to be insufficient to avoid the water deficiency, biochemical processes of plants are activated to maintain the osmotic adjustment and the structure of cell membranes in order to avoid cell dehydration. Decreasing the water potential of leaves induces the accumulation of different osmotic compounds such as sugars, amino acids and quaternary ammonium compounds. The osmotic pressure of cells is increased by the accumulation of osmotic compounds because water movement into the cells and tissues provides the maintenance of turgor [71]. It was found that peas and castor beans exposed to water deficit accumulate a significant amount of soluble sugars and proline [72,73], and the raffinose and sucrose level of leaves are significantly increased by water stress during flowering of snap beans [74]. Action of enzymatic and non-enzymatic antioxidants is intensified to alleviate the oxidative damages in the tissues. Concerted operation of numerous water soluble antioxidant compounds (ACW) contributes to the adaptation of plants to environmental stresses. The level of ACW in the leaves is influenced by stomatal closure because it is related to ascorbic acid redox potential of guard cells [75]. In snap bean genotypes that have a high ACW level in leaves during the flowering and pod development periods, this provides a defence against water deficiency [74].
4.1. Regulation of Water Circulation in Plants under Drought
Many physiological processes are activated to mitigate the water loss of plants (Table 1). Transpiration is restrained as a result of stomatal closure and by decreasing leaf area. Stomata play an important role in the regulation of transpiration and CO2 uptake. Use of light energy gathered by photosynthesis determines the growth and biomass production of plants. In these processes, the stomatal characteristics such as stomatal size, number and ratio of stomata on abaxial and adaxial surfaces significantly affect the C assimilation and water use efficiency (WUE) [76,77]. The higher stomatal density on the abaxial surface of the leaf is related to a higher water use efficiency [78], while those existing on upper epidermis (adaxial surface) of the leaf influenced the water use of plants [15]. Nevertheless, the number of stomata on both epidermis of leaves changes significantly depending on the variety and water supply.
4.1.1. Stomatal Characteristics
More stomata (134–195 stomata mm−2) were observed on the abaxial surface of tomato leaves but it was significantly less (40–62 mm−2) on the adaxial surface of leaves [76]. A significant difference can be demonstrated in stomatal density of leaves between snap beans, green peas and sweet corn grown under non-irrigated and deficit irrigated (50% water deficiency) conditions (Table 2). On the basis of 3 year experiments, on the lower epidermis of leaves the stomata density was significantly higher for snap beans under moderate and severe water stress and it was similarly high for sweet corn only in severe water deficiency, but no difference could be shown for green peas in comparison with the optimal water supplied plants [14,15,88]. On the upper epidermis of leaves more and larger sized stomata can be found for snap beans exposed to drought while there were fewer similar sized stomata for the green peas compared to the irrigated plants (Table 2). However, under water scarcity, significant differences in stomata number and size can be detected between the varieties. Under non-irrigated conditions, the size of stomata on the upper (adaxial) surface of leaves of green-podded bean varieties was smaller by 5–12%, but more of them were found than on optimal water supplied plants. Nevertheless, yellow-podded snap bean varieties had 13–18% larger sized stomata on the adaxial surface of leaves of plants exposed to water deficiency in comparison with the irrigated plants [15]. A larger stomatal density was observed for late ripening green pea varieties [14] and late ripening sweet corn hybrids under water scarcity [88] than for the short duration ones. Nevertheless, the distribution and size of stomata can be different on both areas and surfaces of the same leaf. Various number and sized stomata were detected on different areas of leaves of tomatoes; on the abaxial surface of leaves and their apical and middle areas, larger sized (32–34 µm) and more stomata were found than that on the adaxial surface. The stomata on the apical areas of leaves responded sensitively to water deficiency in that they showed fewer and larger sized stomata on the adaxial surface of leaves than for well-watered plants [68]. A significant correlation between the stomatal density and stomatal conductance (r2 = 0.958) was established in tomatoes. According to this correlation, 130 stomata mm−2 was associated with high stomatal conductance (0.1 mol H2O m−2 s−1) [76]. Others [89] found that the relationship between stomatal density and WUE was positive and the size of stomata correlated negatively with the WUE for grass peas.
4.1.2. Canopy Temperature-Transpiration
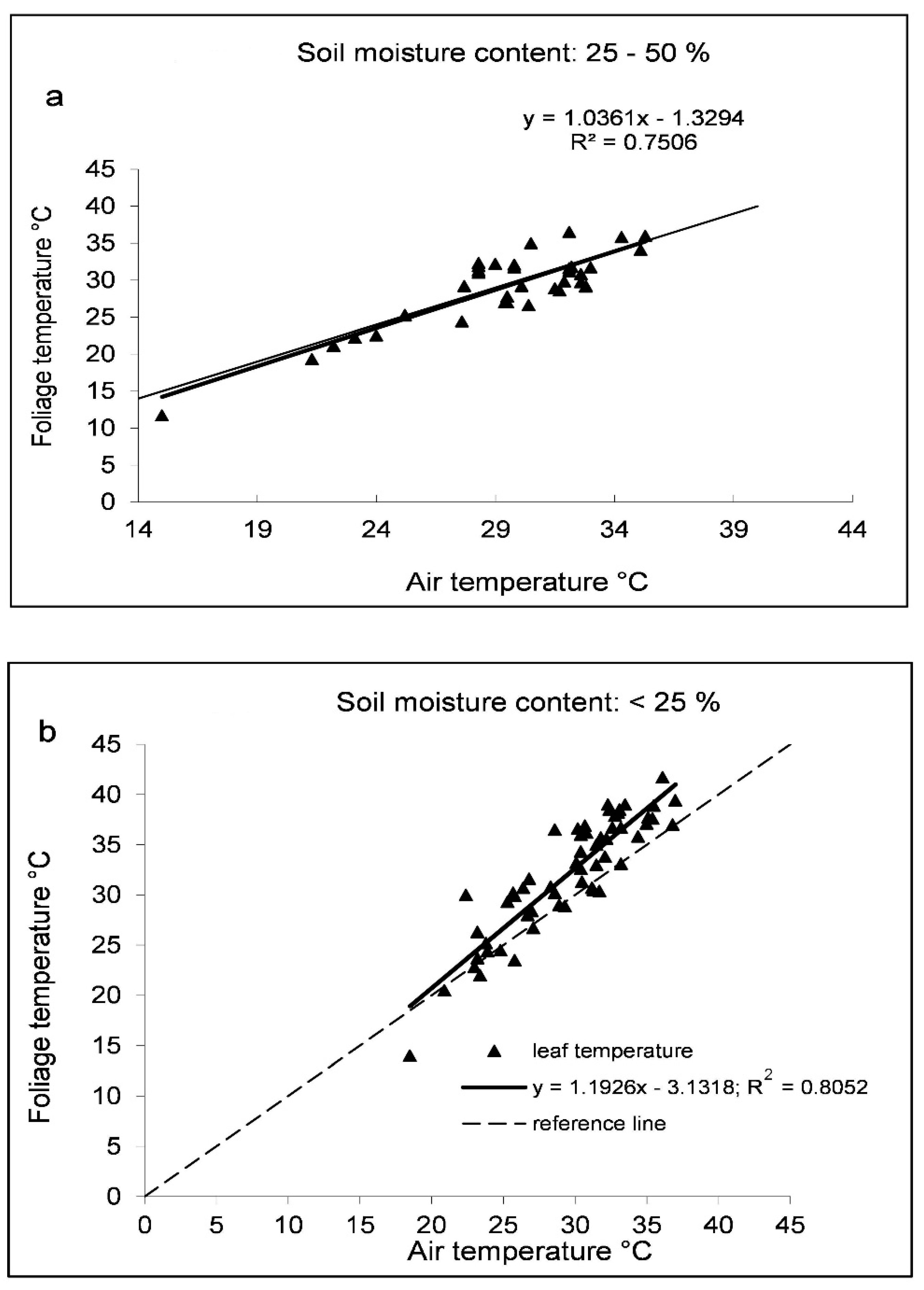
Under high photosynthetically active radiation (PAR) water deficit combined with high temperature results in an increase in leaf temperature and air temperature oscillation (±3–4 °C) due to the opening and closing of stomata [53]. Stomata closure triggers the decrease in the transpiration which contributes to the increase in canopy temperature of plants. One of the tasks of transpiration is to keep the temperature of plants at a favourable level for life processes. Decreasing transpiration causes the temperature of plants to increase. If soil water content is sufficient for the plant stand, the difference between canopy temperature and air temperature is zero or negative, but if the plants suffer from water stress this value is positive. An increase of 1 °C in canopy temperature related to a 10% decrease in the transpiration [31]. Size and stomatal density of genotypes are different thereby the transpiration varies in intensity which correlates with the difference of the canopy temperature of plants. Changes in canopy temperature have often been used to signal water stress [90] to evaluate the drought tolerance of bean genotypes and the difference in canopy temperature and air temperature was used for the real time irrigation [91,92,93,94]. During the daytime the canopy temperature rises along with the daily air temperature and radiation as the available soil water changes. The lowest value of crop water stress index (CWSI) of maize was measured at 10:00 and 11:00 and it was the largest between 12:00 and 13:00 [95]. Under water deficiency, the canopy temperature of both snap beans and tomatoes was higher than the air temperature from 09:00 to 15:00 however, that of tomatoes was higher than the air temperature only at 12:00 and 13:00 [96]. Under water stress conditions, between 09:00 and 15:00, the canopy temperature of snap beans was higher by 3.8 °C than the air temperature while it was lower by 1.6 °C in well-watered plants [93]. When the amount of available moisture in the soil for the plants decreases, then the transpiration is limited depending on the air temperature, which results in an increasing canopy temperature. Under moderate water deficiency, at 25–50% available soil water the canopy temperature of snap beans almost coincided with the air temperature (Figure 2a) that denoted the need for irrigation [93]. Nevertheless, the available soil water below 25% was not able to satisfy the water demands of plants. In this case the cooling of the canopy was not shown by transpiration and the temperature on the foliage surface was higher than the air temperature by 2.5 °C on average, indicating the plants suffered from water stress (Figure 2b) [93].
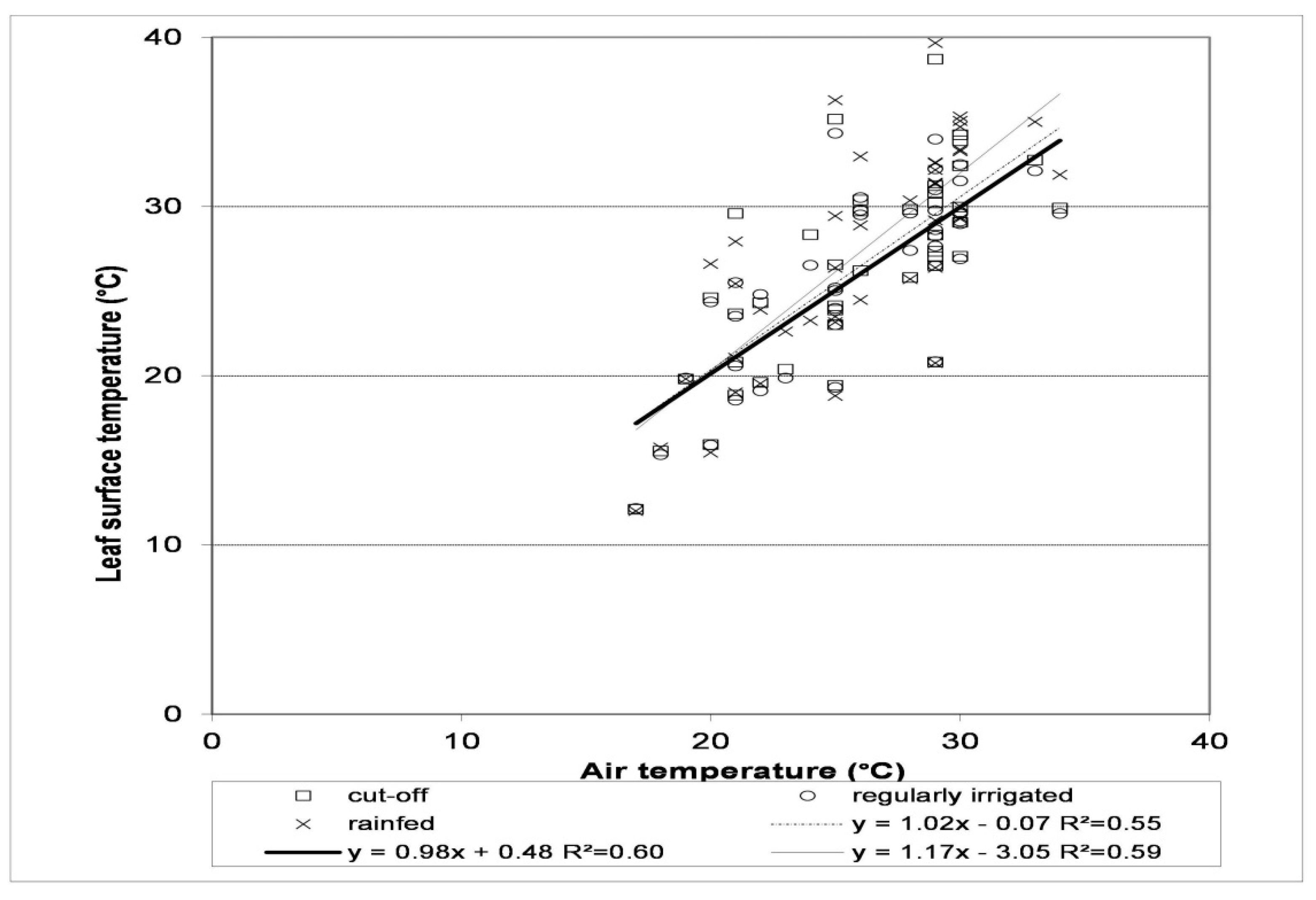
Tomatoes seemed to better use deep soil moisture with deep, strong suction force roots than the shallow rooted snap beans. Under water stress conditions, the canopy temperature of tomatoes was only higher than the air temperature by 1.8 °C, while it was significantly lower (0.6 °C) under optimal water supply conditions [92]. Air temperature had a small impact on the canopy temperature of tomatoes grown under regular irrigation and cut-off stand (i.e., irrigation was stopped 30 days before harvest) (r2 = 0.60; r2 = 0.55), however, the canopy temperature of water stressed plants increased with rising air temperature (r2 = 0.59) (Figure 3) [31]. A close correlation between canopy temperature and leaf water potential of maize was established [80] and the lowest CWSI values were measured between 10:00 and 11:00 and the highest ones between 12:00 and 13:00 [95].
4.1.3. Stomatal Conductance
Stomatal conductance indicates the speed of water vapour evaporation that depends on more plant-specific characteristics such as stomatal density, leaf age and size, guard cell and cell turgor [97]. Stomatal conductance is related to the photosynthetic assimilation rates ensuring an appropriate balance between CO2 uptake for photosynthesis and water loss through transpiration [98]. Variability in photosynthesis capacity can be explained by the CO2 diffusion through stomata and leaf mesophyll which was influenced by the mesophyll thickness and porosity and size of stomata. In drought-acclimated tomato plants the decrease in mesophyll CO2 conductance was due to an increased cell wall thickness [76]. Water stress significantly decreased the transpiration rate (37%) and stomatal conductance (26%) of maize [99]. Nevertheless, the extent of decrease in stomatal conductance depends on the growing period when the water deficiency occurred; at 7 days after anthesis of maize cultivars stomatal conductance decreased by 35% on average but at 21 days after anthesis this decrease was significantly larger (74%) under water deficiency than in well-watered cultivars [100]. In the case of tomatoes grown under non-irrigated conditions, stomatal conductance decreases from 14 to 73% depending on the weather and variety in comparison with the well-watered plants [3,32,87,101] (Table 3).
Under water scarcity, stomatal conductance for both water and CO2 flow decreased by closing the stomata [104], thus it can be said that stomatal resistance increased. The extent of stomatal resistance mainly gives information about the speed of water vapour. Under severe water deficit conditions, stomatal resistance increased by 91% for snap beans, 34% for sweet corn and 12% for green peas in comparison with the well-watered plants (Table 3). The studies shown in Table 3 proved that snap beans responded more intensively to severe water stress than sweet corn and green peas. Flowering and pod development periods of legume crops are the most sensitive to water stress when stomatal resistance changes depending on the varieties and the degree of water stress. Under moderate water deficiency, the late ripening green pea varieties had high stomatal resistance (>3.0 s cm−1), while that of green-podded snap bean varieties was relatively low (0.8–1.2 s cm−1) and yellow-podded snap beans showed different values depending on the varieties (1.0–1.43 s cm−1) [14,15]. During tasselling, the late ripening sweet corn hybrids responded with higher stomatal resistance (3 s cm−1) to medium water deficiency than during the silking period [88].
4.2. Photosynthesis in Drought
The aspects of photosynthesis of selected vegetable crops which can be measured by remote sensing methods and used for the evaluation for drought tolerance of genotypes have to be taken into consideration. In the photosynthesis process the light capture and conversion of light energy to chemical energy is made by photosynthetic pigments in the photochemistry photosystems (PSI, PSII) of leaves. The light energy in the leaf that is not used for photosynthesis is either emitted as fluorescence or released as heat [105]. The efficiency of photosynthesis can be measured by the efficiency of PSII photochemistry or by the amount of photosynthetic pigments [106].
4.2.1. Chlorophyll Fluorescence
Intense dry conditions of soil cause stomatal closure, reduced CO2 mesophyll conductance [107] and decreasing activity of PSII [108], which contributes to the decrease in photosynthesis. Photosystem II (PSII) is highly sensitive to light and drought [60] and the maximum quantum yield of PSII photochemistry (Fv/Fm) indicates an undisturbed or deficient operation of photosynthesis. Chlorophyll a fluorescence is considered to be suitable for the measurement of activity of photosynthesis because environmental stresses significantly affect the emission of chlorophyll fluorescence [109]. For example, UV-B radiation decreased the chlorophyll fluorescence of green peas [110] and ozone stress decreases the Fv/Fm and chlorophyll a concentration of leaves [111]. In higher plants, Fv/Fm fluorescence ranged from 0.78 to 0.84 [112], however this change depended on the variety and intensity of water stress.
In snap beans, the Fv/Fm ratio was relatively high (0.82–0.83) under optimal water supply conditions and it only decreased to 0.80 in the drought sensitive genotype under water stress conditions [102], which proved that chlorophyll fluorescence was not highly sensitive to water deficit.
In dry years, tomatoes grown under non-irrigated conditions had low photosynthetic activity (Fv/Fm = 0.662) and under moderate and optimal water supply conditions the Fv/Fm value ranged between 0.753 and 0.758 [3]. Likewise, the above-mentioned results from Nankishore and Farrell [32] showed a small decrease in Fv/Fm (5.1%) in tomatoes under drought (Table 3).
The maximum efficiency of PSII (Fv/Fm) of well-watered maize plants stayed constant while that of drought stressed plants stayed at control level during the first 2 days then decreased sharply as the soil became drier [103].
Use of Fv/Fm to evaluate the drought tolerance of crops is contradictory. Under controlled conditions, Fv/Fm for pot-grown grapevines decreased when water potential dropped but it seemed to be a good indicator to distinguish the moderate and severe drought stress in the field [113]. Drought stress affected the Fv/Fm parameter of the asparagus bean (Vigna unguiculata L.) [114]. Contrary to these results, no change was detected in the Fv/Fm for strawberries [115] and soybeans [116] grown under drought. Others [117,118] stated that PSII activity expressed by the Fv/Fm of drought tolerant tomato genotypes was less decreased under intensive water stress than sensitive ones. Likewise, Li et al. [119] found that Fv/Fm in drought tolerant barley varieties was higher than those of the drought sensitive group under drought stress. Under 4 day waterlogging conditions, the chlorophyll fluorescence (Fv/Fm) of flooding stress tolerant wax maize hybrids did not change significantly, while the photosynthesis efficiency of sensitive hybrids was relatively low and the Fv/Fm value decreased by 5.2% in comparison with the control [6]. The measurement of chlorophyll fluorescence as a rapid non-destructive method can be easily carried out in the field, thus it can be recommended for screening for drought tolerance [120].
4.2.2. Photosynthetic Pigments
Environmental stresses change not only the activity of the photochemistry apparatus but the chlorophyll concentration in the leaf due to metabolic disturbance [121], whereupon the light absorption decreases. Water also absorbs the radiation in the infrared wavelength of the spectrum and as the water content of leaf decreases, the light absorption decreases and reflectance increases due to the radiative attributes of water [122,123]. Therefore, the water content of leaves and the amount of photosynthetic pigments in leaves both influence the light absorption by leaves. The light absorption of the leaf can be indirectly measured by portable chlorophyll meter. In this way the calculated SPAD values correlated with the chlorophyll content of leaves [124] expresses the efficiency of photosynthesis by the intercepted photosynthetic active radiation. The high SPAD value indicates the low water and chlorophyll concentration simultaneously in the leaf, resulting in a decrease in light absorption and increase in reflectance that is larger in extent in snap beans and smaller in green peas and tomatoes (Table 3). Iturbe-Ormaetxe et al. [125] stated that the decrease in chlorophyll a concentration of leaves was larger (−30%) than that of chlorophyll b (−20.8%) for green peas exposed to water stress than in well-watered plants.
5. Relationship between Drought Stress Markers and Yield
During reproductive periods of plants that are most sensitive to water deficiency, the changes in physiological responses can be used to screen the water stress tolerance of genotypes. During this time the water supply determines the yield production. Stomatal resistance and the relative chlorophyll content of leaves (SPAD) of the individual plants indicate the disturbance of water circulation and photosynthesis. Spectral vegetation indices indicate the absorption of solar energy of the canopy in the visible light spectrum. Health status and water deficit of vegetation can be monitored by different vegetation indices and it can also determine the need for irrigation [126,127,128,129]. The normalized differential vegetation index (NDVI) expresses the ratio of spectral reflectance on the canopy in the infrared and red region and it is used to monitor the effect of water stress on plant growth and forecast biomass [130,131].
The question is how closely the physiological traits are related to water circulation and photosynthesis and can be used to predict the expected yield. Nevertheless, the physiological traits measured during the generative stages of plant species are different (Table 4). On the basis of long-term experiments, stomatal resistance measured during flowering of snap beans and tomatoes correlated with the pod yield of individual plants and weight of tomato fruits under severe drought. A close correlation between the relative chlorophyll content of leaves (SPAD) and weight of tomato fruits and final yield was detected under both mild and severe water deficiency which can be used for selection of genotypes with water stress tolerance. During tasselling of sweet corn, the expected yield of plants can be less predicted by stomatal resistance (47%) and to a higher extent by spectral traits (58–68%) under moderate water deficiency. During flowering of green peas, stomatal resistance and chlorophyll content of leaves showed a close correlation with the expected yield only under severe drought (Table 4).
Other researchers used the normalized differential vegetation index (NDVI) for yield prediction; it was successful for castor beans [132], soybeans [133] and beans [134]. According to Spitkó et al. [38], a medium correlation (r = 0.5–0.6) was detected between NDVI and final yield at 15 days after flowering of maize but not during the flowering period. Different stress indices such as stress degree days (SDD) or crop water stress index (CWSI) can be used to evaluate the water stress tolerance of genotypes [25] for scheduling of irrigation [93] and maybe for prediction of yield. In the case of sweet corn, significant negative correlation was detected between the CWSI and chlorophyll content of leaves (r = 0.802) but for the CWSI, a significant positive (r = 0.478) correlation was observed with the yield [25].
Helyes et al. [31] found a close correlation between the stress degree days (SDD) and yield of tomatoes. If the canopy temperature exceeded the air temperature (at noon), transpiration was reduced, which indicated water stress and resulted in yield reduction and quantity. Figure 4 shows the interrelation between the canopy and air temperature difference values and the yield. In our experiments the correlation was significant at p = 0.01 level with r2 = 0.57 correlation coefficient. High yield per hectare can be achieved if the difference between the cumulative canopy and the air temperature is negative during the growing season.
6. Use of Physiological Characteristics
The use of physiological traits in a breeding program, either directly by selection or stress markers, depends on their genetic correlation with the yield, heritability and genotype × environment interaction [11,135]. Under water stress, high heritability of stomatal resistance, photosynthesis rate and transpiration rate (h2 = 0.91–0.99) was found for Vigna mungo that gives a possibility for successful selection of genotypes [35]. Under severe drought, stomatal conductance and relative chlorophyll content of leaves (SPAD) measured during flowering correlated with the expected yield therefore they are suitable for the selection of individual genotypes for green peas and tomatoes, while the use of these traits for the selection of sweet corn can be efficient only under moderate water stress (Table 4). In the case of snap beans, because the water deficiency has a significant effect on leaf area, the normalized differential vegetation index (NDVI) measured during flowering can predict the expected yield more efficiently than the SPAD value of the leaves of individual plants.
Application of remote sensing techniques makes monitoring the water status of plants and real time irrigation easy [39,136]. The trend in the canopy temperature and the difference between the leaf temperature and air temperature (SDD) can be considered to be the water stress markers of plants [92]. The relationships between the canopy temperature, air temperature and transpiration involving the atmospheric and soil conditions and plant characteristics [40], was used to develop the crop water stress index (CWSI), indicating the need for irrigation. During drought, the decrease in NDVI occurred to a larger extent in snap beans, while it was less in sweet corn and hardly changed in green peas in comparison with optimal water supply conditions (Table 3). This explained why the NDVI was used as spectral indicator for irrigation scheduling mainly in snap beans [136].
In summary, some of the physiological traits influencing the decrease of water loss and biomass production of plants can be used to evaluate the water status of vegetable crops and the water stress tolerance of genotypes. During the generative period, under water deficit conditions, the changes in the stomatal conductance and chlorophyll content of leaves for individual plants is suitable for the estimation the productivity of genotypes. Nevertheless, leaf area of crops should be taken into consideration as they determine the transpiration and their chlorophyll density influences the intensity of photosynthesis and finally the yield. Water stress indices and spectral vegetation indices seemed to be more appropriate in the detection of perceived water deficiency than for the prediction of final yield.
Author Contributions
E.N. planned and wrote the first draft of the review. L.H. contributed to the writing and reviewed the final draft.
Funding
The publication was supported by the Ministry of Human Capacities grant Higher Education Institutional Excellence Program in framework of the water related research of Szent István University and grant number TUDFO/51757/2019-ITM FEKUTSTRAT and EFOP-3.6.3-VEKOP-16-2017-00008. The project is co-financed by the European Union and the European Social Fund.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ray, P. Hi-Tech Horticulture and Climate Change. In Climate Dinamics in Horticultural Science, Principles and Applications; Choudhary, M.L., Patel, V.B., Siddiqui, M.W., Mahdi, S.S., Eds.; Apple Academic Press: Oakville, ON, Canada; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2015; Volume 1, pp. 1–22. [Google Scholar]
- Nemeskéri, E.; Molnár, K.; Rácz, C.; Dobos, A.C.; Helyes, L. Effect of Water Supply on Spectral Traits and Their Relationship with the Productivity of Sweet Corns. Agronomy 2019, 9, 63. [Google Scholar] [CrossRef]
- Nemeskéri, E.; Neményi, A.; Bőcs, A.; Pék, Z.; Helyes, L. Physiological Factors and their Relationship with the Productivity of Processing Tomato under Different Water Supplies. Water 2019, 11, 586. [Google Scholar] [CrossRef]
- Aroca, R.; Porcel, R.; Ruiz-Lozano, J.M. Regulation of root water uptake under abiotic stress conditions. J. Exp. Bot. 2012, 63, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Limami, A.; Diab, H.; Lothier, J. Nitrogen metabolism in plants under low oxygen stress. Planta 2014, 239, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Li, F.H.; Shi, Z.S. Morphological and photosynthetic response of waxy corn inbred line to waterlogging. Photosynthetica 2016, 54, 636–640. [Google Scholar] [CrossRef]
- Tian, L.; Bi, W.; Liu, X.; Sun, L.; Li, J. Effects of waterlogging stress on the physiological response and grain-filling characteristics of spring maize (Zea mays L.) under field conditions. Acta Physiol. Plant. 2019, 41, 63. [Google Scholar] [CrossRef]
- Food and Agriculture Organization. FAOSTAT Crops Production. 2017. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 9 July 2018).
- Heszky, L. Szárazság és a növény kapcsolata. Agrofórum 2007, 18, 37–41. (In Hungarian) [Google Scholar]
- Patanè, C.; Tringali, S.; Sortino, O. Effects of deficit irrigation on biomass, yield, water productivity and fruit quality of processing tomato under semi-arid Mediterranean climate conditions. Sci. Hortic. 2011, 129, 590–596. [Google Scholar] [CrossRef]
- Mir, R.R.; Zaman-Allah, M.; Sreenivasulu, N.; Trethowan, R.; Varshney, R.K. Integrated genomics, physiology and breeding approaches for improving drought tolerance in crops. Theor. Appl. Genet. 2012, 125, 625–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, S.; Shi, W.; Zhang, J. An improved water-use efficiency for maize grown under regulated deficit irrigation. Field Crop. Res. 2000, 67, 207–214. [Google Scholar] [CrossRef]
- Kirda, C. Deficit Irrigation Scheduling Based on Plant Growth Stages Showing Water Stress Tolerance; Deficit Irrigation Practice Water Report 22; FAO: Rome, Italy, 2002; pp. 3–10. [Google Scholar]
- Nemeskéri, E.; Molnár, K.; Vígh, R.; Nagy, J.; Dobos, A. Relationships between stomatal behaviour, spectral traits and water use and productivity of green peas (Pisum sativum L.) in dry seasons. Acta Physiol. Plant. 2015, 37, 1–16. [Google Scholar] [CrossRef]
- Nemeskéri, E.; Molnár, K.; Pék, Z.; Helyes, L. Effect of water supply on the water use-related physiological traits and yield of snap beans in dry seasons. Irrig. Sci. 2018, 36, 143–158. [Google Scholar] [CrossRef]
- Nemeskéri, E.; Molnár, K.; Helyes, L. Relationships of spectral traits with yield and nutritional quality of snap beans (Phaseolus vulgaris L.) in dry seasons. Arch. Agron. Soil Sci. 2018, 64, 1222–1239. [Google Scholar] [CrossRef]
- Fang, X.; Turner, N.C.; Yan, G.; Li, F.; Siddique, K.H.M. Flower numbers, pod production, pollen viability, and pistil function are reduced and flower and pod abortion increased in chickpea (Cicer arietinum L.) under terminal drought. J. Exp. Bot. 2010, 61, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Behboudian, M.H.; Ma, Q.; Turner, N.C.; Palta, J.A. Reactions of chickpea to water stress: Yield and seed composition. J. Sci. Food Agric. 2001, 81, 1288–1291. [Google Scholar] [CrossRef]
- Boutraa, T.; Sanders, F.E. Influence of Water Stress on Grain Yield and Vegetative Growth of Two Cultivars of Bean (Phaseolus vulgaris L.). J. Agron. Crop. Sci. 2001, 187, 251–257. [Google Scholar] [CrossRef]
- Beshir, H.; Bueckert, R.; Tar’An, B. Effect of temporary drought at different growth stages on snap bean pod quality and yield. Afr. Crop. Sci. J. 2016, 24, 317–330. [Google Scholar] [CrossRef]
- Durigon, A.; Evers, J.; Metselaar, K.; Lier, Q.D.J.V. Water Stress Permanently Alters Shoot Architecture in Common Bean Plants. Agronomy 2019, 9, 160. [Google Scholar] [CrossRef]
- Baigorri, H.; Antolín, M.C.; Sánchez-Díaz, M. Reproductive response of two morphologically different pea cultivars to drought. Eur. J. Agron. 1999, 10, 119–128. [Google Scholar] [CrossRef]
- Moser, S.B.; Feil, B.; Jampatong, S.; Stamp, P. Effects of pre-anthesis drought, nitrogen fertilizer rate, and variety on grain yield, yield components, and harvest index of tropical maize. Agric. Water Manag. 2006, 81, 41–58. [Google Scholar] [CrossRef]
- Öktem, A. Effect of water shortage on yield, and protein and mineral compositions of drip-irrigated sweet corn in sustainable agricultural systems. Agric. Water Manag. 2008, 95, 1003–1010. [Google Scholar] [CrossRef]
- Uçak, A.B.; Öktemb, A.; Sezerc, C.; Cengizc, R.; İnald, B. Determination of arid and temperature resistant sweet corn (Zea mays saccharata Sturt) lines. Int. J. Environ. Agric. Res. 2016, 2, 79–88. [Google Scholar]
- Helyes, L.; Varga, G. Irrigation demand of tomato according to the results of three decades. Acta Hortic. 1994, 376, 323–328. [Google Scholar] [CrossRef]
- Helyes, L.; Varga, G.; Dimény, J.; Pék, Z. The simultaneous effect of variety, irrigation and weather on tomato yield. Acta Hortic. 1999, 487, 499–506. [Google Scholar] [CrossRef]
- Patane, C.; Cosentino, S.; Cosentino, S. Effects of soil water deficit on yield and quality of processing tomato under a Mediterranean climate. Agric. Water Manag. 2010, 97, 131–138. [Google Scholar] [CrossRef]
- Pires, R.C.D.M.; Furlani, P.R.; Ribeiro, R.V.; Junior, D.B.; Sakai, E.; Lourenção, A.L.; Neto, A.T. Irrigation frequency and substrate volume effects in the growth and yield of tomato plants under greenhouse conditions. Sci. Agric. 2011, 68, 400–405. [Google Scholar] [CrossRef]
- Bahadur, A.; Chatterjee, A.; Kumar, R.; Singh, M.; Naik, P.S. Physiological and biochemical basis of drought tolerance in vegetables. Veg. Sci. 2011, 38, 1–16. [Google Scholar]
- Helyes, L.; Bőcs, A.; Pék, Z. Effect of water supply on canopy temperature, stomatal conductance and yield quantity of processing tomato (Lycopersicon esculentum Mill.). Int. J. Hortic. Sci. 2010, 16, 13–15. [Google Scholar] [CrossRef]
- Nankishore, A.; Farrell, A.D. The response of contrasting tomato genotypes to combined heat and drought stress. J. Plant Physiol. 2016, 202, 75–82. [Google Scholar] [CrossRef]
- Agbna, G.H.; Dongli, S.; Zhipeng, L.; Elshaikh, N.A.; Guangcheng, S.; Timm, L.C. Effects of deficit irrigation and biochar addition on the growth, yield, and quality of tomato. Sci. Hortic. 2017, 222, 90–101. [Google Scholar] [CrossRef]
- Rodriguez-Ortega, W.; Martinez, V.; Rivero, R.; Cámara-Zapata, J.-M.; Mestre, T.; Garcia-Sanchez, F. Use of a smart irrigation system to study the effects of irrigation management on the agronomic and physiological responses of tomato plants grown under different temperatures regimes. Agric. Water Manag. 2017, 183, 158–168. [Google Scholar] [CrossRef]
- Gurumurthy, S.; Sarkar, B.; Vanaja, M.; Lakshmi, J.; Yadav, S.K.; Maheswari, M. Morpho-physiological and biochemical changes in black gram (Vigna mungo L. Hepper) genotypes under drought stress at flowering stage. Acta Physiol. Plant. 2019, 41, 42. [Google Scholar] [CrossRef]
- Stone, P.J.; Wilson, D.R.; Jamieson, P.D.; Gillespie, R.N. Water deficit effects on sweet corn. Part II. Canopy development. Aust. J. Agric. Res. 2001, 54, 115–126. [Google Scholar] [CrossRef]
- Genc, L.; Inalpulat, M.; Kizil, U.; Mirik, M.; Smith, S.E.; Mendes, M. Determination of water stress with spectral reflectance on sweet corn (Zea mays L.) using classification tree (CT) analysis. Zemdirbyste-Agriculture 2013, 100, 81–90. [Google Scholar] [CrossRef]
- Spitkó, T.; Nagy, Z.; Zsubori, Z.T.; Szőke, C.; Berzy, T.; Pintér, J.; Márton, L. Connection between normalized difference vegetation index and yield in maize. Plant Soil Environ. 2016, 62, 293–298. [Google Scholar] [CrossRef]
- Zhou, J.; Khot, L.R.; Boydston, R.A.; Miklas, P.N.; Porter, L. Low altitude remote sensing technologies for crop stress monitoring: A case study on spatial and temporal monitoring of irrigated pinto bean. Precis. Agric. 2018, 19, 555–569. [Google Scholar] [CrossRef]
- Chatterjee, A.; Solankey, S. Functional Physiology in Drought Tolerance of Vegetable Crops: An Approach to Mitigate Climate Change Impact. In Climate Dynamics in Horticultural Science, Principles and Applications; Choudhary, M.L., Patel, V.B., Siddiqui, M.W., Mahdi, S.S., Eds.; Apple Academic Press: Oakville, ON, Canada; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2015; Volume 1, pp. 149–171. [Google Scholar]
- Cattivelli, L.; Baldi, P.; Crosatti, C.; Di Fonzo, N.; Faccioli, P.; Grossi, M.; Mastrangelo, A.M.; Pecchioni, N.; Stanca, A.M. Chromosome regions and stress-related sequences involved in resistance to abiotic stress in Triticeae. Plant Mol. Biol. 2002, 48, 649–665. [Google Scholar] [CrossRef]
- Ramanjulu, S.; Bartels, D. Drought- and desiccation-induced modulation of gene expression in plants. Plant Cell Environ. 2002, 25, 141–151. [Google Scholar] [CrossRef]
- Zhang, J.; Zheng, H.G.; Aarti, A.; Pantuwan, G.; Nguyen, T.T.; Tripathy, J.N.; Sarial, A.K.; Robin, S.; Babu, R.C.; Nguyen, B.D.; et al. Locating genomic regions associated with components of drought resistance in rice: Comparative mapping within and across species. Theor. Appl. Genet. 2001, 103, 19–29. [Google Scholar] [CrossRef]
- Lanceras, J.C.; Pantuwan, G.; Jongdee, B.; Toojinda, T. Quantitative Trait Loci Associated with Drought Tolerance at Reproductive Stage in Rice. Plant Physiol. 2004, 135, 384–399. [Google Scholar] [CrossRef] [Green Version]
- Cattivelli, L.; Rizza, F.; Badeck, F.-W.; Mazzucotelli, E.; Mastrangelo, A.M.; Francia, E.; Maré, C.; Tondelli, A.; Stanca, A.M. Drought tolerance improvement in crop plants: An integrated view from breeding to genomics. Field Crop. Res. 2008, 105, 1–14. [Google Scholar] [CrossRef]
- De Leonardis, A.M.; Petrarulo, M.; Vita, P.D.; Mastrangelo, A.M. Genetic and Molecular Aspects of Plant Response to Drought in Annual Crop Species. In Advances in Selected Plant Physiology Aspects; Giuseppe, M., Dichio, B., Eds.; InTech Publisher: Rijeka, Croatia, 2012; pp. 45–74. [Google Scholar]
- Kumar, R.; Solankey, S.S.; Singh, M. Breeding for drought tolerance in vegetables. Veg. Sci. 2012, 39, 1–15. [Google Scholar]
- Augé, R.M.; Sylvia, D.M.; Park, S.; Buttery, B.R.; Saxton, A.M.; Moore, J.L.; Cho, K. Partitioning mycorrhizal influence on water relations of Phaseolus vulgaris into soil and plant components. Can. J. Bot. 2004, 82, 503–514. [Google Scholar] [CrossRef]
- Mnasri, B.; Aouani, M.E.; Mhamdi, R. Nodulation and growth of common bean (Phaseolus vulgaris) under water deficiency. Soil Biol. Biochem. 2007, 39, 1744–1750. [Google Scholar] [CrossRef]
- Dalton, D.A.; Moran, J.F.; Iturbe-Ormaetxe, I.; Matamoros, M.A.; Rubio, M.C.; Iturbe-Ormaetxe, I. Reactive oxygen species and antioxidants in legume nodules. Physiol. Plant. 2000, 109, 372–381. [Google Scholar] [Green Version]
- Collados, C.; Barea, J.M.; Ruiz-Lozano, J.M.; Azcón, R. Arbuscular mycorrhizal symbiosis can alleviate drought-induced nodule senescence in soybean plants. New Phytol. 2001, 151, 493–502. [Google Scholar]
- Esfahani, M.N.; Mostajeran, A. Rhizobial strain involvement in symbiosis efficiency of chickpea–rhizobia under drought stress: Plant growth, nitrogen fixation and antioxidant enzyme activities. Acta Physiol. Plant. 2011, 33, 1075–1083. [Google Scholar] [CrossRef]
- Nicholas, S. Plant resistance to environmental stress. Curr. Opin. Biotechnol. 1998, 9, 214–219. [Google Scholar]
- Nemeskéri, E. Water deficiency resistance study on soya and bean cultivars. Acta Agron. Hung. 2001, 49, 83–93. [Google Scholar] [CrossRef]
- Marouelli, W.A.; Silva, W.L.C. Water tension thresholds for processing tomatoes under drip irrigation in Central Brazil. Irrig. Sci. 2007, 25, 411–418. [Google Scholar] [CrossRef]
- Nemeskéri, E.; Horváth, K.; Pék, P.; Helyes, L. Effect of mycorrhizal and bacterial products on the traits related to photosynthesis and fruit quality of tomato under water deficiency conditions. Acta Hortic. 2019, 61–66. [Google Scholar] [CrossRef]
- Nilsen, E.T.; Forseth, E.N., Jr. The role of leaf movements for optimizing photosynthesis in relation to environmental variation. In Advance in Photosynthesis and Respiration, The Leaf: A Platform for Performing Photosynthesis; Adams, W.W., Terashima, I., Eds.; Springer: Basel, Switzerland, 2018; Volume 4, pp. 401–423. [Google Scholar]
- Fernandez, D.; Castrillo, M. Maize leaf rolling initiation. Photosynthetica 1999, 37, 493–497. [Google Scholar] [CrossRef]
- Sağlam, A.; Kadioglu, A.; Demiralay, M.; Terzi, R. Leaf Rolling Reduces Photosynthetic Loss in Maize Under Severe Drought. Acta Bot. Croat. 2014, 73, 315–332. [Google Scholar] [CrossRef]
- Pastenes, C.; Pimentel, P.; Lillo, J. Leaf movements and photoinhibition in relation to water stress in field-grown beans. J. Exp. Bot. 2005, 56, 425–433. [Google Scholar] [CrossRef]
- Gomes, A.M.M.; Lagoa, A.M.M.A.; Medina, C.L.; Machado, E.C.; Machado, M.A. Interactions between leaf water potential, stomatal conductance and abscisic acid content of orange trees submitted to drought stress. Braz. J. Plant Physiol. 2004, 16, 155–161. [Google Scholar] [CrossRef]
- Parry, A.D.; Horgan, R. Abscisic acid biosynthesis in roots. Planta 1992, 187, 185–191. [Google Scholar] [CrossRef]
- Reynolds-Henne, C.E.; Langenegger, A.; Mani, J.; Schenk, N.; Zumsteg, A.; Feller, U. Interactions between temperature, drought and stomatal opening in legumes. Environ. Exp. Bot. 2010, 68, 37–43. [Google Scholar] [CrossRef] [Green Version]
- De Carvalho, M.H.C.; Laffray, D.; Louguet, P. Comparison of the physiological responses of Phaseolus vulgaris and Vigna unguiculata cultivars when submitted to drought conditions. Environ. Exp. Bot. 1998, 40, 197–207. [Google Scholar] [CrossRef]
- Garcia, A.L.; Marcelis, L.; Garcia-Sanchez, F.; Nicolas, N.; Martinez, V. Moderate water stress affects tomato leaf water relations in dependence on the nitrogen supply. Biol. Plant. 2007, 51, 707–712. [Google Scholar] [CrossRef]
- Zhou, R.; Kong, L.; Wu, Z.; Rosenqvist, E.; Wang, Y.; Zhao, L.; Zhao, T.; Ottosen, C.O. Physiological response of tomatoes at drought, heat and their combination followed by recovery. Physiol. Plant. 2019, 165, 144–154. [Google Scholar] [CrossRef]
- Du, W.-J.; Yu, D.-Y.; Fu, S.-X. Analysis of QTLs for the trichome density on the upper and downer surface of leaf blade in soybean [Glycine max (L.) Merr.]. Agric. Sci. China 2009, 8, 529–537. [Google Scholar] [CrossRef]
- Nobel, P.S. Physicochemical and Environmental Plant Physiology; Academic Press: Cambridge, MA, USA, 1991; p. 635. [Google Scholar]
- Sam, O.; Jeréz, E.; Dell’Amico, J.; Ruiz-Sanchez, M.C. Water stress induced changes in anatomy of tomato leaf epidermis. Biol. Plant. 2000, 43, 275–277. [Google Scholar] [CrossRef]
- Sánchez, F.J.; Manzanares, M.; De Andres, E.F.; Tenorio, J.L.; Ayerbe, L. Residual transpiration rate, epicuticular wax load and leaf colour of pea plants in drought conditions. Influence on harvest index and canopy temperature. Eur. J. Agron. 2001, 15, 57–70. [Google Scholar] [CrossRef]
- Gomes, F.P.; Oliva, M.A.; Mielke, M.S.; Almeida, A.-A.F.; Aquino, L.A. Osmotic adjustment, proline accumulation and cell membrane stability in leaves of Cocos nucifera submitted to drought stress. Sci. Hortic. 2010, 126, 379–384. [Google Scholar] [CrossRef]
- Sánchez, F.J.; Manzanares, M.; De Andres, E.F.; Tenorio, J.L.; Ayerbe, L. Turgor maintenance, osmotic adjustment and soluble sugar and proline accumulation in 49 pea cultivars in response to water stress. Field Crop. Res. 1998, 59, 225–235. [Google Scholar] [CrossRef]
- Babita, M.; Maheswari, M.; Rao, L.; Shanker, A.K.; Rao, D.G. Osmotic adjustment, drought tolerance and yield in castor (Ricinus communis L.) hybrids. Environ. Exp. Bot. 2010, 69, 243–249. [Google Scholar] [CrossRef]
- Nemeskéri, E.; Sárdi, E.; Remenyik, J.; Kőszegi, B.; Nagy, P. Study of defensive mechanisms against drought of French bean (Phaseolus vulgaris L.) varieties. Acta Physiol. Plant. 2010, 32, 1125–1134. [Google Scholar] [CrossRef]
- Chen, Z.; Gallie, D.R. The Ascorbic Acid Redox State Controls Guard Cell Signaling and Stomatal Movement. Plant Cell 2004, 16, 1143–1162. [Google Scholar] [CrossRef] [Green Version]
- Galmés, J.; Ochogavía, J.M.; Gago, J.; Roldán, E.J.; Cifre, J.; Conesa, M.A. Leaf responses to drought stress in Mediterranean accessions of Solanum lycopersicum: Anatomical adaptations in relation to gas exchange parameters. Plant Cell Environ. 1913, 36, 920–935. [Google Scholar] [CrossRef]
- Muir, C.D.; Conesa, M.À.; Galmés, J. Independent evolution of ab- and adaxial stomatal density enables adaptation. bioRxiv 2015, 1–25. [Google Scholar] [CrossRef]
- Galmés, J.; Conesa, M.A.; Manuel Ochogavía, J.; Alejandro Perdomo, J.; Francis, D.M.; Ribas-Carbo, M.; Save, R.; Flexas, J.; Medrano, H.; Cifre, J. Physiological and morphological adaptations in relation to water use efficiency in Mediterranean accessions of Solanum lycopersicum. Plant Cell Environ. 2011, 34, 245–260. [Google Scholar] [CrossRef]
- Hardy, J.P.; Anderson, V.J.; Gardner, J.S. Stomatal characteristics, conductance ratios, and drought-induced leaf modifications of semiarid grassland species. Am. J. Bot. 1995, 82, 1–7. [Google Scholar] [CrossRef]
- Dejonge, K.C.; Taghvaeian, S.; Trout, T.J.; Comas, L.H. Comparison of canopy temperature-based water stress indices for maize. Agric. Water Manag. 2015, 156, 51–62. [Google Scholar] [CrossRef]
- Jones, H.G.; Jones, H. Use of thermography for quantitative studies of spatial and temporal variation of stomatal conductance over leaf surfaces. Plant Cell Environ. 1999, 22, 1043–1055. [Google Scholar] [CrossRef] [Green Version]
- Cornic, G.; Lawlor, D.W. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell Environ. 2002, 25, 275–294. [Google Scholar] [Green Version]
- Flagella, Z.; Campanile, R.G.; Stoppelli, M.C.; De Caro, A.; Di Fonzo, N. Drought tolerance of photosynthetic electron transport under CO2-enriched and normal air in cereal species. Physiol. Plant. 1998, 104, 753–759. [Google Scholar] [CrossRef]
- Pol, M.; Gołębiowska, D.; Miklewska, J. Influence of enhanced concentration of carbon dioxide and moderate drought on fluorescence induction in white clover (Trifolium repens L.). Photosynthetica 1999, 37, 537–542. [Google Scholar] [CrossRef]
- Yordanov, I.; Velikova, V.; Tsonev, T. Plant Responses to Drought, Acclimation, and Stress Tolerance. Photosynthetica 2000, 38, 171–186. [Google Scholar] [CrossRef]
- Ghanbari, A.A.; Shakiba, M.R.; Toorchi, M.; Choukan, R. Morpho-physiological responses of common bean leaf to water deficit stress. Eur. J. Exp. Biol. 2013, 3, 487–492. [Google Scholar]
- Guida, G.; Sellami, M.H.; Mistretta, C.; Oliva, M.; Buonomo, R.; De Mascellis, R.; Patanè, C.; Rouphael, Y.; Albrizio, R.; Giorio, P. Agronomical, physiological and fruit quality responses of two Italian long-storage tomato landraces under rain-fed and full irrigation conditions. Agric. Water Manag. 2017, 180, 126–135. [Google Scholar] [CrossRef]
- Nemeskéri, E.; Molnár, K.; Dobos, A.C. Csemegekukorica (Zea mays L. convar. saccharata) sztóma működése, és hatása a növekedésre és terméskomponensekre eltérő vízellátás alatt (Stomatal behaviour and its influence on the growing and yield components of sweet corn (Zea mays L. convar. saccharata). Növénytermelés 2017, 66, 75–95. [Google Scholar]
- Yang, H.M.; Zhang, X.Y.; Wang, G.X. Relationships between stomatal character, photosynthetic character and seed chemical composition in grass pea at different water availabilities. J. Agric. Sci. 2004, 142, 675–681. [Google Scholar] [CrossRef]
- Gonzalez-Dugo, M.P.; Moran, M.S.; Mateos, L.; Bryant, R. Canopy temperature variability as an indicator of crop water stress severity. Irrig. Sci. 2006, 24, 233–240. [Google Scholar] [CrossRef]
- Bonanno, A.R.; Mack, H.J. Use of canopy—Air temperature differentials as a method for scheduling irrigation in snap beans. J. Am. Hortic. Sci. 1983, 108, 826–831. [Google Scholar]
- Helyes, L. Relations among the water supply, foliage temperature and the yield of tomato. Acta Hortic. 1990, 277, 115–122. [Google Scholar] [CrossRef]
- Helyes, L.; Dimény, J.; Varga, G. Az öntözés tervezése a lombfelszín-hőmérséklet alapján (Scheduling of irrigation with canopy temperature). Növénytermelés 2005, 54, 341–350. [Google Scholar]
- Dufková, R. Difference in canopy and air temperature as an indicator of grassland water stress. Soil Water Res. 2006, 1, 127–138. [Google Scholar]
- Taghvaeian, S.; Chávez, J.L.; Hansen, N.C. Infrared Thermometry to Estimate Crop Water Stress Index and Water Use of Irrigated Maize in Northeastern Colorado. Remote Sens. 2012, 4, 3619–3637. [Google Scholar] [CrossRef] [Green Version]
- Helyes, L. A zöldségnövények vízellátottságának és öntözési igényének meghatározása a lombhőmérséklettel. Ph.D. Thesis, Agricultural Unversity of Gödöllő, Gödöllő, Hungary, 1991; p. 123. (In Hungarian). [Google Scholar]
- Jones, H.G. Plants and Microclimate, 2nd ed.; Cambridge University Press: Cambridge, UK, 1992; p. 428. [Google Scholar]
- Lawson, T.; Terashima, I.; Fujita, T.; Wang, Y. Coordination between Photosynthesis and Stomatal Behavior. In The Leaf: A Platform for Performing Photosynthesis; Adams, W.W., Terashima, I., Eds.; Springer: Basel, Switzerland, 2018; pp. 142–156. [Google Scholar]
- Anjum, S.A.; Wang, L.C.; Farooq, M.; Hussain, M.; Xue, L.L.; Zou, C.M. Brassinolide Application Improves the Drought Tolerance in Maize through Modulation of Enzymatic Antioxidants and Leaf Gas Exchange. J. Agron. Crop. Sci. 2011, 197, 177–185. [Google Scholar] [CrossRef]
- Sabagh, A.E.; Barutçular, C.; Islam, M.S. Relationships between stomatal conductance and yield under deficit irrigation in maize (Zea mays L.). J. Exp. Biol. Agric. Sci. 2017, 5, 15–21. [Google Scholar] [CrossRef]
- Helyes, L.; Szuvandzsiev, P.; Neményi, A.; Pék, Z.; Lugasi, A. Different water supply and stomatal conductance correlates with yield quantity and quality parameters. Acta Hortic. 2013, 971, 119–125. [Google Scholar] [CrossRef]
- Tari, I.; Camen, D.; Coradini, G.; Csiszár, J.; Fediuc, E.; Gémes, K.; Lazar, A.; Madosa, E.; Mihacea, S.; Poór, P.; et al. Changes in chlorophyll fluorescence parameters and oxidative stress responses of bush bean genotypes for selecting contrasting acclimation strategies under water stress. Acta Biol. Hung. 2008, 59, 335–345. [Google Scholar] [CrossRef]
- Yan, H.; Wu, L.; Filardo, F.; Yang, X.; Zhao, X.; Fu, D. Chemical and hydraulic signals regulate stomatal behavior and photosynthetic activity in maize during progressive drought. Acta Physiol. Plant. 2017, 39, 125. [Google Scholar] [CrossRef]
- Sing, S.K.; Reddy, K.R. Regulation of photosynthesis, fluorescence, stomatal conductance and water-use efficiency of cowpea [Vigna unguiculata (L.) Walp.] under drought. J. Photochem. Photobiol. B 2011, 105, 40–50. [Google Scholar] [CrossRef]
- Lambrev, P.H.; Miloslavina, Y.; Jahns, P.; Holzwarth, A.R. On the relationship between non-photochemical quenching and photoprotection of Photosystem II. Biochim. Biophys. Acta 2012, 1817, 760–769. [Google Scholar] [CrossRef] [Green Version]
- Bauerle, W.L.; Weston, D.J.; Bowden, J.D.; Dudley, J.B.; Toler, J.E. Leaf absorptance of photosynthetically active radiation in relation to chlorophyll meter estimates among woody plant species. Sci. Hortic. 2004, 101, 169–178. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Escalona, J.M.; Sampol, B.; Medrano, H. Effects of drought on photosynthesis in grapevines under field conditions: An evaluation of stomatal and mesophyll limitations. Funct. Plant Biol. 2002, 29, 461–471. [Google Scholar] [CrossRef]
- Filek, M.; Łabanowska, M.; Kościelniak, J.; Biesaga-Kościelniak, J.; Kurdziel, M.; Szarejko, I.; Hartikainen, H. Characterization of barley leaf tolerance to drought stress by chlorophyll fluorescence and electron paramagnetic resonance studies. J. Agron. Crop. Sci. 2015, 201, 228–240. [Google Scholar] [CrossRef]
- Guo, Y.; Tan, J. Recent advances in the application of chlorophyll a fluorescence from photosystem II. Photochem. Photobiol. 2015, 91, 1–14. [Google Scholar] [CrossRef]
- Nogués, S.; Allen, D.J.; Morison, J.I.L.; Baker, N.R. Ultraviolet-B radiation effects on water relations, leaf development, and photosynthesis in droughted pea plants. Plant Physiol. 1998, 117, 173–181. [Google Scholar] [CrossRef]
- Ismail, I.; Basahi, J.; Hassan, I. Gas exchange and chlorophyll fluorescence of pea (Pisum sativum L.) plants in response to ambient ozone at a rural site in Egypt. Sci. Total Environ. 2014, 497, 585–593. [Google Scholar] [CrossRef]
- Demmig, B.; Björkman, O. Photon yield of O2 evolution and chlorophyll fluorescence characteristics at 77 K among vascular plants of diverse origins. Planta 1987, 170, 489–504. [Google Scholar]
- Zulini, L.; Rubinigg, M.; Zorer, R.; Bertamini, M. Effects of drought stress on chlorophyll fluorescence and photosynthetic pigments in grapevine leaves (vitis vinifera cv. ’white riesling’). Acta Hortic. 2007, 754, 289–294. [Google Scholar] [CrossRef]
- Wang, B.; Liu, Y.; Wu, X.; Lu, Z.; Li, G. The relationship of chlorophyll fluorescence parameters and drought tolerance in asparagus bean seedlings under drought stress. Acta Agric. Zhejiang 2009, 21, 246–249. [Google Scholar]
- Razavi, F.; Pollet, B.; Steppe, K.; Van Labeke, M.C.; Labeke, M.C. Chlorophyll fluorescence as a tool for evaluation of drought stress in strawberry. Photosynthetica 2008, 46, 631–633. [Google Scholar] [CrossRef]
- Ohashi, Y.; Nakayama, N.; Saneoka, H.; Fujita, K. Effects of drought stress on photosynthetic gas exchange, chlorophyll fluorescence and stem diameter of soybean plants. Biol. Plant. 2006, 50, 138–141. [Google Scholar] [CrossRef]
- Srinivasa Rao, N.K.; Bhatt, R.M.; Mascarenhas, J.B.D.; Naren, A. Influence of moisture stress on leaf water status, osmotic potential, chlorophyll fluorescence and solute accumulation in field grown tomato cultivars. Veg. Sci. 1999, 26, 129–132. [Google Scholar]
- Bahadur, A.; Kumar, R.; Mishra, U.; Rai, A.; Singh, M. Physiological approaches for screening of tomato genotypes for moisture stress tolerance. In Proceedings of the National Conference of Plant Physiology (NCPP-2010) BHU, Varanasi, India, 25–27 November 2010; p. 142. [Google Scholar]
- Li, R.-H.; Guo, P.-G.; Michael, B.; Stefania, G.; Salvatore, C. Evaluation of Chlorophyll Content and Fluorescence Parameters as Indicators of Drought Tolerance in Barley. Agric. Sci. China 2006, 5, 751–757. [Google Scholar] [CrossRef]
- Woo, N.S.; Badger, M.R.; Pogson, B.J. A rapid, non-invasive procedure for quantitative assessment of drought survival using chlorophyll fluorescence. Plant Methods 2008, 4, 27. [Google Scholar] [CrossRef]
- Carter, G.A.; Knapp, A.K. Leaf optical properties in higher plants: Linking spectral characteristics to stress and chlorophyll concentration. Am. J. Bot. 2001, 88, 677–684. [Google Scholar] [CrossRef]
- Bowman, W.D. The relationship between leaf water status, gas exchange, and spectral reflectance in cotton leaves. Remote Sens. Environ. 1989, 30, 249–255. [Google Scholar] [CrossRef]
- Huntjr, E.; Rock, B. Detection of changes in leaf water content using Near- and Middle-Infrared reflectances. Remote Sens. Environ. 1989, 30, 43–54. [Google Scholar] [CrossRef]
- Yadava, U.L. A rapid and nondestructive method to determine chlorophyll in intact leaves. Hortic. Sci. 1986, 1, 1449–1450. [Google Scholar]
- Iturbe-Ormaetxe, I. Oxidative Damage in Pea Plants Exposed to Water Deficit or Paraquat. Plant Physiol. 1998, 116, 173–181. [Google Scholar] [CrossRef]
- Anderson, M.C.; Zolin, C.A.; Sentelhas, P.C.; Hain, C.R.; Semmens, K.; Yilmaz, M.T.; Gao, F.; Otkin, J.A.; Tetrault, R. The Evaporative Stress Index as an indicator of agricultural drought in Brazil: An assessment based on crop yield impacts. Remote Sens. Environ. 2016, 174, 82–99. [Google Scholar] [CrossRef]
- Xu, H.; Zhu, S.; Ying, Y.; Jiang, H. Application of multispectral reflectance for early detection of tomato disease. In Proceedings of the Optics for Natural Resources, Agriculture, and Foods, Boston, MA, USA, 1–4 October 2006; Volume 6381. [Google Scholar] [CrossRef]
- Zwart, S.J.; Leclert, L.M.C. A remote sensing-based irrigation performance assessment: A case study of the Office du Niger in Mali. Irrig. Sci. 2010, 28, 371–385. [Google Scholar] [CrossRef]
- Chawade, A.; Van Ham, J.; Blomquist, H.; Bagge, O.; Alexandersson, E.; Ortiz, R. High-Throughput Field-Phenotyping Tools for Plant Breeding and Precision Agriculture. Agronomy 2019, 9, 258. [Google Scholar] [CrossRef]
- Romano, G.; Zia, S.; Spreer, W.; Sanchez, C.; Cairns, J.; Araus, J.L.; Müller, J. Use of thermography for high throughtput phenotyping of tropical maize adaptation in water stress. Comput. Electron. Agric. 2011, 79, 67–74. [Google Scholar] [CrossRef]
- Thenkabail, P.S.; Smith, R.B.; De Pauw, E. Hyperspectral Vegetation Indices and Their Relationships with Agricultural Crop Characteristics. Remote Sens. Environ. 2000, 71, 158–182. [Google Scholar] [CrossRef]
- Li, G.; Zhang, H.; Wu, X.; Shi, C.; Huang, X.; Qin, P. Canopy reflectance in two castor bean varieties (Ricinus communis L.) for growth assessment and yield prediction on coastal saline land of Yancheng District, China. Ind. Crop. Prod. 2011, 33, 395–402. [Google Scholar] [CrossRef]
- Ma, B.L.; Dwyer, L.M.; Costa, C.; Cober, E.R.; Morrison, M.J. Early Prediction of Soybean Yield from Canopy Reflectance Measurements. Agron. J. 2001, 93, 1227–1234. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Rodríguez, M.; Escalante-Estrada, J.A.; Gonzalez, M.T.R.; Reynolds, M.P. Canopy reflectance indices and its relationship with yield in common bean plants (Phaseolus vulgaris L.) with phosphorous supply. Int. J. Agric. Biol. 2006, 8, 203–207. [Google Scholar]
- Scully, B.; Wallace, D.; Viands, D. Heritability and Correlation of Biomass, Growth Rates, Harvest Index, and Phenology to the Yield of Common Beans. J. Am. Soc. Hortic. Sci. 1991, 116, 127–130. [Google Scholar] [CrossRef] [Green Version]
- Köksal, E.S. Hyperspectral reflectance data processing through cluster and principal component analysis for estimating irrigation and yield related indicators. Agric. Water Manag. 2011, 98, 1317–1328. [Google Scholar] [CrossRef]
Figure 1.
Defence mechanism against drought (Leonardis et al. [46]).
Figure 1.
Defence mechanism against drought (Leonardis et al. [46]).

Figure 2.
Relationship between air and canopy temperature for snap beans under water deficit (a) and severe water stress (b). The thick line shows the increase in leaf temperature compared to air temperature (broken line) Source: Helyes et al. [93].
Figure 2.
Relationship between air and canopy temperature for snap beans under water deficit (a) and severe water stress (b). The thick line shows the increase in leaf temperature compared to air temperature (broken line) Source: Helyes et al. [93].

Figure 3.
Relationship between the air and canopy temperature for the Kecskeméti jubileum tomato variety under rain-fed (thin line), cut-off (broken line) and regularly irrigated (thick line) conditions Source: Helyes et al. [31].
Figure 3.
Relationship between the air and canopy temperature for the Kecskeméti jubileum tomato variety under rain-fed (thin line), cut-off (broken line) and regularly irrigated (thick line) conditions Source: Helyes et al. [31].

Figure 4.
Correlation between canopy and air temperature differences and yield Source: [31].
Figure 4.
Correlation between canopy and air temperature differences and yield Source: [31].


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Physiological traits relevant for response to drought.
| Physiological Traits | Effect Relevant for Yield | Alteration under Stress | References |
|---|---|---|---|
| Size and density of stomata | relation to leaf water potential and water consumption | increase/decrease depending on species | Hardy et al. [79], Nemeskéri et al. [14,15] |
| Leaf temperature | relation to transpiration | increase | Helyes et al. [31], DeJonge et al. [80] |
| Stomatal conductance | correlation with water consumption, decrease in individual yield | decrease in diffusion of CO2, stomatal resistance increases | Jones [81], Nemeskéri et al. [14,15] |
| Photosynthetic capacity | modulation of activity enzymes of Calvin cycle | reduction under stress | Lawlor and Cornic [82] |
| Change in chlorophyll fluorescence | alteration of quantum yield of PSII photosystem | decrease in Fv/Fm under severe drought | Flagella et al. [83], Pol et al. [84], Yordanov et al. [85] |
| Chlorophyll content of leaf | decrease in photosynthesis | decrease under stress, relative chlorophyll content (SPAD value) can increase | Nankishore and Farrell [32], Nemeskéri et al. [16] |
| Reduced growth rate | leaf area reduces, biomass decreases | Ghanbari et al. [86], Guida et al. [87] |
Table 2.
Size and density of stomata measured during generative stages of vegetable crops under different water supplies Source: modified from Nemeskéri et al. [14,15,88].
| Species | Water Supply | Lower Epidermis | Upper Epidermis | ||
|---|---|---|---|---|---|
| Stomata mm−2 * | Size of Stomata µ | Stomata mm−2 | Size of Stomata µ | ||
| Snap bean | I0 | 387.79 | 23.72 | 104.81 | 30.51 |
| DI | 374.17 | - | 93.41 | - | |
| WI | 331.22 | 24.90 | 78.61 | 29.64 | |
| average | 364.39 | 24.31 | 92.28 | 30.08 | |
| Green pea | I0 | 214.29 | 25.82 | 165.70 | 25.79 |
| DI | 214.65 | 25.48 | 170.86 | 24.68 | |
| WI | 214.74 | 24.35 | 194.72 | 25.21 | |
| average | 214.56 | 25.22 | 177.10 | 25.23 | |
| Sweet corn | I0 | 145.61 | - | 95.23 | - |
| DI | 140.79 | - | 94.98 | - | |
| WI | 136.13 | - | 93.73 | - | |
| average | 140.84 | 50.04 | 94.65 | 53.22 | |
Table 3.
Physiological traits related to water use and photosynthesis for vegetable crops under optimal water supply (OW) and water stress (WS) conditions.
Table 3.
Physiological traits related to water use and photosynthesis for vegetable crops under optimal water supply (OW) and water stress (WS) conditions.
| Traits | Crops | Units | OW | WS | Difference % | References |
|---|---|---|---|---|---|---|
| Stomatal resistance | green pea | s cm−1 | 2.87 | 3.22 | 12.2 | Nemeskéri et al. [14] |
| snap bean | s cm−1 | 1.33 | 2.54 | 90.9 | Nemeskéri et al. [15] | |
| sweet corn | s cm−1 | 2.13 | 2.85 | 33.8 | Nemeskéri et al. [88] | |
| Stomatal conductance | green pea | mmol m−2 s−1 | 0.57 | 0.32 | −43.9 | Gurumurthy et al. [35] |
| tomato | mmol m−2 s−1 | 1200 | 125 | −89.6 | Nankishore and Farrell [32] | |
| µmol m−2 s−1 | 457.26 | 394.95 | −13.6 | Nemeskéri et al. [3] | ||
| mol m−2 s−1 | 20.2–37.9 | 6.3–10.2 | −68.8 −73.1 | Helyes et al. [101] | ||
| Chlorophyll fluorescence | snap bean | Fv/Fm | 0.80 | 0.78 | −2.5 | Tari et al. [102] |
| maize | Fv/Fm | 0.810 | 0.695 | −14.2 | Yan et al. [103] | |
| tomato | Fv/Fm | 0.785 | 0.745 | −5.1 | Nankishore and Farrell [32] | |
| Fv/Fm | 0.748 | 0.696 | −7.0 | Nemeskéri et al. [3] | ||
| Fq’/Fm’ | 0.4 | 0.25 | −37.5 | Zhou et al. [66] | ||
| Chlorophyll content | green pea | SPAD * | 48.16 | 49.02 | 1.8 | Nemeskéri et al. [14] |
| snap bean | SPAD | 34.57 | 38.94 | 12.6 | Nemeskéri et al. [16] | |
| sweet corn | SPAD | 47.48 | 44.67 | −5.9 | Nemeskéri et al. [2] | |
| tomato | SPAD | 50.97 | 52.63 | 3.3 | Nemeskéri et al. [3] | |
| Vegetation index | green pea | NDVI | 0.679 | 0.693 | 2.3 | Nemeskéri et al. [14] |
| snap bean | NDVI | 0.778 | 0.681 | −12.5 | Nemeskéri et al. [16] | |
| sweet corn | NDVI | 0.743 | 0.711 | −4.3 | Nemeskéri et al. [2] |
* SPAD = relative chlorophyll content of leaves; NDVI = normalized differential vegetation index.
Table 4.
Correlation coefficients between physiological traits measured during flowering and yield under drought.
Table 4.
Correlation coefficients between physiological traits measured during flowering and yield under drought.
| Crops | Water Supply | NI | DI | ||
|---|---|---|---|---|---|
| Traits | Yield g plant−1 | Yield t ha−1 | Yield g plant−1 | Yield t ha−1 | |
| Green peas | SR | 0.3885 | 0.7648 | 0.3541 | 0.4371 |
| SPAD | 0.4685 | 0.7027 | 0.6378 | 0.5301 | |
| NDVI | 0.5550 | 0.7192 | 0.6200 | 0.2891 | |
| Snap beans | SR | 0.6075 | 0.4687 | 0.5249 | 0.7163 |
| SPAD | 0.4326 | 0.4671 | 0.6567 | 0.4385 | |
| NDVI | 0.4251 | 0.7300 | 0.3356 | 0.7665 | |
| Sweet corns * | SR | 0.6184 | 0.5756 | 0.6866 | 0.6214 |
| SPAD | 0.5346 | 0.4614 | 0.8221 | 0.6250 | |
| NDVI | 0.6804 | 0.4619 | 0.7648 | 0.4907 | |
| Tomato | gs | 0.6851 y | 0.7153 | 0.3026 y | 0.3018 |
| SPAD | 0.8655 y | 0.8405 | 0.9256 y | 0.8482 | |
| Fv/Fm | 0.4505 y | 0.3669 | 0.1103 y | 0.0961 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nemeskéri, E.; Helyes, L. Physiological Responses of Selected Vegetable Crop Species to Water Stress. Agronomy 2019, 9, 447. https://doi.org/10.3390/agronomy9080447
AMA Style
Nemeskéri E, Helyes L. Physiological Responses of Selected Vegetable Crop Species to Water Stress. Agronomy. 2019; 9(8):447. https://doi.org/10.3390/agronomy9080447
Chicago/Turabian StyleNemeskéri, Eszter, and Lajos Helyes. 2019. "Physiological Responses of Selected Vegetable Crop Species to Water Stress" Agronomy 9, no. 8: 447. https://doi.org/10.3390/agronomy9080447
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.