Intracellular Environment Improvement of Mycobacterium neoaurum for Enhancing Androst-1,4-Diene-3,17-Dione Production by Manipulating NADH and Reactive Oxygen Species Levels


Abstract
:1. Introduction
2. Results and Discussion
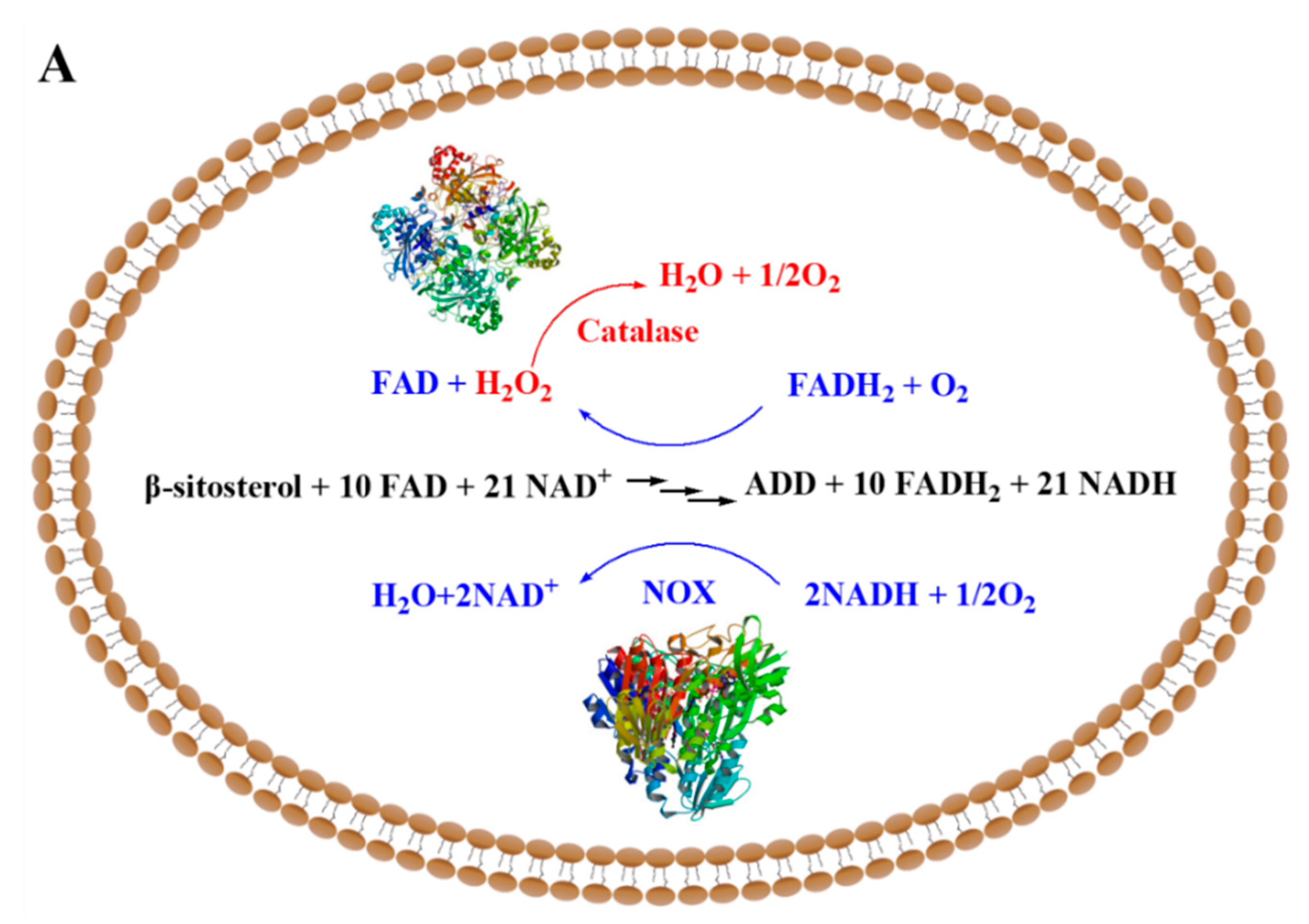
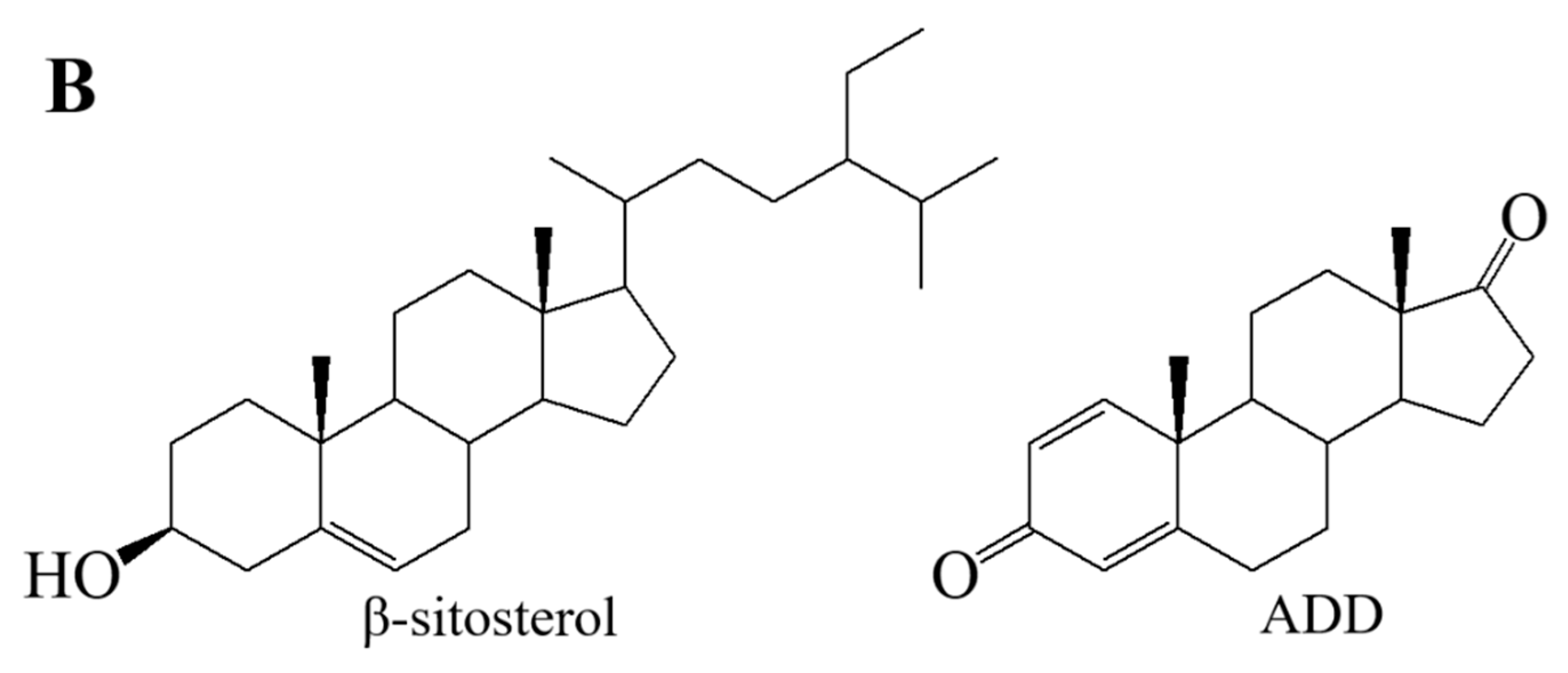
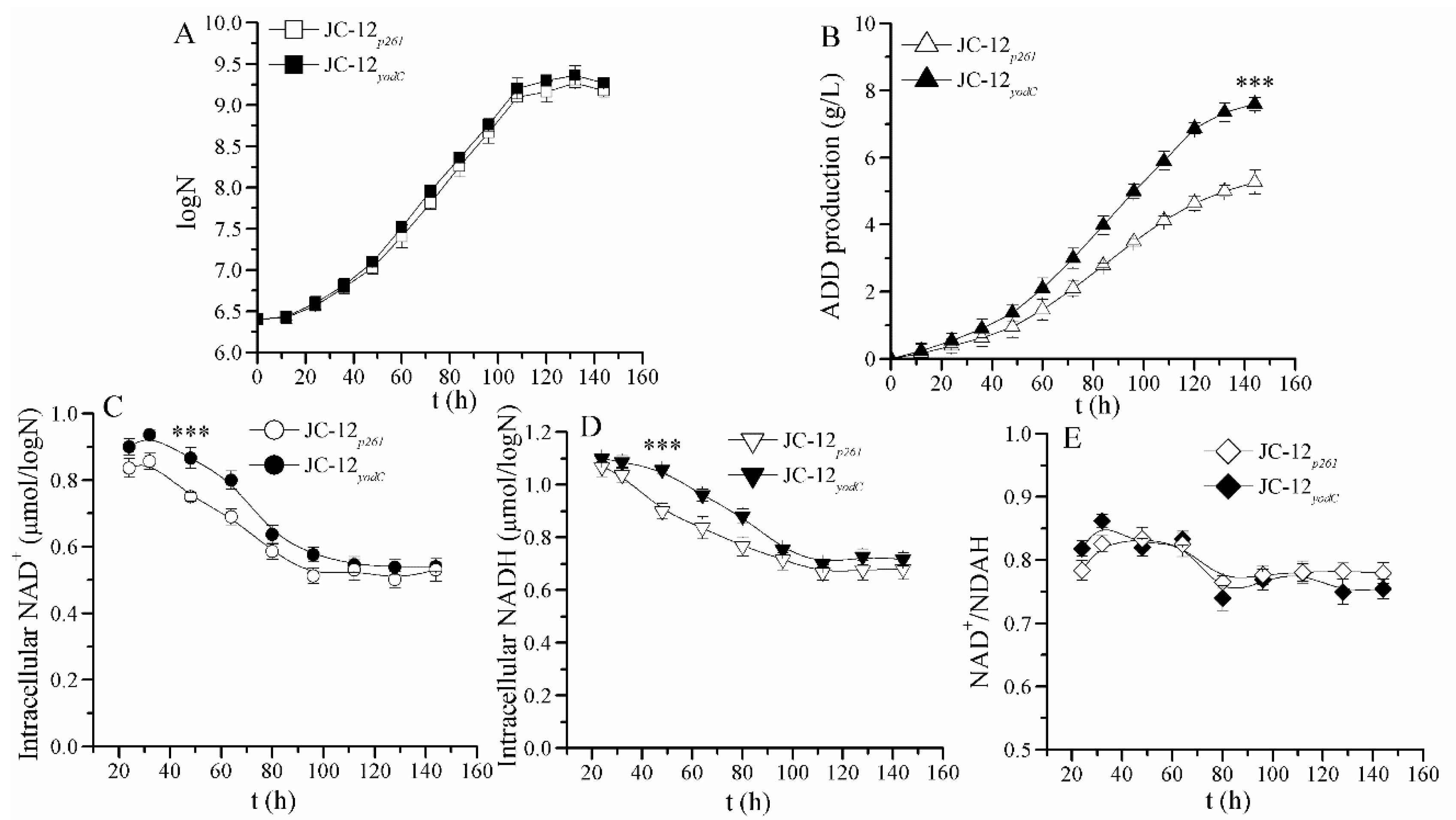
2.1. The Increase in NAD+ Availability by Expressing NOX Resulted in an Improved ADD Yield in the NAD+ Regeneration System of JC-12yodc
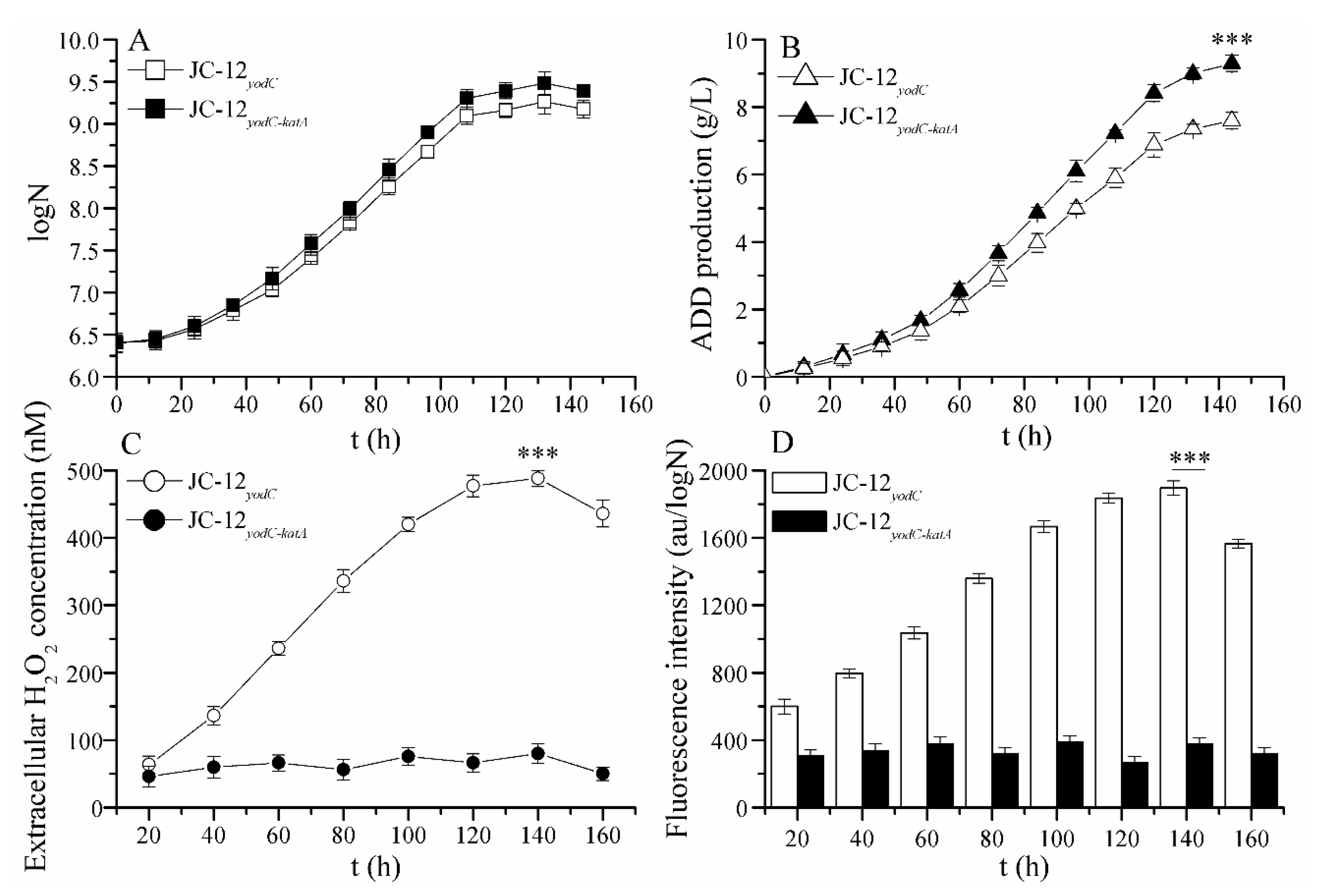
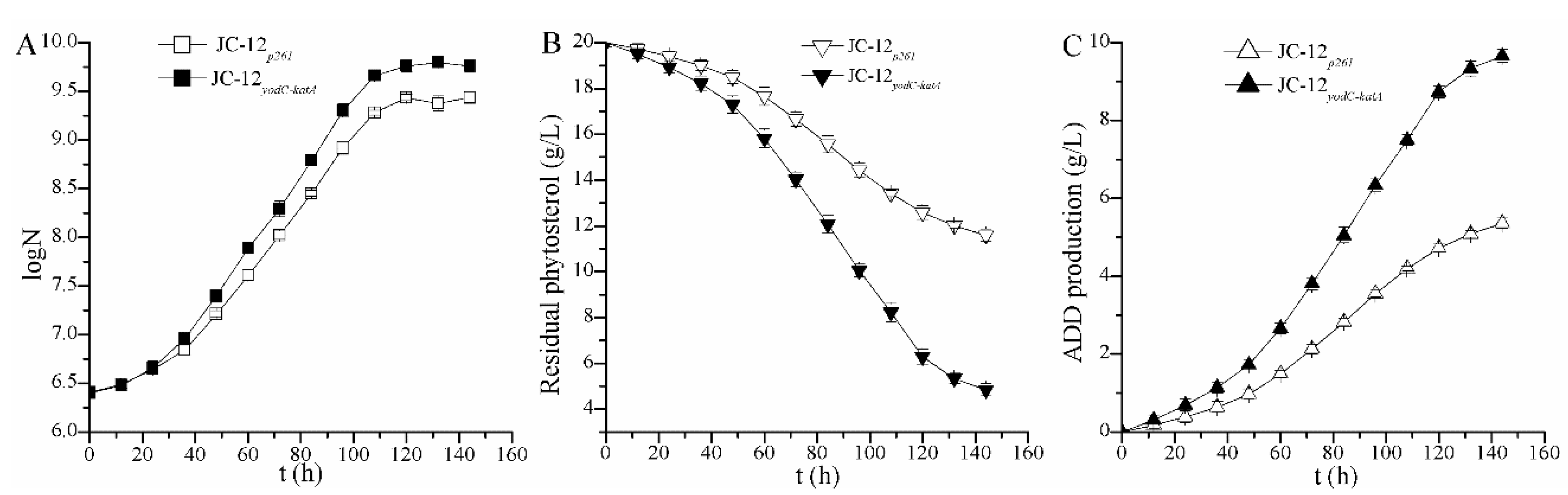
2.2. The Over-Expression of Catalase Eliminated the Toxic Effect of H2O2 Accumulation on Strain Growth and ADD Production
3. Materials and Methods
3.1. Strains and Culture Conditions
3.2. Over-Expression of NOX and Catalase in M. Neoaurum JC-12
3.3. NOX and Catalase Enzyme Activity Assays
3.4. Analytical Methods
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AD | 4-androstene-3,17-dione |
| ADD | androst-1,4-diene-3,17-dione |
| CFU | colony forming units |
| HP-β-CD | hydroxymethyl-β-cyclodextrin |
| H2O2 | hydrogen peroxide |
| ROS | reactive oxygen species |
| NOX | NADH oxidase |
| DCFH-DA | 2′,7′-dichlorofluorescein diacetate |
| GC | gas chromatography |
| HPLC | high performance liquid chromatography |
References
- Donova, M. Microbiotechnologies for steroid production. Microbiol. Aust. 2018, 39, 126–129. [Google Scholar] [CrossRef]
- Malaviya, A.; Gomes, J. Androstenedione production by biotransformation of phytosterols. Bioresour. Technol. 2008, 99, 6725–6737. [Google Scholar] [CrossRef] [PubMed]
- Shao, M.L.; Zhang, X.; Rao, Z.M.; Xu, M.J.; Yang, T.W.; Li, H.; Xu, Z.H.; Yang, S.T. Efficient testosterone production by engineered Pichia pastoris co-expressing human 17β-hydroxysteroid dehydrogenase type 3 and Saccharomyces cerevisiae glucose 6-phosphate dehydrogenase with NADPH regeneration. Green Chem. 2016, 18, 1774–1784. [Google Scholar] [CrossRef]
- Bao, T.; Zhang, X.; Rao, Z.M.; Zhao, X.J.; Zhang, R.Z.; Yang, T.W.; Xu, Z.H.; Yang, S.T. Efficient whole-cell biocatalyst for acetoin production with NAD+ regeneration system through homologous co-expression of 2,3-butanediol dehydrogenase and NADH oxidase in engineered Bacillus subtilis. PLoS ONE 2014, 9, e102951. [Google Scholar] [CrossRef]
- Donova, M.V.; Egorova, O.V. Microbial steroid transformations: Current state and prospects. Appl. Microbiol. Biotechnol. 2012, 94, 1423–1447. [Google Scholar] [CrossRef]
- Josefsen, K.D.; Nordborg, A.; Sletta, H. Bioconversion of Phytosterols into Androstenedione by Mycobacterium. In Microbial Steroids; Springer: New York, NY, USA, 2017; pp. 177–197. [Google Scholar]
- Van der Geize, R.; Yam, K.; Heuser, T.; Wilbrink, M.H.; Hara, H.; Anderton, M.C.; Sim, E.; Dijkhuizen, L.; Davies, J.E.; Mohn, W.W.; et al. A gene cluster encoding cholesterol catabolism in a soil actinomycete provides insight into Mycobacterium tuberculosis survival in macrophages. Proc. Natl. Acad. Sci. USA 2007, 104, 1947–1952. [Google Scholar] [CrossRef]
- Fernández-Cabezón, L.; Galán, B.; García, J.L. New insights on steroid biotechnology. Front. Microbiol. 2018, 9, 958. [Google Scholar] [CrossRef]
- Wei, W.; Wang, F.Q.; Fan, S.Y.; Wei, D.Z. Inactivation and augmentation of the primary 3-ketosteroid-Δ1-dehydrogenase in Mycobacterium neoaurum NwIB-01: Biotransformation of soybean phytosterols to 4-androstene-3,17-dione or 1,4-androstadiene-3,17-dione. Appl. Environ. Microbiol. 2010, 76, 4578–4582. [Google Scholar] [CrossRef]
- Yao, K.; Wang, F.Q.; Zhang, H.C.; Wei, D.Z. Identification and engineering of cholesterol oxidases involved in the initial step of sterols catabolism in Mycobacterium neoaurum. Metab. Eng. 2013, 15, 75–87. [Google Scholar] [CrossRef]
- Wei, W.; Fan, S.Y.; Wang, F.Q.; Wei, D.Z. Accumulation of androstadiene-dione by overexpression of heterologous 3-ketosteroid Δ1-dehydrogenase in Mycobacterium neoaurum NwIB-01. World J. Microbiol. Biotechnol. 2014, 30, 1947–1954. [Google Scholar] [CrossRef]
- Su, L.; Shen, Y.; Zhang, W.; Gao, T.; Shang, Z.; Wang, M. Cofactor engineering to regulate NAD+/NADH ratio with its application to phytosterols biotransformation. Microb. Cell Fact. 2017, 16, 182. [Google Scholar] [CrossRef] [PubMed]
- Shao, M.; Zhang, X.; Rao, Z.; Xu, M.; Yang, T.; Xu, Z.; Yang, S. Identification of steroid C27 monooxygenase isoenzymes involved in sterol catabolism and stepwise pathway engineering of Mycobacterium neoaurum for improved androst-1,4-diene-3,17-dione production. J. Ind. Microbiol. Biotechnol. 2019, 46, 635–647. [Google Scholar] [CrossRef] [PubMed]
- Man, Z.; Rao, Z.; Xu, M.; Guo, J.; Yang, T.; Zhang, X.; Xu, Z. Improvement of the intracellular environment for enhancing L-arginine production of Corynebacterium glutamicum by inactivation of H2O2-forming flavin reductases and optimization of ATP supply. Metab. Eng. 2016, 38, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Angermayr, S.A.; Rovira, A.G.; Hellingwerf, K.J. Metabolic engineering of cyanobacteria for the synthesis of commodity products. Trends Biotechnol. 2015, 33, 352–361. [Google Scholar] [CrossRef] [PubMed]
- Balzer, G.J.; Thakker, C.; Bennett, G.N.; San, K.-Y. Metabolic engineering of Escherichia coli to minimize byproduct formate and improving succinate productivity through increasing NADH availability by heterologous expression of NAD+-dependent formate dehydrogenase. Metab. Eng. 2013, 20, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bao, T.; Zhang, X.; Zhao, X.; Rao, Z.; Yang, T.; Yang, S. Regulation of the NADH pool and NADH/NADPH ratio redistributes acetoin and 2,3-butanediol proportion in Bacillus subtilis. Biochem. J. 2015, 10, 1298–1306. [Google Scholar]
- Szentirmai, A. Microbial physiology of sidechain degradation of sterols. J. Ind. Microbiol. Biotechnol. 1990, 6, 101–115. [Google Scholar] [CrossRef]
- Shao, M.; Sha, Z.; Zhang, X.; Rao, Z.; Xu, M.; Yang, T.; Xu, Z.; Yang, S. Efficient androst-1,4-diene-3,17-dione production by co-expressing 3-ketosteroid-Δ1-dehydrogenase and catalase in Bacillus subtilis. J. Appl. Microbiol. 2017, 122, 119–128. [Google Scholar] [CrossRef]
- Shao, M.L.; Zhang, X.; Rao, Z.M.; Xu, M.; Yang, T.; Li, H.; Xu, Z.; Yang, S. A mutant form of 3-ketosteroid-Δ1-dehydrogenase gives altered androst-1,4-diene-3, 17-dione/androst-4-ene-3,17-dione molar ratios in steroid biotransformations by Mycobacterium neoaurum ST-095. J. Ind. Microbiol. Biotechnol. 2016, 43, 691–701. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, R.Z.; Bao, T.; Rao, Z.M.; Yang, T.W.; Xu, M.J.; Xu, Z.H.; Li, H.Z.; Yang, S.T. The rebalanced pathway significantly enhances acetoin production by disruption of acetoin reductase gene and moderate-expression of a new water-forming NADH oxidase in Bacillus subtilis. Metab. Eng. 2014, 23, 34–41. [Google Scholar] [CrossRef]
- Ezraty, B.; Gennaris, A.; Barras, F.; Collet, J.F. Oxidative stress, protein damage and repair in bacteria. Nat. Rev. Microbiol. 2017, 15, 385. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Imlay, J.A. Cell death from antibiotics without the involvement of reactive oxygen species. Science 2013, 339, 1210–1213. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, J.M.; Kudej, R.K.; LaNoue, K.F.; Vatner, S.F.; Lewandowski, E.D. Limited transfer of cytosolic NADH into mitochondria at high cardiac workload. Am. J. Physiol. Heart Circ. Physiol. 2004, 286, H2237–H2242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Si, M.R.; Zhang, L.; Yang, Z.F.; Xu, Y.X.; Liu, Y.B.; Jiang, C.Y.; Wang, Y.; Shen, X.H.; Liu, S.J. NrdH redoxin enhances resistance to multiple oxidative stresses by acting as a peroxidase cofactor in Corynebacterium glutamicum. Appl. Environ. Microbiol. 2014, 80, 1750–1762. [Google Scholar] [CrossRef] [PubMed]
- Si, M.; Xu, Y.; Wang, T.; Long, M.; Ding, W.; Chen, C.; Guan, X.; Liu, Y.; Wang, Y.; Shen, X. Functional characterization of a mycothiol peroxidase in Corynebacterium glutamicum that uses both mycoredoxin and thioredoxin reducing systems in the response to oxidative stress. Biochem. J. 2015, 469, 45–57. [Google Scholar] [CrossRef]
- Zhang, W.Q.; Shao, M.L.; Rao, Z.M.; Xu, M.J.; Zhang, X.; Yang, T.W.; Li, H.; Xu, Z.H. Bioconversion of 4-androstene-3,17-dione to androst-1,4-diene-3,17-dione by recombinant Bacillus subtilis expressing ksdd gene encoding 3-ketosteroid-Δ1-dehydrogenase from Mycobacterium neoaurum JC-12. J. Steroid Biochem. Mol. Biol. 2013, 135, 36–42. [Google Scholar] [CrossRef]
- Molchanova, M.A.; Andryushina, V.A.; Savinova, T.S.; Stytsenko, T.S.; Rodina, N.V.; Voishvillo, N.E. Preparation of androsta-1,4-diene-3,17-dione from sterols using Mycobacterium neoaurum VKPM Ac-1656 strain. Russ. J. Bioorganic. Chem. 2007, 33, 354–358. [Google Scholar] [CrossRef]
- Duong, S.; Strobel, N.; Buddhadasa, S.; Stockham, K.; Auldist, M.; Wales, B.; Orbell, J.; Cran, M. Rapid measurement of phytosterols in fortified food using gas chromatography with flame ionization detection. Food Chem. 2016, 211, 570–576. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Enzyme Activity * | |
|---|---|---|
| NOX (mU/mg) | Catalase (U/mg) | |
| JC-12p261 | 25.6 ± 1.5 | 27 ± 3 |
| JC-12yodC | 337.2 ± 9.6 | 23 ± 5 |
| JC-12yodC-katA | 312.3 ± 6.3 | 235 ± 6 |
| Strains/Plasmids/Primers | Description | Sources |
|---|---|---|
| Strains | ||
| Escherichia coli | ||
| JM109 | General host for gene cloning | Invitrogen, (Carlsbad, CA, USA) |
| Mycobacterium neoaurum | ||
| JC-12 | Wild type strain, converting sterols to ADD with small amount of AD | Lab storage, (Wuxi, China) |
| JC-12p261 | JC-12 harboring empty plasmid pMV261 | This study |
| JC-12yodC | NOX over-expressed strain of JC-12, harboring plasmid pMV261-yodC | This study |
| JC-12yodC-katA | Catalase over-expressed strain of JC-12yodC, harboring plasmid pMV261-yodC-katA | This study |
| Plasmids | ||
| pMD18-T | E. coli clone vector; AmpR | Novagen, (Madison, WI, USA) |
| pMV261 | Shuttle vector of E. coli and mycobacterium, carrying the heat shock promoter hsp60; KanR | R. Jacobs Jr. |
| pMV261-yodC | pMV261 carrying yodC gene; KanR | This study |
| pMV261-yodC-katA | pMV261-yodC carrying katA gene with its SD sequence inserted after yodC; KanR | This study |
| Primers | ||
| yodC-f | CGGGATCCATGACGAATACTCTGGATG | This study |
| yodC-r | CGGAATTCTTACAGCCAAGTTGATAC | This study |
| katA-SD-f | ACGAAGCTTaagaaggagatataATGCGCGAAAGGAACAGCCC | This study |
| katA-r | ACGAAGCTTCTACTTGACGGCCGCCTC | This study |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shao, M.; Zhao, Y.; Liu, Y.; Yang, T.; Xu, M.; Zhang, X.; Rao, Z. Intracellular Environment Improvement of Mycobacterium neoaurum for Enhancing Androst-1,4-Diene-3,17-Dione Production by Manipulating NADH and Reactive Oxygen Species Levels. Molecules 2019, 24, 3841. https://doi.org/10.3390/molecules24213841
Shao M, Zhao Y, Liu Y, Yang T, Xu M, Zhang X, Rao Z. Intracellular Environment Improvement of Mycobacterium neoaurum for Enhancing Androst-1,4-Diene-3,17-Dione Production by Manipulating NADH and Reactive Oxygen Species Levels. Molecules. 2019; 24(21):3841. https://doi.org/10.3390/molecules24213841
Chicago/Turabian StyleShao, Minglong, Youxi Zhao, Yu Liu, Taowei Yang, Meijuan Xu, Xian Zhang, and Zhiming Rao. 2019. "Intracellular Environment Improvement of Mycobacterium neoaurum for Enhancing Androst-1,4-Diene-3,17-Dione Production by Manipulating NADH and Reactive Oxygen Species Levels" Molecules 24, no. 21: 3841. https://doi.org/10.3390/molecules24213841