
Oxidative Stress Response in Pseudomonas aeruginosa
by
 , ,
, ,
Waleska Stephanie da Cruz Nizer
1 ,
,
Vasily Inkovskiy
1 ,
,
Zoya Versey
1,
Nikola Strempel
2,
Edana Cassol
1 and
Joerg Overhage
1,* 1
Department of Health Sciences, Carleton University, Ottawa, ON K1S 5B6, Canada
2
Institute of Functional Interfaces, Karlsruhe Institute of Technology, 76344 Karlsruhe, Germany
*
Author to whom correspondence should be addressed.
Pathogens 2021, 10(9), 1187; https://doi.org/10.3390/pathogens10091187
Submission received: 9 August 2021
/
Revised: 6 September 2021
/
Accepted: 8 September 2021
/
Published: 14 September 2021
(This article belongs to the Special Issue Pseudomonas aeruginosa Pathogenesis)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Pseudomonas aeruginosa is a Gram-negative environmental and human opportunistic pathogen highly adapted to many different environmental conditions. It can cause a wide range of serious infections, including wounds, lungs, the urinary tract, and systemic infections. The high versatility and pathogenicity of this bacterium is attributed to its genomic complexity, the expression of several virulence factors, and its intrinsic resistance to various antimicrobials. However, to thrive and establish infection, P. aeruginosa must overcome several barriers. One of these barriers is the presence of oxidizing agents (e.g., hydrogen peroxide, superoxide, and hypochlorous acid) produced by the host immune system or that are commonly used as disinfectants in a variety of different environments including hospitals. These agents damage several cellular molecules and can cause cell death. Therefore, bacteria adapt to these harsh conditions by altering gene expression and eliciting several stress responses to survive under oxidative stress. Here, we used PubMed to evaluate the current knowledge on the oxidative stress responses adopted by P. aeruginosa. We will describe the genes that are often differently expressed under oxidative stress conditions, the pathways and proteins employed to sense and respond to oxidative stress, and how these changes in gene expression influence pathogenicity and the virulence of P. aeruginosa. Understanding these responses and changes in gene expression is critical to controlling bacterial pathogenicity and developing new therapeutic agents.
1. Pseudomonas aeruginosa Pathogenicity and Virulence
Pseudomonas aeruginosa is a ubiquitous environmental Gram-negative bacterium. It causes several hospital-acquired infections, such as severe systemic infections, endocarditis, ventilator-associated pneumonia, and urinary tract infections [1,2]. As an opportunistic pathogen, P. aeruginosa successfully causes infections in immunocompromised individuals, patients on mechanical ventilation, elderly people, or patients with comorbidities. [3]. Furthermore, P. aeruginosa is considered to be the leading cause of chronic lung infections in cystic fibrosis (CF) patients, often resulting in lung failure [4,5,6]. In particular, multidrug-resistant (MDR) P. aeruginosa, which causes more than 50,000 healthcare-associated infections in the United States annually, is considered an emerging threat to human health [7].
The pathogenicity and success of P. aeruginosa as a human and environmental pathogen are attributed to several factors, including (i) its ability to develop adaptive and acquired resistance mechanisms, as well as its intrinsic resistance to several classes of antibiotics and disinfectants [8]; (ii) the expression of an arsenal of virulence factors [9]; and (iii) its ability to thrive in different environmental conditions, such as water with very low nutrient content [10].
P. aeruginosa is intrinsically resistant to many commonly used antibiotics due to low outer membrane permeability, which impairs drug influx. [11]. Active efflux pumps also promote the rapid transport of toxic compounds through the membrane [12,13]. Furthermore, the expression of antibiotic-cleaving enzymes also confers resistance to several classes of antibiotics. For instance, P. aeruginosa is inherently resistant to β-lactam antibiotics, such as penicillins and narrow-spectrum cephalosporins, due to the expression of the β-lactamase AmpC [12].
In addition to the intrinsic resistance to several classes of antibiotics, P. aeruginosa, like many other bacteria, can also acquire antibiotic resistance via horizontal gene transfer of plasmid-encoded resistance determinants and genetic mutations [11,12,13]. The development of such mutational resistances is favored by the direct exposure of bacteria to DNA-damaging substances [13], as is the case with different disinfection by-products or antibiotics [14,15], which may lead to the permanent overexpression of resistance-associated genes, such as multidrug efflux pumps [11]. In contrast to intrinsic and acquired resistances, which are usually stable, inheritable, and in general independent of the environment, adaptive resistance mechanisms are specifically induced under certain environmental conditions. They are characterized by changes in gene expression patterns leading to a temporary increase in antimicrobial resistance. An important example of adaptive resistance is the formation of biofilms [11].
In addition to antimicrobial resistance, the pathogenicity of P. aeruginosa is also attributed to the expression of multiple virulence factors. These factors comprise: the expression of (i) surface-bound flagella and type IV pili, which are responsible for bacterial motility and adhesion to a surface [16,17,18]; (ii) the type III secretion system (T3SS), a needle-like complex that allows the direct injection of the toxic effector proteins ExoY, ExoS, ExoT, and ExoU into host cells [19]; (iii) secreted exocompounds, such as proteases, exotoxins, pigments, and rhamnolipids [20,21,22]; (iv) alginate, which is the main component of the capsule that characterizes the mucoid phenotype and represents a common phenotype in chronic CF lung infections [23,24]; and (v) lipopolysaccharide (LPS), which represents a physical barrier against antimicrobials [25]. Most of these virulence factors contribute to the pathogenicity of P. aeruginosa by enabling the pathogen to invade and compromise the host immune responses. For instance, pyocyanin has been reported to induce tissue damage by penetrating biological membranes, inactivating certain enzymes, and inducing apoptotic cell death of neutrophils and epithelial cells via the formation of reactive hydroxyl radicals [26].
The formation of biofilms is not only considered an important virulence factor, but it is also involved in antimicrobial resistance. Biofilms are microbial communities attached to surfaces that serve as protection against harsh environmental conditions. The formation of these sessile structures is induced by diverse factors, such as pH, oxygen availability, and cellular metabolites [27,28]. In this context, studies have shown that P. aeruginosa biofilm formation is stimulated by the presence of sublethal concentrations of antimicrobials, such as the aminoglycosides tobramycin and gentamicin [29,30], β-lactam imipenem [31], the fluoroquinolone ciprofloxacin [30], the disinfectant sodium hypochlorite [32], and the detergent sodium dodecyl sulfate (SDS) [33] (reviewed in [34]). This finding may be of particular clinical interest since sublethal concentrations of antimicrobials have been reported to be used at the beginning or the end of antibiotic therapy [34,35]. Antibiotic resistance mechanisms of P. aeruginosa have previously been described in excellent reviews [11,15,36].
Despite its pathogenicity and adaptability to diverse environmental conditions, P. aeruginosa is continuously confronted by oxidative stress in its natural environments, whether produced endogenously via respiration or exogenously by the host immune system or disinfectants. These agents react with several cellular molecules and can cause irreversible damage to the bacteria, leading to cell death. Therefore, bacteria have developed an arsenal of adaptive mechanisms to respond to and manage oxidative stress damage. In this article, we will start by describing reactive oxygen and chlorine species (ROS and RCS, respectively), their mechanisms of action, and where bacteria encounter them. We will then summarize the currently available knowledge on the strategies adopted by P. aeruginosa to counteract the toxic effects of oxidizing agents. We will describe cellular responses, such as the induction of a mucoid phenotype and production of detoxifying enzymes, as well as the activation of transcriptional regulators and induction of antibiotic resistance by oxidative stress.
2. Oxidative Stress
Oxidizing agents are low molecular-weight molecules that can easily penetrate the membrane or cell wall of bacterial cells and cause intracellular damage. Due to their antimicrobial effect, many oxidizing agents have been used to eradicate pathogens [37,38]. Among the most common examples of oxidizing agents, ROS and RCS, such as hydrogen peroxide (H2O2) and hypochlorous acid (HOCl), are the most widely used disinfectants. Furthermore, they are also produced by the host immune system as a defense against invading pathogens and can also be endogenously produced by bacteria [38]. Although the reactivity of these toxic oxygen species can vary, they share many common modes of action, including the oxidation of (i) proteins, which leads to the disruption of protein and DNA synthesis and enzyme activity; (ii) lipids, leading to membrane destabilization; and (iii) nucleic acids, causing DNA breaks [37,39].
2.1. Reactive Oxygen Species
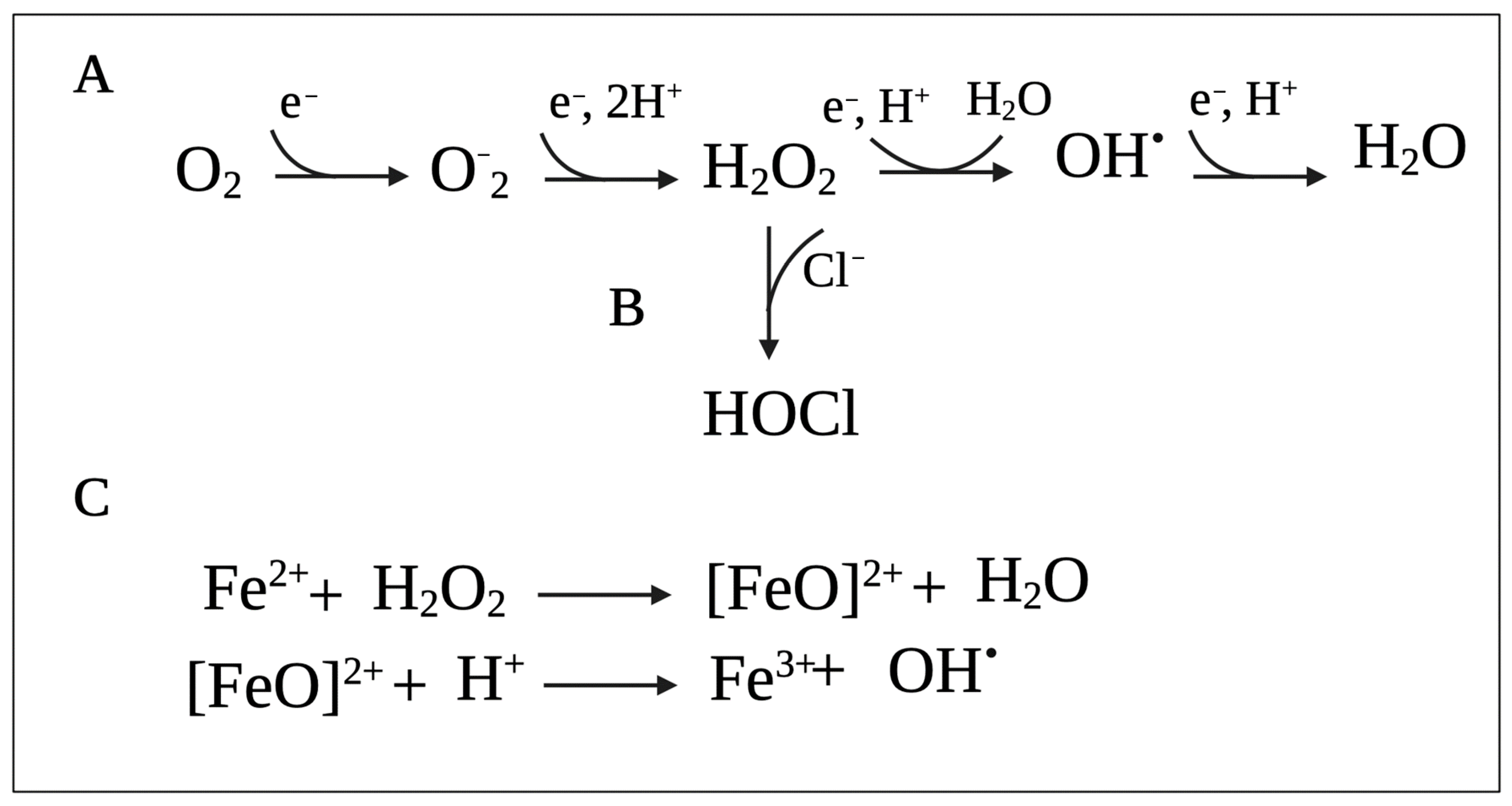
Reactive oxygen species (ROS) (i.e., superoxide (O2−), H2O2, and hydroxyl radicals (HO•)) represent the most common groups of oxidizing agents, which are formed when molecular oxygen (O2) acquires single electrons (Figure 1A) [40,41]. These toxic oxygen species can be produced by the bacteria or by the host immune system or can be found in the environment as disinfectants and antimicrobials.
In bacterial cells, flavoprotein enzymes are believed to be the primary source of endogenous O2− and H2O2 [42,43]. It was shown that ROS is primarily generated by the autoxidation of reduced flavoproteins not involved in the respiratory chain since bacterial mutants lacking respiratory components also produced ROS at comparable rates [40,44]. Examples of these enzymes include NADH dehydrogenase II, used in the aerobic respiratory chain as a primary site of electron transfer to oxygen, and fumarate reductase, which is a terminal oxidase active under anaerobic conditions [45]. Fumarate reductase reacts rapidly with oxygen and is believed to be the major source of oxidative stress when facultative anaerobes enter aerobic environments [40,45]. Importantly, since autoxidation depends on the collision frequency between enzymes and oxygen, higher oxygen concentrations lead to increased ROS production [44].
Studies also suggest that in addition to the traditional mechanisms of action, antibiotic lethality is also mediated (at least in parts) by ROS produced due to alterations in bacterial metabolism, respiration, and iron homeostasis [46]. For example, one study in P. aeruginosa showed that exposure to antibiotics accelerated cell death by promoting the Fenton reaction (Figure 1C), in which HO• is generated by the reaction between ferrous iron (Fe2+) and H2O2 [47]. Additionally, the generation of ROS due to the exposure of E. coli to sublethal concentrations of antibiotics increases their mutation rates due to oxidative damage of DNA and leads to the emergence of MDR [48]. Moreover, Foti et al. (2012) proposed that antibiotic lethality results from a failure to repair closely spaced oxidized nucleotides 8-oxo-deoxyguanosine (8-oxo-dG), leading to the formation of double-stranded breaks [49]. Despite this evidence, exact mechanisms of ROS-dependent antibiotic killing are currently unknown [46].
In addition to the endogenous production of ROS, bacteria are also exposed to exogenous oxygen species. For example, environmental H2O2 can be produced during the chemical oxidation of reduced metals and sulfur at oxic-anoxic surfaces, the reduction of molecular oxygen by flavins and chromophores, the excretion of H2O2 by lactic acid bacteria, as well as during H2O2 production by NADPH oxidases in mammalian and plant phagocytes [38]. In this context, the stimulation of phagocytes by phagocyte particles, such as bacteria, induces an increase in oxygen use, leading to the release of toxic oxygen species. For instance, the concentration of H2O2 during bacterial phagocytosis was increased by more than 50 times compared to basal conditions, in which the concentration of H2O2 was lower than 0.1 nmol/mL per min [50]. Furthermore, it was shown that 107 leukocytes produce approximately 1.03 nmol of O−2 in 15 min [51]. As a disinfectant, a concentration ranging between 5–35% (w/w) is recommended [52].
2.2. Reactive Chlorine Species
Reactive chlorine species (RCS) are comprised of a group of highly reactive compounds that are capable of oxidizing and chlorinating other molecules and include HOCl and chloramines (R2NCl) [53]. Among these, HOCl is the most potent oxidant that easily penetrates bacterial membranes and reacts with most cellular molecules, including sulfur-containing compounds (cysteine, methionine, glutathione), primary and secondary amines, nucleotides, and lipids. A summary of oxidative HOCl reactions and microbial responses to HOCl-induced stress were previously summarized in other reviews [32,53]. The main ingredient of chlorine-based bleach is sodium hypochlorite (NaOCl), which is converted into HOCl in aqueous solution [54].
RCS represent a group of oxidizing agents frequently encountered by bacteria in their native habitats [53]. One of the major sources of RCS in the environment is NaOCl, which is the main ingredient of the most commonly used chlorine-based disinfectant, bleach [32]. Bleach is frequently employed for disinfection purposes in industrial, hospital, and household settings, as well as for water disinfection and wastewater treatment [32,55]. NaOCl is only one of the forms of free chlorine in the aqueous solution, while the other forms include chlorine gas (Cl2), HOCl, and hypochlorite ion (−OCl). The relative concentrations of different forms of chlorine depend on the pH of the solution. At higher pH values (i.e., 8.510), most of the chlorine in solution is from −OCl, while at pHs between 4 and 6, the prevalent species is HOCl, and at acidic pH (lower than 4), the chlorine concentration found in solution is mostly due to the presence of Cl2 [18,48]. Out of these chlorine species, HOCl has the greatest microbicidal action [54].
2.3. Oxidative Stress Generated by the Host Immune System
ROS play an essential role in the host’s innate immune response to microbial pathogens. The main sources of ROS are professional phagocytes, such as neutrophils and macrophages, which are recruited to the site of infection to support early pathogen elimination. They recognize and bind bacterial pathogen-associated molecular patterns (PAMPs) through pattern recognition receptors (PRRs) [56], which activate various microbicidal functions, including the release of inflammatory mediators, the formation of neutrophil extracellular trap (NET), and degranulation [57,58]. It also results in increased phagocytosis and ROS production, which supports bacterial uptake and intracellular killing [53,54]. Microbes and microbial products encounter high concentrations of ROS when they are phagocytosed into intracellular compartments called phagosomes [59,60].
Although PMNs are essential in eliminating P. aeruginosa, in CF patients, they do not clear the infection since this pathogen is known to adapt to their killing mechanisms, such as by avoiding phagocytosis and forming biofilms [61]. Furthermore, the formation of biofilms and a marked inflammatory phase, which includes oxidative damage, in chronic wound infections are associated with poor outcomes [62,63]. In this context, the production of antimicrobial peptides produced by the skin’s innate response plays essential roles in the immune response of chronic wounds. However, the presence of P. aeruginosa biofilms in these infections reduces neutrophil response [62].
The two major systems that generate ROS in immune cells are the NOX2 NADPH oxidase multicomplex and mitochondrial ROS. The NOX family of NADPH oxidases is comprised of membrane-bound isoforms NOX1-5 and DUOX1-2 [64]. The NOX2 NADPH oxidase multicomplex plays a central role in this process by mediating the rapid release of ROS during an immune response, described as respiratory burst. These components assemble in the phagosome membrane to form a multicomplex [65] in response to phagocytosis. Simultaneously, azurophilic granules fuse with the phagosome to release myeloperoxidase (MPO) in the phagosome space [65]. NOX2 catalyzes the reduction of O2 into O2−, which accumulates in the phagosome [66]. The current understanding is that O2− serves as an intermediate molecule and is immediately converted to H2O2 spontaneously by superoxide dismutases (SOD) [65,66]. In addition to NOX2, the mitochondrial electron transport chain (ETC) is a key site for ROS generation. Engagement of cell surface TLRs, TLR-1, -2, and -4 by bacterial ligands lipopeptide, lipoteichoic acid, and LPS, respectively, have been shown to activate the migration of mitochondria to the phagosome membrane in macrophages [67]. ROS is primarily produced when electrons leak from ETC complexes I and III and reduce O2 and generate O2− [68]. Mitochondrial-derived vesicles containing superoxide dismutase-2 (SOD2) are delivered to the phagosome, where mitochondrial O2− dismutases to H2O2 [68,69]. Together, the NOX2 and the mitochondrial ETC amplify H2O2 production within the phagosome.
In addition to mediating antimicrobial effects, H2O2 also serves as a substrate for RCS generation. H2O2 can be oxidized by ferrous iron to generate HO• [66]. However, the phagosome holds high Cl− concentrations [70]. As such, it is accepted that most of the H2O2 is catalyzed by MPO to generate HOCl (Figure 1B) predominantly, and to a lesser extent, HOSCN [71,72,73,74]. These products directly kill the ingested bacteria. In comparison to H2O2, HOCl requires lower concentrations to cause lethal effects in bacteria [53]. For instance, it was shown that 0.2 mmol of HOCl generated by approximately 106 stimulated neutrophils can destroy as many as one million E. coli cells in an extremely short time [75]. Interestingly, Winterbourn et al. [65] have proposed that before HOCl can reach the bacteria, it targets phagosomal proteins to form chloramines. It may seem counterintuitive, but small chloramines, e.g., monochloramine (NH2Cl), have the potential to diffuse into bacterial cells and exert cytotoxic effects [67]. To counteract this issue and improve the effectiveness of HOCl, MPO can selectively bind to cells of some bacterial species to drive HOCl to its intended target [76].
2.4. Mechanisms of Oxidative Cell Damage
Oxidizing agents share many cellular targets, such as thiol groups of proteins, nucleotide bases, and lipids, which can affect ATP, DNA, and protein synthesis, enzyme activity, and membrane integrity [37]. An important aspect of the microbicidal activity of HOCl and ROS is their ability to easily penetrate bacterial membranes due to their neutrality. Inside bacterial cells, they can react with many targets and disrupt multiple cellular processes [40,54]. They can also work in concert to drive bacterial killing [77]. Specifically, studies in E. coli have shown that exposure to HOCl can deplete bacterial antioxidant enzymes, including SOD and glucose-6-phosphate dehydrogenase (G6PD), which potentiates oxidative damage caused by ROS when cells enter an aerobic environment [77].
Oxidative stress induced by ROS can lead to covalent modifications of amino acids, affecting protein structure, including carbonylation of amino acid side chains of proline, arginine, lysine, and threonine, which is irreversible. Carbonyl derivates can also be formed in histidine, lysine, and cysteine as a result of reactions with secondary oxidation products, including carbonyl compounds on carbohydrates and lipids and advanced glycation/lipoxidation end products [78]. Notably, side chains of methionine and cysteine are susceptible to oxidation due to an electron-rich sulfur atom [41].
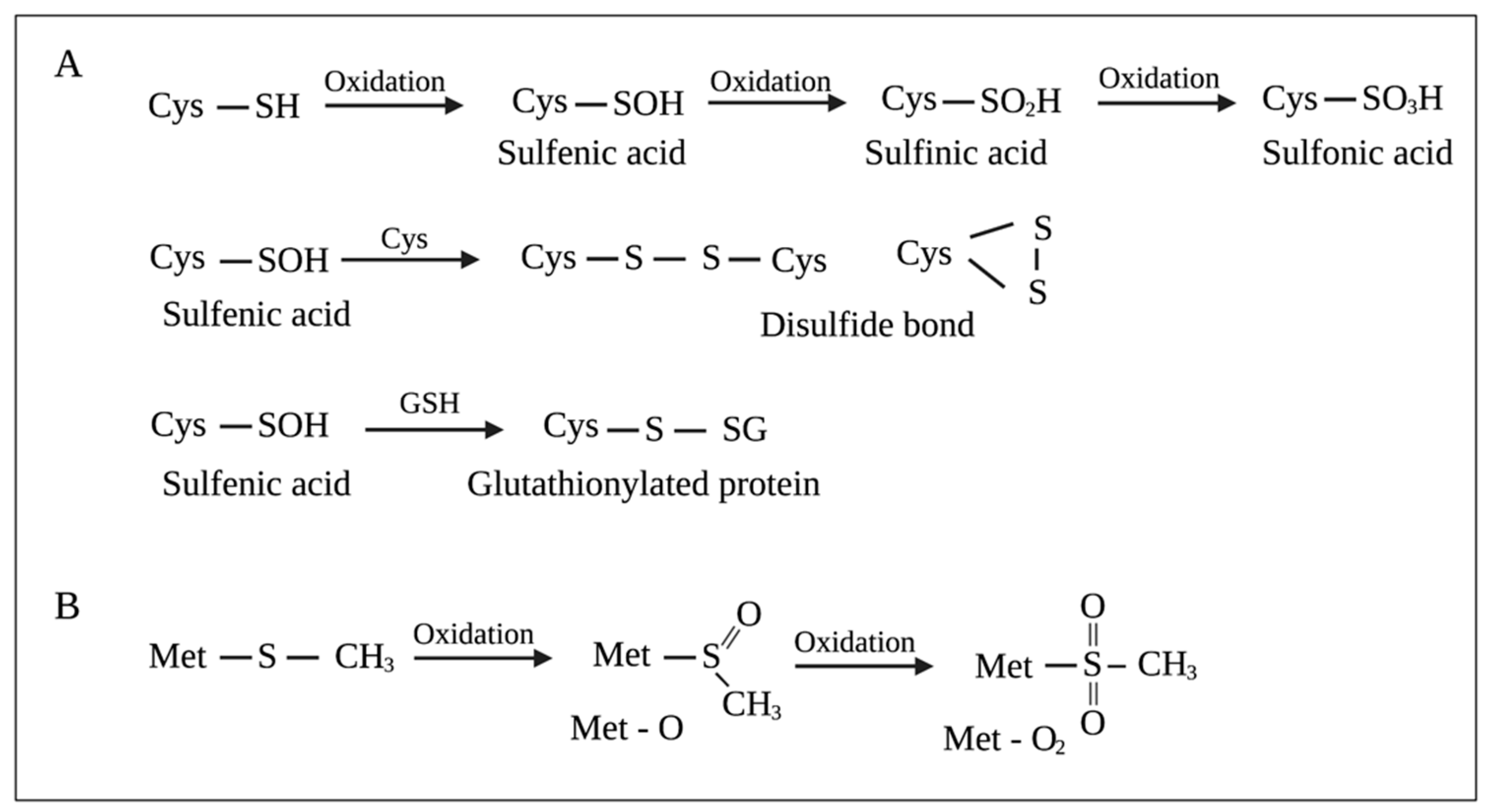
The cysteine (Cys) side chain bears a thiol (–SH) functional group, making it a strong nucleophile. Free Cys has a pKa of approximately 8.6, which can be decreased by 3–4 units in the presence of strong positively charged residues, favoring the deprotonation of thiol (–SH) and the formation of thiolate (–S−) at physiological pH. Thiolates rapidly react with electrophiles and oxidants. Two-electron oxidants, such as H2O2, react with thiolates to produce sulfenic acids (–SOH). One-electron oxidants, O2− and HO•, react with thiolates to produce thiyl radicals (–S•), which can then react with HO• to form –SOH [79]. –SOH can react with Cys residues to either form intra-disulfide or inter-disulfide bonds (-S–S-) or react with glutathione to produce glutathionylated proteins (-S–SG). Additionally, –SOH can be further oxidized to sulfinic acids (–SO2H) and sulfonic acids (–SO3H) [80] (Figure 2A). The oxidation of the thioether (–SR) side chain of methionine (Met) produces methionine sulfoxide (Met-O) (Figure 2B). Further oxidation of Met-O to methionine sulfone (Met-O2) occurs to a much lesser extent [78,81] (Figure 2B).
HOCl can also oxidize proteins, which leads to protein unfolding and irreversible aggregation [82]. Similar to ROS, HOCl most rapidly reacts with Cys and Met in the protein side chains owing to their electron-rich sulfur atom. Moreover, the oxidation of Cys and Met by HOCl and ROS generates the same final products [41]. HOCl can damage membrane proteins involved in energy transduction and transport and cause rapid hydrolysis of ATP [83]. It can also oxidize membrane-bound F1-ATP synthase, leading to the disruption of energy production in bacteria [84]. It can also act on membranes by oxidizing and inhibiting transport proteins or depleting metabolic energy necessary for transport [85].
H2O2 and HOCl also react with iron centers in microbial enzymes, leading to enzyme inactivation. H2O2 is also capable of causing Fe3+ loss and leading to the release of intracellular Fe2+ that can react with H2O2 to produce highly reactive HO•. HOCl can react with the Fe2+ itself, which also produces HO• [54]. The Fenton reaction between Fe2+ ions and H2O2 is shown in Figure 1C.
ROS and HOCl can also damage DNA. H2O2 reaction with the Fe–S cluster in enzymes leads to the release of intracellular iron, which localizes along the phosphodiester backbone of DNA [45]. The reaction of H2O2 with this unincorporated iron leads to the production of HO• that can oxidize ribose moieties or bases and cause a variety of DNA lesions [40]. Guanine residues appear to be particularly susceptible to oxidation due to low reduction potential, which leads to electron movement from guanine to nearby base radicals [86,87]. Hence, the original site of damage is repaired, while lesioned guanine is converted into mutagenic 8-hydroxyguanine, which can base pair with adenine and escape proofread systems [40,88]. Importantly, since iron is required for the Fenton reaction, higher levels of unincorporated cellular iron should increase DNA damage. Indeed, E. coli cells that have disrupted iron metabolism due to mutations in the ferric uptake regulator (fur) exhibited higher levels of DNA damage [89]. In P. aeruginosa, complete fur knockouts have not been obtained due to the essential role of the global-iron regulator [90]. However, P. aeruginosa mutants with altered expression of fur proteins showed increased sensitivity to oxidative stress [91]. Additionally, it was revealed that oxidative stress from O2− causes an increase in the intracellular pool of iron in E. coli, leading to DNA damage [92]. HOCl reactions with DNA, RNA, and polynucleotides can yield unstable chloramines that degrade to nitrogen-centered radicals. These radicals were shown to cause single- and double-stranded DNA breaks [93].
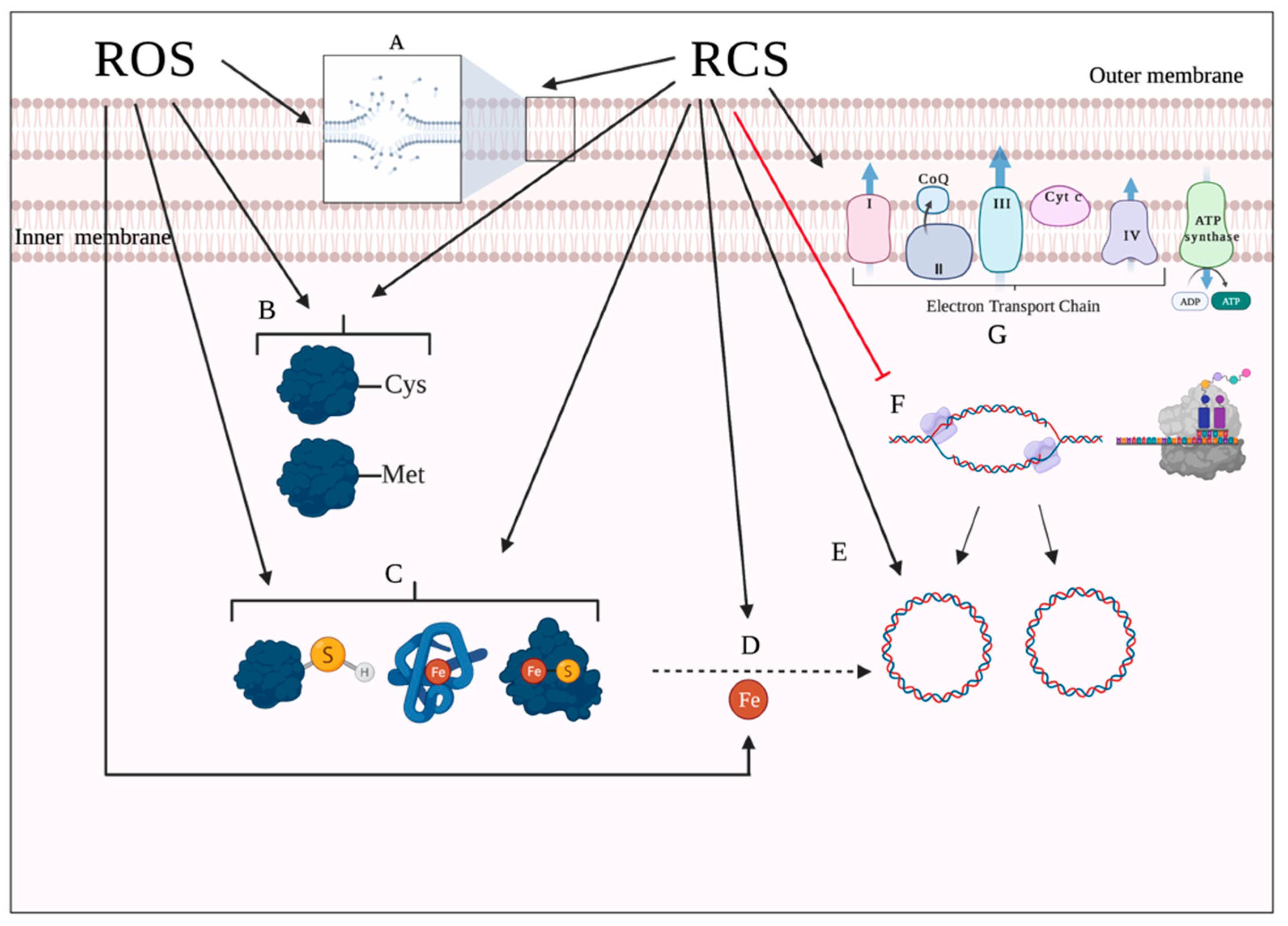
Lipids represent another target site of oxidative damage. The process of lipid damage is called lipid peroxidation and occurs in three steps [29,94]. In the initiation step, a reactive oxygen metabolite, most likely HO•, abstracts a hydrogen atom from a proximal unsaturated lipid (LH), forming a fatty acid radical (L•) and triggering a peroxidation chain. In the presence of oxygen, the fatty acid radical (L•) will react with oxygen to produce peroxyl radical (LOO•) in the propagation stage. The peroxyl radicals (LOO•) can, in turn, abstract hydrogen atoms from nearby unsaturated fatty acids, forming lipid hydroperoxide (LOOH) and fatty acid radical (L•), hence amplifying the reaction chain [94,95,96]. Additionally, LOO• can decompose into aldehydes, damaging membrane proteins [97]. The last step in the process, chain termination, occurs when LOO• interacts either with an antioxidant or another lipid radical [94]. Importantly, the propagation step in the process requires polyunsaturated fatty acids, implying that saturated and monounsaturated fatty acids can only undergo oxidation [40,94]. HOCl can react with the unsaturated bonds in fatty acids and produce chlorohydrins. These molecules are more polar than unmodified lipids and thus can disrupt membrane integrity, which leads to increased membrane permeability and loss of membrane function [54]. Figure 3 summarizes the bacterial targets of ROS and HOCl.
3. Oxidative Stress Responses in P. aeruginosa
Microorganisms are constantly surrounded by oxidizing agents, whether produced endogenously by aerobic respiration or exogenously by the host cell. Therefore, almost all bacterial species have developed several adaptive responses to manage the toxic effects of these agents, including changes in the expression of genes encoding detoxifying proteins and enzymes and the activation of transcriptional regulators [98]. Figure 4 shows a schematic of the main stress responses adopted by P. aeruginosa against ROS and RCS.
3.1. Evaluation of Oxidative Stress Responses of P. aeruginosa by Transcriptomic Studies
Transcriptomic analysis refers to the study of all RNA transcripts produced by the genome of a cell under specific conditions. Through the analysis of gene expression, these genome-wide studies provide insights into the defensive responses adopted by an organism and the possible modes of action of antimicrobial agents [99]. For instance, through the evaluation of gene expression of P. aeruginosa exposed to ortho-phenylphenol (OPP), Nde and colleagues (2008) were able to conclude that a mode of action of OPP is through its interaction with amino acid residues, which induces protein synthesis to overcome their shortage in the cell [100]. Not surprisingly, transcriptomic analyses have been used to evaluate changes in gene expression of P. aeruginosa exposed to different oxidizing agents.
A common finding in transcriptomic studies of the oxidative stress responses of P. aeruginosa is the upregulation of genes involved in protective mechanisms by H2O2 [101,102,103,104], NaOCl [101,105], peracetic acid [106], HOBr, and HOSCN [105]. Among them are detoxifying enzymes, such as katB, PA2826 (glutathione peroxidase), sodM and sodB, ahpB and ahpF; DNA repair systems, including recN, prtN, prtR; heat shock proteins, such as ibpA, clpB, and hscB; and bacterial virulence exoS and exoT genes [101,102,103,104,105,106].
Exposure to NaOCl also induced the expression of genes involved in the transport of sulfur-containing compounds, such as taurine, sulfate, and sulfonate [101,107]. It is consistent with prior studies that showed that due to the reaction of HOCl with sulfur-containing molecules in the cells, the transport of these molecules should be induced to compensate their concentration decreases in the intracellular environment [32].
Another common finding of transcriptomic studies is the downregulation of primary metabolic processes genes in P. aeruginosa cells exposed to peracetic acid [106] and H2O2 [102,103,104]. These genes include energy metabolism (nuo, atp, coxB, coIII, ccoP2, ccoN2, fdxA, hcnA-C, and eutB), ribosomal biogenesis (rps and rpm), purine and pyrimidine synthesis (pur, pyr), fatty acid synthesis (accD, accC, fabABF1), polyamine and uptake genes (speADE and potABCD operon), and the sec-dependent protein translocation pathway (secABDEFGY) [102,103,104,106]. Furthermore, NaOCl exposure downregulated genes involved in carbon metabolism, including the Entner–Doudoroff (ED) and Embden–Meyerhof–Parnas (EMP) pathways and the transport of hexose molecules [101,107]. These results suggest that oxidizing agents provoke a decrease in energy production and metabolism, which could favor the survival of these cells under harsh conditions.
The studies conducted by Palma et al. (2004) and Salunkhe et al., 2005 showed that the exposure of P. aeruginosa to 1 and 10 mM of H2O2, respectively, induced the expression of several genes involved in iron homeostasis. On the other hand, Chang et al. (2005) reported the repression of iron genes regulated by the ferric uptake regulator (Fur) (e.g., pvdS, fpvA, and fptA) [102,103,104]. Iron is important for several cellular processes, including enzymatic activity, the expression of virulence factors, the transport of molecules, and the activation and decomposition of peroxides [108]. The discrepancy in these results could be attributed to the differences in the experimental conditions employed. For instance, Salunkhe et al. (2005) treated P. aeruginosa cells in the stationary phase, while Palma et al. (2004) used cells in the log phase. Therefore, the upregulation of iron-related genes could be an initial adaptation of the cells to H2O2 stress [103]. Moreover, Chang et al. (2005) reported an important finding was the overexpression of the F-, R-, and S-type pyocin induced by H2O2 exposure as a consequence of DNA damage [104].
In another study, Small and colleagues (2007) compared the gene expression profile of P. aeruginosa PAO1 upon exposure to H2O2, NaOCl, and peracetic acid [107]. The authors showed that the expression of a higher number of genes was altered in response to NaOCl treatment, confirming the strong effect of this agent on cellular molecules [32]. Overall, exposure to H2O2 resulted in the lowest number of genes dysregulated compared to the other oxidants. Peracetic acid strongly upregulated the genes glpK and glpD (glycerol-3-phosphatase and glycerol-3-phosphate dehydrogenase, respectively), which suggests that under peracetic acid stress, the cells use glycerol and glycerol-3-phosphate as the substrate for growth. Lastly, H2O2 strongly induced cellular protection genes compared to the other agents [107].
Farrant et al. (2020) evaluated the transcriptional profile of PA14 after exposure to 2.2 mM of NaOCl and 0.8 mM of HOSCN. They found that 70% of the genes upregulated by HOSCN were also induced by NaOCl, indicating that different agents share many bacterial stress responses. These genes belong to the noncoding RNA, protein secretion/export apparatus, and antibiotic resistance and susceptibility [109]. In the study conducted by Groitl et al. (2017), the authors analyzed gene expression changes of P. aeruginosa PA14 treated with HOCl, HOBr, and HOSCN. In addition to the induction of protective enzymes by all three oxidants, the regulator MexT involved in bacterial virulence was also induced by all three treatments, and the regulators Fur and PchR were induced by only HOCl. Another important result obtained by these authors was that HOCl and HOBr upregulated several chaperones and heat shock proteins (e.g., dnaK, ibpA, groES, hskU/V), while HOSCN induced only two chaperones (hslV and grpE). Moreover, HOCl and HOBr also induced several genes related to motility and attachment, chemotaxis, and nucleotide biosynthesis compared to HOSCN. On the other hand, HOSCN induced a higher number of genes in antibiotic resistance or membrane proteins, indicating that the primary mode of action of this agent is its membrane structures [105].
Transcriptomic studies and their similarities provide insights into the adaptive responses adopted by bacteria under oxidative stress. Genes that are mostly regulated in different studies and experimental conditions probably play important roles in bacterial survival under oxidative stress conditions. For instance, most studies revealed the upregulation of detoxifying enzymes, such as catalase and superoxide dismutase, which are well known to be an important protective strategy adopted by bacterial cells to manage oxidative damage. Furthermore, these studies could also help understand bacterial pathogenesis during infection. For example, the induction of genes related to bacterial virulence by oxidative stress could help understand and reduce bacterial pathogenicity during lung infections in CF patients. Overall, the differences obtained among the studies are attributed to the experimental conditions employed, such as the concentration of oxidizing agents, the bacterial strains utilized, the growth phase of the cells during the treatment, and the treatment time.
3.2. Cellular Responses to Oxidative Stress
3.2.1. Antioxidant Enzymes
The removal of oxidizing agents is the primary action taken by cells exposed to oxidative stress [110]. The detoxification of toxic oxygen species is done by specialized enzymes, such as catalase, superoxide dismutase (SOD), peroxidase, and alkyl hydroperoxidase, which catalyze the conversion of these species into less toxic substances. These enzymes are essential for the survival of bacteria under oxidative stress.
SODs are periplasmic enzymes that catalyze the conversion of O2− into H2O2, while catalases and peroxidases are present in the cytoplasm and generate H2O and O2 from H2O2. The removal of toxic oxygen species is connected to the action of these enzymes, in which SODs are considered the first defense involved in superoxide detoxification [111]. P. aeruginosa has two SODs: one that is Fe-cofactored (Fe-SOD; SodB), which is found in all growth conditions, and another one that uses manganese (Mn-SOD; SodM, previously called SodA) as a cofactor, which has been shown to be induced under iron starvation and alginate production [112,113]. Compared to Mn-SOD, Fe-SOD presents a more pronounced effect in P. aeruginosa growth under aerobic conditions, protection against superoxides, and pyocyanin production [112,114]. A significant reduction in catalase production was detected in Fe-SOD mutants compared to Mn-SOD, supporting the roles of this enzyme in oxidative stress protection [114]. Furthermore, Fe-SOD also contributes to the long-term survival of P. aeruginosa within human and murine macrophages, inhibits the activation of autophagy [111], and is involved in the P. aeruginosa infection process of a Bombyx mori model [115].
P. aeruginosa has three catalases (katA, katB, and katC) and four alkyl hydroperoxide reductases (ahpA, ahpB, ahpCF, and ohr) that detoxify H2O2. Of these, katA is considered the major catalase of P. aeruginosa, and its expression is controlled by several systems (e.g., quorum sensing (QS), iron levels, anaerobic regulator (ANR), OxyR, and IscR) [116,117]. It is extremely stable and is expressed continuously throughout all bacterial growth phases, with an observed overexpression detected during the stationary phase. In addition to its antioxidant activity, it is also involved in P. aeruginosa virulence in a Drosophila melanogaster and mice infection models, in which katA mutants presented reduced killing-effects on infected animals compared to the wild-type [118]. On the other hand, the expression of katB is induced by H2O2 stress [117,119]. KatC (also called KatE due to its homology with E. coli catalase KatE) is a temperature-inducible catalase that requires the formation of a disulfide bond for its activity [120]. The role of KatC in cellular processes and protective mechanisms of P. aeruginosa have not been completely investigated [121]. Catalase is also involved in the protection against the H2O2 of P. aeruginosa biofilms by inhibiting the penetration of this oxidizing agent into this structure [119,122,123] and contributing to P. aeruginosa’s resistance in chronic infections [123].
Alkyl hydroperoxide reductase generates alcohol and water from H2O2 and organic hydroperoxide. Similar to katA, ahpA is involved in the oxidative stress response against endogenously produced ROS since it is continually expressed during all phases of aerobic growth. Conversely, AhpCF is overexpressed under exogenous oxidative stress, being considered a protective protein against the damage caused by oxidizing agents [124,125]. In addition to its detoxifying effect, AhpCF has also been shown to possesses chaperone activity [126,127].
3.2.2. Protein Repair Systems
Methionine sulfoxide reductases (Msr) are enzymes that catalyze the reduction of Met-O generated under oxidative conditions to Met. It is conserved and found in several organisms. Most bacterial species, including P. aeruginosa, have two Msr, msrA and msrB [128,129]. The oxidation of Met generates two isomers of Met-O (i.e., Met-(S)-O and Met-(R)-O). MsrA specifically reduces Met-(S)-O, and MrsB reduces Met-(R)-SO [130,131]. In addition to its antioxidant effect, Msr is also involved in the pathogenesis of several bacterial species (reviewed in [130]). In P. aeruginosa, the level of MsrA is kept constant throughout several growth phases under physiological conditions, suggesting that this enzyme is ready to detoxify oxidized Met when the cells are exposed to oxidative stress. On the other hand, the expression of msrB is induced under oxidative stress, such as NaOCl exposure. In this context, msrA, msrB, and msrA msrB double mutants are significantly more susceptible to NaOCl and H2O2 than the wild-type. In addition to the oxidative protection, these mutants also presented reduced virulence in a Drosophila melanogaster model [129].
Oxidizing agents, such as HOCl, HOBr, and HOSCN, cause significant protein unfolding, which leads to unspecific protein aggregation [53,105]. Therefore, it is not surprising that bacteria have established diverse response mechanisms against the proteome damage caused by these agents. In this context, the major response against protein unfolding and aggregation is the activation of chaperones. The activation of these proteins by RCS stress in several bacterial species was reviewed in [32,53,132]. For instance, a study conducted by Groitl et al. (2017) showed that the overproduction of polyP by P. aeruginosa cells under HOCl stress and the substantial protein aggregation when this molecule is absent demonstrates the roles of chaperones in bacterial survival under RCS stress [105]. The roles of polyP as a chaperone have also been described in different studies for different Gram-negative species (reviewed in [32]).
3.2.3. Quorum Sensing (QS)
Quorum sensing (QS) is a mechanism used by bacteria to communicate with each other in response to cell density [133]. Most of the virulence factors produced by P. aeruginosa, including biofilm formation, motility, antibiotic resistance, the response to host immune system, and the secretion of proteases, iron chelators, and efflux pumps, are controlled at least in part by the QS system [134,135]. Furthermore, QS has also been shown to play important roles in the oxidative stress survival of P. aeruginosa. For instance, it regulates rhamnolipid production, known to be involved in oxidative stress responses due to foam formation on the surface of media. This foam prevents the entry of oxygen and, therefore, is considered a protective substance against toxic oxygen species [136,137].
The QS system regulates cell-to-cell communication through the interaction of QS signal molecules (N-acyl-L-homoserine lactones) with transcriptional regulators, which induces the expression of target genes. P. aeruginosa has three major and well-characterized QS systems: las, rhl, and the Pseudomonas quinolone signal (PQS) [138]. The Las system is composed of a transcriptional regulator LasR and the signal molecule N-(3-oxododecanoyl)-L-homoserine lactone (3-oxo-C12HSL), synthesized by the enzyme LasI. Likewise, the rhl system is constituted of the transcriptional regulator RhlR and the signal molecule N-butanoyl-L-homoserine lactone (C4-HSL), synthesized by RhlI synthase. The PQS system is composed of the transcriptional regulator PqsR and the signal molecule 2-heptyl-3-hydroxy-4-quinolone [139,140]. The las system controls the activation of the rhl and PQS systems hierarchically in response to an increase in cell density. An increase in cell population stimulates the synthesis of 3-oxo-C12HSL, which forms a complex with the transcriptional regulator LasR, activating the transcription of the genes lasI and rhlR [141]. PqsR (MvfR) was also shown to exert direct control on LasR and RhlR during early and mid-exponential phases, suggesting an interconnected regulation of these three systems [134].
In addition to its effects on P. aeruginosa’s virulence and pathogenicity, the roles of QS in bacterial survival under oxidative stress have also been demonstrated in several studies. QS controls the expression of antioxidant enzymes [142], and the presence of SodM was not detected in lasI and lasR mutants [143]. The impaired production of detoxifying enzymes by QS mutants explains their susceptibility to oxidizing agents. Furthermore, only cells with functioning QS systems can survive and effectively respond to oxidative stress [144]. The response of LasR to oxidative stress was shown to be due to the formation of a disulfide bond between two cysteine residues in the molecule (Cys201 and Cys203) [145].
Häussler and Becker (2008) showed that pqs mutants were more tolerant to H2O2 than the wild-type, indicating that PQS induces susceptibility of P. aeruginosa to oxidative stress. PQS also decreases the amount of intracellular ROS and exhibits a potent antioxidant effect due to its electron-donating potential and by inducing undamaged bacteria to enter a low metabolic state and, consequently, to become less susceptible. It also has a pro-oxidant effect, in which it also generates ROS through the Fenton reaction [146]. PqsR regulates the key genes of type 6 and type 2 secretion systems (T6SS and T2SS, respectively) at early stages of growth and induces the oxidative stress genes ahpC-ahpF, trxB2-ahpB, and dps, in which these mutants were less tolerant to H2O2 than the wild-type, demonstrating its roles in oxidative stress. Furthermore, this oxidative stress effect provoked higher resistance to the ß-lactam Meropenem than the wild-type [134].
3.2.4. Induction of a Mucoid Phenotype
The colonization of the lungs of CF patients by P. aeruginosa is the leading cause of morbidity and mortality among these individuals [147]. In the earlier stages of infection, P. aeruginosa appears as a nonmucoid bacterium; however, it switches to a mucoid phenotype in the later phases of colonization [148]. The conversion of a nonmucoid to a mucoid phenotype is caused by an overproduction of a capsule composed mainly of the exopolysaccharide alginate and leads to high mortality rates, poor prognosis, and persistent infection in CF patients [23]. Alginate is composed of two uronic acids (i.e., β-D-mannuronic acid and α-L-guluronic acid) linked together through glycosidic bonds [24] and is involved in biofilm formation, bacterial adherence to surfaces, scavenging of ROS, and physical protection against antibiotics, oxidizing agents, and the immune system [149,150,151].
The biosynthesis of alginate is a complex process that involves several genes. It is regulated by the sigma factor σ22 (AlgU, also called AlgT), which is controlled by the MucABCD system [152]. The overproduction of alginate and the consequent conversion to a mucoid phenotype is often due to a mutation in the gene that encodes the anti-σ factor MucA, but it can also occur by the degradation of MucA by the AlgW protease [153,154]. MucA is a transmembrane protein that binds to the alternative sigma factor AlgU, inhibiting the expression of the alginate operon [148,149]. AlgU, in turn, is required for the expression of algD, which is a gene encoding for a GDP-mannose 6-dehydrogenase and is the first gene of the alginate biosynthesis operon and therefore essential for the production of alginate [151].
The conversion to a mucoid phenotype has been described as an adaptive response to toxic ROS. Oxidizing agents, such as H2O2, can induce the overproduction of alginate in nonmucoid P. aeruginosa cells and consequently promote the conversion to the mucoid phenotype [136]. Mathee and colleagues (1999) showed that non-mucoid P. aeruginosa PAO1 grown in biofilms presented the mucoid phenotype and overproduced alginate after exposure to sublethal concentrations of H2O2 (1.0 mM or 2.5 mM) and human polymorphonuclear leukocytes (PMNs), while this phenomenon was not observed in untreated biofilms. It indicates that the overproduction of alginate in P. aeruginosa biofilms is an adaptive response to the immune system and its toxic oxygen species [154]. Other studies have reported that genes involved in alginate biosynthesis pathways are affected by H2O2 and paraquat exposure. For instance, algU mutants were more sensitive to paraquat [151] and hypochlorite [155] than the wild-type, and the mutations of algR impaired the stress response of P. aeruginosa to H2O2 [156].
Boucher and colleagues (1996) demonstrated the roles of the genes algW and mucD on P. aeruginosa survival under H2O2 and paraquat stress and their roles in the conversion to the mucoid phenotype. The authors showed that the inactivation of algW increased the sensitivity of P. aeruginosa PAO1 to H2O2 and paraquat, and mutations on the mucD gene conferred sensitivity to only H2O2 [157]. The difference between the protection levels conferred by these genes against H2O2 and paraquat on the cells can be explained by the fact that paraquat acts mainly in the cytoplasm, while MucD is a periplasmic protein [150,157]. Moreover, a mutation in the genes algW and mucD induced the conversion to the mucoid phenotype since they act as negative regulators of alginate synthesis [157].
3.2.5. Pigment Production
P. aeruginosa secretes several types of extracellular pigments, of which phenazines are the most common. Many P. aeruginosa strains also produce pyocyanin, pyoverdine, pyorubin, and pyomelanin. These pigments are involved in bacterial virulence and survival [158,159]. Among the different types of phenazines secreted by this bacterium, pyocyanin, a blue-green substance, is produced by most P. aeruginosa isolates [160]. It is considered an important virulence factor of P. aeruginosa and plays a vital role in iron metabolism and antimicrobial effect against competitors [161]. Pyoverdine is a fluorescent yellow-green siderophore involved in iron uptake [162], while pyomelanin and pyorubin are non-fluorescent brown and red-brown pigments, respectively [161].
The protective effect of P. aeruginosa pigments against oxidative stress induced by paraquat [159], photodynamic therapy [158], and H2O2 [163] have been reported. The exposure of P. aeruginosa to paraquat increased the growth and production of a dark-brown melanin-like pigment in a P. aeruginosa dark-brown hyperpigmented (HP) strain, suggesting that this dark-brown pigment helped the bacteria to survive under oxidative conditions and might be an oxidative response against superoxide. This hypothesis was confirmed when this pigment, which contains several antioxidant molecules, was added to E. coli cells treated with paraquat, and it protected the cells against the oxidative effect of this superoxide generating agent [159]. The production of pyomelanin is induced by the inactivation of the gene hmgA. The disruption of this gene increased the persistence of the bacteria during lung infection and oxidative tolerance but did not promote tolerance to antibiotics. Furthermore, the supernatant of the hyperproducer mutants protected the wild-type against the effects of H2O2 [163]. Together, these results suggest that the pigments produced by P. aeruginosa cells also contribute to their protection against harsh environmental conditions created by the presence of oxidizing agents, probably by containing detoxifying enzymes.
In addition to the protective effect of P. aeruginosa pigments, some of them are considered toxic to the cells. For instance, pyocyanin generates superoxide and H2O2 by reducing molecular oxygen, which can cause irreversible oxidative damage to the host cells and the bacteria. Therefore, P. aeruginosa adopts several mechanisms to counteract the auto-poisoning effects of pyocyanin, such as inducing the formation of biofilms and decreasing energy production [164], overexpressing antioxidant enzymes, such as catalase and SOD, and blocking the uptake of this phenazine [165]. Meirelles et al. (2018) demonstrated that exposure to pyocyanin strongly induced H2O2 sensing genes regulated by the OxyR transcriptional regulator, including ahpC, ahpB, ahpF, thioredoxin reductase (trxB2 and trxA), and katB. Moreover, pyocyanin also significantly upregulated genes involved in the efflux system (i.e., mexG and mexH of the efflux system mexGHI-ompD and mexEF-oprN), and slightly regulated DNA repair genes (i.e., recA and lexA). Furthermore, three genes belonging to the isc operon (i.e., iscA, iscU, and fdx2) involved in Fe–S cluster biosynthesis were upregulated in the presence of pyocyanin, which is related to the oxidative damage caused by pyocyanin to sulfur-containing compounds. Previous exposure to pyocyanin also induced the cells to enter a persister state when cultured in a nutrient-depleted media. However, when the cells were transferred to nutrient-rich media, they became sensitive again to pyocyanin, indicating that the toxicity of pyocyanin depends on environmental conditions [164]. The production of pyocyanin by P. aeruginosa also induced the persister state in A. baumannii [166].
3.2.6. Carbon Metabolism
According to the growth conditions, P. aeruginosa can adapt and utilize distinct pathways to metabolize different carbon sources, which provides it with the ability to survive in different environmental conditions [167]. For instance, P. aeruginosa PAO1 and other pseudomonads catabolize glucose predominately through the Entner–Doudoroff pathway (EDP) [168], which could be considered a strategy to counteract oxidative stress since this pathway is extremely efficient in generating the NADPH required for the function of several antioxidant responses [168,169].
In addition to providing adaptation to several environmental conditions, studies have shown that other carbon metabolism pathways of P. aeruginosa, such as the glyoxylate shunt (GS), also contribute to bacterial survival under oxidative stress [170,171]. GS is a two-step metabolic route composed of two enzymes (i.e., isocitrate lyase (aceA) and malate synthase (glcB)) and is an alternative to the tricarboxylic acid cycle (TCA). In this pathway, aerobic bacteria metabolize acetate and fatty acids as carbon sources [172]. An example of this kind of environment is the CF sputum, where the concentration of fatty acid is exceptionally high [171].
The P. aeruginosa and E. coli isocitrate lyase have 27% identity, while P. aeruginosa malate synthase shares 59% sequence identity with E. coli malate synthase. In addition to being upregulated by the excess of fatty acid and acetate, aceA expression is induced under H2O2 exposure and iron limitation. At the same time, glcB is overproduced by redox-active compounds and antibiotics exposure. GS was shown to regulate the intracellular amount of iron, and the susceptibility of GS mutants to oxidative stress could be due to the interaction of H2O2 with intracellular iron [171]. Ahn and colleagues (2017) showed that aceA is activated in response to oxidative stress, in which the overproduction of the efflux pump mexE could contribute to the bacterial response to these toxic agents. Furthermore, increased EPS production, biofilm formation, and pyoverdine and pyocyanin synthesis were observed in the aceA mutant [170].
Although the molecular mechanisms behind their roles in the oxidative stress response have not been completely elucidated, other proteins involved in carbon metabolism have also been described to be involved in bacterial survival under oxidizing conditions. For instance, enolase, an enzyme involved in carbon metabolism and an important component of the E. coli RNA degradosome [173], was shown to possesses roles in P. aeruginosa growth and virulence in a murine acute pneumonia model. It was also involved in oxidative stress tolerance by affecting the production of ahpB and ahpC in an OxyR-independent manner [174]. In another study, the authors showed that Crc, a regulator that controls carbon catabolite, participates in the oxidative stress response in P. aeruginosa by controlling the metabolic flux through NADP+-dependent dehydrogenases and maintaining the P. aeruginosa metabolism [175].
3.2.7. Post-Transcriptional Modification
Several RNA transcripts undergo post-transcriptional modification to become a mature and functional molecule, contributing to a more effective translation of RNA into protein [176]. Studies have reported the roles of tRNA methylation, RNases, and small RNAs on oxidative stress response genes. The upregulation of trmB, a gene that encodes a tRNA guanine-N7-methyltransferase that catalyzes 7-Methylguanosine (m7G) modification in tRNA, increases the amounts of m7G modified tRNA, which improves the translation efficiency of Phe and Asp enriched mRNAs, such as katA and katB [177]. Jaroensuk and colleagues (2016) also showed that the methyltransferase TrmJ also plays a role in oxidative stress response by regulating the expression of oxyR-recG, katB, and katE [178].
The endoribonuclease YbeY is involved in ribosome quality control and maturation and post-transcriptional regulation of gene expression [179]. It is also required for bacterial survival in a murine acute pneumonia model. This endoribonuclease has also been shown to play important roles in the oxidative stress response. A mutation in the P. aeruginosa yebY gene reduced the expression of katA and katB, and this mutant also presented higher sensitivity to neutrophiles exposure than the wild-type. YbeY controls the expression of katA through the sigma factor RpoS, in which YbeY affects the rpoS translation by upregulating ReaL, a small RNA (sRNA) that represses rpoS translation. Furthermore, YebY interacts with RpoS and promotes its degradation [180]. The roles of sRNAs on oxidative stress were also shown by Gómez-Lozano et al. (2014). In this study, the authors demonstrated that several antisense small RNAs were differentially expressed under different experimental conditions, including osmotic pressure, oxidative stress, and antibiotic treatment, suggesting that they may play regulatory roles in P. aeruginosa cells [181].
3.2.8. Other Stress Response Mechanisms
As the study of oxidative stress responses adopted by bacteria progresses, several proteins known to be involved in specific pathways have also been recognized for being employed by bacteria as a survival strategy. In P. aeruginosa, several findings have been made in this direction. For instance, studies have shown that lipotoxin F (lptF) is involved in the oxidative stress response and adhesion to human lung epithelial cells, and its expression is stimulated by the conversion to the mucoid phenotype [155]. Furthermore, acyl carrier proteins [182], flavohemoprotein [183], PppA-PpkA kinase [184], flavodoxins [185], and the phnW pyruvate aminotransferase [186], which are involved in fatty acid biosynthesis, nitric-oxide detoxification, protein phosphorylation, electron transfer, and phosphonate degradation, respectively, have also been shown to play important roles in adaptive response and P. aeruginosa survival under oxidizing conditions.
Oxidative stress has also been shown to induce other phenotypic changes in P. aeruginosa, mainly in chronic infection where the immune system and therapeutic agents impose adverse conditions to the survival of pathogens [187]. In this context, resistance to antibiotics, biofilm formation, and mucoid and hypermutable phenotypes play important roles in P. aeruginosa survival under oxidizing environments [187,188]. For instance, the presence of hypermutable P. aeruginosa in the lungs of CF patients was reported to be associated with antibiotic resistance and increased time of infection. This hypermutable phenotype was provoked by DNA mutations induced by oxidizing agents [189]. Furthermore, double-strand breaks caused by oxidative stress in P. aeruginosa biofilms were shown to induce several DNA repair systems, generating mutations that promote the high adaptability and diversity of these communities [190].
3.3. Transcriptional Regulators Involved in P. aeruginosa Oxidative Stress Responses
3.3.1. OxyR: The Master Regulator of Oxidative Stress
OxyR is a 34 kDa H2O2-sensing transcriptional regulator that belongs to the LTTR family (LysR Type Transcriptional Regulators). It is composed of four different domains (i.e., DNA binding domain, H2O2 sensing, transactivation, and tetramerization) [191] and is commonly found in almost all Gram-negative bacteria [192,193,194]. It is also present in some Gram-positive species, such as Streptomyces avermitilis [195] and Staphylococcus aureus [196]. OxyR is the central, most frequently studied, and the best-characterized transcriptional regulator involved in oxidative stress responses by the induction of the transcription of protective genes, such as catalases and alkyl hydroperoxides [197]. In P. aeruginosa, most of the genes involved in response to H2O2 and organic peroxides are controlled by OxyR [192].
The activation of OxyR in P. aeruginosa is done by the oxidation of Cys199 and Cys208. Furthermore, a third cysteine residue (Cys296) might also be involved in OxyR activation. This cysteine residue is not conserved in E. coli and is found only in a few beta-proteobacteria. In addition, P. aeruginosa OxyR presents positive and negative regulation effects [191]. For instance, katA is positively and negatively regulated by OxyR, while katB is positively regulated [117].
In P. aeruginosa, a 34kDa OxyR transcriptional regulator induces the expression of one catalase and two alkyl hydroperoxide reductase (i.e., katB, ahpB, and ahpCF, respectively). It also plays important roles in DNA repair since it is located in an operon with the DNA helicase RecG [125]. In addition to the protection to H2O2, the oxyR-recG operon is also involved in P. aeruginosa protection against paraquat [125] and HOCl [109].
Recent studies have shown that P. aeruginosa OxyR regulates numerous genes involved in stress responses under oxidative stress. Wei and collaborators (2012) demonstrated that OxyR also regulates ~122 genes in response to H2O2 stress [192]. It binds to the promoter of several genes and regulates the expression of those involved in H2O2 detoxification (e.g., katA, katB, ahpB, and ahpCF), iron metabolism (e.g., dps and pvdS), biofilm formation (e.g., pf4 and bdlA), protein synthesis (e.g., rpsL, cca, aspS and alsS), cell wall synthesis (e.g., lpxC), QS (e.g., rsaL), aerobic and anaerobic respiration (e.g., snr1 and cyoA), the transport of small molecules (e.g., PA1541 and PA0185), OxyR reduction (e.g., trxA and trxB), and other regulatory genes (e.g., sRNA). Overall, OxyR represses protein synthesis and oxidative respiration and reduces the expression of iron uptake genes, indicating that bacteria reduce their metabolic activity as a response to oxidative stress. On the other hand, it induces detoxifying defenses, biofilm formation, and the production of sulfur-containing molecules. Detoxifying enzymes and biofilm formation are known to be an important protection for bacterial cells under oxidative conditions. Furthermore, these authors showed that P. aeruginosa OxyR binds to the promoter region of a thioredoxin system (trxA and trxB2), inducing its expression and at the same time also reduces the oxidized OxyR when NAD(P)H is available, indicating an auto-regulatory mechanism [192].
P. aeruginosa OxyR was also shown to affect other phenotypes. For instance, OxyR mutants were unable to swarm on agar plates due to the deficiency in rhamnolipids production. They also presented an increased pyocyanin production, suggesting the roles of this transcriptional regulator in bacterial virulence [198]. In addition to that, the inactivation of the P. aeruginosa OxyR abolished the utilization of pyoverdine, impairing iron acquisition by the cells [199]. These studies show that OxyR is a regulator that plays an essential role in bacterial survival and adaptation by regulating several genes involved in the detoxification of toxic oxygen species, DNA repair, resistance, and virulence.
3.3.2. OhrR and OspR
OhrR (organic peroxide-sensing repressor) is a transcriptional repressor widely found in Gram-positive and Gram-negative bacteria that belongs to the MarR family and controls the transcription of ohr (organic hydroperoxide resistance) [200,201]. P. aeruginosa OhrR acts as a repressor of the expression of ohr and ohrR by biding to their promoter regions. Under oxidative conditions, the formation of a disulfide bond between Cys19, the sensing cysteine, and Cys121 promotes a conformational change in the structure of OhrR, which provokes its dissociation from the promoter and allows the binding of RNA polymerase [202].
Studies have shown the protective effect of OhrR in P. aeruginosa under organic hydroperoxide conditions [202,203]. A P. aeruginosa ohr mutant was susceptible to t-butyl hydroperoxide and cumene hydroperoxide but not to paraquat and H2O2, suggesting that Ohr is a protective protein against organic hydroperoxide. Although the function of Ohr is similar to the function of AhpC-AhpF, its expression is OxyR-independent [204].
OspR (oxidative stress response and pigment production regulator) is a transcriptional regulator homolog to P. aeruginosa OhrR (46.5% identity) [203]. P. aeruginosa OspR is part of the oxidative stress response in this bacterium by the production of a glutathione peroxidase (gpx), which is a protective protein that decomposes peroxides using glutathione [205]. OspR and gpx are strongly induced by organic and lipid hydroperoxides and confer resistance to these compounds and H2O2 [203].
The effect of OspR occurs by its binding to the promoter region of the ospR-gpx operon, which consequently represses the transcription of gpx and ospR. In addition to its roles in oxidative protection of P. aeruginosa cells, OspR is also involved in several cellular pathways, including pigment production, QS, resistance to β-lactam antibiotics in a gpx-independent manner, and virulence in mice [205].
OspR and OhrR are two organic hydroperoxide regulators in P. aeruginosa that belong to the two-cysteine subfamily of the OhrR regulators. Similar to OhrR that has a sensing cysteine (Cys19), which plays essential roles in sensing oxidative stress, Cys24 plays essential roles in the protective effect of OspR [203,205]. Atichartpongkul and colleagues (2016) showed that OspR and OhrR have overlapping roles in the oxidative defense of P. aeruginosa. They demonstrated that OspR binds to the ohr promoter under oxidative stress and regulates its transcription, and OhrR binds to the gpx promoter, however, with lower affinity. Therefore, the ability of OspR to bind to the two promoters (ohr and gpx) highlights its broader protective effect and suggests that the oxidative protection conferred by OhrR and OspR is interconnected [203].
3.3.3. IscR
IscR (iron–sulfur cluster) is a transcriptional regulator that belongs to the Rrf2 family and regulates the expression of genes involved in iron–sulfur ([Fe–S]) cluster biosynthesis. [Fe–S] cluster proteins play essential roles in several cellular processes, including respiration, gene regulation, DNA repair, electron storage, and central metabolism [206,207,208]. In P. aeruginosa only the isc system encoded by the iscRSUA-hscBA-fdx2-iscX operon has been identified [110]. IscR recognizes two different binding motifs: type 1, which is recognized by holo-IscR, and type 2, recognized by both holo- and apo-IscR [209].
IscR has been described to be involved in the stress response of P. aeruginosa to oxidative stress [109,207,210] and bacterial virulence [210]. iscR mutants presented lower KatA, SodA, and SodB production than the wild-type [207,210], and this gene was required for the activity of KatA at the post-translational level [210]. Since the inactivation of iscR provoked a reduction in intracellular iron levels, affecting several iron-dependent pathways [110], and these enzymes require iron to function, the depletion of IscR explains their impaired activity and, consequently, the low tolerance against oxidizing agents.
In addition to its roles in protection against oxidative stress, IscR is also induced under different stress conditions that affect the availability of iron and damage the [Fe–S] cluster, such as iron and thiol depletion and high salt. The genes regulated by iscR that are important for the oxidative stress response have not been completely described [110]; however, a few of them have been studied recently. For instance, fprB, a gene that encodes a ferredoxin NADP(+) reductase, and nfuA, which encodes a [Fe–S] scaffolding protein involved in [Fe–S] cluster biosynthesis, have been described to be regulated by P. aeruginosa IscR. These proteins were significantly induced by oxidative agents, such as cycling agents, peroxide, and organic hydroperoxides, and the respective mutants showed increased sensitivity to oxidative stress [211,212]. fprB is also essential for bacterial survival under osmotic pressure and metal stress [211]. Overall, the susceptibility to oxidative stress observed in these mutants might be associated with the loss of function of the [Fe–S] cluster-containing proteins and transcriptional regulators. In another study, Boonma and collaborators (2017) showed that the Lys-R family transcriptional regulator FinR was activated by NaOCl and paraquat and mediated the expression of fprA under these stressful conditions [213]. Together, these results highlight the role of fprA in maintaining iron homeostasis and bacterial survival under oxidative stress conditions.
3.3.4. Fur
The Ferric uptake regulator (Fur) represses the expression of iron uptake genes when enough iron is available to the cells. It was shown that, in P. aeruginosa, Fur regulates the production of exotoxin A and siderophores [214,215]. In solution, P. aeruginosa Fur is a tetramer and forms dimers in the presence of DNA during its activation. The tetramer form of P. aeruginosa Fur is inactive and is activated by its dissociation into dimers and metal binding [216].
Fur has been described to positively activate P. aeruginosa katA and sodB [217]. Furthermore, Pasqua and collaborators (2017) showed that P. aeruginosa Fur mutants present impaired growth in solid media due to pyochelin production inhibition. However, the mutants did not show impaired biofilm formation and pathogenicity in a Galleria mellonella model, suggesting that Fur is not required for biofilm formation and bacteria pathogenicity in this in vivo model [218].
3.3.5. SoxR
SoxR is a 17kDa transcriptional regulator of the MerR family that controls O2− and NO responses. P. aeruginosa does not have the transcriptional regulator SoxS [219], which is frequently activated by SoxR and induces the transcription of several genes [220,221,222]. Furthermore, while SoxR and OxyR are often considered the primary sensors that control stress responses under oxidative conditions [223], P. aeruginosa SoxR is not essential for the oxidative stress response and antibiotic resistance [224].
Palma et al. (2005) [224] demonstrated that, under paraquat stress, P. aeruginosa SoxR activates six genes that are in three transcriptional subunits by binding to their promoter region. However, it was not activated by H2O2. The three subunits comprise the following genes: (i) PA3718, which encodes an efflux pump of the multiple facilitator superfamily; (ii) PA2274, which is an unknown protein with similarity to the monooxygenase and that has detoxification function; and (iii) four genes in the mexGHI-ompD operon, encoding the multidrug efflux pump of the resistance-nodulation-division superfamily [224]. SoxR might also be involved in P. aeruginosa virulence through the expression of the MexGHI-OmpD system since this efflux pump plays roles in QS [225,226]. In addition, P. aeruginosa SoxR is required for bacterial virulence in mice with pulmonary infection, suggesting the importance of this regulator in P. aeruginosa infections in the host due to the high production of superoxide in the lungs [224].
In addition to responding to paraquat [222,224], P. aeruginosa SoxR is also activated by other redox-active compounds, including pyocyanin, toxoflavin, phenazine, methosulphate, plumbagin, menadione, and menadione sodium bisulfite, indicating that this transcriptional regulator is also involved in the response to several oxidants. Furthermore, its activation by pyocyanin implies that it is also activated by endogenous metabolites, showing that SoxR in P. aeruginosa is involved in other cellular processes [222]. In this regard, Dietrich and colleagues (2006) showed that the expression of mexGHI-ompD, PA2274, and PA3718 was induced by pyocyanin, reinforcing that the activation of SoxR is a protective mechanism against not only paraquat but also to the endogenous production of pyocyanin. The activation of SoxR by pyocyanin occurs under anaerobic conditions, in which there is no formation of superoxide, and in a superoxide-independent manner [226].
3.3.6. Other Transcriptional Regulators
In addition to the transcriptional regulators described in this section, P. aeruginosa stress response studies have also identified other regulatory proteins involved in oxidative stress response. Among them, PA2206 is a novel transcriptional regulator that, along with OxyR, belongs to the LysR-type transcriptional regulators (LTTRs). PA2206 mutant was significantly less tolerant to exogenous oxidative stress and less lethal in a zebrafish model than the wild-type. Furthermore, the oxidative stress response orchestrated by PA2206 is OxyR-independent and involves the direct regulation of PA2214-15, pvds, and PA4881 (gene with unknown function) and approximately 56 other genes, including polyamine metabolism and iron genes [227]. PqrR, a repressor of the pqrABC operon [228], and AnvM (anaerobic and virulence modulator) [229] are also involved in oxidative stress response. PqrR and PqrA were also shown to be induced by H2O2, confirming its role in bacterial survival under peroxide stress [102]. AnvM regulates several genes involved in different pathways, such as oxidoreductase activity, transcription regulation, motility, the response to oxidative stress in the QS system, and pathogenicity [229].
3.4. Oxidative Stress Response Contributes to Antibiotic Resistance of P. aeruginosa
P. aeruginosa is intrinsically resistant to several classes of antibiotics, mainly due to low outer membrane permeability and the expression of several efflux pumps [15]. Bacterial efflux systems are classified into five families: resistance-nodulation-division (RND) family, major facilitator superfamily (MFS), ATP binding cassette (ABC) superfamily, small multidrug resistance (SMR), and multidrug and toxic compound extrusion (MATE) family. Of these, the RDN family is most commonly associated with antibiotic resistance in P. aeruginosa [15,230].
Many of the stress responses elicited by bacteria to counteract the damage caused by oxidative agents contribute to bacterial virulence and resistance, enhancing their survival in the host and the environment. In this context, the oxidative-induced expression of efflux systems has been linked to antibiotic resistance in P. aeruginosa [231,232]. MexR, a member of the MarR family that negatively regulates the expression of the efflux system mexAB-oprM and mexR, is activated upon peroxide oxidation by the formation of a disulfide bond between Cys-30 and Cys-62, which induces a conformational change in the protein and provokes its dissociation from the DNA promoter of the mexAB-oprM operon [231]. mexXY expression depends on the gene PA5471 (armZ) and is involved in the synthesis of the MexXY-OprM system, contributing to aminoglycoside resistance [232]. Nitrosative stress and pentachlorophenol exposure also induced the expression of efflux systems (i.e., mexEF-oprN and mexAB-oprM, respectively) [233,234].
Moreover, P. aeruginosa cells exposed to oxidative stress are often unable to repair all the DNA damage, which increases the mutation frequency in the bacteria, and induces antibiotic resistance mainly due to the increased production of β-lactamase and increased activity of the efflux system MexCD-OprJ [235]. MexAB-OprM, MexXY-OprM, and MexCD-OprJ are important efflux proteins associated with antibiotic resistance in P. aeruginosa [15]. They confer resistance to several classes of antibiotics, such as β -lactams, quinolones, tetracycline, and macrolides [15,236,237]. MexXY-OprM is the only system that has been shown to pump aminoglycosides [15,236]. Therefore, oxidative stress acts as a signal for the expression of the efflux system, which contributes to antibiotic resistance in bacteria.
In addition to the stimulation of efflux pumps, oxidative stress can also induce antibiotic resistance by other mechanisms. NaOCl provokes several damages to membrane structures, which favors the transfer of antibiotic-resistant genes from NaOCl-killed bacteria to other injured bacteria in that environment [238]. Furthermore, the induction of biofilm formation is also considered an adaptive response of bacteria against oxidative stress (Reviewed in [32]). Strempel and collaborators (2017) showed that NaOCl induced the expression of cyclic-di-GMP, a second messenger involved in the transition from planktonic to sessile lifestyle, by inducing the diguanylate synthase PA3177, which, therefore, induced biofilm formation [239,240]. This gene was also associated with antimicrobial resistance in P. aeruginosa biofilms [241].
Studies have shown that the enzymes that are involved in the oxidative stress response also contribute to antibiotic resistance. In this context, Xia and colleagues (2019) showed that oligoribonuclease (Orn) is required for bacterial resistance to aminoglycoside and β-lactam antibiotics and tolerance to oxidative stress by the regulation of the expression of katA at the post-transcriptional level [242]. Orn also contributes to ciprofloxacin tolerance by the overexpression of pyocin [243]. Orn is an exonuclease that hydrolyzes nanoRNAs, which are very short RNAs that contain up to four nucleotides and can be used as primers in the initiation of transcription, resulting in severe alteration in gene expression [244]. It was shown to be required for the expression of the T3SS and bacterial virulence in a mouse acute pneumonia model [245]. Furthermore, Orn also plays essential roles in biofilm formation by inducing the intracellular accumulation of c-di-GMP [246].
AmpR, a major LTTR transcriptional regulator that controls the expression of the β-lactamase ampC [247], was shown to positively regulate QS genes, as well as several other genes involved in P. aeruginosa antibiotic resistance, extracellular products such as alginate and pyoverdine, and the central metabolism. AmpR also regulated small and antisense RNAs, iron uptake by regulating the siderophores pyoverdine and pyochelin expression, heat shock, and oxidative stress. In this regard, AmpR was shown to positively regulate oxidative stress genes, such as the katA, oxyR, and nuo genes [135].
Finally, the induction of a non-culturable state by the prolonged (36h) exposure of P. aeruginosa to H2O2 provoked a decrease in cell turbidity and cultivability of the bacteria [248]. This non-culturable state is also attributed to antibiotic resistance and a tolerance to harsh environmental conditions [32].
4. Final Remarks
The exceptional physiological capabilities of P. aeruginosa allow it to survive in harsh conditions and outgrow other bacteria in resource-limited environments [249]. Furthermore, the persistence and prevalence of P. aeruginosa in clinical settings are explained by the high metabolic capacity of this pathogen, which gives P. aeruginosa the ability to adapt to diverse environments [250]. For example, P. aeruginosa can metabolize several carbon sources and use different electron acceptors [251]. These factors, combined with its high resistance to several classes of antibiotics and disinfectants, increase the incidence and severity of P. aeruginosa infections, and therefore, the morbidity and mortality rates. It has become a public health concern, and in 2017, the World Health Organization classified this Gram-negative pathogen as group 1 priority for the development of new antimicrobials [252].
In its natural environment, P. aeruginosa is continuously exposed to oxidizing agents. Therefore, the survival and successful prevalence of P. aeruginosa in these environments are strictly associated with its ability to develop an arsenal of adaptive mechanisms to counteract the damage caused by these agents. Although several studies have been conducted to evaluate the adaptive repertoire developed by this pathogen to counteract oxidative stress, the complexity and robustness of these responses are still in their infancy, mainly because many of these mechanisms are interconnected with other cell processes. For instance, studies have shown that carbon metabolism pathways, post-transcriptional modifications, quorum sensing, and pigment production also play roles in protecting P. aeruginosa against oxidative stress.
Among the oxidative stress responses described in P. aeruginosa, the expression of detoxifying enzymes, mainly catalases and alkyl hydroperoxides, can be considered one of the most important and prevalent responses under oxidative conditions since their synthesis is controlled by several systems, such as QS and diverse transcriptional regulators. Among the main transcriptional regulators, OxyR is considered the major regulator of oxidative stress responses and has also been shown to regulate genes involved in bacterial virulence. Furthermore, other regulators, including OhrR, OspR, and IscR, have also been shown to possess important roles in defending P. aeruginosa against the irreversible damage caused by oxidizing agents. Recently, many researchers have concentrated efforts on studying regulatory pathways and transcriptional regulators involved in P. aeruginosa’s oxidative stress responses. However, considering that a large amount of its genome is composed of transcriptional regulators, for example, for PAO1, approximately 10% of its genome is composed of these proteins [99], there are still many adaptive mechanisms to be elucidated.
Understanding the defense mechanisms adopted by this model pathogen would shed light on the development and improvement of new therapeutic options to treat P. aeruginosa infections, in which the pathways employed to resist oxidative stress could be targeted by antimicrobials. For instance, azithromycin and macrolides have been shown to downregulate the expression of lipotoxins, such as LptF, and alter biofilms, respectively [253,254]. Furthermore, the glyoxylate shunt, which is not present in humans [255], can also be a potential target for developing new antibiotics. Since these structures are also used as stress responses against oxidative stress, the effectiveness of these antibiotics could be enhanced when treating lung infections of CF patients, in which a high concentration of oxidizing agents is produced, and P. aeruginosa is frequently the causative agent. Furthermore, since many of the stress responses are interconnected to virulence factors, such as the upregulation of pigments and conversion to a mucoid phenotype, the elucidation of these mechanisms could improve vaccine and drug development by compromising the virulence potential of P. aeruginosa [104].
Author Contributions
W.S.d.C.N., V.I., Z.V., N.S., E.C. and J.O. designed, wrote, and revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Carleton University and the Natural Sciences and Engineering Research Council of Canada (NSERC, RGPIN-2019-06335).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
All figures were created with BioRender.com (accessed on 10 September 2021).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Heimesaat, M.M.; Escher, U.; Grunau, A.; Kühl, A.A.; Bereswill, S. Multidrug-Resistant Pseudomonas aeruginosa Accelerate Intestinal, Extra-Intestinal, and Systemic Inflammatory Responses in Human Microbiota-Associated Mice With Subacute Ileitis. Front. Immunol. 2019, 10, 49. [Google Scholar] [CrossRef]
- Mohamed, F.A.; Shaker, G.H.; Askoura, M.M. Oxidative Stress Influences Pseudomonas aeruginosa Susceptibility to Antibiotics and Reduces Its Pathogenesis in Host. Curr. Microbiol. 2020, 77, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Sadikot, R.T.; Blackwell, T.S.; Christman, J.W.; Prince, A.S. Pathogen–Host Interactions in Pseudomonas aeruginosa Pneumonia. Am. J. Respir. Crit. Care Med. 2005, 171, 1209–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nixon, G.M.; Armstrong, D.S.; Carzino, R.; Carlin, J.B.; Olinsky, A.; Robertson, C.F.; Grimwood, K. Clinical Outcome after Early Pseudomonas aeruginosa Infection in Cystic Fibrosis. J. Pediatrics 2001, 138, 699–704. [Google Scholar] [CrossRef] [PubMed]
- Harrison, F. Microbial Ecology of the Cystic Fibrosis Lung. Microbiology 2007, 153, 917–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parkins, M.D.; Somayaji, R.; Waters, V.J. Epidemiology, Biology, and Impact of Clonal Pseudomonas aeruginosa Infections in Cystic Fibrosis. Clin. Microbiol. Rev. 2018, 31, e00019-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassetti, M.; Vena, A.; Croxatto, A.; Righi, E.; Guery, B. How to Manage Pseudomonas aeruginosa Infections. Drugs Context 2018, 7, 1–18. [Google Scholar] [CrossRef]
- Horcajada, J.P.; Montero, M.; Oliver, A.; Sorlí, L.; Luque, S.; Gómez-Zorrilla, S.; Benito, N.; Grau, S. Epidemiology and Treatment of Multidrug-Resistant and Extensively Drug-Resistant Pseudomonas aeruginosa Infections. Clin. Microbiol. Rev. 2019, 32, e00031-19. [Google Scholar] [CrossRef]
- Diggle, S.P.; Whiteley, M. Microbe Profile: Pseudomonas aeruginosa: Opportunistic Pathogen and Lab Rat. Microbiology 2020, 166, 30–33. [Google Scholar] [CrossRef]
- Mao, G.; Song, Y.; Bartlam, M.; Wang, Y. Long-Term Effects of Residual Chlorine on Pseudomonas aeruginosa in Simulated Drinking Water Fed With Low AOC Medium. Front. Microbiol. 2018, 9, 879. [Google Scholar] [CrossRef]
- Breidenstein, E.B.M.; de la Fuente-Núñez, C.; Hancock, R.E.W. Pseudomonas aeruginosa: All Roads Lead to Resistance. Trends Microbiol. 2011, 19, 419–426. [Google Scholar] [CrossRef]
- Poole, K. Pseudomonas aeruginosa: Resistance to the Max. Front. Microbiol. 2011, 2, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breidenstein, E.B.M.; Janot, L.; Strehmel, J.; Fernandez, L.; Taylor, P.K.; Kukavica-Ibrulj, I.; Gellatly, S.L.; Levesque, R.C.; Overhage, J.; Hancock, R.E.W. The Lon Protease Is Essential for Full Virulence in Pseudomonas aeruginosa. PLoS ONE 2012, 7, e49123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, L.; Jiang, T.; Zhang, S.; Yu, X. Exposure to Mutagenic Disinfection Byproducts Leads to Increase of Antibiotic Resistance in Pseudomonas aeruginosa. Environ. Sci. Technol. 2014, 48, 8188–8195. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.-J.; Cheng, Z. Antibiotic Resistance in Pseudomonas aeruginosa: Mechanisms and Alternative Therapeutic Strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef]
- O’Toole, G.A.; Kolter, R. Flagellar and Twitching Motility Are Necessary for Pseudomonas aeruginosa Biofilm Development. Mol. Microbiol. 1998, 30, 295–304. [Google Scholar] [CrossRef]
- Barken, K.B.; Pamp, S.J.; Yang, L.; Gjermansen, M.; Bertrand, J.J.; Klausen, M.; Givskov, M.; Whitchurch, C.B.; Engel, J.N.; Tolker-Nielsen, T. Roles of Type IV Pili, Flagellum-Mediated Motility and Extracellular DNA in the Formation of Mature Multicellular Structures in Pseudomonas aeruginosa Biofilms. Environ. Microbiol. 2008, 10, 2331–2343. [Google Scholar] [CrossRef]
- Persat, A.; Inclan, Y.F.; Engel, J.N.; Stone, H.A.; Gitai, Z. Type IV Pili Mechanochemically Regulate Virulence Factors in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2015, 112, 7563–7568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauser, A.R. The Type III Secretion System of Pseudomonas aeruginosa: Infection by Injection. Nat. Rev. Microbiol. 2009, 7, 654–665. [Google Scholar] [CrossRef] [Green Version]
- Williams, B.J.; Dehnbostel, J.; Blackwell, T.S. Pseudomonas aeruginosa: Host Defence in Lung Diseases. Respirology 2010, 15, 1037–1056. [Google Scholar] [CrossRef]
- Blier, A.-S.; Vieillard, J.; Gerault, E.; Dagorn, A.; Varacavoudin, T.; Le Derf, F.; Orange, N.; Feuilloley, M.; Lesouhaitier, O. Quantification of Pseudomonas aeruginosa Hydrogen Cyanide Production by a Polarographic Approach. J. Microbiol. Methods 2012, 90, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Gellatly, S.L.; Hancock, R.E.W. Pseudomonas aeruginosa: New Insights into Pathogenesis and Host Defenses. Pathog. Dis. 2013, 67, 159–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pritt, B.; O’Brien, L.; Winn, W. Mucoid Pseudomonas in Cystic Fibrosis. Am. J. Clin. Pathol. 2007, 128, 32–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, A.; Anraku, M.; Nakagawa, S.; Ojima, T. Discovery of a Novel Alginate Lyase from Nitratiruptor Sp. SB155-2 Thriving at Deep-Sea Hydrothermal Vents and Identification of the Residues Responsible for Its Heat Stability. J. Biol. Chem. 2016, 291, 15551–15563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, J.D.; Kocíncová, D.; Westman, E.L.; Lam, J.S. Review: Lipopolysaccharide Biosynthesis in Pseudomonas aeruginosa. Innate Immun. 2009, 15, 261–312. [Google Scholar] [CrossRef]
- Hall, S.; McDermott, C.; Anoopkumar-Dukie, S.; McFarland, A.J.; Forbes, A.; Perkins, A.V.; Davey, A.K.; Chess-Williams, R.; Kiefel, M.J.; Arora, D.; et al. Cellular Effects of Pyocyanin, a Secreted Virulence Factor of Pseudomonas aeruginosa. Toxins 2016, 8, 236. [Google Scholar] [CrossRef]
- Kim, D.; Sitepu, I.R.; Hashidoko, Y. Induction of Biofilm Formation in the Betaproteobacterium Burkholderia Unamae CK43B Exposed to Exogenous Indole and Gallic Acid. Appl. Environ. Microbiol. 2013, 79, 4845–4852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jefferson, K.K. What Drives Bacteria to Produce a Biofilm? FEMS Microbiol. Lett. 2004, 236, 163–173. [Google Scholar] [CrossRef]
- Hoffman, L.R.; D’Argenio, D.A.; MacCoss, M.J.; Zhang, Z.; Jones, R.A.; Miller, S.I. Aminoglycoside Antibiotics Induce Bacterial Biofilm Formation. Nature 2005, 436, 1171–1175. [Google Scholar] [CrossRef]
- Linares, J.F.; Gustafsson, I.; Baquero, F.; Martinez, J.L. Antibiotics as Intermicrobial Signaling Agents Instead of Weapons. Proc. Natl. Acad. Sci. USA 2006, 103, 19484–19489. [Google Scholar] [CrossRef] [Green Version]
- Bagge, N.; Schuster, M.; Hentzer, M.; Ciofu, O.; Givskov, M.; Greenberg, E.P.; Høiby, N. Pseudomonas aeruginosa Biofilms Exposed to Imipenem Exhibit Changes in Global Gene Expression and Beta-Lactamase and Alginate Production. Antimicrob. Agents Chemother. 2004, 48, 1175–1187. [Google Scholar] [CrossRef] [Green Version]
- Da Cruz Nizer, W.S.; Inkovskiy, V.; Overhage, J. Surviving Reactive Chlorine Stress: Responses of Gram-Negative Bacteria to Hypochlorous Acid. Microorganisms 2020, 8, 1220. [Google Scholar] [CrossRef]
- Klebensberger, J.; Birkenmaier, A.; Geffers, R.; Kjelleberg, S.; Philipp, B. SiaA and SiaD Are Essential for Inducing Autoaggregation as a Specific Response to Detergent Stress in Pseudomonas aeruginosa. Environ. Microbiol. 2009, 11, 3073–3086. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, J.B. Antibiotic-Induced Biofilm Formation. Int. J. Artif. Organs 2011, 34, 737–751. [Google Scholar] [CrossRef] [PubMed]
- Odenholt, I. Pharmacodynamic Effects of Subinhibitory Antibiotic Concentrations. Int. J. Antimicrob. Agents 2001, 17, 1–8. [Google Scholar] [CrossRef]
- Langendonk, R.F.; Neill, D.R.; Fothergill, J.L. The Building Blocks of Antimicrobial Resistance in Pseudomonas aeruginosa: Implications for Current Resistance-Breaking Therapies. Front. Cell. Infect. Microbiol. 2021, 11, 665759. [Google Scholar] [CrossRef] [PubMed]
- Finnegan, M.; Linley, E.; Denyer, S.P.; McDonnell, G.; Simons, C.; Maillard, J.-Y. Mode of Action of Hydrogen Peroxide and Other Oxidizing Agents: Differences between Liquid and Gas Forms. J. Antimicrob. Chemother. 2010, 65, 2108–2115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imlay, J.A. Where in the World Do Bacteria Experience Oxidative Stress? Environ. Microbiol. 2019, 21, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhao, X. Contribution of Oxidative Damage to Antimicrobial Lethality. Antimicrob. Agents Chemother. 2009, 53, 1395–1402. [Google Scholar] [CrossRef] [Green Version]
- Imlay, J.A. The Molecular Mechanisms and Physiological Consequences of Oxidative Stress: Lessons from a Model Bacterium. Nat. Rev. Microbiol. 2013, 11, 443–454. [Google Scholar] [CrossRef] [Green Version]
- Ezraty, B.; Gennaris, A.; Barras, F.; Collet, J.-F. Oxidative Stress, Protein Damage and Repair in Bacteria. Nat. Rev. Microbiol. 2017, 15, 385–396. [Google Scholar] [CrossRef]
- Imlay, J.A. A Metabolic Enzyme That Rapidly Produces Superoxide, Fumarate Reductase of Escherichia coli(∗). J. Biol. Chem. 1995, 270, 19767–19777. [Google Scholar] [CrossRef]
- Korshunov, S.; Imlay, J.A. Two Sources of Endogenous Hydrogen Peroxide in Escherichia coli. Mol. Microbiol. 2010, 75, 1389–1401. [Google Scholar] [CrossRef] [Green Version]
- Seaver, L.C.; Imlay, J.A. Are Respiratory Enzymes the Primary Sources of Intracellular Hydrogen Peroxide?*. J. Biol. Chem. 2004, 279, 48742–48750. [Google Scholar] [CrossRef] [Green Version]
- Storz, G.; Imlay, J.A. Oxidative Stress. Curr. Opin. Microbiol. 1999, 2, 188–194. [Google Scholar] [CrossRef]
- Dwyer, D.J.; Collins, J.J.; Walker, G.C. Unraveling the Physiological Complexities of Antibiotic Lethality. Annu. Rev. Pharmacol. Toxicol. 2015, 55, 313–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeom, J.; Imlay, J.A.; Park, W. Iron Homeostasis Affects Antibiotic-Mediated Cell Death in Pseudomonas Species*. J. Biol. Chem. 2010, 285, 22689–22695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohanski, M.A.; DePristo, M.A.; Collins, J.J. Sublethal Antibiotic Treatment Leads to Multidrug Resistance via Radical-Induced Mutagenesis. Mol. Cell 2010, 37, 311–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foti, J.J.; Devadoss, B.; Winkler, J.A.; Collins, J.J.; Walker, G.C. Oxidation of the Guanine Nucleotide Pool Underlies Cell Death by Bactericidal Antibiotics. Science 2012, 336, 315–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Root, R.K.; Metcalf, J.; Oshino, N.; Chance, B. H2O2 Release from Human Granulocytes during Phagocytosis. I. Documentation, Quantitation, and Some Regulating Factors. J. Clin. Investig. 1975, 55, 945–955. [Google Scholar] [CrossRef] [Green Version]
- Babior, B.M.; Kipnes, R.S.; Curnutte, J.T. Biological Defense Mechanisms. The production by leukocytes of superoxide, a potential bactericidal agent. J. Clin. Investig. 1973, 52, 741–744. [Google Scholar] [CrossRef]
- Murdoch, L.E.; Bailey, L.; Banham, E.; Watson, F.; Adams, N.M.T.; Chewins, J. Evaluating Different Concentrations of Hydrogen Peroxide in an Automated Room Disinfection System. Lett. Appl. Microbiol. 2016, 63, 178–182. [Google Scholar] [CrossRef]
- Gray, M.J.; Wholey, W.-Y.; Jakob, U. Bacterial Responses to Reactive Chlorine Species. Annu. Rev. Microbiol. 2013, 67, 141–160. [Google Scholar] [CrossRef] [Green Version]
- Fukuzaki, S. Mechanisms of Actions of Sodium Hypochlorite in Cleaning and Disinfection Processes. Biocontrol Sci. 2006, 11, 147–157. [Google Scholar] [CrossRef]
- Rutala, W.A.; Weber, D.J. Uses of Inorganic Hypochlorite (Bleach) in Health-Care Facilities. Clin. Microbiol. Rev. 1997, 10, 597–610. [Google Scholar] [CrossRef]
- Sasai, M.; Yamamoto, M. Pathogen Recognition Receptors: Ligands and Signaling Pathways by Toll-Like Receptors. Int. Rev. Immunol. 2013, 32, 116–133. [Google Scholar] [CrossRef]
- Fujiwara, N.; Kobayashi, K. Macrophages in Inflammation. Curr. Drug Targets Inflamm. Allergy 2005, 4, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Amulic, B.; Cazalet, C.; Hayes, G.L.; Metzler, K.D.; Zychlinsky, A. Neutrophil Function: From Mechanisms to Disease. Annu. Rev. Immunol. 2012, 30, 459–489. [Google Scholar] [CrossRef] [PubMed]
- Underhill, D.M.; Ozinsky, A. Phagocytosis of Microbes: Complexity in Action. Annu. Rev. Immunol. 2002, 20, 825–852. [Google Scholar] [CrossRef] [PubMed]
- Flannagan, R.S.; Cosío, G.; Grinstein, S. Antimicrobial Mechanisms of Phagocytes and Bacterial Evasion Strategies. Nat. Rev. Microbiol. 2009, 7, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Rada, B. Interactions between Neutrophils and Pseudomonas aeruginosa in Cystic Fibrosis. Pathogens 2017, 6, 10. [Google Scholar] [CrossRef]
- Moser, C.; Jensen, P.Ø.; Thomsen, K.; Kolpen, M.; Rybtke, M.; Lauland, A.S.; Trøstrup, H.; Tolker-Nielsen, T. Immune Responses to Pseudomonas aeruginosa Biofilm Infections. Front. Immunol. 2021, 12, 237. [Google Scholar] [CrossRef]
- Versey, Z.; da Cruz Nizer, W.S.; Russell, E.; Zigic, S.; DeZeeuw, K.G.; Marek, J.E.; Overhage, J.; Cassol, E. Biofilm-Innate Immune Interface: Contribution to Chronic Wound Formation. Front. Immunol. 2021, 12, 648554. [Google Scholar] [CrossRef]
- Panday, A.; Sahoo, M.K.; Osorio, D.; Batra, S. NADPH Oxidases: An Overview from Structure to Innate Immunity-Associated Pathologies. Cell. Mol. Immunol. 2015, 12, 5–23. [Google Scholar] [CrossRef] [Green Version]
- Winterbourn, C.C.; Kettle, A.J.; Hampton, M.B. Reactive Oxygen Species and Neutrophil Function. Annu. Rev. Biochem. 2016, 85, 765–792. [Google Scholar] [CrossRef]
- Nguyen, G.T.; Green, E.R.; Mecsas, J. Neutrophils to the ROScue: Mechanisms of NADPH Oxidase Activation and Bacterial Resistance. Front. Cell. Infect. Microbiol. 2017, 7, 373. [Google Scholar] [CrossRef] [PubMed]
- West, A.P.; Brodsky, I.E.; Rahner, C.; Woo, D.K.; Erdjument-Bromage, H.; Tempst, P.; Walsh, M.C.; Choi, Y.; Shadel, G.S.; Ghosh, S. TLR Signalling Augments Macrophage Bactericidal Activity through Mitochondrial ROS. Nature 2011, 472, 476–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, A.P.; Shadel, G.S.; Ghosh, S. Mitochondria in Innate Immune Responses. Nat. Rev. Immunol. 2011, 11, 389–402. [Google Scholar] [CrossRef] [Green Version]
- Abuaita, B.H.; Schultz, T.L.; O’Riordan, M.X. Mitochondria-Derived Vesicles Deliver Antimicrobial Reactive Oxygen Species to Control Phagosome-Localized Staphylococcus aureus. Cell Host Microbe 2018, 24, 625–636.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Painter, R.G.; Wang, G. Direct Measurement of Free Chloride Concentrations in the Phagolysosomes of Human Neutrophils. Anal. Chem. 2006, 78, 3133–3137. [Google Scholar] [CrossRef]
- Klebanoff, S.J.; Kettle, A.J.; Rosen, H.; Winterbourn, C.C.; Nauseef, W.M. Myeloperoxidase: A Front-Line Defender against Phagocytosed Microorganisms. J. Leukoc. Biol. 2013, 93, 185–198. [Google Scholar] [CrossRef] [Green Version]
- Winterbourn, C.C.; Hampton, M.B.; Livesey, J.H.; Kettle, A.J. Modeling the Reactions of Superoxide and Myeloperoxidase in the Neutrophil Phagosome: Implications for Microbial Killing. J. Biol. Chem. 2006, 281, 39860–39869. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Q.; Griffin, D.A.; Barofsky, D.F.; Hurst, J.K. Intraphagosomal Chlorination Dynamics and Yields Determined Using Unique Fluorescent Bacterial Mimics. Chem. Res. Toxicol. 1997, 10, 1080–1089. [Google Scholar] [CrossRef]
- Winterbourn, C.C.; Kettle, A.J. Redox Reactions and Microbial Killing in the Neutrophil Phagosome. Antioxid. Redox Signal. 2013, 18, 642–660. [Google Scholar] [CrossRef]
- Weiss, S.J. Tissue Destruction by Neutrophils. N. Engl. J. Med. 1989, 320, 365–376. [Google Scholar] [CrossRef]
- Allen, R.C.; Stephens, J.T. Myeloperoxidase Selectively Binds and Selectively Kills Microbes. Infect. Immun. 2011, 79, 474–485. [Google Scholar] [CrossRef] [Green Version]
- Dukan, S.; Belkin, S.; Touati, D. Reactive Oxygen Species Are Partially Involved in the Bacteriocidal Action of Hypochlorous Acid. Arch. Biochem. Biophys. 1999, 367, 311–316. [Google Scholar] [CrossRef]
- Nyström, T. Role of Oxidative Carbonylation in Protein Quality Control and Senescence. EMBO J. 2005, 24, 1311–1317. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C.C.; Hampton, M.B. Thiol Chemistry and Specificity in Redox Signaling. Free Radic. Biol. Med. 2008, 45, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P. Kinetics and Mechanisms of Thiol-Disulfide Exchange Covering Direct Substitution and Thiol Oxidation-Mediated Pathways. Antioxid. Redox Signal. 2012, 18, 1623–1641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, M.J. The Oxidative Environment and Protein Damage. Biochim. Biophys. Acta (BBA)—Proteins Proteom. 2005, 1703, 93–109. [Google Scholar] [CrossRef]
- Winter, J.; Ilbert, M.; Graf, P.C.F.; Ozcelik, D.; Jakob, U. Bleach Activates a Redox-Regulated Chaperone by Oxidative Protein Unfolding. Cell 2008, 135, 691–701. [Google Scholar] [CrossRef] [Green Version]
- Barrette, W.C.; Albrich, J.M.; Hurst, J.K. Hypochlorous Acid-Promoted Loss of Metabolic Energy in Escherichia coli. Infect. Immun. 1987, 55, 2518–2525. [Google Scholar] [CrossRef] [Green Version]
- Dukan, S.; Touati, D. Hypochlorous Acid Stress in Escherichia coli: Resistance, DNA Damage, and Comparison with Hydrogen Peroxide Stress. J. Bacteriol. 1996, 178, 6145–6150. [Google Scholar] [CrossRef] [Green Version]
- Albrich, J.M.; Gilbaugh, J.H.; Callahan, K.B.; Hurst, J.K. Effects of the Putative Neutrophil-Generated Toxin, Hypochlorous Acid, on Membrane Permeability and Transport Systems of Escherichia coli. J. Clin. Investig. 1986, 78, 177–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Candeias, L.P.; Steenken, S. Electron Transfer in Di(Deoxy)Nucleoside Phosphates in Aqueous Solution: Rapid Migration of Oxidative Damage (via Adenine) to Guanine. J. Am. Chem. Soc. 1993, 115, 2437–2440. [Google Scholar] [CrossRef]
- Giese, B. Electron Transfer in DNA. Curr. Opin. Chem. Biol. 2002, 6, 612–618. [Google Scholar] [CrossRef]
- Hogg, M.; Wallace, S.S.; Doublié, S. Bumps in the Road: How Replicative DNA Polymerases See DNA Damage. Curr. Opin. Struct. Biol. 2005, 15, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Touati, D.; Jacques, M.; Tardat, B.; Bouchard, L.; Despied, S. Lethal Oxidative Damage and Mutagenesis Are Generated by Iron in Delta Fur Mutants of Escherichia coli: Protective Role of Superoxide Dismutase. J. Bacteriol. 1995, 177, 2305–2314. [Google Scholar] [CrossRef] [Green Version]
- Cornelis, P.; Wei, Q.; Andrews, S.C.; Vinckx, T. Iron Homeostasis and Management of Oxidative Stress Response in Bacteria. Metallomics 2011, 3, 540–549. [Google Scholar] [CrossRef]
- Hassett, D.J.; Sokol, P.A.; Howell, M.L.; Ma, J.-F.; Schweizer, H.T.; Ochsner, U.; Vasil, M.L. Ferric Uptake Regulator (Fur) Mutants of Pseudomonas aeruginosa Demonstrate Defective Siderophore-Mediated Iron Uptake, Altered Aerobic Growth, and Decreased Superoxide Dismutase and Catalase Activities. J. Bacteriol. 1996, 178, 8. [Google Scholar] [CrossRef] [Green Version]
- Keyer, K.; Imlay, J.A. Superoxide Accelerates DNA Damage by Elevating Free-Iron Levels. Proc. Natl. Acad. Sci. USA 1996, 93, 13635–13640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawkins, C.L.; Davies, M.J. Hypochlorite-Induced Damage to DNA, RNA, and Polynucleotides: Formation of Chloramines and Nitrogen-Centered Radicals. Chem. Res. Toxicol. 2002, 15, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Kohen, R.; Nyska, A. Invited Review: Oxidation of Biological Systems: Oxidative Stress Phenomena, Antioxidants, Redox Reactions, and Methods for Their Quantification. Toxicol. Pathol. 2002, 30, 620–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girotti, A.W. Lipid Hydroperoxide Generation, Turnover, and Effector Action in Biological Systems. J. Lipid Res. 1998, 39, 1529–1542. [Google Scholar] [CrossRef]
- Yin, H.; Xu, L.; Porter, N.A. Free Radical Lipid Peroxidation: Mechanisms and Analysis. Chem. Rev. 2011, 111, 5944–5972. [Google Scholar] [CrossRef]
- Sohlenkamp, C.; Geiger, O. Bacterial Membrane Lipids: Diversity in Structures and Pathways. FEMS Microbiol. Rev. 2016, 40, 133–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marles-Wright, J.; Lewis, R.J. Stress Responses of Bacteria. Curr. Opin. Struct. Biol. 2007, 17, 755–760. [Google Scholar] [CrossRef]
- Balasubramanian, D.; Mathee, K. Comparative Transcriptome Analyses of Pseudomonas aeruginosa. Hum. Genom. 2009, 3, 349. [Google Scholar] [CrossRef] [Green Version]
- Nde, C.W.; Jang, H.-J.; Toghrol, F.; Bentley, W.E. Toxicogenomic Response of Pseudomonas aeruginosa to Ortho-Phenylphenol. BMC Genom. 2008, 9, 473. [Google Scholar] [CrossRef] [Green Version]
- Small, D.A.; Chang, W.; Toghrol, F.; Bentley, W.E. Toxicogenomic Analysis of Sodium Hypochlorite Antimicrobial Mechanisms in Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 2007, 74, 176–185. [Google Scholar] [CrossRef]
- Palma, M.; DeLuca, D.; Worgall, S.; Quadri, L.E.N. Transcriptome Analysis of the Response of Pseudomonas aeruginosa to Hydrogen Peroxide. J. Bacteriol. 2004, 186, 248–252. [Google Scholar] [CrossRef] [Green Version]
- Salunkhe, P.; Töpfer, T.; Buer, J.; Tümmler, B. Genome-Wide Transcriptional Profiling of the Steady-State Response of Pseudomonas aeruginosa to Hydrogen Peroxide. J. Bacteriol. 2005, 187, 2565–2572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, W.; Small, D.A.; Toghrol, F.; Bentley, W.E. Microarray Analysis of Pseudomonas aeruginosa Reveals Induction of Pyocin Genes in Response to Hydrogen Peroxide. BMC Genom. 2005, 6, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groitl, B.; Dahl, J.-U.; Schroeder, J.W.; Jakob, U. Pseudomonas aeruginosa Defense Systems against Microbicidal Oxidants. Mol. Microbiol. 2017, 106, 335–350. [Google Scholar] [CrossRef] [Green Version]
- Chang, W.; Small, D.A.; Toghrol, F.; Bentley, W.E. Microarray Analysis of Toxicogenomic Effects of Peracetic Acid on Pseudomonas aeruginosa. Environ. Sci. Technol. 2005, 39, 5893–5899. [Google Scholar] [CrossRef]
- Small, D.A.; Chang, W.; Toghrol, F.; Bentley, W.E. Comparative Global Transcription Analysis of Sodium Hypochlorite, Peracetic Acid, and Hydrogen Peroxide on Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 2007, 76, 1093–1105. [Google Scholar] [CrossRef]
- Kashmiri, Z.N.; Mankar, S.A. Free Radicals and Oxidative Stress in Bacteria. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 34–40. [Google Scholar]
- Farrant, K.V.; Spiga, L.; Davies, J.C.; Williams, H.D. Response of Pseudomonas aeruginosa to the Innate Immune System-Derived Oxidants Hypochlorous Acid and Hypothiocyanous Acid. J. Bacteriol. 2021, 203, 23. [Google Scholar]
- Romsang, A.; Duang-Nkern, J.; Leesukon, P.; Saninjuk, K.; Vattanaviboon, P.; Mongkolsuk, S. The Iron-Sulphur Cluster Biosynthesis Regulator IscR Contributes to Iron Homeostasis and Resistance to Oxidants in Pseudomonas aeruginosa. PLoS ONE 2014, 9, e86763. [Google Scholar] [CrossRef] [PubMed]
- Cavinato, L.; Genise, E.; Luly, F.R.; Di Domenico, E.G.; Del Porto, P.; Ascenzioni, F. Escaping the Phagocytic Oxidative Burst: The Role of SODB in the Survival of Pseudomonas aeruginosa Within Macrophages. Front. Microbiol. 2020, 11, 326. [Google Scholar] [CrossRef]
- Ghorbal, S.K.B.; Maalej, L.; Chourabi, K.; Khefacha, S.; Ouzari, H.-I.; Chatti, A. Antioxidant Defense Mechanisms in Pseudomonas aeruginosa: Role of Iron-Cofactored Superoxide Dismutase in Response to UV-C Radiations. Curr. Microbiol. 2016, 73, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Hassett, D.J.; Woodruff, W.A.; Wozniak, D.J.; Vasil, M.L.; Cohen, M.S.; Ohman, D.E. Cloning and Characterization of the Pseudomonas aeruginosa SodA and SodB Genes Encoding Manganese- and Iron-Cofactored Superoxide Dismutase: Demonstration of Increased Manganese Superoxide Dismutase Activity in Alginate-Producing Bacteria. J. Bacteriol. 1993, 175, 7658–7665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassett, D.J.; Schweizer, H.P.; Ohman, D.E. Pseudomonas aeruginosa SodA and SodB Mutants Defective in Manganese- and Iron-Cofactored Superoxide Dismutase Activity Demonstrate the Importance of the Iron-Cofactored Form in Aerobic Metabolism. J. Bacteriol. 1995, 177, 6330–6337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iiyama, K.; Chieda, Y.; Lee, J.M.; Kusakabe, T.; Yasunaga-Aoki, C.; Shimizu, S. Effect of Superoxide Dismutase Gene Inactivation on Virulence of Pseudomonas aeruginosa PAO1 toward the Silkworm, Bombyx mori. Appl. Environ. Microbiol. 2007, 73, 1569–1575. [Google Scholar] [CrossRef] [Green Version]
- Su, S.; Panmanee, W.; Wilson, J.J.; Mahtani, H.K.; Li, Q.; VanderWielen, B.D.; Makris, T.M.; Rogers, M.; McDaniel, C.; Lipscomb, J.D.; et al. Catalase (KatA) Plays a Role in Protection against Anaerobic Nitric Oxide in Pseudomonas aeruginosa. PLoS ONE 2014, 9, e91813. [Google Scholar] [CrossRef]
- Heo, Y.-J.; Chung, I.-Y.; Cho, W.-J.; Lee, B.-Y.; Kim, J.-H.; Choi, K.-H.; Lee, J.-W.; Hassett, D.J.; Cho, Y.-H. The Major Catalase Gene (KatA) of Pseudomonas aeruginosa PA14 Is under Both Positive and Negative Control of the Global Transactivator OxyR in Response to Hydrogen Peroxide. J. Bacteriol. 2010, 192, 381–390. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-S.; Heo, Y.-J.; Lee, J.K.; Cho, Y.-H. KatA, the Major Catalase, Is Critical for Osmoprotection and Virulence in Pseudomonas aeruginosa PA14. Infect. Immun. 2005, 73, 4399–4403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elkins, J.G.; Hassett, D.J.; Stewart, P.S.; Schweizer, H.P.; McDermott, T.R. Protective Role of Catalase in Pseudomonas aeruginosa Biofilm Resistance to Hydrogen Peroxide. Appl. Environ. Microbiol. 1999, 65, 4594–4600. [Google Scholar] [CrossRef] [Green Version]
- Mossialos, D.; Tavankar, G.R.; Zlosnik, J.E.A.; Williams, H.D. Defects in a Quinol Oxidase Lead to Loss of KatC Catalase Activity in Pseudomonas aeruginosa: KatC Activity Is Temperature Dependent and It Requires an Intact Disulphide Bond Formation System. Biochem. Biophys. Res. Commun. 2006, 341, 697–702. [Google Scholar] [CrossRef]
- Orlandi, V.T.; Martegani, E.; Bolognese, F. Catalase A Is Involved in the Response to Photooxidative Stress in Pseudomonas aeruginosa. Photodiagn. Photodyn. Ther. 2018, 22, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Stewart, P.S.; Roe, F.; Rayner, J.; Elkins, J.G.; Lewandowski, Z.; Ochsner, U.A.; Hassett, D.J. Effect of Catalase on Hydrogen Peroxide Penetration into Pseudomonas aeruginosa Biofilms. Appl. Environ. Microbiol. 2000, 66, 836–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, D.-H.; Choi, Y.-S.; Cho, Y.-H. Unusual Properties of Catalase A (KatA) of Pseudomonas aeruginosa PA14 Are Associated with Its Biofilm Peroxide Resistance. J. Bacteriol. 2008, 190, 2663–2670. [Google Scholar] [CrossRef] [Green Version]
- Hare, N.J.; Scott, N.E.; Shin, E.H.H.; Connolly, A.M.; Larsen, M.R.; Palmisano, G.; Cordwell, S.J. Proteomics of the Oxidative Stress Response Induced by Hydrogen Peroxide and Paraquat Reveals a Novel AhpC-like Protein in Pseudomonas aeruginosa. Proteomics 2011, 11, 3056–3069. [Google Scholar] [CrossRef]
- Ochsner, U.A.; Vasil, M.L.; Alsabbagh, E.; Parvatiyar, K.; Hassett, D.J. Role of the Pseudomonas aeruginosa OxyR-RecG Operon in Oxidative Stress Defense and DNA Repair: OxyR-Dependent Regulation of KatB-AnkB, AhpB, and AhpC-AhpF. J. Bacteriol. 2000, 182, 4533–4544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.T.; Lee, S.S.; Mondal, S.; Tripathi, B.N.; Kim, S.; Lee, K.W.; Hong, S.H.; Bai, H.-W.; Cho, J.-Y.; Chung, B.Y. Enhancement of the Chaperone Activity of Alkyl Hydroperoxide Reductase C from Pseudomonas aeruginosa PAO1 Resulting from a Point-Specific Mutation Confers Heat Tolerance in Escherichia coli. Mol. Cells 2016, 39, 594–602. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.H.; Singh, S.; Tripathi, B.N.; Mondal, S.; Lee, S.; Jung, H.S.; Cho, C.; Kaur, S.; Kim, J.-H.; Lee, S.; et al. Functional Properties and the Oligomeric State of Alkyl Hydroperoxide Reductase Subunit F (AhpF) in Pseudomonas aeruginosa. Protoplasma 2020, 257, 807–817. [Google Scholar] [CrossRef] [PubMed]
- Imlay, J.A. Cellular Defenses against Superoxide and Hydrogen Peroxide. Annu. Rev. Biochem. 2008, 77, 755–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romsang, A.; Atichartpongkul, S.; Trinachartvanit, W.; Vattanaviboon, P.; Mongkolsuk, S. Gene Expression and Physiological Role of Pseudomonas aeruginosa Methionine Sulfoxide Reductases during Oxidative Stress. J. Bacteriol. 2013, 195, 3299–3308. [Google Scholar] [CrossRef] [Green Version]
- Ezraty, B.; Aussel, L.; Barras, F. Methionine Sulfoxide Reductases in Prokaryotes. Biochim. Biophys. Acta (BBA)—Proteins Proteom. 2005, 1703, 221–229. [Google Scholar] [CrossRef]
- Singh, V.K.; Singh, K.; Baum, K. The Role of Methionine Sulfoxide Reductases in Oxidative Stress Tolerance and Virulence of Staphylococcus aureus and Other Bacteria. Antioxidants 2018, 7, 128. [Google Scholar] [CrossRef] [Green Version]
- Sultana, S.; Foti, A.; Dahl, J.-U. Bacterial Defense Systems against the Neutrophilic Oxidant Hypochlorous Acid. Infect. Immun. 2020, 88, e00964-19. [Google Scholar] [CrossRef]
- Pena, R.T.; Blasco, L.; Ambroa, A.; González-Pedrajo, B.; Fernández-García, L.; López, M.; Bleriot, I.; Bou, G.; García-Contreras, R.; Wood, T.K.; et al. Relationship Between Quorum Sensing and Secretion Systems. Front. Microbiol. 2019, 10, 1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maura, D.; Hazan, R.; Kitao, T.; Ballok, A.E.; Rahme, L.G. Evidence for Direct Control of Virulence and Defense Gene Circuits by the Pseudomonas aeruginosa Quorum Sensing Regulator, MvfR. Sci. Rep. 2016, 6, 34083. [Google Scholar] [CrossRef]
- Balasubramanian, D.; Schneper, L.; Kumari, H.; Mathee, K. A Dynamic and Intricate Regulatory Network Determines Pseudomonas aeruginosa Virulence. Nucleic Acids Res. 2013, 41, 1–20. [Google Scholar] [CrossRef]
- Sabra, W.; Kim, E.-J.; Zeng, A.-P. Physiological Responses of Pseudomonas aeruginosa PAO1 to Oxidative Stress in Controlled Microaerobic and Aerobic Cultures. Microbiology 2002, 148, 3195–3202. [Google Scholar] [CrossRef] [Green Version]
- Pacheco, G.J.; Reis, R.S.; Fernandes, A.C.L.B.; da Rocha, S.L.G.; Pereira, M.D.; Perales, J.; Freire, D.M.G. Rhamnolipid Production: Effect of Oxidative Stress on Virulence Factors and Proteome of Pseudomonas aeruginosa PA1. Appl. Microbiol. Biotechnol. 2012, 95, 1519–1529. [Google Scholar] [CrossRef] [PubMed]
- Malešević, M.; Di Lorenzo, F.; Filipić, B.; Stanisavljević, N.; Novović, K.; Senerovic, L.; Polović, N.; Molinaro, A.; Kojić, M.; Jovčić, B. Pseudomonas aeruginosa Quorum Sensing Inhibition by Clinical Isolate Delftia Tsuruhatensis 11304: Involvement of N -Octadecanoylhomoserine Lactones. Sci. Rep. 2019, 9, 16465. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Kong, J.; Dong, B.; Huang, H.; Wang, K.; Wu, L.; Hou, C.; Liang, Y.; Li, B.; Chen, Y. Baicalein Attenuates the Quorum Sensing-Controlled Virulence Factors of Pseudomonas aeruginosa and Relieves the Inflammatory Response in P. Aeruginosa-Infected Macrophages by Downregulating the MAPK and NFκB Signal-Transduction Pathways. Drug Des. Dev. Ther. 2016, 10, 183–203. [Google Scholar] [CrossRef] [Green Version]
- Kievit, T.R.D. Quorum Sensing in Pseudomonas aeruginosa Biofilms. Environ. Microbiol. 2009, 11, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Sakuragi, Y.; Kolter, R. Quorum-Sensing Regulation of the Biofilm Matrix Genes (Pel) of Pseudomonas aeruginosa. J. Bacteriol. 2007, 189, 5383–5386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassett, D.J.; Ma, J.-F.; Elkins, J.G.; McDermott, T.R.; Ochsner, U.A.; West, S.E.H.; Huang, C.-T.; Fredericks, J.; Burnett, S.; Stewart, P.S.; et al. Quorum Sensing in Pseudomonas aeruginosa Controls Expression of Catalase and Superoxide Dismutase Genes and Mediates Biofilm Susceptibility to Hydrogen Peroxide. Mol. Microbiol. 1999, 34, 1082–1093. [Google Scholar] [CrossRef]
- Bollinger, N.; Hassett, D.J.; Iglewski, B.H.; Costerton, J.W.; McDermott, T.R. Gene Expression in Pseudomonas aeruginosa: Evidence of Iron Override Effects on Quorum Sensing and Biofilm-Specific Gene Regulation. J. Bacteriol. 2001, 183, 1990–1996. [Google Scholar] [CrossRef] [Green Version]
- García-Contreras, R.; Nuñez-López, L.; Jasso-Chávez, R.; Kwan, B.W.; Belmont, J.A.; Rangel-Vega, A.; Maeda, T.; Wood, T.K. Quorum Sensing Enhancement of the Stress Response Promotes Resistance to Quorum Quenching and Prevents Social Cheating. ISME J. 2015, 9, 115–125. [Google Scholar] [CrossRef]
- Kafle, P.; Amoh, A.N.; Reaves, J.M.; Suneby, E.G.; Tutunjian, K.A.; Tyson, R.L.; Schneider, T.L. Molecular Insights into the Impact of Oxidative Stress on the Quorum-Sensing Regulator Protein LasR. J. Biol. Chem. 2016, 291, 11776–11786. [Google Scholar] [CrossRef] [Green Version]
- Häussler, S.; Becker, T. The Pseudomonas Quinolone Signal (PQS) Balances Life and Death in Pseudomonas aeruginosa Populations. PLoS Pathog. 2008, 4, e1000166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, J.; Damron, F.H.; Basler, M.; Digiandomenico, A.; Sherman, N.E.; Fox, J.W.; Mekalanos, J.J.; Goldberg, J.B. Comparisons of Two Proteomic Analyses of Non-Mucoid and Mucoid Pseudomonas aeruginosa Clinical Isolates from a Cystic Fibrosis Patient. Front. Microbiol. 2011, 2, 162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pulcrano, G.; Iula, D.V.; Raia, V.; Rossano, F.; Catania, M.R. Different Mutations in MucA Gene of Pseudomonas aeruginosa Mucoid Strains in Cystic Fibrosis Patients and Their Effect on AlgU Gene Expression. New Microbiol. 2012, 35, 295–305. [Google Scholar]
- Qiu, D.; Eisinger, V.M.; Rowen, D.W.; Yu, H.D. Regulated Proteolysis Controls Mucoid Conversion in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2007, 104, 8107–8112. [Google Scholar] [CrossRef] [Green Version]
- Wood, L.F.; Leech, A.J.; Ohman, D.E. Cell Wall-Inhibitory Antibiotics Activate the Alginate Biosynthesis Operon in Pseudomonas aeruginosa: Roles of Σ22 (AlgT) and the AlgW and Prc Proteases. Mol. Microbiol. 2006, 62, 412–426. [Google Scholar] [CrossRef]
- Martin, D.W.; Schurr, M.J.; Yu, H.; Deretic, V. Analysis of Promoters Controlled by the Putative Sigma Factor AlgU Regulating Conversion to Mucoidy in Pseudomonas aeruginosa: Relationship to Sigma E and Stress Response. J. Bacteriol. 1994, 176, 6688–6696. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Lou, X.; Xu, Y.; Teng, X.; Liu, R.; Zhang, Q.; Wu, W.; Wang, Y.; Bartlam, M. Structural Basis for the Recognition of MucA by MucB and AlgU in Pseudomonas aeruginosa. FEBS J. 2019, 286, 4982–4994. [Google Scholar] [CrossRef]
- Damron, F.H.; Yu, H.D. Pseudomonas aeruginosa MucD Regulates the Alginate Pathway through Activation of MucA Degradation via MucP Proteolytic Activity. J. Bacteriol. 2011, 193, 286–291. [Google Scholar] [CrossRef] [Green Version]
- Mathee, K.; Ciofu, O.; Sternberg, C.; Lindum, P.W.; Campbell, J.I.A.; Jensen, P.; Johnsen, A.H.; Givskov, M.; Ohman, D.E.; Søren, M.; et al. Mucoid Conversion of Pseudomonas aeruginosa by Hydrogen Peroxide: A Mechanism for Virulence Activation in the Cystic Fibrosis Lung. Microbiology 1999, 145, 1349–1357. [Google Scholar] [CrossRef] [Green Version]
- Damron, F.H.; Napper, J.; Teter, M.A.; Yu, H.D. Lipotoxin F of Pseudomonas aeruginosa Is an AlgU-Dependent and Alginate-Independent Outer Membrane Protein Involved in Resistance to Oxidative Stress and Adhesion to A549 Human Lung Epithelia. Microbiology 2009, 155, 1028–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crespo, A.; Pedraz, L.; Van Der Hofstadt, M.; Gomila, G.; Torrents, E. Regulation of Ribonucleotide Synthesis by the Pseudomonas aeruginosa Two-Component System AlgR in Response to Oxidative Stress. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef]
- Boucher, J.C.; Martinez-Salazar, J.; Schurr, M.J.; Mudd, M.H.; Yu, H.; Deretic, V. Two Distinct Loci Affecting Conversion to Mucoidy in Pseudomonas aeruginosa in Cystic Fibrosis Encode Homologs of the Serine Protease HtrA. J. Bacteriol. 1996, 178, 511–523. [Google Scholar] [CrossRef] [Green Version]
- Orlandi, V.T.; Bolognese, F.; Chiodaroli, L.; Tolker-Nielsen, T.; Barbieri, P. Pigments Influence the Tolerance of Pseudomonas aeruginosa PAO1 to Photodynamically Induced Oxidative Stress. Microbiology 2015, 161, 2298–2309. [Google Scholar] [CrossRef]
- Thippakorn, C.; Isarankura-Na-Ayudhya, C.; Pannengpetch, S.; Isarankura-Na-Ayudhya, P.; Schaduangrat, N.; Nantasenamat, C.; Prachayasittikul, V. Oxidative Responses and Defense Mechanism of Hyperpigmented P. aeruginosa as Characterized by Proteomics and Metabolomics. EXCLI J. 2018, 17, 544–562. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Renga Thavasi, T.; Jayalakshmi, S. Phenazine Pigments from Pseudomonas aeruginosa and Their Application as Antibacterial Agent and Food Colourants. Res. J. Microbiol. 2008, 3, 122–128. [Google Scholar] [CrossRef] [Green Version]
- Jayaseelan, S.; Ramaswamy, D.; Dharmaraj, S. Pyocyanin: Production, Applications, Challenges and New Insights. World J. Microbiol. Biotechnol. 2014, 30, 1159–1168. [Google Scholar] [CrossRef]
- Visca, P.; Imperi, F.; Lamont, I.L. Pyoverdine Siderophores: From Biogenesis to Biosignificance. Trends Microbiol. 2007, 15, 22–30. [Google Scholar] [CrossRef]
- Rodríguez-Rojas, A.; Mena, A.; Martín, S.; Borrell, N.; Oliver, A.; Blázquez, J. Inactivation of the HmgA Gene of Pseudomonas aeruginosa Leads to Pyomelanin Hyperproduction, Stress Resistance and Increased Persistence in Chronic Lung Infection. Microbiology 2009, 155, 1050–1057. [Google Scholar] [CrossRef] [Green Version]
- Meirelles, L.A.; Newman, D.K. Both Toxic and Beneficial Effects of Pyocyanin Contribute to the Lifecycle of Pseudomonas aeruginosa. Mol. Microbiol. 2018, 110, 995–1010. [Google Scholar] [CrossRef] [Green Version]
- Hassett, D.J.; Charniga, L.; Bean, K.; Ohman, D.E.; Cohen, M.S. Response of Pseudomonas aeruginosa to Pyocyanin: Mechanisms of Resistance, Antioxidant Defenses, and Demonstration of a Manganese-Cofactored Superoxide Dismutase. Infect. Immun. 1992, 60, 328–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhargava, N.; Sharma, P.; Capalash, N. Pyocyanin Stimulates Quorum Sensing-Mediated Tolerance to Oxidative Stress and Increases Persister Cell Populations in Acinetobacter Baumannii. Infect. Immun. 2014, 82, 3417–3425. [Google Scholar] [CrossRef] [Green Version]
- Dolan, S.K.; Kohlstedt, M.; Trigg, S.; Ramirez, P.V.; Kaminski, C.F.; Wittmann, C.; Welch, M. Contextual Flexibility in Pseudomonas aeruginosa Central Carbon Metabolism during Growth in Single Carbon Sources. mBio 2020, 11, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger, A.; Dohnt, K.; Tielen, P.; Jahn, D.; Becker, J.; Wittmann, C. Robustness and Plasticity of Metabolic Pathway Flux among Uropathogenic Isolates of Pseudomonas aeruginosa. PLoS ONE 2014, 9, e88368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chavarría, M.; Nikel, P.I.; Pérez-Pantoja, D.; de Lorenzo, V. The Entner–Doudoroff Pathway Empowers Pseudomonas Putida KT2440 with a High Tolerance to Oxidative Stress. Environ. Microbiol. 2013, 15, 1772–1785. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.; Jung, J.; Jang, I.-A.; Madsen, E.L.; Park, W. Role of Glyoxylate Shunt in Oxidative Stress Response. J. Biol. Chem. 2016, 291, 11928–11938. [Google Scholar] [CrossRef] [Green Version]
- Ha, S.; Shin, B.; Park, W. Lack of Glyoxylate Shunt Dysregulates Iron Homeostasis in Pseudomonas aeruginosa. Microbiology 2018, 164, 587–599. [Google Scholar] [CrossRef]
- Koedooder, C.; Guéneuguès, A.; Van Geersdaële, R.; Vergé, V.; Bouget, F.-Y.; Labreuche, Y.; Obernosterer, I.; Blain, S. The Role of the Glyoxylate Shunt in the Acclimation to Iron Limitation in Marine Heterotrophic Bacteria. Front. Mar. Sci. 2018, 5, 435. [Google Scholar] [CrossRef] [Green Version]
- Morita, T.; Kawamoto, H.; Mizota, T.; Inada, T.; Aiba, H. Enolase in the RNA Degradosome Plays a Crucial Role in the Rapid Decay of Glucose Transporter MRNA in the Response to Phosphosugar Stress in Escherichia coli. Mol. Microbiol. 2004, 54, 1063–1075. [Google Scholar] [CrossRef] [PubMed]
- Weng, Y.; Chen, F.; Liu, Y.; Zhao, Q.; Chen, R.; Pan, X.; Liu, C.; Cheng, Z.; Jin, S.; Jin, Y.; et al. Pseudomonas aeruginosa Enolase Influences Bacterial Tolerance to Oxidative Stresses and Virulence. Front. Microbiol. 2016, 7, 1999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corona, F.; Martínez, J.L.; Nikel, P.I. The Global Regulator Crc Orchestrates the Metabolic Robustness Underlying Oxidative Stress Resistance in Pseudomonas aeruginosa. Environ. Microbiol. 2019, 21, 898–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picard, F.; Dressaire, C.; Girbal, L.; Cocaign-Bousquet, M. Examination of Post-Transcriptional Regulations in Prokaryotes by Integrative Biology. Comptes Rendus Biol. 2009, 332, 958–973. [Google Scholar] [CrossRef]
- Thongdee, N.; Jaroensuk, J.; Atichartpongkul, S.; Chittrakanwong, J.; Chooyoung, K.; Srimahaeak, T.; Chaiyen, P.; Vattanaviboon, P.; Mongkolsuk, S.; Fuangthong, M. TrmB, a TRNA M7G46 Methyltransferase, Plays a Role in Hydrogen Peroxide Resistance and Positively Modulates the Translation of KatA and KatB MRNAs in Pseudomonas aeruginosa. Nucleic Acids Res. 2019, 47, 9271–9281. [Google Scholar] [CrossRef] [Green Version]
- Jaroensuk, J.; Atichartpongkul, S.; Chionh, Y.H.; Wong, Y.H.; Liew, C.W.; McBee, M.E.; Thongdee, N.; Prestwich, E.G.; DeMott, M.S.; Mongkolsuk, S.; et al. Methylation at Position 32 of TRNA Catalyzed by TrmJ Alters Oxidative Stress Response in Pseudomonas aeruginosa. Nucleic Acids Res. 2016, 44, 10834–10848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Summer, S.; Smirnova, A.; Gabriele, A.; Toth, U.; Fasemore, A.M.; Förstner, K.U.; Kuhn, L.; Chicher, J.; Hammann, P.; Mitulović, G.; et al. YBEY Is an Essential Biogenesis Factor for Mitochondrial Ribosomes. Nucleic Acids Res. 2020, 48, 9762–9786. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Weng, Y.; Xu, C.; Wang, D.; Pan, X.; Tian, Z.; Xia, B.; Li, H.; Chen, R.; Liu, C.; et al. Endoribonuclease YbeY Is Essential for RNA Processing and Virulence in Pseudomonas aeruginosa. mBio 2020, 11. [Google Scholar] [CrossRef]
- Gómez-Lozano, M.; Marvig, R.L.; Tulstrup, M.V.; Molin, S. Expression of Antisense Small RNAs in Response to Stress in Pseudomonas aeruginosa. BMC Genom. 2014, 15, 783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Wang, B.; Gruber, J.D.; Zhang, Y.-M.; Davies, C. Acyl Carrier Protein 3 Is Involved in Oxidative Stress Response in Pseudomonas aeruginosa. Front. Microbiol. 2018, 9, 2244. [Google Scholar] [CrossRef] [PubMed]
- Koskenkorva-Frank, T.S.; Kallio, P.T. Induction of Pseudomonas aeruginosa Fhp and FhpR by Reactive Oxygen Species. Can. J. Microbiol. 2009, 55, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Goldová, J.; Ulrych, A.; Hercík, K.; Branny, P. A Eukaryotic-Type Signalling System of Pseudomonas aeruginosa Contributes to Oxidative Stress Resistance, Intracellular Survival and Virulence. BMC Genom. 2011, 12, 437. [Google Scholar] [CrossRef] [Green Version]
- Moyano, A.J.; Tobares, R.A.; Rizzi, Y.S.; Krapp, A.R.; Mondotte, J.A.; Bocco, J.L.; Saleh, M.-C.; Carrillo, N.; Smania, A.M. A Long-Chain Flavodoxin Protects Pseudomonas aeruginosa from Oxidative Stress and Host Bacterial Clearance. PLoS Genet. 2014, 10, e1004163. [Google Scholar] [CrossRef]
- Panmanee, W.; Charoenlap, N.; Atichartpongkul, S.; Mahavihakanont, A.; Whiteside, M.D.; Winsor, G.; Brinkman, F.S.L.; Mongkolsuk, S.; Hassett, D.J. The OxyR-Regulated PhnW Gene Encoding 2-Aminoethylphosphonate:Pyruvate Aminotransferase Helps Protect Pseudomonas aeruginosa from Tert-Butyl Hydroperoxide. PLoS ONE 2017, 12, e0189066. [Google Scholar] [CrossRef] [Green Version]
- Ciofu, O.; Mandsberg, L.F.; Wang, H.; Høiby, N. Phenotypes Selected during Chronic Lung Infection in Cystic Fibrosis Patients: Implications for the Treatment of Pseudomonas aeruginosa Biofilm Infections. FEMS Immunol. Med. Microbiol. 2012, 65, 215–225. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Barat, L.; Ciofu, O.; Kragh, K.N.; Pressler, T.; Johansen, U.; Motos, A.; Torres, A.; Hoiby, N. Phenotypic Shift in Pseudomonas aeruginosa Populations from Cystic Fibrosis Lungs after 2-Week Antipseudomonal Treatment. J. Cyst. Fibros. 2017, 16, 222–229. [Google Scholar] [CrossRef] [Green Version]
- Ciofu, O.; Riis, B.; Pressler, T.; Poulsen, H.E.; Høiby, N. Occurrence of Hypermutable Pseudomonas Aeruginosa in Cystic Fibrosis Patients Is Associated with the Oxidative Stress Caused by Chronic Lung Inflammation. Antimicrob. Agents Chemother. 2005, 49, 2276–2282. [Google Scholar] [CrossRef] [Green Version]
- Colque, C.A.; Albarracín Orio, A.G.; Feliziani, S.; Marvig, R.L.; Tobares, A.R.; Johansen, H.K.; Molin, S.; Smania, A.M. Hypermutator Pseudomonas aeruginosa Exploits Multiple Genetic Pathways To Develop Multidrug Resistance during Long-Term Infections in the Airways of Cystic Fibrosis Patients. Antimicrob. Agents Chemother. 2020, 64, e02142-19. [Google Scholar] [CrossRef]
- Bae, H.-W.; Cho, Y.-H. Mutational Analysis of Pseudomonas aeruginosa OxyR to Define the Regions Required for Peroxide Resistance and Acute Virulence. Res. Microbiol. 2012, 163, 55–63. [Google Scholar] [CrossRef]
- Wei, Q.; Le Minh, P.N.; Dötsch, A.; Hildebrand, F.; Panmanee, W.; Elfarash, A.; Schulz, S.; Plaisance, S.; Charlier, D.; Hassett, D.; et al. Global Regulation of Gene Expression by OxyR in an Important Human Opportunistic Pathogen. Nucleic Acids Res. 2012, 40, 4320–4333. [Google Scholar] [CrossRef]
- Papp-Szabò, E.; Firtel, M.; Josephy, P.D. Comparison of the Sensitivities of Salmonella Typhimurium OxyR and KatG Mutants to Killing by Human Neutrophils. Infect. Immun. 1994, 62, 2662–2668. [Google Scholar] [CrossRef] [Green Version]
- Storz, G.; Tartaglia, L.A. OxyR: A Regulator of Antioxidant Genes. J. Nutr. 1992, 122, 627–630. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Sun, M.; Cheng, Y.; Yang, R.; Wen, Y.; Chen, Z.; Li, J. OxyR Is a Key Regulator in Response to Oxidative Stress in Streptomyces Avermitilis. Microbiology 2016, 162, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, K.; Ohniwa, R.L.; Kim, J.; Maruyama, A.; Ohta, T.; Takeyasu, K. Bacterial Nucleoid Dynamics: Oxidative Stress Response in Staphylococcus aureus. Genes Cells 2006, 11, 409–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubbs, J.M.; Mongkolsuk, S. Peroxide-Sensing Transcriptional Regulators in Bacteria. J. Bacteriol. 2012, 194, 5495–5503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinckx, T.; Wei, Q.; Matthijs, S.; Cornelis, P. The Pseudomonas aeruginosa Oxidative Stress Regulator OxyR Influences Production of Pyocyanin and Rhamnolipids: Protective Role of Pyocyanin. Microbiology 2010, 156, 678–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinckx, T.; Matthijs, S.; Cornelis, P. Loss of the Oxidative Stress Regulator OxyR in Pseudomonas aeruginosa PAO1 Impairs Growth under Iron-Limited Conditions. FEMS Microbiol. Lett. 2008, 288, 258–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, M.; Lyu, M.; Wen, Y.; Song, Y.; Li, J.; Chen, Z. Organic Peroxide-Sensing Repressor OhrR Regulates Organic Hydroperoxide Stress Resistance and Avermectin Production in Streptomyces avermitilis. Front. Microbiol. 2018, 9, 1398. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.-Y.; Shin, J.-H.; Roe, J.-H. Dual Role of OhrR as a Repressor and an Activator in Response to Organic Hydroperoxides in Streptomyces Coelicolor. J. Bacteriol. 2007, 189, 6284–6292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atichartpongkul, S.; Fuangthong, M.; Vattanaviboon, P.; Mongkolsuk, S. Analyses of the Regulatory Mechanism and Physiological Roles of Pseudomonas aeruginosa OhrR, a Transcription Regulator and a Sensor of Organic Hydroperoxides. J. Bacteriol. 2010, 192, 2093–2101. [Google Scholar] [CrossRef] [Green Version]
- Atichartpongkul, S.; Vattanaviboon, P.; Wisitkamol, R.; Jaroensuk, J.; Mongkolsuk, S.; Fuangthong, M. Regulation of Organic Hydroperoxide Stress Response by Two OhrR Homologs in Pseudomonas aeruginosa. PLoS ONE 2016, 11, e0161982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ochsner, U.A.; Hassett, D.J.; Vasil, M.L. Genetic and Physiological Characterization Of ohr, Encoding a Protein Involved in Organic Hydroperoxide Resistance in Pseudomonas aeruginosa. J. Bacteriol. 2001, 183, 773–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, L.; Murray, T.S.; Kazmierczak, B.I.; He, C. Pseudomonas aeruginosa OspR Is an Oxidative Stress Sensing Regulator That Affects Pigment Production, Antibiotic Resistance and Dissemination during Infection. Mol. Microbiol. 2010, 75, 76–91. [Google Scholar] [CrossRef] [Green Version]
- Crack, J.C.; Green, J.; Hutchings, M.I.; Thomson, A.J.; Le Brun, N.E. Bacterial Iron-Sulfur Regulatory Proteins As Biological Sensor-Switches. Antioxid. Redox Signal. 2012, 17, 1215–1231. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.-S.; Shin, D.-H.; Chung, I.-Y.; Kim, S.-H.; Heo, Y.-J.; Cho, Y.-H. Identification of Pseudomonas aeruginosa Genes Crucial for Hydrogen Peroxide Resistance. J. Microbiol. Biotechnol. 2007, 17, 1344–1352. [Google Scholar]
- Saninjuk, K.; Romsang, A.; Duang-nkern, J.; Vattanaviboon, P.; Mongkolsuk, S. Transcriptional Regulation of the Pseudomonas aeruginosa Iron-Sulfur Cluster Assembly Pathway by Binding of IscR to Multiple Sites. PLoS ONE 2019, 14, e0218385. [Google Scholar] [CrossRef] [Green Version]
- Miller, H.K.; Auerbuch, V. Bacterial Iron-Sulfur Cluster Sensors in Mammalian Pathogens. Metallomics 2015, 7, 943–956. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-H.; Lee, B.-Y.; Lau, G.W.; Cho, Y.-H. IscR Modulates Catalase A (KatA) Activity, Peroxide Resistance and Full Virulence of Pseudomonas aeruginosa PA14. J. Microbiol. Biotechnol. 2009, 19, 1520–1526. [Google Scholar] [CrossRef] [Green Version]
- Romsang, A.; Duangnkern, J.; Wirathorn, W.; Vattanaviboon, P.; Mongkolsuk, S. Pseudomonas aeruginosa IscR-Regulated Ferredoxin NADP(+) Reductase Gene (FprB) Functions in Iron-Sulfur Cluster Biogenesis and Multiple Stress Response. PLoS ONE 2015, 10, e0134374. [Google Scholar] [CrossRef] [Green Version]
- Romsang, A.; Duangnkern, J.; Saninjuk, K.; Vattanaviboon, P.; Mongkolsuk, S. Pseudomonas aeruginosa NfuA: Gene Regulation and Its Physiological Roles in Sustaining Growth under Stress and Anaerobic Conditions and Maintaining Bacterial Virulence. PLoS ONE 2018, 13, e0202151. [Google Scholar] [CrossRef]
- Boonma, S.; Romsang, A.; Duangnkern, J.; Atichartpongkul, S.; Trinachartvanit, W.; Vattanaviboon, P.; Mongkolsuk, S. The FinR-Regulated Essential Gene FprA, Encoding Ferredoxin NADP+ Reductase: Roles in Superoxide-Mediated Stress Protection and Virulence of Pseudomonas aeruginosa. PLoS ONE 2017, 12, e0172071. [Google Scholar] [CrossRef]
- Prince, R.W.; Cox, C.D.; Vasil, M.L. Coordinate Regulation of Siderophore and Exotoxin A Production: Molecular Cloning and Sequencing of the Pseudomonas aeruginosa Fur Gene. J. Bacteriol. 1993, 175, 2589–2598. [Google Scholar] [CrossRef] [Green Version]
- Barton, H.A.; Johnson, Z.; Cox, C.D.; Vasil, A.I.; Vasil, M.L. Ferric Uptake Regulator Mutants of Pseudomonas aeruginosa with Distinct Alterations in the Iron-Dependent Repression of Exotoxin A and Siderophores in Aerobic and Microaerobic Environments. Mol. Microbiol. 1996, 21, 1001–1017. [Google Scholar] [CrossRef]
- Pérard, J.; Covès, J.; Castellan, M.; Solard, C.; Savard, M.; Miras, R.; Galop, S.; Signor, L.; Crouzy, S.; Michaud-Soret, I.; et al. Quaternary Structure of Fur Proteins, a New Subfamily of Tetrameric Proteins. Biochemistry 2016, 55, 1503–1515. [Google Scholar] [CrossRef] [PubMed]
- Wilderman, P.J.; Sowa, N.A.; FitzGerald, D.J.; FitzGerald, P.C.; Gottesman, S.; Ochsner, U.A.; Vasil, M.L. Identification of Tandem Duplicate Regulatory Small RNAs in Pseudomonas aeruginosa Involved in Iron Homeostasis. Proc. Natl. Acad. Sci. USA 2004, 101, 9792–9797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasqua, M.; Visaggio, D.; Lo Sciuto, A.; Genah, S.; Banin, E.; Visca, P.; Imperi, F. Ferric Uptake Regulator Fur Is Conditionally Essential in Pseudomonas aeruginosa. J. Bacteriol. 2017, 199, e00472-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, U.; Jin, S. Expression of the SoxR Gene of Pseudomonas aeruginosa Is Inducible during Infection of Burn Wounds in Mice and Is Required To Cause Efficient Bacteremia. Infect. Immun. 1999, 67, 5324–5331. [Google Scholar] [CrossRef] [Green Version]
- Koo, M.-S.; Lee, J.-H.; Rah, S.-Y.; Yeo, W.-S.; Lee, J.-W.; Lee, K.-L.; Koh, Y.-S.; Kang, S.-O.; Roe, J.-H. A Reducing System of the Superoxide Sensor SoxR in Escherichia coli. EMBO J. 2003, 22, 2614–2622. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, K. Sensing Mechanisms in the Redox-Regulated, [2Fe–2S] Cluster-Containing, Bacterial Transcriptional Factor SoxR. Acc. Chem. Res. 2017, 50, 1672–1678. [Google Scholar] [CrossRef]
- Singh, A.K.; Shin, J.-H.; Lee, K.-L.; Imlay, J.A.; Roe, J.-H. Comparative Study of SoxR Activation by Redox-Active Compounds. Mol. Microbiol. 2013, 90, 983–996. [Google Scholar] [CrossRef] [Green Version]
- Pomposiello, P.J.; Demple, B. Redox-Operated Genetic Switches: The SoxR and OxyR Transcription Factors. Trends Biotechnol. 2001, 19, 109–114. [Google Scholar] [CrossRef]
- Palma, M.; Zurita, J.; Ferreras, J.A.; Worgall, S.; Larone, D.H.; Shi, L.; Campagne, F.; Quadri, L.E.N. Pseudomonas aeruginosa SoxR Does Not Conform to the Archetypal Paradigm for SoxR-Dependent Regulation of the Bacterial Oxidative Stress Adaptive Response. Infect. Immun. 2005, 73, 2958–2966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aendekerk, S.; Ghysels, B.; Cornelis, P.; Baysse, C. Characterization of a New Efflux Pump, MexGHI-OpmD, from Pseudomonas aeruginosa That Confers Resistance to Vanadium. Microbiology 2002, 148, 2371–2381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietrich, L.E.P.; Price-Whelan, A.; Petersen, A.; Whiteley, M.; Newman, D.K. The Phenazine Pyocyanin Is a Terminal Signalling Factor in the Quorum Sensing Network of Pseudomonas aeruginosa. Mol. Microbiol. 2006, 61, 1308–1321. [Google Scholar] [CrossRef] [PubMed]
- Reen, F.J.; Haynes, J.M.; Mooij, M.J.; O’Gara, F. A Non-Classical LysR-Type Transcriptional Regulator PA2206 Is Required for an Effective Oxidative Stress Response in Pseudomonas aeruginosa. PLoS ONE 2013, 8, e54479. [Google Scholar] [CrossRef] [Green Version]
- Rungrassamee, W.; Ryan, K.C.; Maroney, M.J.; Pomposiello, P.J. The PqrR Transcriptional Repressor of Pseudomonas aeruginosa Transduces Redox Signals via an Iron-Containing Prosthetic Group. J. Bacteriol. 2009, 191, 6709–6721. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhou, C.; Pu, Q.; Wu, Q.; Tan, S.; Shao, X.; Zhang, W.; Xie, Y.; Li, R.; Yu, X.; et al. Pseudomonas aeruginosa Regulatory Protein AnvM Controls Pathogenicity in Anaerobic Environments and Impacts Host Defense. mBio 2019, 10, e01362-19. [Google Scholar] [CrossRef] [Green Version]
- Issa, K.H.B.; Phan, G.; Broutin, I. Functional Mechanism of the Efflux Pumps Transcription Regulators From Pseudomonas aeruginosa Based on 3D Structures. Front. Mol. Biosci. 2018, 5, 57. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Hu, J.; Chen, P.R.; Lan, L.; Li, Z.; Hicks, L.M.; Dinner, A.R.; He, C. The Pseudomonas aeruginosa Multidrug Efflux Regulator MexR Uses an Oxidation-Sensing Mechanism. Proc. Natl. Acad. Sci. USA 2008, 105, 13586–13591. [Google Scholar] [CrossRef] [Green Version]
- Fraud, S.; Poole, K. Oxidative Stress Induction of the MexXY Multidrug Efflux Genes and Promotion of Aminoglycoside Resistance Development in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2011, 55, 1068–1074. [Google Scholar] [CrossRef] [Green Version]
- Fetar, H.; Gilmour, C.; Klinoski, R.; Daigle, D.M.; Dean, C.R.; Poole, K. MexEF-OprN Multidrug Efflux Operon of Pseudomonas aeruginosa: Regulation by the MexT Activator in Response to Nitrosative Stress and Chloramphenicol. Antimicrob. Agents Chemother. 2011, 55, 508–514. [Google Scholar] [CrossRef] [Green Version]
- Muller, J.F.; Stevens, A.M.; Craig, J.; Love, N.G. Transcriptome Analysis Reveals That Multidrug Efflux Genes Are Upregulated To Protect Pseudomonas aeruginosa from Pentachlorophenol Stress. Appl. Environ. Microbiol. 2007, 73, 4550–4558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandsberg, L.F.; Ciofu, O.; Kirkby, N.; Christiansen, L.E.; Poulsen, H.E.; Høiby, N. Antibiotic Resistance in Pseudomonas aeruginosa Strains with Increased Mutation Frequency Due to Inactivation of the DNA Oxidative Repair System. Antimicrob. Agents Chemother. 2009, 53, 2483–2491. [Google Scholar] [CrossRef] [Green Version]
- Masuda, N.; Sakagawa, E.; Ohya, S.; Gotoh, N.; Tsujimoto, H.; Nishino, T. Substrate Specificities of MexAB-OprM, MexCD-OprJ, and MexXY-OprM Efflux Pumps in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2000, 44, 3322–3327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laborda, P.; Alcalde-Rico, M.; Blanco, P.; Martínez, J.L.; Hernando-Amado, S. Novel Inducers of the Expression of Multidrug Efflux Pumps That Trigger Pseudomonas aeruginosa Transient Antibiotic Resistance. Antimicrob. Agents Chemother. 2019, 63, 15. [Google Scholar] [CrossRef]
- Jin, M.; Liu, L.; Wang, D.; Yang, D.; Liu, W.; Yin, J.; Yang, Z.; Wang, H.; Qiu, Z.; Shen, Z.; et al. Chlorine Disinfection Promotes the Exchange of Antibiotic Resistance Genes across Bacterial Genera by Natural Transformation. ISME J. 2020, 14, 1847–1856. [Google Scholar] [CrossRef]
- Valentini, M.; Filloux, A. Biofilms and Cyclic Di-GMP (c-Di-GMP) Signaling: Lessons from Pseudomonas aeruginosa and Other Bacteria. J. Biol. Chem. 2016, 291, 12547–12555. [Google Scholar] [CrossRef] [Green Version]
- Strempel, N.; Nusser, M.; Neidig, A.; Brenner-Weiss, G.; Overhage, J. The Oxidative Stress Agent Hypochlorite Stimulates C-Di-GMP Synthesis and Biofilm Formation in Pseudomonas Aeruginosa. Front. Microbiol. 2017, 8, 2311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poudyal, B.; Sauer, K. The PA3177 Gene Encodes an Active Diguanylate Cyclase That Contributes to Biofilm Antimicrobial Tolerance but Not Biofilm Formation by Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2018, 62, e01049-18. [Google Scholar] [CrossRef] [Green Version]
- Xia, B.; Li, M.; Tian, Z.; Chen, G.; Liu, C.; Xia, Y.; Jin, Y.; Bai, F.; Cheng, Z.; Jin, S.; et al. Oligoribonuclease Contributes to Tolerance to Aminoglycoside and β-Lactam Antibiotics by Regulating KatA in Pseudomonas Aeruginosa. Antimicrob. Agents Chemother. 2019, 63, e00212-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Chen, G.; Liu, Y.; Jin, Y.; Cheng, Z.; Liu, Y.; Yang, L.; Jin, S.; Wu, W. Pseudomonas aeruginosa Oligoribonuclease Contributes to Tolerance to Ciprofloxacin by Regulating Pyocin Biosynthesis. Antimicrob. Agents Chemother. 2017, 61, e02256-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldman, S.R.; Sharp, J.S.; Vvedenskaya, I.O.; Livny, J.; Dove, S.L.; Nickels, B.E. NanoRNAs Prime Transcription Initiation In Vivo. Mol. Cell 2011, 42, 817–825. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Zhao, Q.; Zhu, F.; Chen, R.; Jin, Y.; Liu, C.; Pan, X.; Jin, S.; Wu, W.; Cheng, Z. Oligoribonuclease Is Required for the Type III Secretion System and Pathogenesis of Pseudomonas Aeruginosa. Microbiol. Res. 2016, 188–189, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Orr, M.W.; Donaldson, G.P.; Severin, G.B.; Wang, J.; Sintim, H.O.; Waters, C.M.; Lee, V.T. Oligoribonuclease Is the Primary Degradative Enzyme for PGpG in Pseudomonas aeruginosa That Is Required for Cyclic-Di-GMP Turnover. Proc. Natl. Acad. Sci. USA 2015, 112, E5048–E5057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balasubramanian, D.; Kumari, H.; Mathee, K. Pseudomonas aeruginosa AmpR: An Acute-Chronic Switch Regulator. Pathog. Dis. 2015, 73, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhury, N.; Nur, I.; Noor, R. Comparative Study of Oxidative Stress Response in Pseudomonas aeruginosa (Subp01) and Pseudomonas Fluorescence (Subp02) Upon External Stimulation by Hydrogen Peroxide. EC Microbiol. 2019, 15, 653–660. [Google Scholar]
- Kiewitz, C.; Tümmler, B. Sequence Diversity of Pseudomonas aeruginosa: Impact on Population Structure and Genome Evolution. J. Bacteriol. 2000, 182, 3125–3135. [Google Scholar] [CrossRef] [Green Version]
- Frimmersdorf, E.; Horatzek, S.; Pelnikevich, A.; Wiehlmann, L.; Schomburg, D. How Pseudomonas aeruginosa Adapts to Various Environments: A Metabolomic Approach. Environ. Microbiol. 2010, 12, 1734–1747. [Google Scholar] [CrossRef]
- Moradali, M.F.; Ghods, S.; Rehm, B.H.A. Pseudomonas aeruginosa Lifestyle: A Paradigm for Adaptation, Survival, and Persistence. Front. Cell. Infect. Microbiol. 2017, 7, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. WHO Publishes List of Bacteria for Which New Antibiotics Are Urgently Needed. Available online: https://www.who.int/news/item/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (accessed on 5 May 2021).
- Skindersoe, M.E.; Alhede, M.; Phipps, R.; Yang, L.; Jensen, P.O.; Rasmussen, T.B.; Bjarnsholt, T.; Tolker-Nielsen, T.; Høiby, N.; Givskov, M. Effects of Antibiotics on Quorum Sensing in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2008, 52, 3648–3663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wozniak, D.J.; Keyser, R. Effects of Subinhibitory Concentrations of Macrolide Antibiotics on Pseudomonas aeruginosa. Chest 2004, 125, 62S–69S. [Google Scholar] [CrossRef]
- Puckett, S.; Trujillo, C.; Wang, Z.; Eoh, H.; Ioerger, T.R.; Krieger, I.; Sacchettini, J.; Schnappinger, D.; Rhee, K.Y.; Ehrt, S. Glyoxylate Detoxification Is an Essential Function of Malate Synthase Required for Carbon Assimilation in Mycobacterium Tuberculosis. Proc. Natl. Acad. Sci. USA 2017, 114, E2225–E2232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Series of reactions leading to the production of (A) reactive oxygen species (ROS) from molecular oxygen (O2) and (B) hypochlorous acid (HOCl). (C) Fenton reaction between ferrous iron (Fe2+) and hydrogen peroxide (H2O2) leading to the production of hydroxyl radical (HO•). Superoxide: O2−, hydrogen peroxide: H2O2, hydroxyl radicals: HO•, H2O: water, e−: electron, H+: hydrogen ion. This figure was created with BioRender.com (accessed on 10 September 2021).
Figure 1.
Series of reactions leading to the production of (A) reactive oxygen species (ROS) from molecular oxygen (O2) and (B) hypochlorous acid (HOCl). (C) Fenton reaction between ferrous iron (Fe2+) and hydrogen peroxide (H2O2) leading to the production of hydroxyl radical (HO•). Superoxide: O2−, hydrogen peroxide: H2O2, hydroxyl radicals: HO•, H2O: water, e−: electron, H+: hydrogen ion. This figure was created with BioRender.com (accessed on 10 September 2021).

Figure 2.
Oxidation of (A) cysteine (Cys) and (B) methionine (Met) residues on proteins by reactive oxygen species (ROS). GSH: glutathione, Met-O: Methionine sulfoxide, Met-O2: methionine sulfone. This figure was created with BioRender.com (accessed on 10 September 2021).
Figure 2.
Oxidation of (A) cysteine (Cys) and (B) methionine (Met) residues on proteins by reactive oxygen species (ROS). GSH: glutathione, Met-O: Methionine sulfoxide, Met-O2: methionine sulfone. This figure was created with BioRender.com (accessed on 10 September 2021).

Figure 3.
Bacterial targets of reactive oxygen species (ROS) and hypochlorous acid (HOCl), a reactive chlorine species (RCS) with potent antimicrobial effect. (A) Lipids represent a common oxidative target for both ROS and HOCl. Both oxidative agents can also induce lipid peroxidation. (B) Sulfur-containing side chains of methionine and cysteine are particularly prone to oxidation by ROS and RCS. (C) ROS and HOCl actively react with Fe–S clusters and heme proteins in enzymes, which often leads to the release of intracellular iron. (D) H2O2 and HOCl can react with the intracellular iron released after oxidation of Fe–S clusters. This reaction produces HO• that oxidatively damage DNA. (E) HOCl can directly damage DNA through deamination, oxidation, and formation of single-stranded breaks and cross-links. (F) HOCl can also inhibit DNA and protein synthesis by the inactivation of proteins. (G) HOCl can also interfere with the electron transport chain, decreasing energy production. H2O2: hydrogen peroxide; HO•: hydroxyl radicals. This figure was created with BioRender.com (accessed on 10 September 2021).
Figure 3.
Bacterial targets of reactive oxygen species (ROS) and hypochlorous acid (HOCl), a reactive chlorine species (RCS) with potent antimicrobial effect. (A) Lipids represent a common oxidative target for both ROS and HOCl. Both oxidative agents can also induce lipid peroxidation. (B) Sulfur-containing side chains of methionine and cysteine are particularly prone to oxidation by ROS and RCS. (C) ROS and HOCl actively react with Fe–S clusters and heme proteins in enzymes, which often leads to the release of intracellular iron. (D) H2O2 and HOCl can react with the intracellular iron released after oxidation of Fe–S clusters. This reaction produces HO• that oxidatively damage DNA. (E) HOCl can directly damage DNA through deamination, oxidation, and formation of single-stranded breaks and cross-links. (F) HOCl can also inhibit DNA and protein synthesis by the inactivation of proteins. (G) HOCl can also interfere with the electron transport chain, decreasing energy production. H2O2: hydrogen peroxide; HO•: hydroxyl radicals. This figure was created with BioRender.com (accessed on 10 September 2021).

Figure 4.
Simplified schematic overview of the interconnected oxidative stress responses adopted by Pseudomonas aeruginosa. (A) Production of pigments. (B) Transcriptional regulators activated by oxidizing agents: OxyR, which has been shown to regulate the expression of several phenotypes, such as the formation of biofilm and induction of small RNAs, in addition to the induction of genes involved in oxidative detoxification (katB, ahpCF, and ahpB); Fur and IscR, which play roles in the expression of detoxifying enzymes; OspR and OhrR, which are homolog regulators that present interconnected activity; and SoxR, which has been shown to activate six genes that are in three transcriptional subunits (PA3718, which encodes an efflux pump of the multiple facilitator superfamily; PA2274, an unknown protein; and four genes in the mexGHI-ompD operon). (C) Activation of chaperones and the Methionine sulfoxide reductase (Msr) system. (D) Antibiotic resistance due to increased mutation frequency, horizontal gene transfer, expression of efflux pumps, and persister state. (E) Induction of quorum sensing genes, which are involved in rhamnolipid production and detoxifying enzymes. (F) Detoxification of toxic oxygen and chlorine species by antioxidant enzymes, such as catalases (katA and katB), peroxidases, superoxide dismutase (sodM and sodB), and alkyl hydroperoxide reductase (ahpA, ahpB, ahpCF, ohr). (G) Genes involved in carbon metabolism, more specifically in the glyoxylate shunt and the Entner–Doudoroff pathaway were involved in P. aeruginosa survival under oxidative stress. (H) Methylation and induction of small RNAs. (I) Induction of the mucoid phenotype due to the overexpression of alginate. Msr: Methionine sulfoxide reductase; SOD: superoxide dismutase; ahp: alkyl hydroperoxide reductase. This figure was created with BioRender.com (accessed on 10 September 2021).
Figure 4.
Simplified schematic overview of the interconnected oxidative stress responses adopted by Pseudomonas aeruginosa. (A) Production of pigments. (B) Transcriptional regulators activated by oxidizing agents: OxyR, which has been shown to regulate the expression of several phenotypes, such as the formation of biofilm and induction of small RNAs, in addition to the induction of genes involved in oxidative detoxification (katB, ahpCF, and ahpB); Fur and IscR, which play roles in the expression of detoxifying enzymes; OspR and OhrR, which are homolog regulators that present interconnected activity; and SoxR, which has been shown to activate six genes that are in three transcriptional subunits (PA3718, which encodes an efflux pump of the multiple facilitator superfamily; PA2274, an unknown protein; and four genes in the mexGHI-ompD operon). (C) Activation of chaperones and the Methionine sulfoxide reductase (Msr) system. (D) Antibiotic resistance due to increased mutation frequency, horizontal gene transfer, expression of efflux pumps, and persister state. (E) Induction of quorum sensing genes, which are involved in rhamnolipid production and detoxifying enzymes. (F) Detoxification of toxic oxygen and chlorine species by antioxidant enzymes, such as catalases (katA and katB), peroxidases, superoxide dismutase (sodM and sodB), and alkyl hydroperoxide reductase (ahpA, ahpB, ahpCF, ohr). (G) Genes involved in carbon metabolism, more specifically in the glyoxylate shunt and the Entner–Doudoroff pathaway were involved in P. aeruginosa survival under oxidative stress. (H) Methylation and induction of small RNAs. (I) Induction of the mucoid phenotype due to the overexpression of alginate. Msr: Methionine sulfoxide reductase; SOD: superoxide dismutase; ahp: alkyl hydroperoxide reductase. This figure was created with BioRender.com (accessed on 10 September 2021).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
da Cruz Nizer, W.S.; Inkovskiy, V.; Versey, Z.; Strempel, N.; Cassol, E.; Overhage, J. Oxidative Stress Response in Pseudomonas aeruginosa. Pathogens 2021, 10, 1187. https://doi.org/10.3390/pathogens10091187
AMA Style
da Cruz Nizer WS, Inkovskiy V, Versey Z, Strempel N, Cassol E, Overhage J. Oxidative Stress Response in Pseudomonas aeruginosa. Pathogens. 2021; 10(9):1187. https://doi.org/10.3390/pathogens10091187
Chicago/Turabian Styleda Cruz Nizer, Waleska Stephanie, Vasily Inkovskiy, Zoya Versey, Nikola Strempel, Edana Cassol, and Joerg Overhage. 2021. "Oxidative Stress Response in Pseudomonas aeruginosa" Pathogens 10, no. 9: 1187. https://doi.org/10.3390/pathogens10091187
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.