
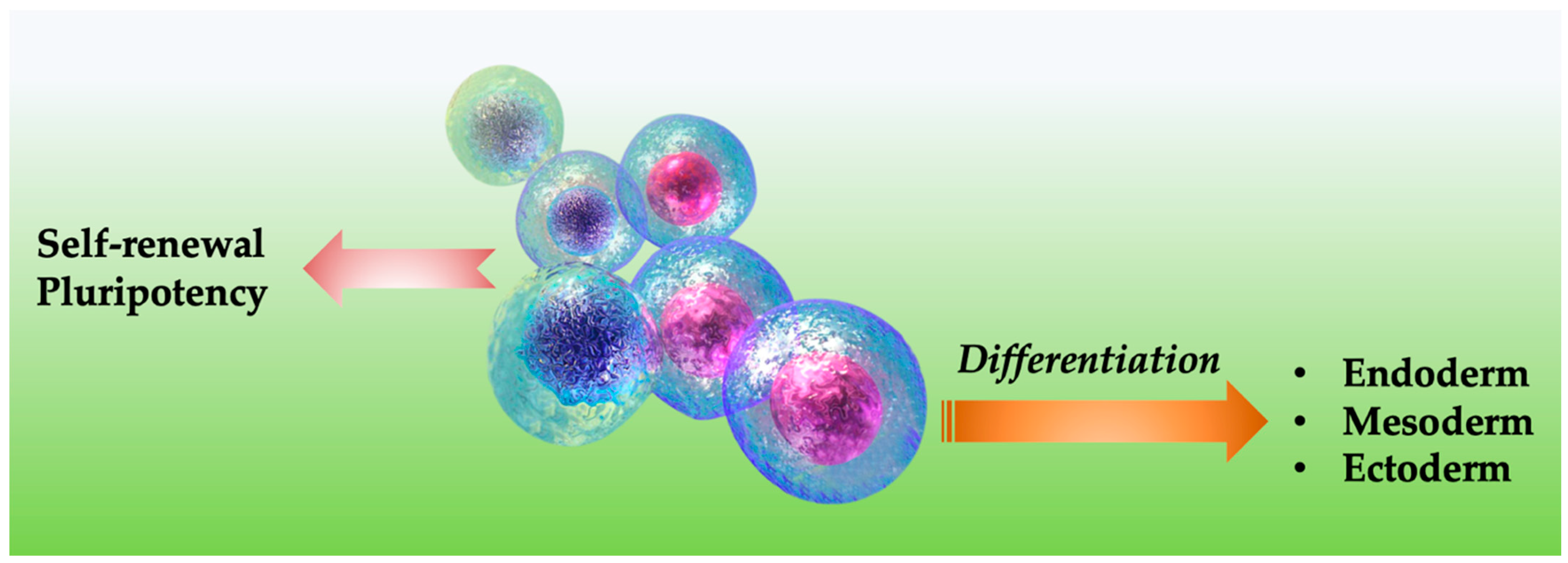
Molecular Mechanisms Underlying Pluripotency and Self-Renewal of Embryonic Stem Cells


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
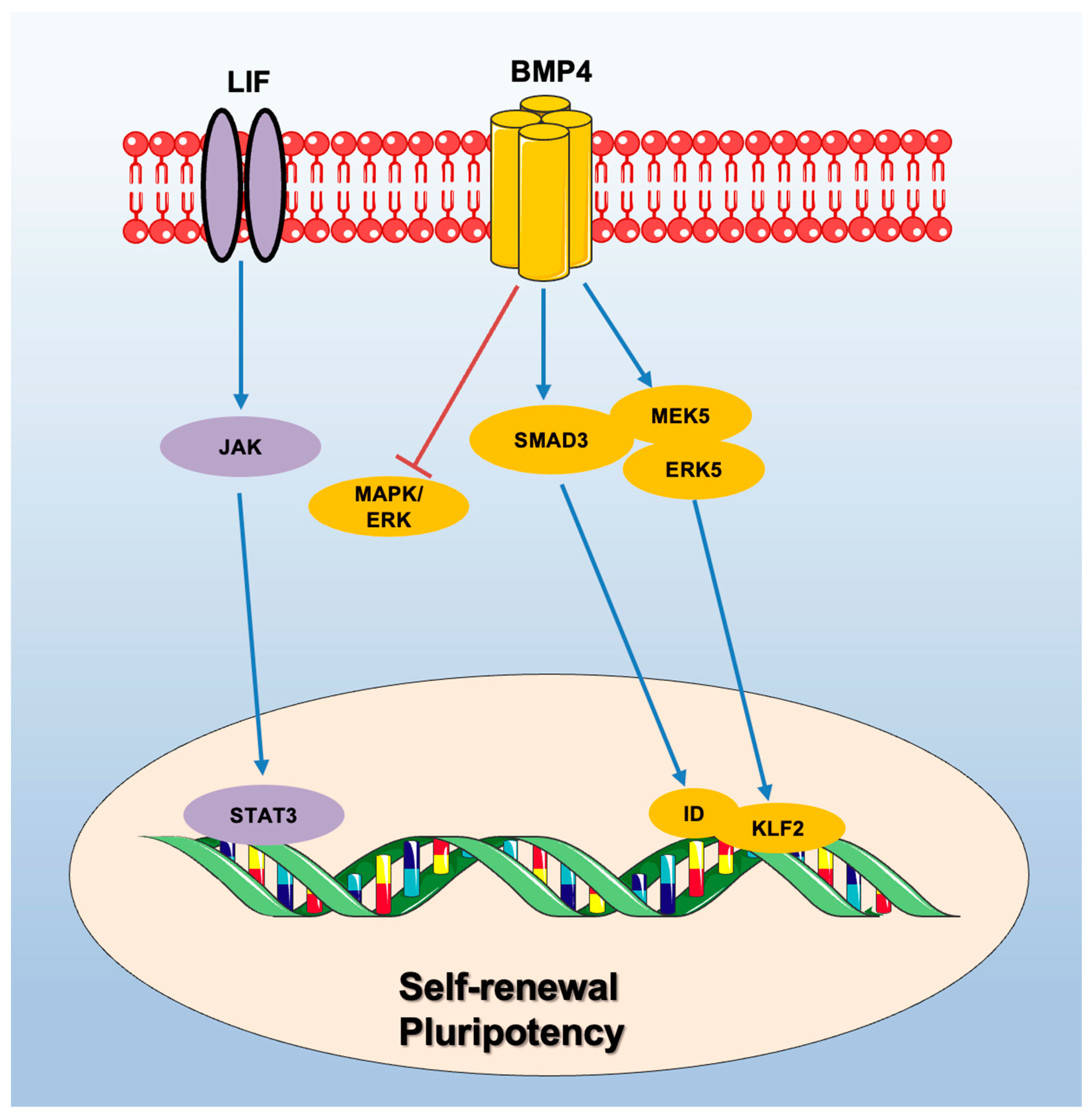
2. Murine ESCs: Signaling Pathways Supporting Self-Renewal and Pluripotency
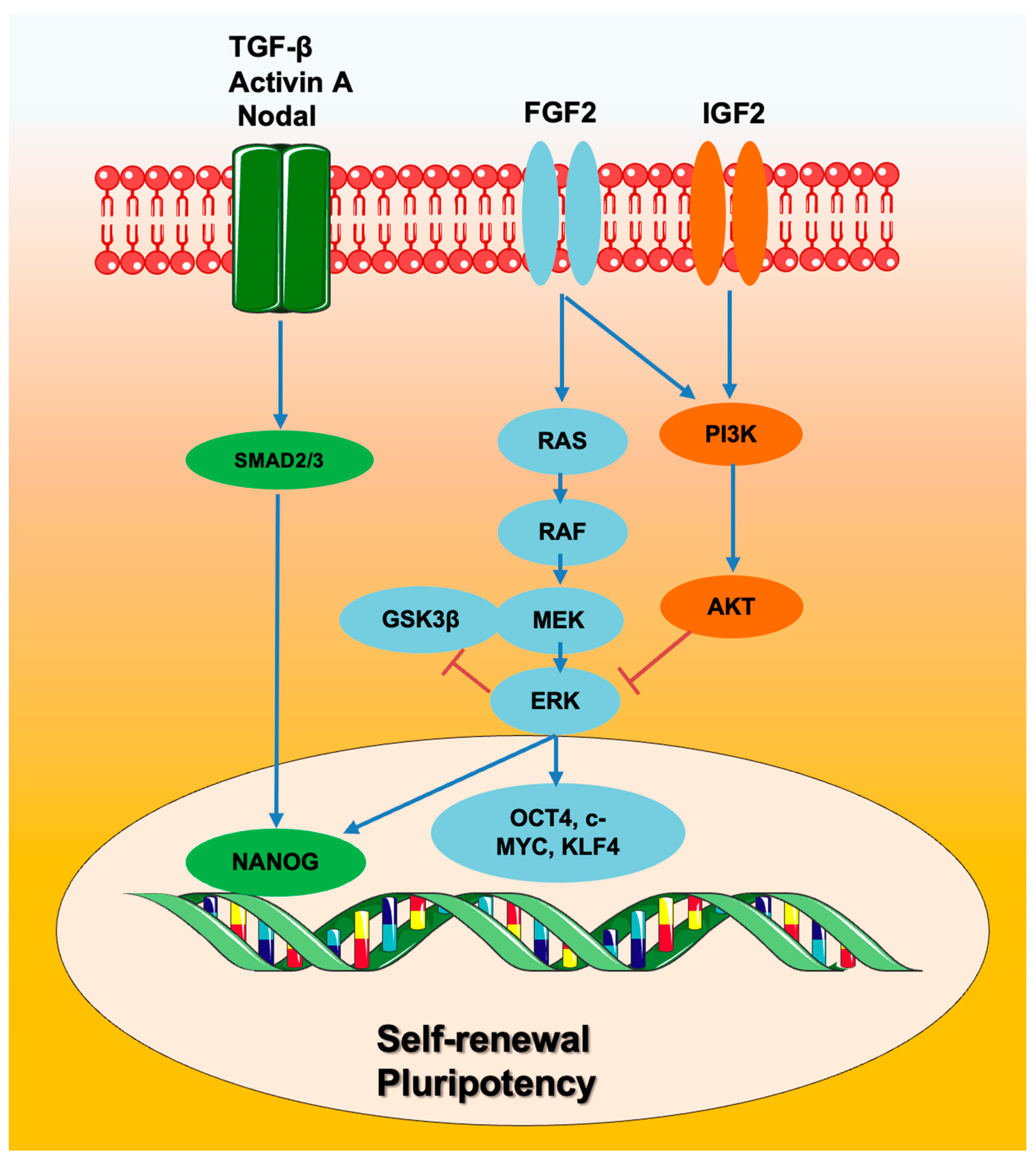
3. Human ESCs: Signaling Pathways Involved in Self-Renewal and Pluripotency
4. Epigenetic and Chromatin Modifications of Self-Renewal and Pluripotency in ESCs
5. Naïve Pluripotency
6. Controversial Issues
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Balboa, D.; Iworima, D.G.; Kieffer, T.J. Human Pluripotent Stem Cells to Model Islet Defects in Diabetes. Front. Endocrinol. 2021, 12, 642152. [Google Scholar] [CrossRef]
- Hanna, J.H.; Saha, K.; Jaenisch, R. Pluripotency and Cellular Reprogramming: Facts, Hypotheses, Unresolved Issues. Cell 2010, 143, 508–525. [Google Scholar] [CrossRef]
- Bogliotti, Y.S.; Wu, J.; Vilarino, M.; Okamura, D.; Soto, D.A.; Zhong, C.; Sakurai, M.; Sampaio, R.V.; Suzuki, K.; Belmonte, J.C.I.; et al. Efficient derivation of stable primed pluripotent embryonic stem cells from bovine blastocysts. Proc. Natl. Acad. Sci. USA 2018, 115, 2090–2095. [Google Scholar] [CrossRef]
- Thomson, J.A.; Itskovitz-Eldor, J.; Shapiro, S.S.; Waknitz, M.A.; Swiergiel, J.J.; Marshall, V.S.; Jones, J.M. Embryonic Stem Cell Lines Derived from Human Blastocysts. Science 1998, 282, 1145–1147. [Google Scholar] [CrossRef]
- Tachibana, M.; Amato, P.; Sparman, M.; Gutierrez, N.M.; Tippner-Hedges, R.; Ma, H.; Kang, E.; Fulati, A.; Lee, H.-S.; Sritanaudomchai, H.; et al. Human Embryonic Stem Cells Derived by Somatic Cell Nuclear Transfer. Cell 2013, 153, 1228–1238. [Google Scholar] [CrossRef]
- Dekel, C.; Morey, R.; Hanna, J.; Laurent, L.C.; Ben-Yosef, D.; Amir, H. Stabilization of hESCs in two distinct substates along the continuum of pluripotency. iScience 2022, 25, 105469. [Google Scholar] [CrossRef]
- Martin, G.R. Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells. Proc. Natl. Acad. Sci. USA 1981, 78, 7634–7638. [Google Scholar] [CrossRef]
- Evans, M.J.; Kaufman, M.H. Establishment in culture of pluripotential cells from mouse embryos. Nature 1981, 292, 154–156. [Google Scholar] [CrossRef]
- Plusa, B.; Hadjantonakis, A.-K. Embryonic stem cell identity grounded in the embryo. Nature 2014, 16, 502–504. [Google Scholar] [CrossRef]
- Czechanski, A.; Byers, C.; Greenstein, I.; Schrode, N.; Donahue, L.R.; Hadjantonakis, A.-K.; Reinholdt, L.G. Derivation and characterization of mouse embryonic stem cells from permissive and nonpermissive strains. Nat. Protoc. 2014, 9, 559–574. [Google Scholar] [CrossRef]
- Varzideh, F.; Mone, P.; Santulli, G. Bioengineering Strategies to Create 3D Cardiac Constructs from Human Induced Pluripotent Stem Cells. Bioengineering 2022, 9, 168. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef]
- Yu, J.; Vodyanik, M.A.; Smuga-Otto, K.; Antosiewicz-Bourget, J.; Frane, J.L.; Tian, S.; Nie, J.; Jonsdottir, G.A.; Ruotti, V.; Stewart, R.; et al. Induced Pluripotent Stem Cell Lines Derived from Human Somatic Cells. Science 2007, 318, 1917–1920. [Google Scholar] [CrossRef]
- Park, I.-H.; Zhao, R.; West, J.A.; Yabuuchi, A.; Huo, H.; Ince, T.A.; Lerou, P.H.; Lensch, M.W.; Daley, G.Q. Reprogramming of human somatic cells to pluripotency with defined factors. Nature 2008, 451, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.; Plath, K. The roles of the reprogramming factors Oct4, Sox2 and Klf4 in resetting the somatic cell epigenome during induced pluripotent stem cell generation. Genome Biol. 2012, 13, 251. [Google Scholar] [CrossRef]
- Zhu, S.; Li, W.; Zhou, H.; Wei, W.; Ambasudhan, R.; Lin, T.; Kim, J.; Zhang, K.; Ding, S. Reprogramming of Human Primary Somatic Cells by OCT4 and Chemical Compounds. Cell Stem Cell 2010, 7, 651–655. [Google Scholar] [CrossRef] [PubMed]
- Plath, K.; Lowry, W.E. Progress in understanding reprogramming to the induced pluripotent state. Nat. Rev. Genet. 2011, 12, 253–265. [Google Scholar] [CrossRef]
- Dalton, S. Signaling networks in human pluripotent stem cells. Curr. Opin. Cell Biol. 2013, 25, 241–246. [Google Scholar] [CrossRef]
- Chen, X.; Xu, H.; Yuan, P.; Fang, F.; Huss, M.; Vega, V.B.; Wong, E.; Orlov, Y.L.; Zhang, W.; Jiang, J.; et al. Integration of External Signaling Pathways with the Core Transcriptional Network in Embryonic Stem Cells. Cell 2008, 133, 1106–1117. [Google Scholar] [CrossRef]
- Hall, J.; Guo, G.; Wray, J.; Eyres, I.; Nichols, J.; Grotewold, L.; Morfopoulou, S.; Humphreys, P.; Mansfield, W.; Walker, R.; et al. Oct4 and LIF/Stat3 Additively Induce Krüppel Factors to Sustain Embryonic Stem Cell Self-Renewal. Cell Stem Cell 2009, 5, 597–609. [Google Scholar] [CrossRef]
- Ng, H.-H.; Surani, A. The transcriptional and signalling networks of pluripotency. Nature 2011, 13, 490–496. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Lei, A.; Tian, L.; Wang, X.; Correia, C.; Weiskittel, T.; Li, H.; Trounson, A.; Fu, Q.; Yao, K.; et al. Strategies for Genetically Engineering Hypoimmunogenic Universal Pluripotent Stem Cells. iScience 2020, 23, 101162. [Google Scholar] [CrossRef] [PubMed]
- Weidgang, C.E.; Seufferlein, T.; Kleger, A.; Mueller, M. Pluripotency Factors on Their Lineage Move. Stem Cells Int. 2016, 2016, 6838253. [Google Scholar] [CrossRef] [PubMed]
- Pauklin, S.; Vallier, L. The Cell-Cycle State of Stem Cells Determines Cell Fate Propensity. Cell 2013, 155, 135–147. [Google Scholar] [CrossRef]
- Avery, K.; Avery, S.; Shepherd, J.; Heath, P.R.; Moore, H. Sphingosine-1-Phosphate Mediates Transcriptional Regulation of Key Targets Associated with Survival, Proliferation, and Pluripotency in Human Embryonic Stem Cells. Stem Cells Dev. 2008, 17, 1195–1206. [Google Scholar] [CrossRef]
- Ramírez, M.A.; Pericuesta, E.; Fernandez-Gonzalez, R.; Moreira, P.; Pintado, B.; Gutierrez-Adan, A. Transcriptional and post-transcriptional regulation of retrotransposons IAP and MuERV-L affect pluripotency of mice ES cells. Reprod. Biol. Endocrinol. 2006, 4, 55. [Google Scholar] [CrossRef]
- Zhang, J.; Tam, W.-L.; Tong, G.Q.; Wu, Q.; Chan, H.-Y.; Soh, B.-S.; Lou, Y.; Yang, J.; Ma, Y.; Chai, L.; et al. Sall4 modulates embryonic stem cell pluripotency and early embryonic development by the transcriptional regulation of Pou5f1. Nature 2006, 8, 1114–1123. [Google Scholar] [CrossRef]
- Shi, G.; Jin, Y. Role of Oct4 in maintaining and regaining stem cell pluripotency. Stem Cell Res. Ther. 2010, 1, 39. [Google Scholar] [CrossRef]
- Joshi, R.; Buchanan, J.C.; Paruchuri, S.; Morris, N.; Tavana, H. Molecular Analysis of Stromal Cells-Induced Neural Differentiation of Mouse Embryonic Stem Cells. PLoS ONE 2016, 11, e0166316. [Google Scholar] [CrossRef]
- George, M.N.; Leavens, K.F.; Gadue, P. Genome Editing Human Pluripotent Stem Cells to Model β-Cell Disease and Unmask Novel Genetic Modifiers. Front. Endocrinol. 2021, 12, 682625. [Google Scholar] [CrossRef]
- Yao, X.; Dani, V.; Dani, C. Human Pluripotent Stem Cells: A Relevant Model to Identify Pathways Governing Thermogenic Adipocyte Generation. Front. Endocrinol. 2019, 10, 932. [Google Scholar] [CrossRef]
- Sewell, W.; Lin, R.-Y. Generation of Thyroid Follicular Cells from Pluripotent Stem Cells: Potential for Regenerative Medicine. Front. Endocrinol. 2014, 5, 96. [Google Scholar] [CrossRef] [PubMed]
- Posabella, A.; Alber, A.B.; Undeutsch, H.J.; Droeser, R.A.; Hollenberg, A.N.; Ikonomou, L.; Kotton, D.N. Derivation of Thyroid Follicular Cells from Pluripotent Stem Cells: Insights from Development and Implications for Regenerative Medicine. Front. Endocrinol. 2021, 12, 666565. [Google Scholar] [CrossRef] [PubMed]
- Hackett, J.A.; Surani, M.A. Regulatory Principles of Pluripotency: From the Ground State Up. Cell Stem Cell 2014, 15, 416–430. [Google Scholar] [CrossRef] [PubMed]
- Geng, T.; Zhang, D.; Jiang, W. Epigenetic Regulation of Transition Among Different Pluripotent States: Concise Review. Stem Cells 2019, 37, 1372–1380. [Google Scholar] [CrossRef]
- Udomlumleart, T.; Hu, S.; Garg, S. Lineages of embryonic stem cells show non-Markovian state transitions. iScience 2021, 24, 102879. [Google Scholar] [CrossRef]
- Wang, R.; Guo, Y.-L. Transient inhibition of cell proliferation does not compromise self-renewal of mouse embryonic stem cells. Exp. Cell Res. 2012, 318, 2094–2104. [Google Scholar] [CrossRef]
- Devika, A.S.; Montebaur, A.; Saravanan, S.; Bhushan, R.; Koch, F.; Sudheer, S. Human ES Cell Culture Conditions Fail to Preserve the Mouse Epiblast State. Stem Cells Int. 2021, 2021, 8818356. [Google Scholar] [CrossRef]
- Tremble, K.C.; Stirparo, G.G.; Bates, L.E.; Maskalenka, K.; Stuart, H.T.; Jones, K.; Andersson-Rolf, A.; Radzisheuskaya, A.; Koo, B.-K.; Bertone, P.; et al. Sox2 modulation increases naïve pluripotency plasticity. iScience 2021, 24, 102153. [Google Scholar] [CrossRef]
- Liang, W.; Han, P.; Kim, E.H.; Mak, J.; Zhang, R.; Torrente, A.G.; Goldhaber, J.I.; Marbán, E.; Cho, H.C. Canonical Wnt signaling promotes pacemaker cell specification of cardiac mesodermal cells derived from mouse and human embryonic stem cells. Stem Cells 2020, 38, 352–368. [Google Scholar] [CrossRef]
- Pecori, F.; Kondo, N.; Ogura, C.; Miura, T.; Kume, M.; Minamijima, Y.; Yamamoto, K.; Nishihara, S. Site-specific O-GlcNAcylation of Psme3 maintains mouse stem cell pluripotency by impairing P-body homeostasis. Cell Rep. 2021, 36, 109361. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.-Y.; Huang, W.-H.; Chen, H.-C.; Meir, Y.-J.J. Capturing Pluripotency and Beyond. Cells 2021, 10, 3558. [Google Scholar] [CrossRef]
- Pollini, D.; Loffredo, R.; Maniscalco, F.; Cardano, M.; Micaelli, M.; Bonomo, I.; Licata, N.V.; Peroni, D.; Tomaszewska, W.; Rossi, A.; et al. Multilayer and MATR3-dependent regulation of mRNAs maintains pluripotency in human induced pluripotent stem cells. iScience 2021, 24, 102197. [Google Scholar] [CrossRef]
- Beyer, T.A.; Narimatsu, M.; Weiss, A.; David, L.; Wrana, J.L. The TGFβ superfamily in stem cell biology and early mammalian embryonic development. Biochim. Biophys. Acta 2013, 1830, 2268–2279. [Google Scholar] [CrossRef]
- Sakaki-Yumoto, M.; Katsuno, Y.; Derynck, R. TGF-β family signaling in stem cells. Biochim. et Biophys. Acta 2013, 1830, 2280–2296. [Google Scholar] [CrossRef]
- Watabe, T.; Miyazono, K. Roles of TGF-β family signaling in stem cell renewal and differentiation. Cell Res. 2009, 19, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Ying, Q.-L.; Nichols, J.; Chambers, I.; Smith, A. BMP Induction of Id Proteins Suppresses Differentiation and Sustains Embryonic Stem Cell Self-Renewal in Collaboration with STAT3. Cell 2003, 115, 281–292. [Google Scholar] [CrossRef]
- Li, Z.; Fei, T.; Zhang, J.; Zhu, G.; Wang, L.; Lu, D.; Chi, X.; Teng, Y.; Hou, N.; Yang, X.; et al. BMP4 Signaling Acts via Dual-Specificity Phosphatase 9 to Control ERK Activity in Mouse Embryonic Stem Cells. Cell Stem Cell 2012, 10, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, M.; Koinuma, D.; Mizutani, A.; Kawasaki, N.; Holmborn, K.; Sundqvist, A.; Tsutsumi, S.; Watabe, T.; Aburatani, H.; Heldin, C.-H.; et al. BMP Sustains Embryonic Stem Cell Self-Renewal through Distinct Functions of Different Krüppel-like Factors. Stem Cell Rep. 2016, 6, 64–73. [Google Scholar] [CrossRef]
- Richter, A.; Valdimarsdottir, L.; Hrafnkelsdottir, H.E.; Runarsson, J.F.; Omarsdottir, A.R.; Oostwaard, D.W.-V.; Mummery, C.; Valdimarsdottir, G. BMP4 Promotes EMT and Mesodermal Commitment in Human Embryonic Stem Cells via SLUG and MSX2. Stem Cells 2014, 32, 636–648. [Google Scholar] [CrossRef]
- Chen, L.; Yang, M.; Dawes, J.; Khillan, J.S. Suppression of ES cell differentiation by retinol (vitamin A) via the overexpression of Nanog. Differentiation 2007, 75, 682–693. [Google Scholar] [CrossRef] [PubMed]
- Chambers, I.; Colby, D.; Robertson, M.; Nichols, J.; Lee, S.; Tweedie, S.; Smith, A. Functional Expression Cloning of Nanog, a Pluripotency Sustaining Factor in Embryonic Stem Cells. Cell 2003, 113, 643–655. [Google Scholar] [CrossRef] [PubMed]
- Chambers, I. The Molecular Basis of Pluripotency in Mouse Embryonic Stem Cells. Cloning Stem Cells 2004, 6, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Wang, H.; Pan, G.; Geng, Y.; Guo, Y.; Pei, D. Regulation of the Pluripotency Marker Rex-1 by Nanog and Sox2. J. Biol. Chem. 2006, 281, 23319–23325. [Google Scholar] [CrossRef]
- Festuccia, N.; Osorno, R.; Halbritter, F.; Karwacki-Neisius, V.; Navarro, P.; Colby, D.; Wong, F.; Yates, A.; Tomlinson, S.R.; Chambers, I. Esrrb Is a Direct Nanog Target Gene that Can Substitute for Nanog Function in Pluripotent Cells. Cell Stem Cell 2012, 11, 477–490. [Google Scholar] [CrossRef]
- He, R.; Xhabija, B.; Al-Qanber, B.; Kidder, B.L. OCT4 supports extended LIF-independent self-renewal and maintenance of transcriptional and epigenetic networks in embryonic stem cells. Sci. Rep. 2017, 7, 16360. [Google Scholar] [CrossRef]
- Närvä, E.; Taskinen, M.E.; Lilla, S.; Isomursu, A.; Pietilä, M.; Weltner, J.; Isola, J.; Sihto, H.; Joensuu, H.; Zanivan, S.; et al. MASTL is enriched in cancerous and pluripotent stem cells and influences OCT1/OCT4 levels. iScience 2022, 25, 104459. [Google Scholar] [CrossRef] [PubMed]
- Abdelalim, E.M.; Tooyama, I. NPR-A regulates self-renewal and pluripotency of embryonic stem cells. Cell Death Dis. 2011, 2, e127. [Google Scholar] [CrossRef]
- Theret, M.; Gsaier, L.; Schaffer, B.; Juban, G.; Ben Larbi, S.; Weiss-Gayet, M.; Bultot, L.; Collodet, C.; Foretz, M.; Desplanches, D.; et al. AMPK α1- LDH pathway regulates muscle stem cell self-renewal by controlling metabolic homeostasis. EMBO J. 2017, 36, 1946–1962. [Google Scholar] [CrossRef]
- Pei, S.; Minhajuddin, M.; Adane, B.; Khan, N.; Stevens, B.M.; Mack, S.C.; Lai, S.; Rich, J.N.; Inguva, A.; Shannon, K.M.; et al. AMPK/FIS1-Mediated Mitophagy Is Required for Self-Renewal of Human AML Stem Cells. Cell Stem Cell 2018, 23, 86–100.e6. [Google Scholar] [CrossRef]
- Alba, G.; Martínez, R.; Postigo-Corrales, F.; López, S.; Santa-María, C.; Jiménez, G.A.; Cahuana, G.M.; Soria, B.; Bedoya, F.J.; Tejedo, J.R. AICAR Stimulates the Pluripotency Transcriptional Complex in Embryonic Stem Cells Mediated by PI3K, GSK3β, and β-Catenin. ACS Omega 2020, 5, 20270–20282. [Google Scholar] [CrossRef]
- Jankauskas, S.S.; Mone, P.; Avvisato, R.; Varzideh, F.; De Gennaro, S.; Salemme, L.; Macina, G.; Kansakar, U.; Cioppa, A.; Frullone, S.; et al. miR-181c targets Parkin and SMAD7 in human cardiac fibroblasts: Validation of differential microRNA expression in patients with diabetes and heart failure with preserved ejection fraction. Mech. Ageing Dev. 2023; in press. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Gu, S.; Li, W.; Sun, C.; Chen, F.; Xiao, M.; Wang, L.; Xu, D.; Li, Y.; Ding, C.; et al. Smad7 enables STAT3 activation and promotes pluripotency independent of TGF-β signaling. Proc. Natl. Acad. Sci. USA 2017, 114, 10113–10118. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.-C.; Liao, P.-N.; Ho, A.-S.; Lim, K.-H.; Chang, J.; Su, Y.-W.; Chen, C.G.-S.; Chiang, Y.-W.; Yang, B.-L.; Lin, H.-C.; et al. STAT3 exacerbates survival of cancer stem-like tumorspheres in EGFR-positive colorectal cancers: RNAseq analysis and therapeutic screening. J. Biomed. Sci. 2018, 25, 60. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-J.; Min, T.-R.; Chi, G.-Y.; Choi, Y.-H.; Park, S.-H. Induction of apoptosis by morusin in human non-small cell lung cancer cells by suppression of EGFR/STAT3 activation. Biochem. Biophys. Res. Commun. 2018, 505, 194–200. [Google Scholar] [CrossRef]
- Prenzel, N.; Fischer, O.M.; Streit, S.; Hart, S.; Ullrich, A. The epidermal growth factor receptor family as a central element for cellular signal transduction and diversification. Endocr. Relat. Cancer 2001, 8, 11–31. [Google Scholar] [CrossRef]
- Yu, M.; Wei, Y.; Xu, K.; Liu, S.; Ma, L.; Pei, Y.; Hu, Y.; Liu, Z.; Zhang, X.; Wang, B.; et al. EGFR deficiency leads to impaired self-renewal and pluripotency of mouse embryonic stem cells. PeerJ 2019, 7, e6314. [Google Scholar] [CrossRef]
- Xia, R.M.; Yao, D.B.; Cai, X.M.; Xu, X.Q. LHPP-Mediated Histidine Dephosphorylation Suppresses the Self-Renewal of Mouse Embryonic Stem Cells. Front. Cell Dev. Biol. 2021, 9, 638815. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhang, Y.; Wang, X.; Wang, X.; Ye, S.-D. Inhibition of protein kinase D by CID755673 promotes maintenance of the pluripotency of embryonic stem cells. Development 2020, 147, dev185264. [Google Scholar] [CrossRef]
- Park, S.; Kwon, W.; Kim, H.-Y.; Ji, Y.R.; Kim, D.; Kim, W.; Han, J.E.; Cho, G.-J.; Yun, S.; Kim, M.O.; et al. Knockdown of Maged1 inhibits cell cycle progression and causes cell death in mouse embryonic stem cells. Differentiation 2022, 125, 18–26. [Google Scholar] [CrossRef]
- Ju, H.; Lee, S.; Lee, J.; Ghil, S. Necdin modulates osteogenic cell differentiation by regulating Dlx5 and MAGE-D1. Biochem. Biophys. Res. Commun. 2017, 489, 109–115. [Google Scholar] [CrossRef]
- Chen, X.; Wu, R.; Feng, S.; Gu, B.; Dai, L.; Zhang, M.; Zhao, X. Single cell derived murine embryonic stem cell clones stably express Rex1-specific green fluorescent protein and their differentiation study. Biochem. Biophys. Res. Commun. 2007, 362, 467–473. [Google Scholar] [CrossRef]
- Li, T.; Kon, N.; Jiang, L.; Tan, M.; Ludwig, T.; Zhao, Y.; Baer, R.; Gu, W. Tumor Suppression in the Absence of p53-Mediated Cell-Cycle Arrest, Apoptosis, and Senescence. Cell 2012, 149, 1269–1283. [Google Scholar] [CrossRef]
- Aubrey, B.J.; Kelly, G.L.; Janic, A.; Herold, M.J.; Strasser, A. How does p53 induce apoptosis and how does this relate to p53-mediated tumour suppression? Cell Death Differ. 2018, 25, 104–113. [Google Scholar] [CrossRef]
- Fu, X.; Wu, S.; Li, B.; Xu, Y.; Liu, J. Functions of p53 in pluripotent stem cells. Protein Cell 2020, 11, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.; Chao, C.; Saito, S.; Mazur, S.J.; Murphy, M.; Appella, E.; Xu, Y. p53 induces differentiation of mouse embryonic stem cells by suppressing Nanog expression. Nat. Cell Biol. 2005, 7, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; He, Y.; Dubois, W.; Wu, X.; Shi, J.; Huang, J. Distinct Regulatory Mechanisms and Functions for p53-Activated and p53-Repressed DNA Damage Response Genes in Embryonic Stem Cells. Mol. Cell 2012, 46, 30–42. [Google Scholar] [CrossRef]
- Yang, A.; Shi, G.; Zhou, C.; Lu, R.; Li, H.; Sun, L.; Jin, Y. Nucleolin Maintains Embryonic Stem Cell Self-renewal by Suppression of p53 Protein-dependent Pathway. J. Biol. Chem. 2011, 286, 43370–43382. [Google Scholar] [CrossRef]
- Xiao, L.; Yuan, X.; Sharkis, S.J. Activin A Maintains Self-Renewal and Regulates Fibroblast Growth Factor, Wnt, and Bone Morphogenic Protein Pathways in Human Embryonic Stem Cells. Stem Cells 2006, 24, 1476–1486. [Google Scholar] [CrossRef] [PubMed]
- Beattie, G.M.; Lopez, A.D.; Bucay, N.; Hinton, A.; Firpo, M.T.; King, C.C.; Hayek, A. Activin A Maintains Pluripotency of Human Embryonic Stem Cells in the Absence of Feeder Layers. Stem Cells 2005, 23, 489–495. [Google Scholar] [CrossRef]
- Massagué, J.; Seoane, J.; Wotton, D. Smad transcription factors. Genes Dev. 2005, 19, 2783–2810. [Google Scholar] [CrossRef]
- Vallier, L.; Mendjan, S.; Brown, S.; Chng, Z.; Teo, A.; Smithers, L.E.; Trotter, M.W.B.; Cho, C.H.-H.; Martinez, A.; Rugg-Gunn, P.; et al. Activin/Nodal signalling maintains pluripotency by controlling Nanog expression. Development 2009, 136, 1339–1349. [Google Scholar] [CrossRef] [PubMed]
- Bertero, A.; Madrigal, P.; Galli, A.; Hubner, N.C.; Moreno, I.; Burks, D.; Brown, S.; Pedersen, R.A.; Gaffney, D.; Mendjan, S.; et al. Activin/Nodal signaling and NANOG orchestrate human embryonic stem cell fate decisions by controlling the H3K4me3 chromatin mark. Genes Dev. 2015, 29, 702–717. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.-H.; Sampsell-Barron, T.L.; Gu, F.; Root, S.; Peck, R.M.; Pan, G.; Yu, J.; Antosiewicz-Bourget, J.; Tian, S.; Stewart, R.; et al. NANOG Is a Direct Target of TGFβ/Activin-Mediated SMAD Signaling in Human ESCs. Cell Stem Cell 2008, 3, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.; Teo, A.; Pauklin, S.; Hannan, N.; Cho, C.H.-H.; Lim, B.; Vardy, L.; Dunn, N.R.; Trotter, M.; Pedersen, R.; et al. Activin/Nodal Signaling Controls Divergent Transcriptional Networks in Human Embryonic Stem Cells and in Endoderm Progenitors. Stem Cells 2011, 29, 1176–1185. [Google Scholar] [CrossRef]
- Sakaki-Yumoto, M.; Liu, J.; Ramalho-Santos, M.; Yoshida, N.; Derynck, R. Smad2 Is Essential for Maintenance of the Human and Mouse Primed Pluripotent Stem Cell State. J. Biol. Chem. 2013, 288, 18546–18560. [Google Scholar] [CrossRef]
- Chng, Z.; Teo, A.; Pedersen, R.A.; Vallier, L. SIP1 Mediates Cell-Fate Decisions between Neuroectoderm and Mesendoderm in Human Pluripotent Stem Cells. Cell Stem Cell 2010, 6, 59–70. [Google Scholar] [CrossRef]
- Tsuneyoshi, N.; Tan, E.K.; Sadasivam, A.; Poobalan, Y.; Sumi, T.; Nakatsuji, N.; Suemori, H.; Dunn, N.R. The SMAD2/3 corepressor SNON maintains pluripotency through selective repression of mesendodermal genes in human ES cells. Genes Dev. 2012, 26, 2471–2476. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.V.; Dixon, G.; Verma, N.; Rosen, B.P.; Gordillo, M.; Luo, R.; Xu, C.; Wang, Q.; Soh, C.-L.; Yang, D.; et al. Genome-scale screens identify JNK–JUN signaling as a barrier for pluripotency exit and endoderm differentiation. Nat. Genet. 2019, 51, 999–1010. [Google Scholar] [CrossRef]
- Tsai, Z.-Y.; Singh, S.; Yu, S.-L.; Kao, L.-P.; Chen, B.-Z.; Ho, B.-C.; Yang, P.-C.; Li, S.S.-L. Identification of microRNAs regulated by activin A in human embryonic stem cells. J. Cell. Biochem. 2010, 109, 93–102. [Google Scholar] [CrossRef]
- Tomizawa, M.; Shinozaki, F.; Sugiyama, T.; Yamamoto, S.; Sueishi, M.; Yoshida, T. Activin A is essential for Feeder-free culture of human induced pluripotent stem cells. J. Cell. Biochem. 2013, 114, 584–588. [Google Scholar] [CrossRef]
- D'Amour, K.A.; Agulnick, A.D.; Eliazer, S.; Kelly, O.G.; Kroon, E.; Baetge, E.E. Efficient differentiation of human embryonic stem cells to definitive endoderm. Nat. Biotechnol. 2005, 23, 1534–1541. [Google Scholar] [CrossRef]
- Cheng, C.-L.; Yang, S.-C.; Lai, C.-Y.; Wang, C.-K.; Chang, C.-F.; Lin, C.-Y.; Chen, W.-J.; Lin, P.-Y.; Wu, H.-C.; Ma, N.; et al. CXCL14 Maintains hESC Self-Renewal through Binding to IGF-1R and Activation of the IGF-1R Pathway. Cells 2020, 9, 1706. [Google Scholar] [CrossRef]
- Timilsina, S.; Kirsch-Mangu, T.; Werth, S.; Shepard, B.; Ma, T.; Villa-Diaz, L.G. Enhanced self-renewal of human pluripotent stem cells by simulated microgravity. npj Microgravity 2022, 8, 22. [Google Scholar] [CrossRef]
- Levenstein, M.E.; Ludwig, T.E.; Xu, R.-H.; Llanas, R.A.; VanDenHeuvel-Kramer, K.; Manning, D.; Thomson, J.A. Basic Fibroblast Growth Factor Support of Human Embryonic Stem Cell Self-Renewal. Stem Cells 2006, 24, 568–574. [Google Scholar] [CrossRef]
- Greber, B.; Lehrach, H.; Adjaye, J. Fibroblast Growth Factor 2 Modulates Transforming Growth Factor β Signaling in Mouse Embryonic Fibroblasts and Human ESCs (hESCs) to Support hESC Self-Renewal. Stem Cells 2007, 25, 455–464. [Google Scholar] [CrossRef]
- Li, J.; Wang, G.; Wang, C.; Zhao, Y.; Zhang, H.; Tan, Z.; Song, Z.; Ding, M.; Deng, H. MEK/ERK signaling contributes to the maintenance of human embryonic stem cell self-renewal. Differentiation 2007, 75, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, L.; Hughes, O.; Yung, S.; Hyslop, L.; Stewart, R.; Wappler, I.; Peters, H.; Walter, T.; Stojkovic, P.; Evans, J.; et al. The role of PI3K/AKT, MAPK/ERK and NFκβ signalling in the maintenance of human embryonic stem cell pluripotency and viability highlighted by transcriptional profiling and functional analysis. Hum. Mol. Genet. 2006, 15, 1894–1913. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.B.; Kim, J.S.; Kwon, H.-J.; Nam, K.H.; Youn, H.S.; Sok, D.-E.; Lee, Y.; Hu, F.; Wang, X.; Liang, G.; et al. Basic Fibroblast Growth Factor Activates ERK and Induces c-Fos in Human Embryonic Stem Cell Line MizhES1. Stem Cells Dev. 2005, 14, 395–401. [Google Scholar] [CrossRef]
- Paling, N.R.D.; Wheadon, H.; Bone, H.K.; Welham, M.J. Regulation of Embryonic Stem Cell Self-renewal by Phosphoinositide 3-Kinase-dependent Signaling. J. Biol. Chem. 2004, 279, 48063–48070. [Google Scholar] [CrossRef] [PubMed]
- Storm, M.P.; Bone, H.K.; Beck, C.G.; Bourillot, P.-Y.; Schreiber, V.; Damiano, T.; Nelson, A.; Savatier, P.; Welham, M.J. Regulation of Nanog Expression by Phosphoinositide 3-Kinase-dependent Signaling in Murine Embryonic Stem Cells. J. Biol. Chem. 2007, 282, 6265–6273. [Google Scholar] [CrossRef]
- Storm, M.P.; Kumpfmueller, B.; Thompson, B.; Kolde, R.; Vilo, J.; Hummel, O.; Schulz, H.; Welham, M.J. Characterization of the Phosphoinositide 3-Kinase-Dependent Transcriptome in Murine Embryonic Stem Cells: Identification of Novel Regulators of Pluripotency. Stem Cells 2009, 27, 764–775. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Ripoll, Y.; Bone, H.K.; Owen, T.; Guedes, A.M.V.; Abranches, E.; Kumpfmueller, B.; Spriggs, R.V.; Henrique, D.; Welham, M.J. Glycogen Synthase Kinase-3 Inhibition Enhances Translation of Pluripotency-Associated Transcription Factors to Contribute to Maintenance of Mouse Embryonic Stem Cell Self-Renewal. PLoS ONE 2013, 8, e60148. [Google Scholar] [CrossRef]
- Romorini, L.; Garate, X.; Neiman, G.; Luzzani, C.; Furmento, V.A.; Guberman, A.S.; Sevlever, G.E.; Scassa, M.E.; Miriuka, S.G. AKT/GSK3β signaling pathway is critically involved in human pluripotent stem cell survival. Sci. Rep. 2016, 6, 35660. [Google Scholar] [CrossRef] [PubMed]
- Galarreta, A.; Valledor, P.; Fernandez-Capetillo, O.; Lecona, E. Coordinating DNA Replication and Mitosis through Ubiquitin/SUMO and CDK1. Int. J. Mol. Sci. 2021, 22, 8796. [Google Scholar] [CrossRef]
- Gallo, D.; Young, J.T.F.; Fourtounis, J.; Martino, G.; Álvarez-Quilón, A.; Bernier, C.; Duffy, N.M.; Papp, R.; Roulston, A.; Stocco, R.; et al. CCNE1 amplification is synthetic lethal with PKMYT1 kinase inhibition. Nature 2022, 604, 749–756. [Google Scholar] [CrossRef]
- Yu, J.; Raia, P.; Ghent, C.M.; Raisch, T.; Sadian, Y.; Cavadini, S.; Sabale, P.M.; Barford, D.; Raunser, S.; Morgan, D.O.; et al. Structural basis of human separase regulation by securin and CDK1–cyclin B1. Nature 2021, 596, 138–142. [Google Scholar] [CrossRef] [PubMed]
- Ulu, A.; Oh, W.; Zuo, Y.; Frost, J.A. Cdk1 phosphorylation negatively regulates the activity of Net1 towards RhoA during mitosis. Cell. Signal. 2021, 80, 109926. [Google Scholar] [CrossRef]
- Wang, X.Q.; Lo, C.M.; Chen, L.; Ngan, E.; Xu, A.; Poon, R. CDK1-PDK1-PI3K/Akt signaling pathway regulates embryonic and induced pluripotency. Cell Death Differ. 2017, 24, 38–48. [Google Scholar] [CrossRef]
- Zhang, W.W.; Zhang, X.J.; Liu, H.X.; Chen, J.; Ren, Y.H.; Huang, D.G.; Zou, X.H.; Xiao, W. Cdk1 is required for the self-renewal of mouse embryonic stem cells. J. Cell. Biochem. 2011, 112, 942–948. [Google Scholar] [CrossRef]
- Neganova, I.; Tilgner, K.; Buskin, A.; Paraskevopoulou, I.; Atkinson, S.P.; Peberdy, D.; Passos, J.F.; Lako, M. CDK1 plays an important role in the maintenance of pluripotency and genomic stability in human pluripotent stem cells. Cell Death Dis. 2014, 5, e1508. [Google Scholar] [CrossRef]
- Munoz-Descalzo, S.; Rué, P.; Faunes, F.; Hayward, P.; Jakt, L.M.; Balayo, T.; Garcia-Ojalvo, J.; Arias, A.M. A competitive protein interaction network buffers Oct4-mediated differentiation to promote pluripotency in embryonic stem cells. Mol. Syst. Biol. 2013, 9, 694. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Rao, S.; Chu, J.; Shen, X.; Levasseur, D.N.; Theunissen, T.W.; Orkin, S.H. A protein interaction network for pluripotency of embryonic stem cells. Nature 2006, 444, 364–368. [Google Scholar] [CrossRef]
- Li, L.; Wang, J.; Hou, J.; Wu, Z.; Zhuang, Y.; Lu, M.; Zhang, Y.; Zhou, X.; Li, Z.; Xiao, W.; et al. Cdk1 interplays with Oct4 to repress differentiation of embryonic stem cells into trophectoderm. FEBS Lett. 2012, 586, 4100–4107. [Google Scholar] [CrossRef]
- Palmieri, S.L.; Peter, W.; Hess, H.; Schöler, H. Oct-4 Transcription Factor Is Differentially Expressed in the Mouse Embryo during Establishment of the First Two Extraembryonic Cell Lineages Involved in Implantation. Dev. Biol. 1994, 166, 259–267. [Google Scholar] [CrossRef]
- Zhou, C.; Yang, X.; Sun, Y.; Yu, H.; Zhang, Y.; Jin, Y. Comprehensive profiling reveals mechanisms of SOX2-mediated cell fate specification in human ESCs and NPCs. Cell Res. 2016, 26, 171–189. [Google Scholar] [CrossRef]
- Davidson, K.C.; Adams, A.M.; Goodson, J.M.; McDonald, C.E.; Potter, J.C.; Berndt, J.D.; Biechele, T.L.; Taylor, R.J.; Moon, R.T. Wnt/β-catenin signaling promotes differentiation, not self-renewal, of human embryonic stem cells and is repressed by Oct4. Proc. Natl. Acad. Sci. USA 2012, 109, 4485–4490. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, X.; Xu, X.; Kyba, M.; Cooney, A.J. Germ Cell Nuclear Factor (GCNF) Represses Oct4 Expression and Globally Modulates Gene Expression in Human Embryonic Stem (hES) Cells. J. Biol. Chem. 2016, 291, 8644–8652. [Google Scholar] [CrossRef] [PubMed]
- Ying, Q.-L.; Wray, J.; Nichols, J.; Batlle-Morera, L.; Doble, B.; Woodgett, J.; Cohen, P.; Smith, A. The ground state of embryonic stem cell self-renewal. Nature 2008, 453, 519–523. [Google Scholar] [CrossRef]
- Martisova, A.; Holcakova, J.; Izadi, N.; Sebuyoya, R.; Hrstka, R.; Bartosik, M. DNA Methylation in Solid Tumors: Functions and Methods of Detection. Int. J. Mol. Sci. 2021, 22, 4247. [Google Scholar] [CrossRef]
- Wu, W.; Hill, S.E.; Nathan, W.J.; Paiano, J.; Callen, E.; Wang, D.; Shinoda, K.; van Wietmarschen, N.; Colón-Mercado, J.M.; Zong, D.; et al. Neuronal enhancers are hotspots for DNA single-strand break repair. Nature 2021, 593, 440–444. [Google Scholar] [CrossRef] [PubMed]
- Gambardella, J.; Jankauskas, S.; Kansakar, U.; Varzideh, F.; Avvisato, R.; Prevete, N.; Sidoli, S.; Mone, P.; Wang, X.; Lombardi, A.; et al. Ketone bodies rescue mitochondrial dysfunction via epigenetic remodeling. JACC Basic Transl. Sci. 2023; in press. [Google Scholar]
- Mittelstaedt, N.N.; Becker, A.L.; de Freitas, D.N.; Zanin, R.F.; Stein, R.T.; de Souza, A.P.D. DNA Methylation and Immune Memory Response. Cells 2021, 10, 2943. [Google Scholar] [CrossRef]
- Światowy, W.J.; Drzewiecka, H.; Kliber, M.; Sąsiadek, M.; Karpiński, P.; Pławski, A.; Jagodziński, P.P. Physical Activity and DNA Methylation in Humans. Int. J. Mol. Sci. 2021, 22, 12989. [Google Scholar] [CrossRef]
- Mattei, A.L.; Bailly, N.; Meissner, A. DNA methylation: A historical perspective. Trends Genet. 2022, 38, 676–707. [Google Scholar] [CrossRef] [PubMed]
- Lappalainen, T.; Greally, J.M. Associating cellular epigenetic models with human phenotypes. Nat. Rev. Genet. 2017, 18, 441–451. [Google Scholar] [CrossRef]
- Shilatifard, A. The COMPASS Family of Histone H3K4 Methylases: Mechanisms of Regulation in Development and Disease Pathogenesis. Annu. Rev. Biochem. 2012, 81, 65–95. [Google Scholar] [CrossRef] [PubMed]
- Lanzuolo, C.; Orlando, V. Memories from the Polycomb Group Proteins. Annu. Rev. Genet. 2012, 46, 561–589. [Google Scholar] [CrossRef]
- Lauberth, S.M.; Nakayama, T.; Wu, X.; Ferris, A.L.; Tang, Z.; Hughes, S.H.; Roeder, R.G. H3K4me3 Interactions with TAF3 Regulate Preinitiation Complex Assembly and Selective Gene Activation. Cell 2013, 152, 1021–1036. [Google Scholar] [CrossRef]
- Simon, J.A.; Kingston, R.E. Occupying Chromatin: Polycomb Mechanisms for Getting to Genomic Targets, Stopping Transcriptional Traffic, and Staying Put. Mol. Cell 2013, 49, 808–824. [Google Scholar] [CrossRef]
- Bernstein, B.E.; Mikkelsen, T.S.; Xie, X.; Kamal, M.; Huebert, D.J.; Cuff, J.; Fry, B.; Meissner, A.; Wernig, M.; Plath, K.; et al. A Bivalent Chromatin Structure Marks Key Developmental Genes in Embryonic Stem Cells. Cell 2006, 125, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Pan, G.; Tian, S.; Nie, J.; Yang, C.; Ruotti, V.; Wei, H.; Jonsdottir, G.A.; Stewart, R.; Thomson, J.A. Whole-Genome Analysis of Histone H3 Lysine 4 and Lysine 27 Methylation in Human Embryonic Stem Cells. Cell Stem Cell 2007, 1, 299–312. [Google Scholar] [CrossRef]
- Zhao, X.D.; Han, X.; Chew, J.L.; Liu, J.; Chiu, K.P.; Choo, A.; Orlov, Y.L.; Sung, W.-K.; Shahab, A.; Kuznetsov, V.A.; et al. Whole-Genome Mapping of Histone H3 Lys4 and 27 Trimethylations Reveals Distinct Genomic Compartments in Human Embryonic Stem Cells. Cell Stem Cell 2007, 1, 286–298. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Fan, C.; Zhang, J.L.; Zhang, S.Q. Expression specificity and compensation effect of Ash2l-1/Ash2l-2 in mouse embryonic stem cells. Yi Chuan 2018, 40, 237–249. [Google Scholar] [CrossRef]
- Heurtier, V.; Owens, N.; Gonzalez, I.; Mueller, F.; Proux, C.; Mornico, D.; Clerc, P.; Dubois, A.; Navarro, P. The molecular logic of Nanog-induced self-renewal in mouse embryonic stem cells. Nat. Commun. 2019, 10, 1109. [Google Scholar] [CrossRef] [PubMed]
- Olariu, V.; Lövkvist, C.; Sneppen, K. Nanog, Oct4 and Tet1 interplay in establishing pluripotency. Sci. Rep. 2016, 6, 25438. [Google Scholar] [CrossRef]
- Sohni, A.; Bartoccetti, M.; Khoueiry, R.; Spans, L.; Velde, J.V.; De Troyer, L.; Pulakanti, K.; Claessens, F.; Rao, S.; Koh, K.P. Dynamic Switching of Active Promoter and Enhancer Domains Regulates Tet1 and Tet2 Expression during Cell State Transitions between Pluripotency and Differentiation. Mol. Cell. Biol. 2015, 35, 1026–1042. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Chen, J.; Li, K.; Wu, T.; Huang, B.; Liu, W.; Kou, X.; Zhang, Y.; Huang, H.; Jiang, Y.; et al. Replacement of Oct4 by Tet1 during iPSC Induction Reveals an Important Role of DNA Methylation and Hydroxymethylation in Reprogramming. Cell Stem Cell 2013, 12, 453–469. [Google Scholar] [CrossRef]
- Lee, J.; Wu, Y.; Harada, B.T.; Li, Y.; Zhao, J.; He, C.; Ma, Y.; Wu, X. N6 -methyladenosine modification of lncRNA Pvt1 governs epidermal stemness. EMBO J. 2021, 40, e106276. [Google Scholar] [CrossRef]
- Sepich-Poore, C.; Zheng, Z.; Schmitt, E.; Wen, K.; Zhang, Z.S.; Cui, X.-L.; Dai, Q.; Zhu, A.C.; Zhang, L.; Castillo, A.S.; et al. The METTL5-TRMT112 N6-methyladenosine methyltransferase complex regulates mRNA translation via 18S rRNA methylation. J. Biol. Chem. 2022, 298, 101590. [Google Scholar] [CrossRef]
- Wen, J.; Lv, R.; Ma, H.; Shen, H.; He, C.; Wang, J.; Jiao, F.; Liu, H.; Yang, P.; Tan, L.; et al. Zc3h13 Regulates Nuclear RNA m6A Methylation and Mouse Embryonic Stem Cell Self-Renewal. Mol. Cell 2018, 69, 1028–1038.e6. [Google Scholar] [CrossRef] [PubMed]
- Tsumura, A.; Hayakawa, T.; Kumaki, Y.; Takebayashi, S.-I.; Sakaue, M.; Matsuoka, C.; Shimotohno, K.; Ishikawa, F.; Li, E.; Ueda, H.R.; et al. Maintenance of self-renewal ability of mouse embryonic stem cells in the absence of DNA methyltransferases Dnmt1, Dnmt3a and Dnmt3b. Genes Cells 2006, 11, 805–814. [Google Scholar] [CrossRef]
- Dahariya, S.; Raghuwanshi, S.; Thamodaran, V.; Velayudhan, S.R.; Gutti, R.K. Role of Long non-coding RNAs in Human Induced Pluripotent Stem Cells derived Megakaryocytes: A p53, HOTAIRM1 and miR-125b interaction study. J. Pharmacol. Exp. Ther. 2022. [Google Scholar] [CrossRef]
- Rosa, A.; Ballarino, M. Long Noncoding RNA Regulation of Pluripotency. Stem Cells Int. 2016, 2016, 1797692. [Google Scholar] [CrossRef] [PubMed]
- Fico, A.; Fiorenzano, A.; Pascale, E.; Patriarca, E.J.; Minchiotti, G. Long non-coding RNA in stem cell pluripotency and lineage commitment: Functions and evolutionary conservation. Cell. Mol. Life Sci. 2019, 76, 1459–1471. [Google Scholar] [CrossRef]
- Zhong, L.; Mu, H.; Wen, B.; Zhang, W.; Wei, Q.; Gao, G.; Han, J.; Cao, S. Long non-coding RNAs involved in the regulatory network during porcine pre-implantation embryonic development and iPSC induction. Sci. Rep. 2018, 8, 6649. [Google Scholar] [CrossRef]
- Hunkler, H.J.; Groß, S.; Thum, T.; Bär, C. Non-coding RNAs: Key regulators of reprogramming, pluripotency, and cardiac cell specification with therapeutic perspective for heart regeneration. Cardiovasc. Res. 2022, 118, 3071–3084. [Google Scholar] [CrossRef]
- Chen, J.; Wang, Y.; Wang, C.; Hu, J.-F.; Li, W. LncRNA Functions as a New Emerging Epigenetic Factor in Determining the Fate of Stem Cells. Front. Genet. 2020, 11, 277. [Google Scholar] [CrossRef]
- Zhao, C.; Xie, W.; Zhu, H.; Zhao, M.; Liu, W.; Wu, Z.; Wang, L.; Bin Zhu, B.; Li, S.; Zhou, Y.; et al. LncRNAs and their RBPs: How to influence the fate of stem cells? Stem Cell Res. Ther. 2022, 13, 175. [Google Scholar] [CrossRef]
- Mondal, T.; Kanduri, C. Noncoding RNA scaffolds in pluripotency. Circ. Res. 2012, 110, 1162–1165. [Google Scholar] [CrossRef]
- Guttman, M.; Donaghey, J.; Carey, B.W.; Garber, M.; Grenier, J.K.; Munson, G.; Young, G.; Lucas, A.B.; Ach, R.; Bruhn, L.; et al. lincRNAs act in the circuitry controlling pluripotency and differentiation. Nature 2011, 477, 295–300. [Google Scholar] [CrossRef]
- Ng, S.-Y.; Johnson, R.; Stanton, L.W. Human long non-coding RNAs promote pluripotency and neuronal differentiation by association with chromatin modifiers and transcription factors. EMBO J. 2012, 31, 522–533. [Google Scholar] [CrossRef]
- Shi, X.; Wu, Y.; Ai, Z.; Liu, X.; Yang, L.; Du, J.; Shao, J.; Guo, Z.; Zhang, Y. AICAR Sustains J1 Mouse Embryonic Stem Cell Self-Renewal and Pluripotency by Regulating Transcription Factor and Epigenetic Modulator Expression. Cell. Physiol. Biochem. 2013, 32, 459–475. [Google Scholar] [CrossRef]
- Jia, J.; Zheng, X.; Hu, G.; Cui, K.; Zhang, J.; Zhang, A.; Jiang, H.; Lu, B.; Yates, J., 3rd; Liu, C.; et al. Regulation of Pluripotency and Self- Renewal of ESCs through Epigenetic- Threshold Modulation and mRNA Pruning. Cell 2012, 151, 576–589. [Google Scholar] [CrossRef] [PubMed]
- Guo, G.; Yang, J.; Nichols, J.; Hall, J.S.; Eyres, I.; Mansfield, W.; Smith, A. Klf4 reverts developmentally programmed restriction of ground state pluripotency. Development 2009, 136, 1063–1069. [Google Scholar] [CrossRef] [PubMed]
- Hanna, J.; Markoulaki, S.; Mitalipova, M.; Cheng, A.W.; Cassady, J.P.; Staerk, J.; Carey, B.W.; Lengner, C.J.; Foreman, R.; Love, J.; et al. Metastable Pluripotent States in NOD-Mouse-Derived ESCs. Cell Stem Cell 2009, 4, 513–524. [Google Scholar] [CrossRef]
- Okamura, D.; Chikushi, M.; Chigi, Y.; Shiogai, N.; Jafar, S.; Wu, J. Stepwise conversion methods between ground states pluripotency from naïve to primed. Biochem. Biophys. Res. Commun. 2021, 574, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Tiana, M.; Lopez-Jimenez, E.; de Aja, J.S.; Barral, A.; Victorino, J.; Badia-Careaga, C.; Rollan, I.; Rouco, R.; Santos, E.; Sanchez-Iranzo, H.; et al. Pluripotency factors regulate the onset of Hox cluster activation in the early embryo. Sci. Adv. 2022, 8, eabo3583. [Google Scholar] [CrossRef]
- Minchiotti, G.; D’aniello, C.; Fico, A.; De Cesare, D.; Patriarca, E.J. Capturing Transitional Pluripotency through Proline Metabolism. Cells 2022, 11, 2125. [Google Scholar] [CrossRef]
- Romayor, I.; Herrera, L.; Burón, M.; Martin-Inaraja, M.; Prieto, L.; Etxaniz, J.; Inglés-Ferrándiz, M.; Pineda, J.R.; Eguizabal, C. A Comparative Study of Cell Culture Conditions during Conversion from Primed to Naive Human Pluripotent Stem Cells. Biomedicines 2022, 10, 1358. [Google Scholar] [CrossRef]
- Yousefi, M.; Marashi, S.-A.; Sharifi-Zarchi, A.; Taleahmad, S. The metabolic network model of primed/naive human embryonic stem cells underlines the importance of oxidation-reduction potential and tryptophan metabolism in primed pluripotency. Cell Biosci. 2019, 9, 71. [Google Scholar] [CrossRef]
- Tsogtbaatar, E.; Landin, C.; Minter-Dykhouse, K.; Folmes, C.D.L. Energy Metabolism Regulates Stem Cell Pluripotency. Front. Cell Dev. Biol. 2020, 8, 87. [Google Scholar] [CrossRef]
- Takahashi, S.; Kobayashi, S.; Hiratani, I. Epigenetic differences between naïve and primed pluripotent stem cells. Cell. Mol. Life Sci. 2018, 75, 1191–1203. [Google Scholar] [CrossRef] [PubMed]
- Ghimire, S.; Van der Jeught, M.; Neupane, J.; Roost, M.S.; Anckaert, J.; Popovic, M.; Van Nieuwerburgh, F.; Mestdagh, P.; Vandesompele, J.; Deforce, D.; et al. Comparative analysis of naive, primed and ground state pluripotency in mouse embryonic stem cells originating from the same genetic background. Sci. Rep. 2018, 8, 5884. [Google Scholar] [CrossRef] [PubMed]
- Yamaji, M.; Ueda, J.; Hayashi, K.; Ohta, H.; Yabuta, Y.; Kurimoto, K.; Nakato, R.; Yamada, Y.; Shirahige, K.; Saitou, M. PRDM14 Ensures Naive Pluripotency through Dual Regulation of Signaling and Epigenetic Pathways in Mouse Embryonic Stem Cells. Cell Stem Cell 2013, 12, 368–382. [Google Scholar] [CrossRef]
- Chia, N.-Y.; Chan, Y.-S.; Feng, B.; Lu, X.; Orlov, Y.L.; Moreau, D.; Kumar, P.; Yang, L.; Jiang, J.; Lau, M.-S.; et al. A genome-wide RNAi screen reveals determinants of human embryonic stem cell identity. Nature 2010, 468, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Ginis, I.; Luo, Y.; Miura, T.; Thies, S.; Brandenberger, R.; Gerecht-Nir, S.; Amit, M.; Hoke, A.; Carpenter, M.K.; Itskovitz-Eldor, J.; et al. Differences between human and mouse embryonic stem cells. Dev. Biol. 2004, 269, 360–380. [Google Scholar] [CrossRef]
- Smith, Z.D.; Chan, M.M.; Humm, K.C.; Karnik, R.; Mekhoubad, S.; Regev, A.; Eggan, K.; Meissner, A. DNA methylation dynamics of the human preimplantation embryo. Nature 2014, 511, 611–615. [Google Scholar] [CrossRef]
- O’Leary, T.; Heindryckx, B.; Lierman, S.; van Bruggen, D.; Goeman, J.J.; Vandewoestyne, M.; Deforce, D.; Lopes, S.M.C.D.S.; De Sutter, P. Tracking the progression of the human inner cell mass during embryonic stem cell derivation. Nat. Biotechnol. 2012, 30, 278–282. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Aksoy, I.; Gonnot, F.; Osteil, P.; Aubry, M.; Hamela, C.; Rognard, C.; Hochard, A.; Voisin, S.; Fontaine, E.; et al. Reinforcement of STAT3 activity reprogrammes human embryonic stem cells to naive-like pluripotency. Nat. Commun. 2015, 6, 7095. [Google Scholar] [CrossRef]
- Chan, Y.-S.; Göke, J.; Ng, J.-H.; Lu, X.; Gonzales, K.A.; Tan, C.-P.; Tng, W.-Q.; Hong, Z.-Z.; Lim, Y.-S.; Ng, H.-H. Induction of a Human Pluripotent State with Distinct Regulatory Circuitry that Resembles Preimplantation Epiblast. Cell Stem Cell 2013, 13, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Guo, G.; von Meyenn, F.; Rostovskaya, M.; Clarke, J.; Dietmann, S.; Baker, D.; Sahakyan, A.; Myers, S.; Bertone, P.; Reik, W.; et al. Epigenetic resetting of human pluripotency. Development 2017, 144, 2748–2763. [Google Scholar] [CrossRef] [PubMed]
- Guo, G.; von Meyenn, F.; Santos, F.; Chen, Y.; Reik, W.; Bertone, P.; Smith, A.; Nichols, J. Naive Pluripotent Stem Cells Derived Directly from Isolated Cells of the Human Inner Cell Mass. Stem Cell Rep. 2016, 6, 437–446. [Google Scholar] [CrossRef]
- Gafni, O.; Weinberger, L.; Mansour, A.A.; Manor, Y.S.; Chomsky, E.; Ben-Yosef, D.; Kalma, Y.; Viukov, S.; Maza, I.; Zviran, A.; et al. Derivation of novel human ground state naive pluripotent stem cells. Nature 2013, 504, 282–286. [Google Scholar] [CrossRef] [PubMed]
- Theunissen, T.; Friedli, M.; He, Y.; Planet, E.; O’neil, R.C.; Markoulaki, S.; Pontis, J.; Wang, H.; Iouranova, A.; Imbeault, M.; et al. Molecular Criteria for Defining the Naive Human Pluripotent State. Cell Stem Cell 2016, 19, 502–515. [Google Scholar] [CrossRef]
- Takashima, Y.; Guo, G.; Loos, R.; Nichols, J.; Ficz, G.; Krueger, F.; Oxley, D.; Santos, F.; Clarke, J.; Mansfield, W.; et al. Resetting Transcription Factor Control Circuitry toward Ground-State Pluripotency in Human. Cell 2014, 158, 1254–1269. [Google Scholar] [CrossRef]
- Qin, H.; Hejna, M.; Liu, Y.; Percharde, M.; Wossidlo, M.; Blouin, L.; Durruthy-Durruthy, J.; Wong, P.; Qi, Z.; Yu, J.; et al. YAP Induces Human Naive Pluripotency. Cell Rep. 2016, 14, 2301–2312. [Google Scholar] [CrossRef]
- Liu, X.; Nefzger, C.; Rossello, F.J.; Chen, J.; Knaupp, A.; Firas, J.; Ford, E.; Pflueger, J.; Paynter, J.M.; Chy, H.S.; et al. Comprehensive characterization of distinct states of human naive pluripotency generated by reprogramming. Nat. Methods 2017, 14, 1055–1062. [Google Scholar] [CrossRef]
- Hanna, J.; Cheng, A.W.; Saha, K.; Kim, J.; Lengner, C.J.; Soldner, F.; Cassady, J.P.; Muffat, J.; Carey, B.W.; Jaenisch, R. Human embryonic stem cells with biological and epigenetic characteristics similar to those of mouse ESCs. Proc. Natl. Acad. Sci. USA 2010, 107, 9222–9227. [Google Scholar] [CrossRef]
- Fischer, L.A.; Khan, S.A.; Theunissen, T.W. Induction of Human Naïve Pluripotency Using 5i/L/A Medium. Methods Mol. Biol. 2022, 2416, 13–28. [Google Scholar] [CrossRef]
- Theunissen, T.W.; Powell, B.E.; Wang, H.; Mitalipova, M.; Faddah, D.A.; Reddy, J.; Fan, Z.P.; Maetzel, D.; Ganz, K.; Shi, L.; et al. Systematic Identification of Culture Conditions for Induction and Maintenance of Naive Human Pluripotency. Cell Stem Cell 2014, 15, 471–487. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.W.; Joo, J.Y.; Hong, Y.J.; Kim, J.S.; Song, H.; Lee, J.W.; Wu, G.; Schöler, H.R.; Do, J.T. Distinct Enhancer Activity of Oct4 in Naive and Primed Mouse Pluripotency. Stem Cell Rep. 2016, 7, 911–926. [Google Scholar] [CrossRef] [PubMed]
- Bayerl, J.; Ayyash, M.; Shani, T.; Manor, Y.S.; Gafni, O.; Massarwa, R.; Kalma, Y.; Aguilera-Castrejon, A.; Zerbib, M.; Amir, H.; et al. Principles of signaling pathway modulation for enhancing human naive pluripotency induction. Cell Stem Cell 2021, 28, 1549–1565.e12. [Google Scholar] [CrossRef] [PubMed]
- Pastor, W.A.; Chen, D.; Liu, W.; Kim, R.; Sahakyan, A.; Lukianchikov, A.; Plath, K.; Jacobsen, S.E.; Clark, A.T. Naive Human Pluripotent Cells Feature a Methylation Landscape Devoid of Blastocyst or Germline Memory. Cell Stem Cell 2016, 18, 323–329. [Google Scholar] [CrossRef]
- Khan, S.A.; Park, K.-M.; Fischer, L.A.; Dong, C.; Lungjangwa, T.; Jimenez, M.; Casalena, D.; Chew, B.; Dietmann, S.; Auld, D.S.; et al. Probing the signaling requirements for naive human pluripotency by high-throughput chemical screening. Cell Rep. 2021, 35, 109233. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-H.; Hao, J.; Zhang, C.; Duan, F.-F.; Chiu, Y.-T.; Shi, M.; Huang, X.; Yang, J.; Cao, H.; Wang, Y. KLF17 promotes human naive pluripotency through repressing MAPK3 and ZIC2. Sci. China Life Sci. 2022, 65, 1985–1997. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhang, X.; Yi, L.; Hou, Z.; Chen, J.; Kou, X.; Zhao, Y.; Wang, H.; Sun, X.-F.; Jiang, C.; et al. Naïve Induced Pluripotent Stem Cells Generated From β-Thalassemia Fibroblasts Allow Efficient Gene Correction With CRISPR/Cas9. Stem Cells Transl. Med. 2016, 5, 267. [Google Scholar] [CrossRef]
- Rossant, J.; Tam, P.P. New Insights into Early Human Development: Lessons for Stem Cell Derivation and Differentiation. Cell Stem Cell 2017, 20, 18–28. [Google Scholar] [CrossRef]
- Zimmerlin, L.; Park, T.S.; Zambidis, E.T.; Ward, E.; Twaroski, K.; Tolar, J.; Birbrair, A.; Borges, I.D.T.; Sena, I.F.G.; Almeida, G.G.; et al. Capturing Human Naïve Pluripotency in the Embryo and in the Dish. Stem Cells Dev. 2017, 26, 1141–1161. [Google Scholar] [CrossRef]
- Zhou, J.; Hu, J.; Wang, Y.; Gao, S. Induction and application of human naive pluripotency. Cell Rep. 2023, 42, 112379. [Google Scholar] [CrossRef]
- Bi, Y.; Tu, Z.; Zhang, Y.; Yang, P.; Guo, M.; Zhu, X.; Zhao, C.; Zhou, J.; Wang, H.; Wang, Y.; et al. Identification of ALPPL2 as a Naive Pluripotent State-Specific Surface Protein Essential for Human Naive Pluripotency Regulation. Cell Rep. 2020, 30, 3917–3931.e5. [Google Scholar] [CrossRef]
- Ai, Z.; Xiang, X.; Xiang, Y.; Szczerbinska, I.; Qian, Y.; Xu, X.; Ma, C.; Su, Y.; Gao, B.; Shen, H.; et al. Krüppel-like factor 5 rewires NANOG regulatory network to activate human naive pluripotency specific LTR7Ys and promote naive pluripotency. Cell Rep. 2022, 40, 111240. [Google Scholar] [CrossRef] [PubMed]
- Göke, J.; Ng, H.H. CTRL + INSERT: Retrotransposons and their contribution to regulation and innovation of the transcriptome. EMBO Rep. 2016, 17, 1131–1144. [Google Scholar] [CrossRef] [PubMed]
- Göke, J.; Lu, X.; Chan, Y.-S.; Ng, H.-H.; Ly, L.-H.; Sachs, F.; Szczerbinska, I. Dynamic Transcription of Distinct Classes of Endogenous Retroviral Elements Marks Specific Populations of Early Human Embryonic Cells. Cell Stem Cell 2015, 16, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Avilion, A.A.; Nicolis, S.K.; Pevny, L.H.; Perez, L.; Vivian, N.; Lovell-Badge, R. Multipotent cell lineages in early mouse development depend on SOX2 function. Genes Dev. 2003, 17, 126–140. [Google Scholar] [CrossRef]
- Masui, S.; Nakatake, Y.; Toyooka, Y.; Shimosato, D.; Yagi, R.; Takahashi, K.; Okochi, H.; Okuda, A.; Matoba, R.; Sharov, A.A.; et al. Pluripotency governed by Sox2 via regulation of Oct3/4 expression in mouse embryonic stem cells. Nat. Cell Biol. 2007, 9, 625–635. [Google Scholar] [CrossRef]
- Khan, S.A.; Audergon, P.N.C.B.; Payer, B. X-chromosome activity in naive human pluripotent stem cells-are we there yet? Stem Cell Investig. 2017, 4, 54. [Google Scholar] [CrossRef]
- Chour, T.; Tian, L.; Lau, E.; Thomas, D.; Itzhaki, I.; Malak, O.; Zhang, J.Z.; Qin, X.; Wardak, M.; Liu, Y.; et al. Method for selective ablation of undifferentiated human pluripotent stem cell populations for cell-based therapies. J. Clin. Investig. 2021, 6, e142000. [Google Scholar] [CrossRef]
- Mallanna, S.K.; Rizzino, A. Systems biology provides new insights into the molecular mechanisms that control the fate of embryonic stem cells. J. Cell. Physiol. 2012, 227, 27–34. [Google Scholar] [CrossRef]
- Huang, G.; Ye, S.; Zhou, X.; Liu, D.; Ying, Q.-L. Molecular basis of embryonic stem cell self-renewal: From signaling pathways to pluripotency network. Cell. Mol. Life Sci. 2015, 72, 1741–1757. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varzideh, F.; Gambardella, J.; Kansakar, U.; Jankauskas, S.S.; Santulli, G. Molecular Mechanisms Underlying Pluripotency and Self-Renewal of Embryonic Stem Cells. Int. J. Mol. Sci. 2023, 24, 8386. https://doi.org/10.3390/ijms24098386
Varzideh F, Gambardella J, Kansakar U, Jankauskas SS, Santulli G. Molecular Mechanisms Underlying Pluripotency and Self-Renewal of Embryonic Stem Cells. International Journal of Molecular Sciences. 2023; 24(9):8386. https://doi.org/10.3390/ijms24098386
Chicago/Turabian StyleVarzideh, Fahimeh, Jessica Gambardella, Urna Kansakar, Stanislovas S. Jankauskas, and Gaetano Santulli. 2023. "Molecular Mechanisms Underlying Pluripotency and Self-Renewal of Embryonic Stem Cells" International Journal of Molecular Sciences 24, no. 9: 8386. https://doi.org/10.3390/ijms24098386