
The Interplay between Gut Microbiota and Parkinson’s Disease: Implications on Diagnosis and Treatment
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Parkinson’s Disease and Gut Microbiota: Links and Mechanisms
2.1. Gut Microbiota Dysregulation in PD
2.2. Gut Permeability in PD
3. Parkinson’s Disease and Gut Microbiota: Biomarkers and Drug Interactions
3.1. Gut Microbiota-Based PD Biomarkers
3.2. Gut Microbiota-Drug Interactions
4. Parkinson’s Disease and Gut Microbiota: Therapeutic Approaches
4.1. Gut Microbiota-Based PD Interventions: Antibiotics
4.2. Gut Microbiota-Based PD Interventions: Probiotics
4.2.1. Preclinical Studies on Probiotics Supplementation in PD
4.2.2. Clinical Studies on Probiotics Supplementation in PD
4.3. Gut Microbiota-Based PD Interventions: Prebiotics
4.4. Gut Microbiota-Based PD Interventions: Diet
4.4.1. Mediterranean Diet
4.4.2. Ketogenic Diet
4.4.3. Omega-3 Fatty Acids
4.5. Gut Microbiota-Based PD Interventions: Fecal Microbiota Transplantation
5. Oral and Nasal Microbiota: Other Important Districts Involved in the Disease
6. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sousa-Fraguas, M.C.; Rodríguez-Fuentes, G.; Conejo, N.M. Frailty and Cognitive Impairment in Parkinson’s Disease: A Systematic Review. Neurol. Sci. 2022, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Leta, V.; Urso, D.; Batzu, L.; Lau, Y.H.; Mathew, D.; Boura, I.; Raeder, V.; Falup-Pecurariu, C.; van Wamelen, D.; Ray Chaudhuri, K. Viruses, Parkinsonism and Parkinson’s Disease: The Past, Present and Future. J. Neural Transm. 2022, 129, 1119–1132. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Wang, X.; Pan, F.; Mao, Z. The Relationship between Parkinson’s Disease and Gastrointestinal Diseases. Front. Aging Neurosci. 2022, 14, 955919. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Berardelli, A.; Bhattacharya, A.; Bologna, M.; Chen, K.-H.S.; Fasano, A.; Helmich, R.C.; Hutchison, W.D.; Kamble, N.; Kühn, A.A.; et al. Clinical Neurophysiology of Parkinson’s Disease and Parkinsonism. Clin. Neurophysiol. Pract 2022, 7, 201–227. [Google Scholar] [CrossRef] [PubMed]
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The Gut-Brain Axis: Interactions between Enteric Microbiota, Central and Enteric Nervous Systems. Ann. Gastroenterol. 2015, 28, 203–209. [Google Scholar]
- Klann, E.M.; Dissanayake, U.; Gurrala, A.; Farrer, M.; Shukla, A.W.; Ramirez-Zamora, A.; Mai, V.; Vedam-Mai, V. The Gut–Brain Axis and Its Relation to Parkinson’s Disease: A Review. Front. Aging Neurosci. 2022, 13, 782082. [Google Scholar] [CrossRef] [PubMed]
- Thangaleela, S.; Sivamaruthi, B.S.; Kesika, P.; Bharathi, M.; Chaiyasut, C. Role of the Gut–Brain Axis, Gut Microbial Composition, Diet, and Probiotic Intervention in Parkinson’s Disease. Microorganisms 2022, 10, 1544. [Google Scholar] [CrossRef] [PubMed]
- Sherman, M.P.; Zaghouani, H.; Niklas, V. Gut Microbiota, the Immune System, and Diet Influence the Neonatal Gut–Brain Axis. Pediatr. Res. 2015, 77, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.; Jheeta, S.; Fuentes, H.V.; Palacios-Pérez, M. Feeding Our Microbiota: Stimulation of the Immune/Semiochemical System and the Potential Amelioration of Non-Communicable Diseases. Life 2022, 12, 1197. [Google Scholar] [CrossRef] [PubMed]
- Hoyles, L.; Snelling, T.; Umlai, U.-K.; Nicholson, J.K.; Carding, S.R.; Glen, R.C.; McArthur, S. Microbiome–Host Systems Interactions: Protective Effects of Propionate upon the Blood–Brain Barrier. Microbiome 2018, 6, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, S.M.-S.; Mohajeri, M.H. The Role of Gut Bacterial Metabolites in Brain Development, Aging and Disease. Nutrients 2021, 13, 732. [Google Scholar] [CrossRef] [PubMed]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids From Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirayama, M.; Ohno, K. Parkinson’s Disease and Gut Microbiota. Ann. Nutr. Metab. 2021, 77, 28–35. [Google Scholar] [CrossRef]
- Grathwohl, S.A.; Steiner, J.A.; Britschgi, M.; Brundin, P. Mind the Gut: Secretion of α-Synuclein by Enteric Neurons. J. Neurochem. 2013, 125, 487–490. [Google Scholar] [CrossRef] [PubMed]
- Sampson, T.R.; Challis, C.; Jain, N.; Moiseyenko, A.; Ladinsky, M.S.; Shastri, G.G.; Thron, T.; Needham, B.D.; Horvath, I.; Debelius, J.W.; et al. A Gut Bacterial Amyloid Promotes α-Synuclein Aggregation and Motor Impairment in Mice. Elife 2020, 9, e53111. [Google Scholar] [CrossRef] [PubMed]
- Mulak, A. Brain-Gut-Microbiota Axis in Parkinson’s Disease. World J. Gastroenterol. 2015, 21, 10609. [Google Scholar] [CrossRef] [PubMed]
- Schemann, M.; Neunlist, M. The Human Enteric Nervous System. Neurogastroenterol. Motil. 2004, 16, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Szurszewski, J.H. Physiology of Mammalian Prevertebral Ganglia. Annu. Rev. Physiol. 1981, 43, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Dogra, N.; Mani, R.J.; Katare, D.P. The Gut-Brain Axis: Two Ways Signaling in Parkinson’s Disease. Cell Mol. Neurobiol. 2022, 42, 315–332. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.G.; Stribinskis, V.; Rane, M.J.; Demuth, D.R.; Gozal, E.; Roberts, A.M.; Jagadapillai, R.; Liu, R.; Choe, K.; Shivakumar, B.; et al. Exposure to the Functional Bacterial Amyloid Protein Curli Enhances Alpha-Synuclein Aggregation in Aged Fischer 344 Rats and Caenorhabditis Elegans. Sci. Rep. 2016, 6, 34477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murros, K.E.; Huynh, V.A.; Takala, T.M.; Saris, P.E.J. Desulfovibrio Bacteria Are Associated With Parkinson’s Disease. Front. Cell Infect Microbiol. 2021, 11, 652617. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.-Q.; Wang, Y.-L.; Yuan, B.-S.; Yuan, X.; Hou, X.-O.; Bian, J.-S.; Liu, C.-F.; Hu, L.-F. Impaired CBS-H2S Signaling Axis Contributes to MPTP-Induced Neurodegeneration in a Mouse Model of Parkinson’s Disease. Brain Behav. Immun. 2018, 67, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Murros, K.E. Hydrogen Sulfide Produced by Gut Bacteria May Induce Parkinson’s Disease. Cells 2022, 11, 978. [Google Scholar] [CrossRef] [PubMed]
- Dinan, K.; Dinan, T.G. Gut Microbes and Neuropathology: Is There a Causal Nexus? Pathogens 2022, 11, 796. [Google Scholar] [CrossRef]
- Ettinger, S. Diet, Gut Microbiome, and Cognitive Decline. Curr. Nutr. Rep. 2022. [Google Scholar] [CrossRef]
- Costa, H.N.; Esteves, A.R.; Empadinhas, N.; Cardoso, S.M. Parkinson’s Disease: A Multisystem Disorder. Neurosci. Bull 2022. [Google Scholar] [CrossRef]
- Choi, J.; Kim, B.-R.; Akuzum, B.; Chang, L.; Lee, J.-Y.; Kwon, H.-K. TREGking From Gut to Brain: The Control of Regulatory T Cells Along the Gut-Brain Axis. Front. Immunol. 2022, 13, 3193. [Google Scholar] [CrossRef]
- Pascale, A.; Marchesi, N.; Marelli, C.; Coppola, A.; Luzi, L.; Govoni, S.; Giustina, A.; Gazzaruso, C. Microbiota and Metabolic Diseases. Endocrine 2018, 61, 357–371. [Google Scholar] [CrossRef]
- Pascale, A.; Marchesi, N.; Govoni, S.; Barbieri, A. Targeting the Microbiota in Pharmacology of Psychiatric Disorders. Pharmacol. Res. 2020, 157, 104856. [Google Scholar] [CrossRef]
- Varesi, A.; Pierella, E.; Romeo, M.; Piccini, G.B.; Alfano, C.; Bjørklund, G.; Oppong, A.; Ricevuti, G.; Esposito, C.; Chirumbolo, S.; et al. The Potential Role of Gut Microbiota in Alzheimer’s Disease: From Diagnosis to Treatment. Nutrients 2022, 14, 668. [Google Scholar] [CrossRef]
- Cattaneo, A.; Cattane, N.; Galluzzi, S.; Provasi, S.; Lopizzo, N.; Festari, C.; Ferrari, C.; Guerra, U.P.; Paghera, B.; Muscio, C.; et al. Association of Brain Amyloidosis with Pro-Inflammatory Gut Bacterial Taxa and Peripheral Inflammation Markers in Cognitively Impaired Elderly. Neurobiol. Aging 2017, 49, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Vogt, N.M.; Kerby, R.L.; Dill-McFarland, K.A.; Harding, S.J.; Merluzzi, A.P.; Johnson, S.C.; Carlsson, C.M.; Asthana, S.; Zetterberg, H.; Blennow, K.; et al. Gut Microbiome Alterations in Alzheimer’s Disease. Sci. Rep. 2017, 7, 13537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varesi, A.; Deumer, U.-S.; Ananth, S.; Ricevuti, G. The Emerging Role of Gut Microbiota in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS): Current Evidence and Potential Therapeutic Applications. J. Clin. Med. 2021, 10, 5077. [Google Scholar] [CrossRef] [PubMed]
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Ostolaza, H.; Martín, C. Pathophysiology of Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2020, 21, 6275. [Google Scholar] [CrossRef]
- Abenavoli, L.; Scarpellini, E.; Colica, C.; Boccuto, L.; Salehi, B.; Sharifi-Rad, J.; Aiello, V.; Romano, B.; de Lorenzo, A.; Izzo, A.A.; et al. Gut Microbiota and Obesity: A Role for Probiotics. Nutrients 2019, 11, 2690. [Google Scholar] [CrossRef] [Green Version]
- Gerhardt, S.; Mohajeri, M. Changes of Colonic Bacterial Composition in Parkinson’s Disease and Other Neurodegenerative Diseases. Nutrients 2018, 10, 708. [Google Scholar] [CrossRef] [Green Version]
- Weiss, G.A.; Hennet, T. Mechanisms and Consequences of Intestinal Dysbiosis. Cell. Mol. Life Sci. 2017, 74, 2959–2977. [Google Scholar] [CrossRef] [Green Version]
- Schwabe, R.F.; Jobin, C. The Microbiome and Cancer. Nat. Rev. Cancer 2013, 13, 800–812. [Google Scholar] [CrossRef] [Green Version]
- Perez-Pardo, P.; Dodiya, H.B.; Engen, P.A.; Naqib, A.; Forsyth, C.B.; Green, S.J.; Garssen, J.; Keshavarzian, A.; Kraneveld, A.D. Gut Bacterial Composition in a Mouse Model of Parkinson’s Disease. Benef. Microbes 2018, 9, 799–814. [Google Scholar] [CrossRef] [Green Version]
- Sampson, T.R.; Debelius, J.W.; Thron, T.; Janssen, S.; Shastri, G.G.; Ilhan, Z.E.; Challis, C.; Schretter, C.E.; Rocha, S.; Gradinaru, V.; et al. Gut Microbiota Regulate Motor Deficits and Neuroinflammation in a Model of Parkinson’s Disease. Cell 2016, 167, 1469–1480.e12. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.-F.; Shen, Y.-Q. Dysbiosis of Gut Microbiota and Microbial Metabolites in Parkinson’s Disease. Ageing Res. Rev. 2018, 45, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Qian, Y.; Xu, S.; Song, Y.; Xiao, Q. Longitudinal Analysis of Fecal Microbiome and Pathologic Processes in a Rotenone Induced Mice Model of Parkinson’s Disease. Front. Aging Neurosci. 2018, 9, 441. [Google Scholar] [CrossRef] [Green Version]
- Lai, F.; Jiang, R.; Xie, W.; Liu, X.; Tang, Y.; Xiao, H.; Gao, J.; Jia, Y.; Bai, Q. Intestinal Pathology and Gut Microbiota Alterations in a Methyl-4-Phenyl-1,2,3,6-Tetrahydropyridine (MPTP) Mouse Model of Parkinson’s Disease. Neurochem. Res. 2018, 43, 1986–1999. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Huan, F.; Wang, J.; Xie, X.; Yu, G.; Wang, X.; Jiang, L.; Gao, R.; Xiao, H.; Ding, H.; et al. 1-Methyl-4-Phenyl-1,2,3,6-Tetrahydropyridine Induced Parkinson’s Disease in Mouse: Potential Association between Neurotransmitter Disturbance and Gut Microbiota Dysbiosis. ACS Chem. Neurosci. 2020, 11, 3366–3376. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Nishiwaki, H.; Ito, M.; Iwaoka, K.; Takahashi, K.; Suzuki, Y.; Taguchi, K.; Yamahara, K.; Tsuboi, Y.; Kashihara, K.; et al. Altered Gut Microbiota in Parkinson’s Disease Patients with Motor Complications. Park. Relat. Disord. 2022, 95, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Cassani, E.; Barichella, M.; Cancello, R.; Cavanna, F.; Iorio, L.; Cereda, E.; Bolliri, C.; Zampella Maria, P.; Bianchi, F.; Cestaro, B.; et al. Increased Urinary Indoxyl Sulfate (Indican): New Insights into Gut Dysbiosis in Parkinson’s Disease. Park. Relat. Disord. 2015, 21, 389–393. [Google Scholar] [CrossRef]
- Zhang, F.; Yue, L.; Fang, X.; Wang, G.; Li, C.; Sun, X.; Jia, X.; Yang, J.; Song, J.; Zhang, Y.; et al. Altered Gut Microbiota in Parkinson’s Disease Patients/Healthy Spouses and Its Association with Clinical Features. Park. Relat. Disord. 2020, 81, 84–88. [Google Scholar] [CrossRef]
- Wallen, Z.D.; Appah, M.; Dean, M.N.; Sesler, C.L.; Factor, S.A.; Molho, E.; Zabetian, C.P.; Standaert, D.G.; Payami, H. Characterizing Dysbiosis of Gut Microbiome in PD: Evidence for Overabundance of Opportunistic Pathogens. NPJ Park. Dis. 2020, 6, 11. [Google Scholar] [CrossRef]
- Nishiwaki, H.; Hamaguchi, T.; Ito, M.; Ishida, T.; Maeda, T.; Kashihara, K.; Tsuboi, Y.; Ueyama, J.; Shimamura, T.; Mori, H.; et al. Short-Chain Fatty Acid-Producing Gut Microbiota Is Decreased in Parkinson’s Disease but Not in Rapid-Eye-Movement Sleep Behavior Disorder. mSystems 2020, 5, e00797-20. [Google Scholar] [CrossRef]
- Keshavarzian, A.; Green, S.J.; Engen, P.A.; Voigt, R.M.; Naqib, A.; Forsyth, C.B.; Mutlu, E.; Shannon, K.M. Colonic Bacterial Composition in Parkinson’s Disease. Mov. Disord. 2015, 30, 1351–1360. [Google Scholar] [CrossRef]
- Lubomski, M.; Xu, X.; Holmes, A.J.; Muller, S.; Yang, J.Y.H.; Davis, R.L.; Sue, C.M. The Gut Microbiome in Parkinson’s Disease: A Longitudinal Study of the Impacts on Disease Progression and the Use of Device-Assisted Therapies. Front. Aging Neurosci. 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Nishiwaki, H.; Ito, M.; Ishida, T.; Hamaguchi, T.; Maeda, T.; Kashihara, K.; Tsuboi, Y.; Ueyama, J.; Shimamura, T.; Mori, H.; et al. Meta-Analysis of Gut Dysbiosis in Parkinson’s Disease. Mov. Disord. 2020, 35, 1626–1635. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.; Yue, Y.; He, T.; Huang, C.; Qu, B.; Lv, W.; Lai, H.-Y. The Association Between the Gut Microbiota and Parkinson’s Disease, a Meta-Analysis. Front. Aging Neurosci. 2021, 13, 636545. [Google Scholar] [CrossRef] [PubMed]
- Romano, S.; Savva, G.M.; Bedarf, J.R.; Charles, I.G.; Hildebrand, F.; Narbad, A. Meta-Analysis of the Parkinson’s Disease Gut Microbiome Suggests Alterations Linked to Intestinal Inflammation. NPJ Park. Dis. 2021, 7, 27. [Google Scholar] [CrossRef]
- Wallen, Z.D.; Stone, W.J.; Factor, S.A.; Molho, E.; Zabetian, C.P.; Standaert, D.G.; Payami, H. Exploring Human-Genome Gut-Microbiome Interaction in Parkinson’s Disease. NPJ Park. Dis. 2021, 7, 74. [Google Scholar] [CrossRef]
- Ji, T.; Huang, H.; Liu, J.; Peng, T.; Zhou, X.; Tan, Q.; Yuan, J.; Hua, H.; Ding, S.; Liu, H. Leveraging Sequence-based Faecal Microbial Community Survey Data to Identify Alterations in Gut Microbiota among Patients with Parkinson’s Disease. Eur. J. Neurosci. 2021, 53, 687–696. [Google Scholar] [CrossRef]
- Sokol, H.; Pigneur, B.; Watterlot, L.; Lakhdari, O.; Bermúdez-Humarán, L.G.; Gratadoux, J.-J.; Blugeon, S.; Bridonneau, C.; Furet, J.-P.; Corthier, G.; et al. Faecalibacterium Prausnitzii Is an Anti-Inflammatory Commensal Bacterium Identified by Gut Microbiota Analysis of Crohn Disease Patients. Proc. Natl. Acad. Sci. USA 2008, 105, 16731–16736. [Google Scholar] [CrossRef] [Green Version]
- Lenoir, M.; Martín, R.; Torres-Maravilla, E.; Chadi, S.; González-Dávila, P.; Sokol, H.; Langella, P.; Chain, F.; Bermúdez-Humarán, L.G. Butyrate Mediates Anti-Inflammatory Effects of Faecalibacterium Prausnitzii in Intestinal Epithelial Cells through Dact3. Gut Microbes 2020, 12, 1826748. [Google Scholar] [CrossRef] [PubMed]
- Cosma-Grigorov, A.; Meixner, H.; Mrochen, A.; Wirtz, S.; Winkler, J.; Marxreiter, F. Changes in Gastrointestinal Microbiome Composition in PD: A Pivotal Role of Covariates. Front. Neurol. 2020, 11, 1041. [Google Scholar] [CrossRef]
- Xie, A.; Ensink, E.; Li, P.; Gordevičius, J.; Marshall, L.L.; George, S.; Pospisilik, J.A.; Aho, V.T.E.; Houser, M.C.; Pereira, P.A.B.; et al. Bacterial Butyrate in Parkinson’s Disease Is Linked to Epigenetic Changes and Depressive Symptoms. Mov. Disord. 2022, 37, 1644–1653. [Google Scholar] [CrossRef]
- Tan, A.H.; Chong, C.W.; Lim, S.; Yap, I.K.S.; Teh, C.S.J.; Loke, M.F.; Song, S.; Tan, J.Y.; Ang, B.H.; Tan, Y.Q.; et al. Gut Microbial Ecosystem in Parkinson Disease: New Clinicobiological Insights from Multi-Omics. Ann. Neurol. 2021, 89, 546–559. [Google Scholar] [CrossRef]
- Chen, S.-J.; Chen, C.-C.; Liao, H.-Y.; Lin, Y.-T.; Wu, Y.-W.; Liou, J.-M.; Wu, M.-S.; Kuo, C.-H.; Lin, C.-H. Association of Fecal and Plasma Levels of Short-Chain Fatty Acids With Gut Microbiota and Clinical Severity in Patients With Parkinson Disease. Neurology 2022, 98, e848–e858. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wu, X.; Hu, X.; Wang, T.; Liang, S.; Duan, Y.; Jin, F.; Qin, B. Structural Changes of Gut Microbiota in Parkinson’s Disease and Its Correlation with Clinical Features. Sci. China Life Sci. 2017, 60, 1223–1233. [Google Scholar] [CrossRef] [PubMed]
- Mao, L.; Zhang, Y.; Tian, J.; Sang, M.; Zhang, G.; Zhou, Y.; Wang, P. Cross-Sectional Study on the Gut Microbiome of Parkinson’s Disease Patients in Central China. Front. Microbiol. 2021, 12, 728479. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Wang, P.; Chen, Z.; Sui, X.; Xie, X.; Zhang, J. Alteration of the Fecal Microbiota in North-Eastern Han Chinese Population with Sporadic Parkinson’s Disease. Neurosci. Lett. 2019, 707, 134297. [Google Scholar] [CrossRef] [PubMed]
- Kenna, J.E.; Chua, E.G.; Bakeberg, M.; Tay, A.; McGregor, S.; Gorecki, A.; Horne, M.; Marshall, B.; Mastaglia, F.L.; Anderton, R.S. Changes in the Gut Microbiome and Predicted Functional Metabolic Effects in an Australian Parkinson’s Disease Cohort. Front. Neurosci. 2021, 15, 756951. [Google Scholar] [CrossRef]
- Parada Venegas, D.; de la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [Green Version]
- Lin, A.; Zheng, W.; He, Y.; Tang, W.; Wei, X.; He, R.; Huang, W.; Su, Y.; Huang, Y.; Zhou, H.; et al. Gut Microbiota in Patients with Parkinson’s Disease in Southern China. Park. Relat. Disord. 2018, 53, 82–88. [Google Scholar] [CrossRef]
- Barichella, M.; Severgnini, M.; Cilia, R.; Cassani, E.; Bolliri, C.; Caronni, S.; Ferri, V.; Cancello, R.; Ceccarani, C.; Faierman, S.; et al. Unraveling Gut Microbiota in Parkinson’s Disease and Atypical Parkinsonism. Mov. Disord. 2019, 34, 396–405. [Google Scholar] [CrossRef]
- Li, Z.; Lu, G.; Luo, E.; Wu, B.; Li, Z.; Guo, J.; Xia, Z.; Zheng, C.; Su, Q.; Zeng, Y.; et al. Oral, Nasal, and Gut Microbiota in Parkinson’s Disease. Neuroscience 2022, 480, 65–78. [Google Scholar] [CrossRef]
- Pietrucci, D.; Cerroni, R.; Unida, V.; Farcomeni, A.; Pierantozzi, M.; Mercuri, N.B.; Biocca, S.; Stefani, A.; Desideri, A. Dysbiosis of Gut Microbiota in a Selected Population of Parkinson’s Patients. Park. Relat. Disord. 2019, 65, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Hopfner, F.; Künstner, A.; Müller, S.H.; Künzel, S.; Zeuner, K.E.; Margraf, N.G.; Deuschl, G.; Baines, J.F.; Kuhlenbäumer, G. Gut Microbiota in Parkinson Disease in a Northern German Cohort. Brain Res. 2017, 1667, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Yang, X.; Xu, S.; Wu, C.; Song, Y.; Qin, N.; Chen, S.-D.; Xiao, Q. Alteration of the Fecal Microbiota in Chinese Patients with Parkinson’s Disease. Brain Behav. Immun. 2018, 70, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Vascellari, S.; Melis, M.; Palmas, V.; Pisanu, S.; Serra, A.; Perra, D.; Santoru, M.L.; Oppo, V.; Cusano, R.; Uva, P.; et al. Clinical Phenotypes of Parkinson’s Disease Associate with Distinct Gut Microbiota and Metabolome Enterotypes. Biomolecules 2021, 11, 144. [Google Scholar] [CrossRef]
- Ransmayr, G.; Poewe, W.; Plörer, S.; Gerstenbrand, F.; Leidlmair, K.; Mayr, U. Prognostic Implications of the Motor Symptoms of Parkinson’s Disease with Respect to Clinical, Computertomographic and Psychometric Parameters. J. Neural. Transm. 1986, 67, 1–14. [Google Scholar] [CrossRef]
- Rajput, A.H.; Pahwa, R.; Pahwa, P.; Rajput, A. Prognostic Significance of the Onset Mode in Parkinsonism. Neurology 1993, 43, 829–830. [Google Scholar] [CrossRef] [PubMed]
- Zetusky, W.J.; Jankovic, J.; Pirozzolo, F.J. The Heterogeneity of Parkinson’s Disease: Clinical and Prognostic Implications. Neurology 1985, 35, 522–526. [Google Scholar] [CrossRef]
- Li, C.; Cui, L.; Yang, Y.; Miao, J.; Zhao, X.; Zhang, J.; Cui, G.; Zhang, Y. Gut Microbiota Differs Between Parkinson’s Disease Patients and Healthy Controls in Northeast China. Front. Mol. Neurosci. 2019, 12, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidal-Martinez, G.; Chin, B.; Camarillo, C.; Herrera, G.V.; Yang, B.; Sarosiek, I.; Perez, R.G. A Pilot Microbiota Study in Parkinson’s Disease Patients versus Control Subjects, and Effects of FTY720 and FTY720-Mitoxy Therapies in Parkinsonian and Multiple System Atrophy Mouse Models. J. Park. Dis. 2020, 10, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Heintz-Buschart, A.; Pandey, U.; Wicke, T.; Sixel-Döring, F.; Janzen, A.; Sittig-Wiegand, E.; Trenkwalder, C.; Oertel, W.H.; Mollenhauer, B.; Wilmes, P. The Nasal and Gut Microbiome in Parkinson’s Disease and Idiopathic Rapid Eye Movement Sleep Behavior Disorder. Mov. Disord. 2018, 33, 88–98. [Google Scholar] [CrossRef] [Green Version]
- Zapała, B.; Stefura, T.; Wójcik-Pędziwiatr, M.; Kabut, R.; Bałajewicz-Nowak, M.; Milewicz, T.; Dudek, A.; Stój, A.; Rudzińska-Bar, M. Differences in the Composition of Gut Microbiota between Patients with Parkinson’s Disease and Healthy Controls: A Cohort Study. J. Clin. Med. 2021, 10, 5698. [Google Scholar] [CrossRef]
- Derrien, M.; Vaughan, E.E.; Plugge, C.M.; de Vos, W.M. Akkermansia Muciniphila Gen. Nov., Sp. Nov., a Human Intestinal Mucin-Degrading Bacterium. Int. J. Syst. Evol. Microbiol. 2004, 54, 1469–1476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jangi, S.; Gandhi, R.; Cox, L.M.; Li, N.; von Glehn, F.; Yan, R.; Patel, B.; Mazzola, M.A.; Liu, S.; Glanz, B.L.; et al. Alterations of the Human Gut Microbiome in Multiple Sclerosis. Nat. Commun. 2016, 7, 12015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganesh, B.P.; Klopfleisch, R.; Loh, G.; Blaut, M. Commensal Akkermansia Muciniphila Exacerbates Gut Inflammation in Salmonella Typhimurium-Infected Gnotobiotic Mice. PLoS ONE 2013, 8, e74963. [Google Scholar] [CrossRef]
- Amorim Neto, D.P.; Bosque, B.P.; Pereira de Godoy, J.V.; Rodrigues, P.V.; Meneses, D.D.; Tostes, K.; Costa Tonoli, C.C.; Faustino de Carvalho, H.; González-Billault, C.; de Castro Fonseca, M. Akkermansia Muciniphila Induces Mitochondrial Calcium Overload and α -Synuclein Aggregation in an Enteroendocrine Cell Line. iScience 2022, 25, 103908. [Google Scholar] [CrossRef]
- Vandeputte, D.; Falony, G.; Vieira-Silva, S.; Tito, R.Y.; Joossens, M.; Raes, J. Stool Consistency Is Strongly Associated with Gut Microbiota Richness and Composition, Enterotypes and Bacterial Growth Rates. Gut 2016, 65, 57–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gobert, A.P.; Sagrestani, G.; Delmas, E.; Wilson, K.T.; Verriere, T.G.; Dapoigny, M.; Del’homme, C.; Bernalier-Donadille, A. The Human Intestinal Microbiota of Constipated-Predominant Irritable Bowel Syndrome Patients Exhibits Anti-Inflammatory Properties. Sci. Rep. 2016, 6, 39399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skrzydło-Radomańska, B.; Prozorow-Król, B.; Cichoż-Lach, H.; Majsiak, E.; Bierła, J.B.; Kosikowski, W.; Szczerbiński, M.; Gantzel, J.; Cukrowska, B. The Effectiveness of Synbiotic Preparation Containing Lactobacillus and Bifidobacterium Probiotic Strains and Short Chain Fructooligosaccharides in Patients with Diarrhea Predominant Irritable Bowel Syndrome—A Randomized Double-Blind, Placebo-Controlled Study. Nutrients 2020, 12, 1999. [Google Scholar] [CrossRef]
- Minato, T.; Maeda, T.; Fujisawa, Y.; Tsuji, H.; Nomoto, K.; Ohno, K.; Hirayama, M. Progression of Parkinson’s Disease Is Associated with Gut Dysbiosis: Two-Year Follow-up Study. PLoS ONE 2017, 12, e0187307. [Google Scholar] [CrossRef] [Green Version]
- Bolliri, C.; Fontana, A.; Cereda, E.; Barichella, M.; Cilia, R.; Ferri, V.; Caronni, S.; Calandrella, D.; Morelli, L.; Pezzoli, G. Gut Microbiota in Monozygotic Twins Discordant for Parkinson’s Disease. Ann. Neurol. 2022, 92, 631–636. [Google Scholar] [CrossRef]
- Porras, A.M.; Shi, Q.; Zhou, H.; Callahan, R.; Montenegro-Bethancourt, G.; Solomons, N.; Brito, I.L. Geographic Differences in Gut Microbiota Composition Impact Susceptibility to Enteric Infection. Cell Rep. 2021, 36, 109457. [Google Scholar] [CrossRef]
- Gupta, V.K.; Paul, S.; Dutta, C. Geography, Ethnicity or Subsistence-Specific Variations in Human Microbiome Composition and Diversity. Front. Microbiol. 2017, 8, 1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, Z.-Q.; Shen, L.-L.; Li, W.-W.; Fu, X.; Zeng, F.; Gui, L.; Lü, Y.; Cai, M.; Zhu, C.; Tan, Y.-L.; et al. Gut Microbiota Is Altered in Patients with Alzheimer’s Disease. J. Alzheimer’s Dis. 2018, 63, 1337–1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Y.; Pedersen, O. Gut Microbiota in Human Metabolic Health and Disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Oca, P.; Robles-Vera, I.; Sánchez-Roncero, A.; Escrivá, F.; Pérez-Vizcaíno, F.; Duarte, J.; Álvarez, C.; Fernández-Millán, E. Gut DYSBIOSIS and Altered Barrier Function Precedes the Appearance of Metabolic Syndrome in a Rat Model of Nutrient-Induced Catch-up Growth. J. Nutr. Biochem. 2020, 81, 108383. [Google Scholar] [CrossRef] [PubMed]
- Hertel, J.; Harms, A.C.; Heinken, A.; Baldini, F.; Thinnes, C.C.; Glaab, E.; Vasco, D.A.; Pietzner, M.; Stewart, I.D.; Wareham, N.J.; et al. Integrated Analyses of Microbiome and Longitudinal Metabolome Data Reveal Microbial-Host Interactions on Sulfur Metabolism in Parkinson’s Disease. Cell Rep. 2019, 29, 1767–1777.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, P.A.B.; Trivedi, D.K.; Silverman, J.; Duru, I.C.; Paulin, L.; Auvinen, P.; Scheperjans, F. Multiomics Implicate Gut Microbiota in Altered Lipid and Energy Metabolism in Parkinson’s Disease. NPJ Park. Dis. 2022, 8, 39. [Google Scholar] [CrossRef]
- Li, P.; Killinger, B.A.; Ensink, E.; Beddows, I.; Yilmaz, A.; Lubben, N.; Lamp, J.; Schilthuis, M.; Vega, I.E.; Woltjer, R.; et al. Gut Microbiota Dysbiosis Is Associated with Elevated Bile Acids in Parkinson’s Disease. Metabolites 2021, 11, 29. [Google Scholar] [CrossRef]
- Rosario, D.; Bidkhori, G.; Lee, S.; Bedarf, J.; Hildebrand, F.; le Chatelier, E.; Uhlen, M.; Ehrlich, S.D.; Proctor, G.; Wüllner, U.; et al. Systematic Analysis of Gut Microbiome Reveals the Role of Bacterial Folate and Homocysteine Metabolism in Parkinson’s Disease. Cell Rep. 2021, 34, 108807. [Google Scholar] [CrossRef]
- Romo-Vaquero, M.; Fernández-Villalba, E.; Gil-Martinez, A.-L.; Cuenca-Bermejo, L.; Espín, J.C.; Herrero, M.T.; Selma, M.V. Urolithins: Potential Biomarkers of Gut Dysbiosis and Disease Stage in Parkinson’s Patients. Food Funct. 2022, 13, 6306–6316. [Google Scholar] [CrossRef]
- Cirstea, M.S.; Yu, A.C.; Golz, E.; Sundvick, K.; Kliger, D.; Radisavljevic, N.; Foulger, L.H.; Mackenzie, M.; Huan, T.; Finlay, B.B.; et al. Microbiota Composition and Metabolism Are Associated With Gut Function in Parkinson’s Disease. Mov. Disord. 2020, 35, 1208–1217. [Google Scholar] [CrossRef]
- Yan, Z.; Yang, F.; Cao, J.; Ding, W.; Yan, S.; Shi, W.; Wen, S.; Yao, L. Alterations of Gut Microbiota and Metabolome with Parkinson’s Disease. Microb Pathog. 2021, 160, 105187. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-J.; Chen, C.-C.; Liao, H.-Y.; Wu, Y.-W.; Liou, J.-M.; Wu, M.-S.; Kuo, C.-H.; Lin, C.-H. Alteration of Gut Microbial Metabolites in the Systemic Circulation of Patients with Parkinson’s Disease. J. Park. Dis. 2022, 12, 1219–1230. [Google Scholar] [CrossRef] [PubMed]
- Voigt, R.M.; Wang, Z.; Brown, J.M.; Engen, P.A.; Naqib, A.; Goetz, C.G.; Hall, D.A.; Metman, L.V.; Shaikh, M.; Forsyth, C.B.; et al. Gut Microbial Metabolites in Parkinson’s Disease: Association with Lifestyle, Disease Characteristics, and Treatment Status. Neurobiol. Dis. 2022, 170, 105780. [Google Scholar] [CrossRef]
- Arrona Cardoza, P.; Spillane, M.B.; Morales Marroquin, E. Alzheimer’s Disease and Gut Microbiota: Does Trimethylamine N-Oxide (TMAO) Play a Role? Nutr. Rev. 2022, 80, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Kritsilis, M.; Rizou, S.V.; Koutsoudaki, P.; Evangelou, K.; Gorgoulis, V.; Papadopoulos, D. Ageing, Cellular Senescence and Neurodegenerative Disease. Int. J. Mol. Sci. 2018, 19, 2937. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Ke, Y.; Zhan, R.; Liu, C.; Zhao, M.; Zeng, A.; Shi, X.; Ji, L.; Cheng, S.; Pan, B.; et al. Trimethylamine-N-Oxide Promotes Brain Aging and Cognitive Impairment in Mice. Aging Cell 2018, 17, e12768. [Google Scholar] [CrossRef] [PubMed]
- Unger, M.M.; Spiegel, J.; Dillmann, K.-U.; Grundmann, D.; Philippeit, H.; Bürmann, J.; Faßbender, K.; Schwiertz, A.; Schäfer, K.-H. Short Chain Fatty Acids and Gut Microbiota Differ between Patients with Parkinson’s Disease and Age-Matched Controls. Park. Relat. Disord. 2016, 32, 66–72. [Google Scholar] [CrossRef]
- Huang, T.; Shi, H.; Xu, Y.; Ji, L. The Gut Microbiota Metabolite Propionate Ameliorates Intestinal Epithelial Barrier Dysfunction-Mediated Parkinson’s Disease via the AKT Signaling Pathway. Neuroreport 2021, 32, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Jiang, Z.; Pu, Y.; Chen, S.; Wang, T.; Wang, Y.; Xu, X.; Wang, S.; Jin, M.; Yao, Y.; et al. Serum Short-Chain Fatty Acids and Its Correlation with Motor and Non-Motor Symptoms in Parkinson’s Disease Patients. BMC Neurol. 2022, 22, 13. [Google Scholar] [CrossRef]
- Erny, D.; Hrabě de Angelis, A.L.; Jaitin, D.; Wieghofer, P.; Staszewski, O.; David, E.; Keren-Shaul, H.; Mahlakoiv, T.; Jakobshagen, K.; Buch, T.; et al. Host Microbiota Constantly Control Maturation and Function of Microglia in the CNS. Nat. Neurosci. 2015, 18, 965–977. [Google Scholar] [CrossRef] [PubMed]
- Ostendorf, F.; Metzdorf, J.; Gold, R.; Haghikia, A.; Tönges, L. Propionic Acid and Fasudil as Treatment against Rotenone Toxicity in an In Vitro Model of Parkinson’s Disease. Molecules 2020, 25, 2502. [Google Scholar] [CrossRef] [PubMed]
- Patnala, R.; Arumugam, T.V.; Gupta, N.; Dheen, S.T. HDAC Inhibitor Sodium Butyrate-Mediated Epigenetic Regulation Enhances Neuroprotective Function of Microglia During Ischemic Stroke. Mol. Neurobiol. 2017, 54, 6391–6411. [Google Scholar] [CrossRef] [PubMed]
- Rane, P.; Shields, J.; Heffernan, M.; Guo, Y.; Akbarian, S.; King, J.A. The Histone Deacetylase Inhibitor, Sodium Butyrate, Alleviates Cognitive Deficits in Pre-Motor Stage PD. Neuropharmacology 2012, 62, 2409–2412. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Kanthasamy, A.; Harischandra, D.S.; Kondru, N.; Ghosh, A.; Panicker, N.; Anantharam, V.; Rana, A.; Kanthasamy, A.G. Histone Hyperacetylation Up-Regulates Protein Kinase Cδ in Dopaminergic Neurons to Induce Cell Death. J. Biol. Chem. 2014, 289, 34743–34767. [Google Scholar] [CrossRef] [Green Version]
- Qiao, C.-M.; Sun, M.-F.; Jia, X.-B.; Li, Y.; Zhang, B.-P.; Zhao, L.-P.; Shi, Y.; Zhou, Z.-L.; Zhu, Y.-L.; Cui, C.; et al. Sodium Butyrate Exacerbates Parkinson’s Disease by Aggravating Neuroinflammation and Colonic Inflammation in MPTP-Induced Mice Model. Neurochem. Res. 2020, 45, 2128–2142. [Google Scholar] [CrossRef] [PubMed]
- van IJzendoorn, S.C.D.; Derkinderen, P. The Intestinal Barrier in Parkinson’s Disease: Current State of Knowledge. J. Park. Dis. 2019, 9, S323–S329. [Google Scholar] [CrossRef] [Green Version]
- Groschwitz, K.R.; Hogan, S.P. Intestinal Barrier Function: Molecular Regulation and Disease Pathogenesis. J. Allergy Clin. Immunol. 2009, 124, 3–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, M.; Kolodziejczyk, A.A.; Thaiss, C.A.; Elinav, E. Dysbiosis and the Immune System. Nat. Rev. Immunol. 2017, 17, 219–232. [Google Scholar] [CrossRef]
- Stadlbauer, V.; Engertsberger, L.; Komarova, I.; Feldbacher, N.; Leber, B.; Pichler, G.; Fink, N.; Scarpatetti, M.; Schippinger, W.; Schmidt, R.; et al. Dysbiosis, Gut Barrier Dysfunction and Inflammation in Dementia: A Pilot Study. BMC Geriatr. 2020, 20, 248. [Google Scholar] [CrossRef] [PubMed]
- Michielan, A.; D’Incà, R. Intestinal Permeability in Inflammatory Bowel Disease: Pathogenesis, Clinical Evaluation, and Therapy of Leaky Gut. Mediat. Inflamm 2015, 2015, 628157. [Google Scholar] [CrossRef] [PubMed]
- Hanning, N.; Edwinson, A.L.; Ceuleers, H.; Peters, S.A.; de Man, J.G.; Hassett, L.C.; de Winter, B.Y.; Grover, M. Intestinal Barrier Dysfunction in Irritable Bowel Syndrome: A Systematic Review. Therap. Adv. Gastroenterol. 2021, 14, 175628482199358. [Google Scholar] [CrossRef] [PubMed]
- Baizabal-Carvallo, J.F.; Alonso-Juarez, M. The Link between Gut Dysbiosis and Neuroinflammation in Parkinson’s Disease. Neuroscience 2020, 432, 160–173. [Google Scholar] [CrossRef] [PubMed]
- Anderson, G.; Seo, M.; Berk, M.; Carvalho, A.; Maes, M. Gut Permeability and Microbiota in Parkinson’s Disease: Role of Depression, Tryptophan Catabolites, Oxidative and Nitrosative Stress and Melatonergic Pathways. Curr. Pharm. Des. 2016, 22, 6142–6151. [Google Scholar] [CrossRef]
- Seguella, L.; Sarnelli, G.; Esposito, G. Leaky Gut, Dysbiosis, and Enteric Glia Activation: The Trilogy behind the Intestinal Origin of Parkinson’s Disease. Neural Regen. Res. 2020, 15, 1037. [Google Scholar] [CrossRef]
- Tan, A.H.; Lim, S.Y.; Lang, A.E. The Microbiome–Gut–Brain Axis in Parkinson Disease—from Basic Research to the Clinic. Nat. Rev. Neurol. 2022, 18, 476–495. [Google Scholar] [CrossRef]
- Loffredo, L.; Ettorre, E.; Zicari, A.M.; Inghilleri, M.; Nocella, C.; Perri, L.; Spalice, A.; Fossati, C.; de Lucia, M.C.; Pigozzi, F.; et al. Oxidative Stress and Gut-Derived Lipopolysaccharides in Neurodegenerative Disease: Role of NOX2. Oxid Med. Cell Longev. 2020, 2020, 8630275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drobny, A.; Ngo, P.A.; Neurath, M.F.; Zunke, F.; López-Posadas, R. Molecular Communication Between Neuronal Networks and Intestinal Epithelial Cells in Gut Inflammation and Parkinson’s Disease. Front. Med. 2021, 8, 8630275. [Google Scholar] [CrossRef]
- Pellegrini, C.; D’Antongiovanni, V.; Miraglia, F.; Rota, L.; Benvenuti, L.; di Salvo, C.; Testa, G.; Capsoni, S.; Carta, G.; Antonioli, L.; et al. Enteric α-Synuclein Impairs Intestinal Epithelial Barrier through Caspase-1-Inflammasome Signaling in Parkinson’s Disease before Brain Pathology. NPJ Park. Dis. 2022, 8, 9. [Google Scholar] [CrossRef]
- Clairembault, T.; Leclair-Visonneau, L.; Coron, E.; Bourreille, A.; le Dily, S.; Vavasseur, F.; Heymann, M.-F.; Neunlist, M.; Derkinderen, P. Structural Alterations of the Intestinal Epithelial Barrier in Parkinson’s Disease. Acta Neuropathol. Commun. 2015, 3, 12. [Google Scholar] [CrossRef] [Green Version]
- Gorecki, A.M.; Preskey, L.; Bakeberg, M.C.; Kenna, J.E.; Gildenhuys, C.; MacDougall, G.; Dunlop, S.A.; Mastaglia, F.L.; Akkari, P.A.; Koengten, F.; et al. Altered Gut Microbiome in Parkinson’s Disease and the Influence of Lipopolysaccharide in a Human α-Synuclein Over-Expressing Mouse Model. Front. Neurosci. 2019, 13, 839. [Google Scholar] [CrossRef]
- Schwiertz, A.; Spiegel, J.; Dillmann, U.; Grundmann, D.; Bürmann, J.; Faßbender, K.; Schäfer, K.-H.; Unger, M.M. Fecal Markers of Intestinal Inflammation and Intestinal Permeability Are Elevated in Parkinson’s Disease. Park. Relat. Disord. 2018, 50, 104–107. [Google Scholar] [CrossRef]
- Dumitrescu, L.; Marta, D.; Dănău, A.; Lefter, A.; Tulbă, D.; Cozma, L.; Manole, E.; Gherghiceanu, M.; Ceafalan, L.C.; Popescu, B.O. Serum and Fecal Markers of Intestinal Inflammation and Intestinal Barrier Permeability Are Elevated in Parkinson’s Disease. Front. Neurosci. 2021, 15, 689723. [Google Scholar] [CrossRef]
- Mulak, A.; Koszewicz, M.; Panek-Jeziorna, M.; Koziorowska-Gawron, E.; Budrewicz, S. Fecal Calprotectin as a Marker of the Gut Immune System Activation Is Elevated in Parkinson’s Disease. Front. Neurosci. 2019, 13, 992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Ai, P.; He, X.; Mo, C.; Zhang, Y.; Xu, S.; Lai, Y.; Qian, Y.; Xiao, Q. Parkinson’s Disease Is Associated with Impaired Gut–Blood Barrier for Short-Chain Fatty Acids. Mov. Disord. 2022, 37, 1634–1643. [Google Scholar] [CrossRef]
- Schoultz, I.; Keita, Å.V. The Intestinal Barrier and Current Techniques for the Assessment of Gut Permeability. Cells 2020, 9, 1909. [Google Scholar] [CrossRef]
- Mishra, A.; Makharia, G.K. Techniques of Functional and Motility Test: How to Perform and Interpret Intestinal Permeability. J. Neurogastroenterol. Motil 2012, 18, 443–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salat-Foix, D.; Tran, K.; Ranawaya, R.; Meddings, J.; Suchowersky, O. Increased Intestinal Permeability and Parkinson Disease Patients: Chicken or Egg? Can. J. Neurol. Sci. / J. Can. Des Sci. Neurol. 2012, 39, 185–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, K.N.; King, D.; Billington, D.; Barrett, J.A. Intestinal Permeability and Orocaecal Transit Time in Elderly Patients with Parkinson’s Disease. Postgrad. Med. J. 1996, 72, 164–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, X.-Y.; Yan, J.-T.; Zhang, X.-L.; Zhu, J.-X. Gastrointestinal Non-Motor Dysfunction in Parkinson’s Disease Model Rats with 6-Hydroxydopamine. Physiol. Res. 2019, 68, 295–303. [Google Scholar] [CrossRef]
- Jackson, A.; Engen, P.A.; Forsyth, C.B.; Shaikh, M.; Naqib, A.; Wilber, S.; Frausto, D.M.; Raeisi, S.; Green, S.J.; Bradaric, B.D.; et al. Intestinal Barrier Dysfunction in the Absence of Systemic Inflammation Fails to Exacerbate Motor Dysfunction and Brain Pathology in a Mouse Model of Parkinson’s Disease. Front. Neurol. 2022, 13, 689723. [Google Scholar] [CrossRef]
- Mohammad, S.; Thiemermann, C. Role of Metabolic Endotoxemia in Systemic Inflammation and Potential Interventions. Front. Immunol. 2021, 11, 594150. [Google Scholar] [CrossRef]
- Bhattacharyya, D.; Mohite, G.M.; Krishnamoorthy, J.; Gayen, N.; Mehra, S.; Navalkar, A.; Kotler, S.A.; Ratha, B.N.; Ghosh, A.; Kumar, R.; et al. Lipopolysaccharide from Gut Microbiota Modulates α-Synuclein Aggregation and Alters Its Biological Function. ACS Chem. Neurosci. 2019, 10, 2229–2236. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Wu, X.; Block, M.L.; Liu, Y.; Breese, G.R.; Hong, J.-S.; Knapp, D.J.; Crews, F.T. Systemic LPS Causes Chronic Neuroinflammation and Progressive Neurodegeneration. Glia 2007, 55, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, S.; Goto, S.; Tsuji, H.; Okuno, T.; Asahara, T.; Nomoto, K.; Shibata, A.; Fujisawa, Y.; Minato, T.; Okamoto, A.; et al. Intestinal Dysbiosis and Lowered Serum Lipopolysaccharide-Binding Protein in Parkinson’s Disease. PLoS ONE 2015, 10, e0142164. [Google Scholar] [CrossRef] [Green Version]
- Forsyth, C.B.; Shannon, K.M.; Kordower, J.H.; Voigt, R.M.; Shaikh, M.; Jaglin, J.A.; Estes, J.D.; Dodiya, H.B.; Keshavarzian, A. Increased Intestinal Permeability Correlates with Sigmoid Mucosa Alpha-Synuclein Staining and Endotoxin Exposure Markers in Early Parkinson’s Disease. PLoS ONE 2011, 6, e28032. [Google Scholar] [CrossRef] [Green Version]
- Pal, G.D.; Shaikh, M.; Forsyth, C.B.; Ouyang, B.; Keshavarzian, A.; Shannon, K.M. Abnormal Lipopolysaccharide Binding Protein as Marker of Gastrointestinal Inflammation in Parkinson Disease. Front. Neurosci. 2015, 9, 306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutsmann, T.; Müller, M.; Carroll, S.F.; MacKenzie, R.C.; Wiese, A.; Seydel, U. Dual Role of Lipopolysaccharide (LPS)-Binding Protein in Neutralization of LPS and Enhancement of LPS-Induced Activation of Mononuclear Cells. Infect Immun. 2001, 69, 6942–6950. [Google Scholar] [CrossRef] [Green Version]
- Minter, R.M.; Bi, X.; Ben-Josef, G.; Wang, T.; Hu, B.; Arbabi, S.; Hemmila, M.R.; Wang, S.C.; Remick, D.G.; Su, G.L. LPS-Binding Protein Mediates LPS-Induced Liver Injury and Mortality in the Setting of Biliary Obstruction. Am. J. Physiol.-Gastrointest. Liver Physiol. 2009, 296, G45–G54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.-H.; Chen, C.-C.; Chiang, H.-L.; Liou, J.-M.; Chang, C.-M.; Lu, T.-P.; Chuang, E.Y.; Tai, Y.-C.; Cheng, C.; Lin, H.-Y.; et al. Altered Gut Microbiota and Inflammatory Cytokine Responses in Patients with Parkinson’s Disease. J. Neuroinflamm. 2019, 16, 129. [Google Scholar] [CrossRef]
- Su, Y.; Liu, N.; Zhang, Z.; Li, H.; Ma, J.; Yuan, Y.; Shi, M.; Liu, J.; Zhao, Z.; Zhang, Z.; et al. Cholecystokinin and Glucagon-like Peptide-1 Analogues Regulate Intestinal Tight Junction, Inflammation, Dopaminergic Neurons and α-Synuclein Accumulation in the Colon of Two Parkinson’s Disease Mouse Models. Eur. J. Pharmacol. 2022, 926, 175029. [Google Scholar] [CrossRef]
- Tasselli, M.; Chaumette, T.; Paillusson, S.; Monnet, Y.; Lafoux, A.; Huchet-Cadiou, C.; Aubert, P.; Hunot, S.; Derkinderen, P.; Neunlist, M. Effects of Oral Administration of Rotenone on Gastrointestinal Functions in Mice. Neurogastroenterol. Motil. 2013, 25, e183–e193. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Yan, X.; Guo, J.; Xu, Q.; Tang, B.; Sun, Q. Recent Advances in Biomarkers for Parkinson’s Disease. Front. Aging Neurosci. 2018, 10, 305. [Google Scholar] [CrossRef]
- Li, T.; Le, W. Biomarkers for Parkinson’s Disease: How Good Are They? Neurosci. Bull 2020, 36, 183–194. [Google Scholar] [CrossRef]
- Delenclos, M.; Jones, D.R.; McLean, P.J.; Uitti, R.J. Biomarkers in Parkinson’s Disease: Advances and Strategies. Park. Relat. Disord. 2016, 22, S106–S110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farrokhi, V.; Nemati, R.; Nichols, F.C.; Yao, X.; Anstadt, E.; Fujiwara, M.; Grady, J.; Wakefield, D.; Castro, W.; Donaldson, J.; et al. Bacterial Lipodipeptide, Lipid 654, Is a Microbiome-Associated Biomarker for Multiple Sclerosis. Clin. Transl. Immunol. 2013, 2, e8. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Yang, X.; Xu, S.; Huang, P.; Li, B.; Du, J.; He, Y.; Su, B.; Xu, L.-M.; Wang, L.; et al. Gut Metagenomics-Derived Genes as Potential Biomarkers of Parkinson’s Disease. Brain 2020, 143, 2474–2489. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.; Rahmani, E.; Garcia, E.; Jacobs, J.P. Gastrointestinal Symptoms Are Predictive of Trajectories of Cognitive Functioning in de Novo Parkinson’s Disease. Park. Relat. Disord. 2020, 72, 7–12. [Google Scholar] [CrossRef]
- Fricova, D.; Harsanyiova, J.; Kralova Trancikova, A. Alpha-Synuclein in the Gastrointestinal Tract as a Potential Biomarker for Early Detection of Parkinson’s Disease. Int. J. Mol. Sci. 2020, 21, 8666. [Google Scholar] [CrossRef]
- Cilia, R.; Piatti, M.; Cereda, E.; Bolliri, C.; Caronni, S.; Ferri, V.; Cassani, E.; Bonvegna, S.; Ferrarese, C.; Zecchinelli, A.L.; et al. Does Gut Microbiota Influence the Course of Parkinson’s Disease? A 3-Year Prospective Exploratory Study in de Novo Patients. J. Park. Dis. 2021, 11, 159–170. [Google Scholar] [CrossRef]
- Pietrucci, D.; Teofani, A.; Unida, V.; Cerroni, R.; Biocca, S.; Stefani, A.; Desideri, A. Can Gut Microbiota Be a Good Predictor for Parkinson’s Disease? A Machine Learning Approach. Brain Sci. 2020, 10, 242. [Google Scholar] [CrossRef]
- Guo, X.; Tang, P.; Hou, C.; Chong, L.; Zhang, X.; Liu, P.; Chen, L.; Liu, Y.; Zhang, L.; Li, R. Integrated Microbiome and Host Transcriptome Profiles Link Parkinson’s Disease to Blautia Genus: Evidence From Feces, Blood, and Brain. Front. Microbiol. 2022, 13. [Google Scholar] [CrossRef]
- Lubomski, M.; Xu, X.; Holmes, A.J.; Muller, S.; Yang, J.Y.H.; Davis, R.L.; Sue, C.M. Nutritional Intake and Gut Microbiome Composition Predict Parkinson’s Disease. Front. Aging Neurosci. 2022, 14. [Google Scholar] [CrossRef]
- Chung, S.J.; Rim, J.H.; Ji, D.; Lee, S.; Yoo, H.S.; Jung, J.H.; Baik, K.; Choi, Y.; Ye, B.S.; Sohn, Y.H.; et al. Gut Microbiota-Derived Metabolite Trimethylamine N-Oxide as a Biomarker in Early Parkinson’s Disease. Nutrition 2021, 83, 111090. [Google Scholar] [CrossRef]
- He, X.; Qian, Y.; Xu, S.; Zhang, Y.; Mo, C.; Guo, W.; Yang, X.; Xiao, Q. Plasma Short-Chain Fatty Acids Differences in Multiple System Atrophy from Parkinson’s Disease. J. Park. Dis. 2021, 11, 1167–1176. [Google Scholar] [CrossRef]
- Nishiwaki, H.; Ito, M.; Hamaguchi, T.; Maeda, T.; Kashihara, K.; Tsuboi, Y.; Ueyama, J.; Yoshida, T.; Hanada, H.; Takeuchi, I.; et al. Short Chain Fatty Acids-Producing and Mucin-Degrading Intestinal Bacteria Predict the Progression of Early Parkinson’s Disease. NPJ Park. Dis. 2022, 8, 65. [Google Scholar] [CrossRef]
- Cerroni, R.; Pietrucci, D.; Teofani, A.; Chillemi, G.; Liguori, C.; Pierantozzi, M.; Unida, V.; Selmani, S.; Mercuri, N.B.; Stefani, A. Not Just a Snapshot: An Italian Longitudinal Evaluation of Stability of Gut Microbiota Findings in Parkinson’s Disease. Brain Sci. 2022, 12, 739. [Google Scholar] [CrossRef]
- Plassais, J.; Gbikpi-Benissan, G.; Figarol, M.; Scheperjans, F.; Gorochov, G.; Derkinderen, P.; Cervino, A.C.L. Gut Microbiome Alpha-Diversity Is Not a Marker of Parkinson’s Disease and Multiple Sclerosis. Brain Commun. 2021, 3, fcab113. [Google Scholar] [CrossRef]
- Boertien, J.M.; van der Zee, S.; Chrysou, A.; Gerritsen, M.J.J.; Jansonius, N.M.; Spikman, J.M.; van Laar, T.; Verwey, N.A.; van Harten, B.; Portman, A.T.; et al. Study Protocol of the DUtch PARkinson Cohort (DUPARC): A Prospective, Observational Study of de Novo Parkinson’s Disease Patients for the Identification and Validation of Biomarkers for Parkinson’s Disease Subtypes, Progression and Pathophysiology. BMC Neurol. 2020, 20, 245. [Google Scholar] [CrossRef]
- Falony, G.; Joossens, M.; Vieira-Silva, S.; Wang, J.; Darzi, Y.; Faust, K.; Kurilshikov, A.; Bonder, M.J.; Valles-Colomer, M.; Vandeputte, D.; et al. Population-Level Analysis of Gut Microbiome Variation. Science (1979) 2016, 352, 560–564. [Google Scholar] [CrossRef]
- Zhernakova, A.; Kurilshikov, A.; Bonder, M.J.; Tigchelaar, E.F.; Schirmer, M.; Vatanen, T.; Mujagic, Z.; Vila, A.V.; Falony, G.; Vieira-Silva, S.; et al. Population-Based Metagenomics Analysis Reveals Markers for Gut Microbiome Composition and Diversity. Science (1979) 2016, 352, 565–569. [Google Scholar] [CrossRef] [PubMed]
- Collins, S.L.; Patterson, A.D. The Gut Microbiome: An Orchestrator of Xenobiotic Metabolism. Acta Pharm. Sin. B 2020, 10, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; He, J.; Jia, W. The Influence of Gut Microbiota on Drug Metabolism and Toxicity. Expert Opin. Drug Metab. Toxicol. 2016, 12, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Vich Vila, A.; Collij, V.; Sanna, S.; Sinha, T.; Imhann, F.; Bourgonje, A.R.; Mujagic, Z.; Jonkers, D.M.A.E.; Masclee, A.A.M.; Fu, J.; et al. Impact of Commonly Used Drugs on the Composition and Metabolic Function of the Gut Microbiota. Nat. Commun. 2020, 11, 362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brüssow, H. Parkinson Disease, Levodopa and the Gut Microbiota—When Microbiology Meets Pharmacology. Environ. Microbiol. 2020, 22, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Keshavarzian, A.; Engen, P.; Bonvegna, S.; Cilia, R. The Gut Microbiome in Parkinson’s Disease: A Culprit or a Bystander? Prog. Brain Res. 2020, 252, 357–450. [Google Scholar]
- Zhang, X.; Han, Y.; Huang, W.; Jin, M.; Gao, Z. The Influence of the Gut Microbiota on the Bioavailability of Oral Drugs. Acta Pharm. Sin. B 2021, 11, 1789–1812. [Google Scholar] [CrossRef]
- Lubomski, M.; Xu, X.; Holmes, A.J.; Yang, J.Y.H.; Sue, C.M.; Davis, R.L. The Impact of Device-Assisted Therapies on the Gut Microbiome in Parkinson’s Disease. J. Neurol. 2022, 269, 780–795. [Google Scholar] [CrossRef]
- Salat, D.; Tolosa, E. Levodopa in the Treatment of Parkinson’s Disease: Current Status and New Developments. J. Park. Dis. 2013, 3, 255–269. [Google Scholar] [CrossRef] [Green Version]
- van Kessel, S.P.; Frye, A.K.; El-Gendy, A.O.; Castejon, M.; Keshavarzian, A.; van Dijk, G.; el Aidy, S. Gut Bacterial Tyrosine Decarboxylases Restrict Levels of Levodopa in the Treatment of Parkinson’s Disease. Nat. Commun. 2019, 10, 310. [Google Scholar] [CrossRef] [Green Version]
- Maini Rekdal, V.; Bess, E.N.; Bisanz, J.E.; Turnbaugh, P.J.; Balskus, E.P. Discovery and Inhibition of an Interspecies Gut Bacterial Pathway for Levodopa Metabolism. Science (1979) 2019, 364, eaau6323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jameson, K.G.; Hsiao, E.Y. A Novel Pathway for Microbial Metabolism of Levodopa. Nat. Med. 2019, 25, 1195–1197. [Google Scholar] [CrossRef]
- Hitchings, R.; Kelly, L. Drug Metabolism as a Community Effort. Cell Metab. 2019, 30, 235–237. [Google Scholar] [CrossRef]
- van Kessel, S.P.; de Jong, H.R.; Winkel, S.L.; van Leeuwen, S.S.; Nelemans, S.A.; Permentier, H.; Keshavarzian, A.; el Aidy, S. Gut Bacterial Deamination of Residual Levodopa Medication for Parkinson’s Disease. BMC Biol. 2020, 18, 137. [Google Scholar] [CrossRef] [PubMed]
- Weis, S.; Schwiertz, A.; Unger, M.M.; Becker, A.; Faßbender, K.; Ratering, S.; Kohl, M.; Schnell, S.; Schäfer, K.-H.; Egert, M. Effect of Parkinson’s Disease and Related Medications on the Composition of the Fecal Bacterial Microbiota. NPJ Park. Dis. 2019, 5, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melis, M.; Vascellari, S.; Santoru, M.L.; Oppo, V.; Fabbri, M.; Sarchioto, M.; Murgia, D.; Zibetti, M.; Lopiano, L.; Serra, A.; et al. Gut Microbiota and Metabolome Distinctive Features in Parkinson Disease: Focus on Levodopa and Levodopa-carbidopa Intrajejunal Gel. Eur. J. Neurol. 2021, 28, 1198–1209. [Google Scholar] [CrossRef] [PubMed]
- Palacios, N.; Hannoun, A.; Flahive, J.; Ward, D.; Goostrey, K.; Deb, A.; Smith, K.M. Effect of Levodopa Initiation on the Gut Microbiota in Parkinson’s Disease. Front. Neurol. 2021, 12, 574529. [Google Scholar] [CrossRef]
- van Kessel, S.P.; Auvinen, P.; Scheperjans, F.; el Aidy, S. Gut Bacterial Tyrosine Decarboxylase Associates with Clinical Variables in a Longitudinal Cohort Study of Parkinsons Disease. NPJ Park. Dis. 2021, 7, 115. [Google Scholar] [CrossRef]
- Bedarf, J.R.; Hildebrand, F.; Coelho, L.P.; Sunagawa, S.; Bahram, M.; Goeser, F.; Bork, P.; Wüllner, U. Functional Implications of Microbial and Viral Gut Metagenome Changes in Early Stage L-DOPA-Naïve Parkinson’s Disease Patients. Genome Med. 2017, 9, 39. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.-J.; Liang, C.-Y.; Yang, L.-Q.; Ren, S.-M.; Xia, Y.-M.; Cui, L.; Li, X.-F.; Gao, B.-L. Association of Parkinson’s Disease With Microbes and Microbiological Therapy. Front. Cell Infect. Microbiol. 2021, 11, 619354. [Google Scholar] [CrossRef]
- van Kessel, S.P.; Bullock, A.; van Dijk, G.; el Aidy, S. Parkinson’s Disease Medication Alters Small Intestinal Motility and Microbiota Composition in Healthy Rats. mSystems 2022, 7. [Google Scholar] [CrossRef] [PubMed]
- Gray, R.; Patel, S.; Ives, N.; Rick, C.; Woolley, R.; Muzerengi, S.; Gray, A.; Jenkinson, C.; McIntosh, E.; Wheatley, K.; et al. Long-Term Effectiveness of Adjuvant Treatment With Catechol-O-Methyltransferase or Monoamine Oxidase B Inhibitors Compared With Dopamine Agonists Among Patients With Parkinson Disease Uncontrolled by Levodopa Therapy. JAMA Neurol. 2022, 79, 131. [Google Scholar] [CrossRef] [PubMed]
- Ness, J.; Hoth, A.; Barnett, M.J.; Shorr, R.I.; Kaboli, P.J. Anticholinergic Medications in Community-Dwelling Older Veterans: Prevalence of Anticholinergic Symptoms, Symptom Burden, and Adverse Drug Events. Am. J. Geriatr. Pharm. 2006, 4, 42–51. [Google Scholar] [CrossRef]
- Kaakkola, S. Clinical Pharmacology, Therapeutic Use and Potential of COMT Inhibitors in Parkinson’s Disease. Drugs 2000, 59, 1233–1250. [Google Scholar] [CrossRef] [PubMed]
- Scheperjans, F.; Aho, V.; Pereira, P.A.B.; Koskinen, K.; Paulin, L.; Pekkonen, E.; Haapaniemi, E.; Kaakkola, S.; Eerola-Rautio, J.; Pohja, M.; et al. Gut Microbiota Are Related to Parkinson’s Disease and Clinical Phenotype. Mov. Disord. 2015, 30, 350–358. [Google Scholar] [CrossRef]
- Hill-Burns, E.M.; Debelius, J.W.; Morton, J.T.; Wissemann, W.T.; Lewis, M.R.; Wallen, Z.D.; Peddada, S.D.; Factor, S.A.; Molho, E.; Zabetian, C.P.; et al. Parkinson’s Disease and Parkinson’s Disease Medications Have Distinct Signatures of the Gut Microbiome. Mov. Disord. 2017, 32, 739–749. [Google Scholar] [CrossRef] [Green Version]
- Aho, V.T.E.; Pereira, P.A.B.; Voutilainen, S.; Paulin, L.; Pekkonen, E.; Auvinen, P.; Scheperjans, F. Gut Microbiota in Parkinson’s Disease: Temporal Stability and Relations to Disease Progression. EBioMedicine 2019, 44, 691–707. [Google Scholar] [CrossRef] [Green Version]
- Gordin, A.; Kaakkola, S.; Teräväinen, H. Clinical Advantages of COMT Inhibition with Entacapone ? A Review. J. Neural. Transm. 2004, 111, 1343–1363. [Google Scholar] [CrossRef]
- Grün, D.; Zimmer, V.C.; Kauffmann, J.; Spiegel, J.; Dillmann, U.; Schwiertz, A.; Faßbender, K.; Fousse, M.; Unger, M.M. Impact of Oral COMT-Inhibitors on Gut Microbiota and Short Chain Fatty Acids in Parkinson’s Disease. Park. Relat. Disord. 2020, 70, 20–22. [Google Scholar] [CrossRef] [Green Version]
- Fu, S.-C.; Lee, C.-H.; Hsieh, Y.-C.; Wu, P.-H.; Lin, S.-H.; Wang, H. A Pilot Study Exploring the Association of Entacapone, Gut Microbiota, and the Subsequent Side Effects in Patients With Parkinson’s Disease. Front. Cell Infect. Microbiol. 2022, 12, 837019. [Google Scholar] [CrossRef]
- Enright, E.F.; Gahan, C.G.M.; Joyce, S.A.; Griffin, B.T. The Impact of the Gut Microbiota on Drug Metabolism and Clinical Outcome. Yale J. Biol. Med. 2016, 89, 375–382. [Google Scholar] [PubMed]
- Sun, C.; Chen, L.; Shen, Z. Mechanisms of Gastrointestinal Microflora on Drug Metabolism in Clinical Practice. Saudi Pharm. J. 2019, 27, 1146–1156. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Shan, C.; Zhuang, S.; Zhuang, Q.; Ghosh, A.; Zhu, K.; Kong, X.; Wang, S.; Gong, Y.; Yang, Y.; et al. Gut Microbiota-Derived Propionate Mediates the Neuroprotective Effect of Osteocalcin in a Mouse Model of Parkinson’s Disease. Microbiome 2021, 9, 34. [Google Scholar] [CrossRef]
- Wang, Y.; Tong, Q.; Ma, S.-R.; Zhao, Z.-X.; Pan, L.-B.; Cong, L.; Han, P.; Peng, R.; Yu, H.; Lin, Y.; et al. Oral Berberine Improves Brain Dopa/Dopamine Levels to Ameliorate Parkinson’s Disease by Regulating Gut Microbiota. Signal Transduct Target Ther. 2021, 6, 77. [Google Scholar] [CrossRef] [PubMed]
- Rothstein, J.D.; Patel, S.; Regan, M.R.; Haenggeli, C.; Huang, Y.H.; Bergles, D.E.; Jin, L.; Dykes Hoberg, M.; Vidensky, S.; Chung, D.S.; et al. β-Lactam Antibiotics Offer Neuroprotection by Increasing Glutamate Transporter Expression. Nature 2005, 433, 73–77. [Google Scholar] [CrossRef]
- Lipski, J.; Wan, C.K.; Bai, J.Z.; Pi, R.; Li, D.; Donnelly, D. Neuroprotective Potential of Ceftriaxone in in Vitro Models of Stroke. Neuroscience 2007, 146, 617–629. [Google Scholar] [CrossRef] [PubMed]
- Burton, A. Antibiotics for Neurological Ailments? Lancet Neurol. 2005, 4, 142. [Google Scholar] [CrossRef]
- Pradhan, S.; Madke, B.; Kabra, P.; Singh, A. Anti-Inflammatory and Immunomodulatory Effects of Antibiotics and Their Use in Dermatology. Indian J. Dermatol. 2016, 61, 469. [Google Scholar] [CrossRef] [PubMed]
- Stoilova, T.; Colombo, L.; Forloni, G.; Tagliavini, F.; Salmona, M. A New Face for Old Antibiotics: Tetracyclines in Treatment of Amyloidoses. J. Med. Chem. 2013, 56, 5987–6006. [Google Scholar] [CrossRef]
- Van Vuuren, M.J.; Nell, T.A.; Carr, J.A.; Kell, D.B.; Pretorius, E. Iron Dysregulation and Inflammagens Related to Oral and Gut Health Are Central to the Development of Parkinson’s Disease. Biomolecules 2020, 11, 30. [Google Scholar] [CrossRef] [PubMed]
- Bortolanza, M.; Nascimento, G.C.; Socias, S.B.; Ploper, D.; Chehín, R.N.; Raisman-Vozari, R.; Del-Bel, E. Tetracycline Repurposing in Neurodegeneration: Focus on Parkinson’s Disease. J. Neural. Transm. 2018, 125, 1403–1415. [Google Scholar] [CrossRef] [PubMed]
- Yadav, N.; Thakur, A.K.; Shekhar, N. Ayushi Potential of Antibiotics for the Treatment and Management of Parkinson’s Disease: An Overview. Curr. Drug Res. Rev. 2021, 13, 166–171. [Google Scholar] [CrossRef]
- Hurkacz, M.; Dobrek, L.; Wiela-Hojeńska, A. Antibiotics and the Nervous System—Which Face of Antibiotic Therapy Is Real, Dr. Jekyll (Neurotoxicity) or Mr. Hyde (Neuroprotection)? Molecules 2021, 26, 7456. [Google Scholar] [CrossRef] [PubMed]
- Balducci, C.; Forloni, G. Doxycycline for Alzheimer’s Disease: Fighting β-Amyloid Oligomers and Neuroinflammation. Front. Pharmacol. 2019, 10, 738. [Google Scholar] [CrossRef] [Green Version]
- Lorente-Picón, M.; Laguna, A. New Avenues for Parkinson’s Disease Therapeutics: Disease-Modifying Strategies Based on the Gut Microbiota. Biomolecules 2021, 11, 433. [Google Scholar] [CrossRef]
- Pu, Y.; Chang, L.; Qu, Y.; Wang, S.; Zhang, K.; Hashimoto, K. Antibiotic-Induced Microbiome Depletion Protects against MPTP-Induced Dopaminergic Neurotoxicity in the Brain. Aging 2019, 11, 6915–6929. [Google Scholar] [CrossRef] [PubMed]
- Koutzoumis, D.N.; Vergara, M.; Pino, J.; Buddendorff, J.; Khoshbouei, H.; Mandel, R.J.; Torres, G.E. Alterations of the Gut Microbiota with Antibiotics Protects Dopamine Neuron Loss and Improve Motor Deficits in a Pharmacological Rodent Model of Parkinson’s Disease. Exp. Neurol. 2020, 325, 113159. [Google Scholar] [CrossRef]
- Cui, C.; Hong, H.; Shi, Y.; Zhou, Y.; Qiao, C.-M.; Zhao, W.-J.; Zhao, L.-P.; Wu, J.; Quan, W.; Niu, G.-Y.; et al. Vancomycin Pretreatment on MPTP-Induced Parkinson’s Disease Mice Exerts Neuroprotection by Suppressing Inflammation Both in Brain and Gut. J. Neuroimmune Pharmacol. 2022. [Google Scholar] [CrossRef]
- Baizabal-Carvallo, J.F.; Alonso-Juarez, M.; Fekete, R. Intestinal Decontamination Therapy for Dyskinesia and Motor Fluctuations in Parkinson’s Disease. Front. Neurol. 2021, 12, 729961. [Google Scholar] [CrossRef]
- Dănău, A.; Dumitrescu, L.; Lefter, A.; Tulbă, D.; Popescu, B.O. Small Intestinal Bacterial Overgrowth as Potential Therapeutic Target in Parkinson’s Disease. Int. J. Mol. Sci. 2021, 22, 11663. [Google Scholar] [CrossRef] [PubMed]
- Ramprasad, C.; Douglas, J.Y.; Moshiree, B. Parkinson’s Disease and Current Treatments for Its Gastrointestinal Neurogastromotility Effects. Curr. Treat Options Gastroenterol 2018, 16, 489–510. [Google Scholar] [CrossRef] [PubMed]
- Fasano, A.; Bove, F.; Gabrielli, M.; Petracca, M.; Zocco, M.A.; Ragazzoni, E.; Barbaro, F.; Piano, C.; Fortuna, S.; Tortora, A.; et al. The Role of Small Intestinal Bacterial Overgrowth in Parkinson’s Disease. Mov. Disord. 2013, 28, 1241–1249. [Google Scholar] [CrossRef]
- Pekary, A.E.; Sattin, A. Rifaximin Modulates TRH and TRH-like Peptide Expression throughout the Brain and Peripheral Tissues of Male Rats. BMC Neurosci. 2022, 23, 9. [Google Scholar] [CrossRef]
- DiBaise, J.K.; Crowell, M.D.; Driver-Dunckley, E.; Mehta, S.H.; Hoffman-Snyder, C.; Lin, T.; Adler, C.H. Weight Loss in Parkinson’s Disease: No Evidence for Role of Small Intestinal Bacterial Overgrowth. J. Park. Dis. 2018, 8, 571–581. [Google Scholar] [CrossRef]
- Yimer, E.M.; Hishe, H.Z.; Tuem, K.B. Repurposing of the β-Lactam Antibiotic, Ceftriaxone for Neurological Disorders: A Review. Front. Neurosci. 2019, 13, 236. [Google Scholar] [CrossRef] [Green Version]
- Tai, C.-H.; Bellesi, M.; Chen, A.-C.; Lin, C.-L.; Li, H.-H.; Lin, P.-J.; Liao, W.-C.; Hung, C.-S.; Schwarting, R.K.; Ho, Y.-J. A New Avenue for Treating Neuronal Diseases: Ceftriaxone, an Old Antibiotic Demonstrating Behavioral Neuronal Effects. Behavioural. Brain Res. 2019, 364, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Yamada, J.; Jinno, S. Alterations in Neuronal Survival and Glial Reactions after Axotomy by Ceftriaxone and Minocycline in the Mouse Hypoglossal Nucleus. Neurosci. Lett. 2011, 504, 295–300. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.; Qu, S. Ceftriaxone Protects Astrocytes from MPP+ via Suppression of NF-ΚB/JNK/c-Jun Signaling. Mol. Neurobiol. 2015, 52, 78–92. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; He, K.-J.; Hu, H.; Zhang, J.-B.; Dong, L.-G.; Li, D.; Chen, J.; Mao, C.-J.; Wang, F.; Liu, C.-F. Induction of Parkinsonian-Like Changes via Targeted Downregulation of Astrocytic Glutamate Transporter GLT-1 in the Striatum. J. Park. Dis. 2022, 12, 295–314. [Google Scholar] [CrossRef]
- Ruzza, P.; Siligardi, G.; Hussain, R.; Marchiani, A.; Islami, M.; Bubacco, L.; Delogu, G.; Fabbri, D.; Dettori, M.A.; Sechi, M.; et al. Ceftriaxone Blocks the Polymerization of α-Synuclein and Exerts Neuroprotective Effects in Vitro. ACS Chem. Neurosci. 2014, 5, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Smaga, I.; Fierro, D.; Mesa, J.; Filip, M.; Knackstedt, L.A. Molecular Changes Evoked by the Beta-Lactam Antibiotic Ceftriaxone across Rodent Models of Substance Use Disorder and Neurological Disease. Neurosci. Biobehav. Rev. 2020, 115, 116–130. [Google Scholar] [CrossRef]
- Zhou, X.; Lu, J.; Wei, K.; Wei, J.; Tian, P.; Yue, M.; Wang, Y.; Hong, D.; Li, F.; Wang, B.; et al. Neuroprotective Effect of Ceftriaxone on MPTP-Induced Parkinson’s Disease Mouse Model by Regulating Inflammation and Intestinal Microbiota. Oxid Med. Cell. Longev. 2021, 2021, 9424582. [Google Scholar] [CrossRef]
- Kaur, B.; Prakash, A. Ceftriaxone Attenuates Glutamate-Mediated Neuro-Inflammation and Restores BDNF in MPTP Model of Parkinson’s Disease in Rats. Pathophysiology 2017, 24, 71–79. [Google Scholar] [CrossRef]
- Bisht, R.; Kaur, B.; Gupta, H.; Prakash, A. Ceftriaxone Mediated Rescue of Nigral Oxidative Damage and Motor Deficits in MPTP Model of Parkinson’s Disease in Rats. Neurotoxicology 2014, 44, 71–79. [Google Scholar] [CrossRef]
- Brahmachari, S. Induction of Glial Fibrillary Acidic Protein Expression in Astrocytes by Nitric Oxide. J. Neurosci. 2006, 26, 4930–4939. [Google Scholar] [CrossRef] [Green Version]
- Jurga, A.M.; Paleczna, M.; Kuter, K.Z. Overview of General and Discriminating Markers of Differential Microglia Phenotypes. Front. Cell. Neurosci. 2020, 14, 198. [Google Scholar] [CrossRef]
- Ho, S.-C.; Hsu, C.-C.; Pawlak, C.R.; Tikhonova, M.A.; Lai, T.-J.; Amstislavskaya, T.G.; Ho, Y.-J. Effects of Ceftriaxone on the Behavioral and Neuronal Changes in an MPTP-Induced Parkinson’s Disease Rat Model. Behavioural. Brain Res. 2014, 268, 177–184. [Google Scholar] [CrossRef]
- Hsu, C.-Y.; Hung, C.-S.; Chang, H.-M.; Liao, W.-C.; Ho, S.-C.; Ho, Y.-J. Ceftriaxone Prevents and Reverses Behavioral and Neuronal Deficits in an MPTP-Induced Animal Model of Parkinson’s Disease Dementia. Neuropharmacology 2015, 91, 43–56. [Google Scholar] [CrossRef]
- Hsieh, M.-H.; Meng, W.-Y.; Liao, W.-C.; Weng, J.-C.; Li, H.-H.; Su, H.-L.; Lin, C.-L.; Hung, C.-S.; Ho, Y.-J. Ceftriaxone Reverses Deficits of Behavior and Neurogenesis in an MPTP-Induced Rat Model of Parkinson’s Disease Dementia. Brain Res. Bull 2017, 132, 129–138. [Google Scholar] [CrossRef]
- Chotibut, T.; Davis, R.W.; Arnold, J.C.; Frenchek, Z.; Gurwara, S.; Bondada, V.; Geddes, J.W.; Salvatore, M.F. Ceftriaxone Increases Glutamate Uptake and Reduces Striatal Tyrosine Hydroxylase Loss in 6-OHDA Parkinson’s Model. Mol. Neurobiol. 2014, 49, 1282–1292. [Google Scholar] [CrossRef] [Green Version]
- Weng, J.-C.; Tikhonova, M.A.; Chen, J.-H.; Shen, M.-S.; Meng, W.-Y.; Chang, Y.-T.; Chen, K.-H.; Liang, K.-C.; Hung, C.-S.; Amstislavskaya, T.G.; et al. Ceftriaxone Prevents the Neurodegeneration and Decreased Neurogenesis Seen in a Parkinson’s Disease Rat Model: An Immunohistochemical and MRI Study. Behav. Brain Res. 2016, 305, 126–139. [Google Scholar] [CrossRef]
- Leung, T.C.H.; Lui, C.N.P.; Chen, L.W.; Yung, W.H.; Chan, Y.S.; Yung, K.K.L. Ceftriaxone Ameliorates Motor Deficits and Protects Dopaminergic Neurons in 6-Hydroxydopamine-Lesioned Rats. ACS Chem. Neurosci. 2012, 3, 22–30. [Google Scholar] [CrossRef]
- Kelsey, J.E.; Neville, C. The Effects of the β-Lactam Antibiotic, Ceftriaxone, on Forepaw Stepping and l-DOPA-Induced Dyskinesia in a Rodent Model of Parkinson’s Disease. Psychopharmacology 2014, 231, 2405–2415. [Google Scholar] [CrossRef]
- Chotibut, T.; Meadows, S.; Kasanga, E.A.; McInnis, T.; Cantu, M.A.; Bishop, C.; Salvatore, M.F. Ceftriaxone Reduces L-Dopa-Induced Dyskinesia Severity in 6-Hydroxydopamine Parkinson’s Disease Model. Mov. Disord. 2017, 32, 1547–1556. [Google Scholar] [CrossRef]
- Karaman, I.; Kizilay-Ozfidan, G.; Karadag, C.H.; Ulugol, A. Lack of Effect of Ceftriaxone, a GLT-1 Transporter Activator, on Spatial Memory in Mice. Pharmacol. Biochem. Behav. 2013, 108, 61–65. [Google Scholar] [CrossRef]
- Huang, C.-K.; Chang, Y.-T.; Amstislavskaya, T.G.; Tikhonova, M.A.; Lin, C.-L.; Hung, C.-S.; Lai, T.-J.; Ho, Y.-J. Synergistic Effects of Ceftriaxone and Erythropoietin on Neuronal and Behavioral Deficits in an MPTP-Induced Animal Model of Parkinson’s Disease Dementia. Behav. Brain Res. 2015, 294, 198–207. [Google Scholar] [CrossRef]
- Garrido-Mesa, N.; Zarzuelo, A.; Gálvez, J. Minocycline: Far beyond an Antibiotic. Br. J. Pharmacol. 2013, 169, 337–352. [Google Scholar] [CrossRef] [Green Version]
- Cankaya, S.; Cankaya, B.; Kilic, U.; Kilic, E.; Yulug, B. The Therapeutic Role of Minocycline in Parkinson’s Disease. Drugs Context 2019, 8, 212553. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.; Xie, L.; Stevenson, F.F.; Melov, S.; di Monte, D.A.; Andersen, J.K. Nigrostriatal Dopaminergic Neurodegeneration in the Weaver Mouse Is Mediated via Neuroinflammation and Alleviated by Minocycline Administration. J. Neurosci. 2006, 26, 11644–11651. [Google Scholar] [CrossRef] [Green Version]
- Inamdar, A.A.; Chaudhuri, A.; O’Donnell, J. The Protective Effect of Minocycline in a Paraquat-Induced Parkinson’s Disease Model in Drosophila Is Modified in Altered Genetic Backgrounds. Park. Dis. 2012, 2012, 212553. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-S.; Suh, Y.-H. Minocycline and Neurodegenerative Diseases. Behav. Brain Res. 2009, 196, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.; Le, W.D.; Jankovic, J. Minocycline and Other Tetracycline Derivatives: A Neuroprotective Strategy in Parkinson’s Disease and Huntington’s Disease. Clin. Neuropharmacol 2003, 26, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Yong, V.W.; Wells, J.; Giuliani, F.; Casha, S.; Power, C.; Metz, L.M. The Promise of Minocycline in Neurology. Lancet Neurol. 2004, 3, 744–751. [Google Scholar] [CrossRef]
- Fan, L.; Wang, T.-L.; Xu, Y.C.; Ma, Y.H.; Ye, W.G. Minocycline May Be Useful to Prevent/Treat Postoperative Cognitive Decline in Elderly Patients. Med. Hypotheses 2011, 76, 733–736. [Google Scholar] [CrossRef]
- Blum, D.; Chtarto, A.; Tenenbaum, L.; Brotchi, J.; Levivier, M. Clinical Potential of Minocycline for Neurodegenerative Disorders. Neurobiol. Dis. 2004, 17, 359–366. [Google Scholar] [CrossRef]
- Thomas, M.; Le, W. Minocycline: Neuroprotective Mechanisms in Parkinsons Disease. Curr. Pharm. Des. 2004, 10, 679–686. [Google Scholar] [CrossRef]
- Zemke, D.; Majid, A. The Potential of Minocycline for Neuroprotection in Human Neurologic Disease. Clin. Neuropharmacol 2004, 27, 293–298. [Google Scholar] [CrossRef]
- Ruan, Z.; Zhang, D.; Huang, R.; Sun, W.; Hou, L.; Zhao, J.; Wang, Q. Microglial Activation Damages Dopaminergic Neurons through MMP-2/-9-Mediated Increase of Blood-Brain Barrier Permeability in a Parkinson’s Disease Mouse Model. Int. J. Mol. Sci. 2022, 23, 2793. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Q.; Yu, R.; Zhang, Q.; Zhang, Z.; Li, H.; Ren, C.; Yang, R.; Niu, H. Minocycline Inhibition of Microglial Rescues Nigrostriatal Dopaminergic Neurodegeneration Caused by Mutant Alpha-Synuclein Overexpression. Aging 2020, 12, 14232–14243. [Google Scholar] [CrossRef]
- Zhang, D.; Li, S.; Hou, L.; Jing, L.; Ruan, Z.; Peng, B.; Zhang, X.; Hong, J.-S.; Zhao, J.; Wang, Q. Microglial Activation Contributes to Cognitive Impairments in Rotenone-Induced Mouse Parkinson’s Disease Model. J. Neuroinflamm. 2021, 18, 4. [Google Scholar] [CrossRef]
- Tomás-Camardiel, M.; Rite, I.; Herrera, A.J.; de Pablos, R.M.; Cano, J.; Machado, A.; Venero, J.L. Minocycline Reduces the Lipopolysaccharide-Induced Inflammatory Reaction, Peroxynitrite-Mediated Nitration of Proteins, Disruption of the Blood–Brain Barrier, and Damage in the Nigral Dopaminergic System. Neurobiol. Dis. 2004, 16, 190–201. [Google Scholar] [CrossRef]
- Wu, D.C.; Jackson-Lewis, V.; Vila, M.; Tieu, K.; Teismann, P.; Vadseth, C.; Choi, D.-K.; Ischiropoulos, H.; Przedborski, S. Blockade of Microglial Activation Is Neuroprotective in the 1-Methyl-4-Phenyl-1,2,3,6-Tetrahydropyridine Mouse Model of Parkinson Disease. J. Neurosci. 2002, 22, 1763–1771. [Google Scholar] [CrossRef]
- Kumar, V.; Singh, B.K.; Chauhan, A.K.; Singh, D.; Patel, D.K.; Singh, C. Minocycline Rescues from Zinc-Induced Nigrostriatal Dopaminergic Neurodegeneration: Biochemical and Molecular Interventions. Mol. Neurobiol. 2016, 53, 2761–2777. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Ma, Z.; Lin, S.; Dodel, R.C.; Gao, F.; Bales, K.R.; Triarhou, L.C.; Chernet, E.; Perry, K.W.; Nelson, D.L.G.; et al. Minocycline Prevents Nigrostriatal Dopaminergic Neurodegeneration in the MPTP Model of Parkinson’s Disease. Proc. Natl. Acad. Sci. USA 2001, 98, 14669–14674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, B.-P.; Le, L.; Xu, L.-J.; Xiao, P.-G. Minocycline Inhibits ICAD Degradation and the NF-ΚB Activation Induced by 6-OHDA in PC12 Cells. Brain Res. 2014, 1586, 1–11. [Google Scholar] [CrossRef]
- Radad, K.; Moldzio, R.; Rausch, W.-D. Minocycline Protects Dopaminergic Neurons Against Long-Term Rotenone Toxicity. Can. J. Neurol. Sci. J. Can. Des Sci. Neurol. 2010, 37, 81–85. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Wei, X.; Xu, Y.; Yan, C.; Dodel, R.; Zhang, Y.; Liu, J.; Klaunig, J.E.; Farlow, M.; Du, Y. Minocycline Blocks 6-Hydroxydopamine-Induced Neurotoxicity and Free Radical Production in Rat Cerebellar Granule Neurons. Life Sci. 2003, 72, 1635–1641. [Google Scholar] [CrossRef]
- Cronin, A.; Grealy, M. Neuroprotective and Neuro-Restorative Effects of Minocycline and Rasagiline in a Zebrafish 6-Hydroxydopamine Model of Parkinson’s Disease. Neuroscience 2017, 367, 34–46. [Google Scholar] [CrossRef]
- Maneshian, M.; Nasirinezhad, F.; Mohammadi, F.; Behzadi, M.; Asadi-Shekaari, M.; Shabani, M. Minocycline Mitigation of Tremor Syndrome and Defect of Cognitive and Balance Induced by Harmaline. Basic Clin. Neurosci. J. 2021, 12, 255–268. [Google Scholar] [CrossRef]
- Quintero, E.M.; Willis, L.; Singleton, R.; Harris, N.; Huang, P.; Bhat, N.; Granholm, A.-C. Behavioral and Morphological Effects of Minocycline in the 6-Hydroxydopamine Rat Model of Parkinson’s Disease. Brain Res. 2006, 1093, 198–207. [Google Scholar] [CrossRef]
- Sun, C.; Wang, Y.; Mo, M.; Song, C.; Wang, X.; Chen, S.; Liu, Y. Minocycline Protects against Rotenone-Induced Neurotoxicity Correlating with Upregulation of Nurr1 in a Parkinson’s Disease Rat Model. Biomed Res. Int. 2019, 2019, 6843265. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.-L.; Lee, Y.-C.; Yang, Y.-C.; Kuo, T.-Y.; Huang, N.-K. Minocycline Prevents Paraquat-Induced Cell Death through Attenuating Endoplasmic Reticulum Stress and Mitochondrial Dysfunction. Toxicol. Lett. 2012, 209, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Gomez, F.J.; Galindo, M.F.; Gomez-Lazaro, M.; González-García, C.; Ceña, V.; Aguirre, N.; Jordán, J. Involvement of Mitochondrial Potential and Calcium Buffering Capacity in Minocycline Cytoprotective Actions. Neuroscience 2005, 133, 959–967. [Google Scholar] [CrossRef]
- Dixit, A.; Srivastava, G.; Verma, D.; Mishra, M.; Singh, P.K.; Prakash, O.; Singh, M.P. Minocycline, Levodopa and MnTMPyP Induced Changes in the Mitochondrial Proteome Profile of MPTP and Maneb and Paraquat Mice Models of Parkinson’s Disease. Biochim. Et Biophys. Acta (BBA)-Mol. Basis Dis. 2013, 1832, 1227–1240. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Sugama, S.; Chirichigno, J.W.; Gregorio, J.; Lorenzl, S.; Shin, D.H.; Browne, S.E.; Shimizu, Y.; Joh, T.H.; Beal, M.F.; et al. Minocycline Enhances MPTP Toxicity to Dopaminergic Neurons. J. Neurosci. Res. 2003, 74, 278–285. [Google Scholar] [CrossRef]
- Diguet, E.; Fernagut, P.-O.; Wei, X.; Du, Y.; Rouland, R.; Gross, C.; Bezard, E.; Tison, F. Deleterious Effects of Minocycline in Animal Models of Parkinson’s Disease and Huntington’s Disease. Eur. J. Neurosci. 2004, 19, 3266–3276. [Google Scholar] [CrossRef]
- NINDS NET-PD Investigators. A Pilot Clinical Trial of Creatine and Minocycline in Early Parkinson Disease. Clin. Neuropharmacol 2008, 31, 141–150. [Google Scholar] [CrossRef] [Green Version]
- NINDS NET-PD Investigators. A Randomized, Double-Blind, Futility Clinical Trial of Creatine and Minocycline in Early Parkinson Disease. Neurology 2006, 66, 664–671. [Google Scholar] [CrossRef]
- Rahmani, M.; Negro Álvarez, S.E.; Hernández, E.B. The Potential Use of Tetracyclines in Neurodegenerative Diseases and the Role of Nano-Based Drug Delivery Systems. Eur. J. Pharm. Sci. 2022, 175, 106237. [Google Scholar] [CrossRef]
- Santa-Cecília, F.V.; Socias, B.; Ouidja, M.O.; Sepulveda-Diaz, J.E.; Acuña, L.; Silva, R.L.; Michel, P.P.; Del-Bel, E.; Cunha, T.M.; Raisman-Vozari, R. Doxycycline Suppresses Microglial Activation by Inhibiting the P38 MAPK and NF-KB Signaling Pathways. Neurotox. Res. 2016, 29, 447–459. [Google Scholar] [CrossRef]
- Cho, Y.; Son, H.J.; Kim, E.-M.; Choi, J.H.; Kim, S.T.; Ji, I.J.; Choi, D.H.; Joh, T.H.; Kim, Y.S.; Hwang, O. Doxycycline Is Neuroprotective Against Nigral Dopaminergic Degeneration by a Dual Mechanism Involving MMP-3. Neurotox. Res. 2009, 16, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Meijide, A.; Parrales, V.; Vasili, E.; González-Lizárraga, F.; König, A.; Lázaro, D.F.; Lannuzel, A.; Haik, S.; del Bel, E.; Chehín, R.; et al. Doxycycline Inhibits α-Synuclein-Associated Pathologies in Vitro and in Vivo. Neurobiol. Dis. 2021, 151, 105256. [Google Scholar] [CrossRef]
- Do Amaral, L.; dos Santos, N.A.G.; Sisti, F.M.; del Bel, E.; Santos, A.C. dos The Antibiotic Doxycycline Mimics the NGF Signaling in PC12 Cells: A Relevant Mechanism for Neuroprotection. Chem. Biol. Interact 2021, 341, 109454. [Google Scholar] [CrossRef] [PubMed]
- González-Lizárraga, F.; Socías, S.B.; Ávila, C.L.; Torres-Bugeau, C.M.; Barbosa, L.R.S.; Binolfi, A.; Sepúlveda-Díaz, J.E.; Del-Bel, E.; Fernandez, C.O.; Papy-Garcia, D.; et al. Repurposing Doxycycline for Synucleinopathies: Remodelling of α-Synuclein Oligomers towards Non-Toxic Parallel Beta-Sheet Structured Species. Sci. Rep. 2017, 7, 41755. [Google Scholar] [CrossRef] [Green Version]
- Lazzarini, M.; Martin, S.; Mitkovski, M.; Vozari, R.R.; Stühmer, W.; Bel, E. del Doxycycline Restrains Glia and Confers Neuroprotection in a 6-OHDA Parkinson Model. Glia 2013, 61, 1084–1100. [Google Scholar] [CrossRef]
- Zhang, G.-B.; Feng, Y.-H.; Wang, P.-Q.; Song, J.-H.; Wang, P.; Wang, S.-A. A Study on the Protective Role of Doxycycline upon Dopaminergic Neuron of LPS-PD Rat Model Rat. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 3468–3474. [Google Scholar]
- Bortolanza, M.; Nascimento, G.C.; Raisman-Vozari, R.; Del-Bel, E. Doxycycline and Its Derivative, COL-3, Decrease Dyskinesia Induced by L-DOPA in Hemiparkinsonian Rats. Br. J. Pharmacol. 2021, 178, 2595–2616. [Google Scholar] [CrossRef]
- Bi, W.; Zhu, L.; Jing, X.; Liang, Y.; Tao, E. Rifampicin and Parkinson’s Disease. Neurol. Sci. 2013, 34, 137–141. [Google Scholar] [CrossRef]
- Yulug, B.; Hanoglu, L.; Kilic, E.; Schabitz, W.R. Rifampicin: An Antibiotic with Brain Protective Function. Brain Res. Bull 2014, 107, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Jing, X.; Chen, Y.; Liang, Y.; Lei, M.; Peng, S.; Zhou, T.; Zheng, D.; Zeng, Z.; Wu, X.; et al. Rifampicin Pre-Treatment Inhibits the Toxicity of Rotenone-Induced PC12 Cells by Enhancing Sumoylation Modification of α-Synuclein. Biochem. Biophys. Res. Commun. 2017, 485, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhu, M.; Rajamani, S.; Uversky, V.N.; Fink, A.L. Rifampicin Inhibits α-Synuclein Fibrillation and Disaggregates Fibrils. Chem. Biol. 2004, 11, 1513–1521. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Wei, C.; Xu, C.; Bennett, M.C.; Zhang, G.; Li, F.; Tao, E. Rifampicin Protects PC12 Cells against MPP+-Induced Apoptosis and Inhibits the Expression of an α-Synuclein Multimer. Brain Res. 2007, 1139, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Zhou, T.; Chen, Y.; Lin, D.; Jing, X.; Peng, S.; Zheng, D.; Zeng, Z.; Lei, M.; Wu, X.; et al. Rifampicin Inhibits Rotenone-Induced Microglial Inflammation via Enhancement of Autophagy. Neurotoxicology 2017, 63, 137–145. [Google Scholar] [CrossRef]
- Acuña, L.; Hamadat, S.; Corbalán, N.S.; González-Lizárraga, F.; Dos-Santos-Pereira, M.; Rocca, J.; Díaz, J.S.; Del-Bel, E.; Papy-García, D.; Chehín, R.N.; et al. Rifampicin and Its Derivative Rifampicin Quinone Reduce Microglial Inflammatory Responses and Neurodegeneration Induced In Vitro by α-Synuclein Fibrillary Aggregates. Cells 2019, 8, 776. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Zheng, D.; Peng, S.; Lin, D.; Jing, X.; Zeng, Z.; Chen, Y.; Huang, K.; Xie, Y.; Zhou, T.; et al. Rifampicin Attenuates Rotenone-Treated Microglia Inflammation via Improving Lysosomal Function. Toxicol. Vitr. 2020, 63, 104690. [Google Scholar] [CrossRef]
- Wu, X.; Liang, Y.; Jing, X.; Lin, D.; Chen, Y.; Zhou, T.; Peng, S.; Zheng, D.; Zeng, Z.; Lei, M.; et al. Rifampicin Prevents SH-SY5Y Cells from Rotenone-Induced Apoptosis via the PI3K/Akt/GSK-3β/CREB Signaling Pathway. Neurochem. Res. 2018, 43, 886–893. [Google Scholar] [CrossRef]
- Jing, X.; Shi, Q.; Bi, W.; Zeng, Z.; Liang, Y.; Wu, X.; Xiao, S.; Liu, J.; Yang, L.; Tao, E. Rifampicin Protects PC12 Cells from Rotenone-Induced Cytotoxicity by Activating GRP78 via PERK-EIF2α-ATF4 Pathway. PLoS ONE 2014, 9, e92110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oida, Y.; Kitaichi, K.; Nakayama, H.; Ito, Y.; Fujimoto, Y.; Shimazawa, M.; Nagai, H.; Hara, H. Rifampicin Attenuates the MPTP-Induced Neurotoxicity in Mouse Brain. Brain Res. 2006, 1082, 196–204. [Google Scholar] [CrossRef]
- Yurtsever, İ.; Üstündağ, Ü.V.; Ünal, İ.; Ateş, P.S.; Emekli-Alturfan, E. Rifampicin Decreases Neuroinflammation to Maintain Mitochondrial Function and Calcium Homeostasis in Rotenone-Treated Zebrafish. Drug Chem. Toxicol. 2022, 45, 1544–1551. [Google Scholar] [CrossRef]
- Modi, S.R.; Collins, J.J.; Relman, D.A. Antibiotics and the Gut Microbiota. J. Clin. Investig. 2014, 124, 4212–4218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markulin, I.; Matasin, M.; Turk, V.E.; Salković-Petrisic, M. Challenges of Repurposing Tetracyclines for the Treatment of Alzheimer’s and Parkinson’s Disease. J. Neural. Transm. 2022, 129, 773–804. [Google Scholar] [CrossRef] [PubMed]
- Avagliano, C.; Coretti, L.; Lama, A.; Pirozzi, C.; de Caro, C.; de Biase, D.; Turco, L.; Mollica, M.P.; Paciello, O.; Calignano, A.; et al. Dual-Hit Model of Parkinson’s Disease: Impact of Dysbiosis on 6-Hydroxydopamine-Insulted Mice—Neuroprotective and Anti-Inflammatory Effects of Butyrate. Int. J. Mol. Sci. 2022, 23, 6367. [Google Scholar] [CrossRef] [PubMed]
- Ternák, G.; Kuti, D.; Kovács, K.J. Dysbiosis in Parkinson’s Disease Might Be Triggered by Certain Antibiotics. Med. Hypotheses 2020, 137, 109564. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Lau, C.Y.; Ma, F.; Zheng, C. Genome-Wide Screen Identifies Curli Amyloid Fibril as a Bacterial Component Promoting Host Neurodegeneration. Proc. Natl. Acad. Sci. USA 2021, 118, e2106504118. [Google Scholar] [CrossRef]
- Roe, K. An Alternative Explanation for Alzheimer’s Disease and Parkinson’s Disease Initiation from Specific Antibiotics, Gut Microbiota Dysbiosis and Neurotoxins. Neurochem. Res. 2022, 47, 517–530. [Google Scholar] [CrossRef]
- Mertsalmi, T.H.; Pekkonen, E.; Scheperjans, F. Antibiotic Exposure and Risk of Parkinson’s Disease in Finland: A Nationwide Case-Control Study. Mov. Disord. 2020, 35, 431–442. [Google Scholar] [CrossRef]
- Palacios, N.; O’Reilly, É.J.; Schwarzschild, M.A.; Ascherio, A. Long-Term Use of Antibiotics and Risk of Parkinson’s Disease in the Nurses’ Health Study. Park. Dis. 2020, 2020, 4038375. [Google Scholar] [CrossRef] [Green Version]
- Morelli, L.; Capurso, L. FAO/WHO Guidelines on Probiotics. J. Clin. Gastroenterol. 2012, 46, S1–S2. [Google Scholar] [CrossRef]
- Alfonsetti, M.; Castelli, V.; d’Angelo, M. Are We What We Eat? Impact of Diet on the Gut–Brain Axis in Parkinson’s Disease. Nutrients 2022, 14, 380. [Google Scholar] [CrossRef]
- Cristofori, F.; Dargenio, V.N.; Dargenio, C.; Miniello, V.L.; Barone, M.; Francavilla, R. Anti-Inflammatory and Immunomodulatory Effects of Probiotics in Gut Inflammation: A Door to the Body. Front. Immunol. 2021, 12, 578386. [Google Scholar] [CrossRef]
- Gazerani, P. Probiotics for Parkinson’s Disease. Int. J. Mol. Sci. 2019, 20, 4121. [Google Scholar] [CrossRef] [Green Version]
- Gupta, V.; Garg, R. PROBIOTICS. Indian J. Med. Microbiol. 2009, 27, 202–209. [Google Scholar] [CrossRef]
- Reid, G.; Younes, J.A.; van der Mei, H.C.; Gloor, G.B.; Knight, R.; Busscher, H.J. Microbiota Restoration: Natural and Supplemented Recovery of Human Microbial Communities. Nat. Rev. Microbiol. 2011, 9, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Nandwana, V.; Nandwana, N.K.; Das, Y.; Saito, M.; Panda, T.; Das, S.; Almaguel, F.; Hosmane, N.S.; Das, B.C. The Role of Microbiome in Brain Development and Neurodegenerative Diseases. Molecules 2022, 27, 3402. [Google Scholar] [CrossRef] [PubMed]
- Peterson, C.T. Dysfunction of the Microbiota-Gut-Brain Axis in Neurodegenerative Disease: The Promise of Therapeutic Modulation With Prebiotics, Medicinal Herbs, Probiotics, and Synbiotics. J. Evid Based Integr. Med. 2020, 25, 2515690X20957225. [Google Scholar] [CrossRef]
- Becker, A.; Pierre Schmartz, G.; Gröger, L.; Grammes, N.; Galata, V.; Philippeit, H.; Weiland, J.; Ludwig, N.; Meese, E.; Tierling, S.; et al. Effects of Resistant Starch on Symptoms, Fecal Markers and Gut Microbiota in Parkinson’s Disease–the RESISTA-PD Trial. Genom. Proteom. Bioinform. 2021. [Google Scholar] [CrossRef]
- Jiang, J.; Chu, C.; Wu, C.; Wang, C.; Zhang, C.; Li, T.; Zhai, Q.; Yu, L.; Tian, F.; Chen, W. Efficacy of Probiotics in Multiple Sclerosis: A Systematic Review of Preclinical Trials and Meta-Analysis of Randomized Controlled Trials. Food Funct. 2021, 12, 2354–2377. [Google Scholar] [CrossRef]
- Mirzaei, H.; Sedighi, S.; Kouchaki, E.; Barati, E.; Dadgostar, E.; Aschner, M.; Tamtaji, O.R. Probiotics and the Treatment of Parkinson’s Disease: An Update. Cell. Mol. Neurobiol. 2021. [Google Scholar] [CrossRef]
- Socała, K.; Doboszewska, U.; Szopa, A.; Serefko, A.; Włodarczyk, M.; Zielińska, A.; Poleszak, E.; Fichna, J.; Wlaź, P. The Role of Microbiota-Gut-Brain Axis in Neuropsychiatric and Neurological Disorders. Pharmacol. Res. 2021, 172, 105840. [Google Scholar] [CrossRef]
- Xiang, S.; Ji, J.-L.; Li, S.; Cao, X.-P.; Xu, W.; Tan, L.; Tan, C.-C. Efficacy and Safety of Probiotics for the Treatment of Alzheimer’s Disease, Mild Cognitive Impairment, and Parkinson’s Disease: A Systematic Review and Meta-Analysis. Front. Aging Neurosci. 2022, 14. [Google Scholar] [CrossRef]
- Hsiao, E.Y.; McBride, S.W.; Hsien, S.; Sharon, G.; Hyde, E.R.; McCue, T.; Codelli, J.A.; Chow, J.; Reisman, S.E.; Petrosino, J.F.; et al. Microbiota Modulate Behavioral and Physiological Abnormalities Associated with Neurodevelopmental Disorders. Cell 2013, 155, 1451–1463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonfili, L.; Cecarini, V.; Berardi, S.; Scarpona, S.; Suchodolski, J.S.; Nasuti, C.; Fiorini, D.; Boarelli, M.C.; Rossi, G.; Eleuteri, A.M. Microbiota Modulation Counteracts Alzheimer’s Disease Progression Influencing Neuronal Proteolysis and Gut Hormones Plasma Levels. Sci. Rep. 2017, 7, 2426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akbari, E.; Asemi, Z.; Daneshvar Kakhaki, R.; Bahmani, F.; Kouchaki, E.; Tamtaji, O.R.; Hamidi, G.A.; Salami, M. Effect of Probiotic Supplementation on Cognitive Function and Metabolic Status in Alzheimer’s Disease: A Randomized, Double-Blind and Controlled Trial. Front. Aging Neurosci. 2016, 8, 256. [Google Scholar] [CrossRef]
- Abildgaard, A.; Elfving, B.; Hokland, M.; Wegener, G.; Lund, S. Probiotic Treatment Reduces Depressive-like Behaviour in Rats Independently of Diet. Psychoneuroendocrinology 2017, 79, 40–48. [Google Scholar] [CrossRef]
- Srivastav, S.; Neupane, S.; Bhurtel, S.; Katila, N.; Maharjan, S.; Choi, H.; Hong, J.T.; Choi, D.-Y. Probiotics Mixture Increases Butyrate, and Subsequently Rescues the Nigral Dopaminergic Neurons from MPTP and Rotenone-Induced Neurotoxicity. J. Nutr. Biochem. 2019, 69, 73–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, C.J.K.; Milev, R. The Effects of Probiotics on Depressive Symptoms in Humans: A Systematic Review. Ann. Gen. Psychiatry 2017, 16. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Lee, I.-S.; Braun, C.; Enck, P. Effect of Probiotics on Central Nervous System Functions in Animals and Humans: A Systematic Review. J. Neurogastroenterol. Motil. 2016, 22, 589–605. [Google Scholar] [CrossRef] [Green Version]
- Magistrelli, L.; Amoruso, A.; Mogna, L.; Graziano, T.; Cantello, R.; Pane, M.; Comi, C. Probiotics May Have Beneficial Effects in Parkinson’s Disease: In Vitro Evidence. Front. Immunol. 2019, 10, 969. [Google Scholar] [CrossRef] [Green Version]
- Ghyselinck, J.; Verstrepen, L.; Moens, F.; van den Abbeele, P.; Bruggeman, A.; Said, J.; Smith, B.; Barker, L.A.; Jordan, C.; Leta, V.; et al. Influence of Probiotic Bacteria on Gut Microbiota Composition and Gut Wall Function in an In-Vitro Model in Patients with Parkinson’s Disease. Int. J. Pharm. X 2021, 3, 100087. [Google Scholar] [CrossRef] [PubMed]
- Cheon, M.-J.; Lee, N.-K.; Paik, H.-D. Neuroprotective Effects of Heat-Killed Lactobacillus Plantarum 200655 Isolated from Kimchi Against Oxidative Stress. Probiotics Antimicrob Proteins 2021, 13, 788–795. [Google Scholar] [CrossRef]
- Castelli, V.; d’Angelo, M.; Lombardi, F.; Alfonsetti, M.; Antonosante, A.; Catanesi, M.; Benedetti, E.; Palumbo, P.; Cifone, M.G.; Giordano, A.; et al. Effects of the Probiotic Formulation SLAB51 in in Vitro and in Vivo Parkinson’s Disease Models. Aging 2020, 12, 4641–4659. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.-F.; Cheng, Y.-F.; You, S.-T.; Kuo, W.-C.; Huang, C.-W.; Chiou, J.-J.; Hsu, C.-C.; Hsieh-Li, H.-M.; Wang, S.; Tsai, Y.-C. Lactobacillus Plantarum PS128 Alleviates Neurodegenerative Progression in 1-Methyl-4-Phenyl-1,2,3,6-Tetrahydropyridine-Induced Mouse Models of Parkinson’s Disease. Brain Behav. Immun. 2020, 90, 26–46. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, T.-H.; Kuo, C.-W.; Hsieh, K.-H.; Shieh, M.-J.; Peng, C.-W.; Chen, Y.-C.; Chang, Y.-L.; Huang, Y.-Z.; Chen, C.-C.; Chang, P.-K.; et al. Probiotics Alleviate the Progressive Deterioration of Motor Functions in a Mouse Model of Parkinson’s Disease. Brain Sci. 2020, 10, 206. [Google Scholar] [CrossRef]
- Sun, J.; Li, H.; Jin, Y.; Yu, J.; Mao, S.; Su, K.-P.; Ling, Z.; Liu, J. Probiotic Clostridium Butyricum Ameliorated Motor Deficits in a Mouse Model of Parkinson’s Disease via Gut Microbiota-GLP-1 Pathway. Brain Behav. Immun. 2021, 91, 703–715. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhao, Z.; Zhao, L.; Zhao, Y.; Yang, G.; Wang, C.; Gao, L.; Niu, C.; Li, S. Lactobacillus Plantarum DP189 Reduces α-SYN Aggravation in MPTP-Induced Parkinson’s Disease Mice via Regulating Oxidative Damage, Inflammation, and Gut Microbiota Disorder. J. Agric. Food Chem. 2022, 70, 1163–1173. [Google Scholar] [CrossRef] [PubMed]
- Cassani, E.; Privitera, G.; Pezzoli, G.; Pusani, C.; Madio, C.; Iorio, L.; Barichella, M. Use of Probiotics for the Treatment of Constipation in Parkinson’s Disease Patients. Minerva Gastroenterol. Dietol. 2011, 57, 117–121. [Google Scholar] [PubMed]
- Georgescu, D.; Ancusa, O.; Georgescu, L.; Ionita, I.; Reisz, D. Nonmotor Gastrointestinal Disorders in Older Patients with Parkinson’s Disease: Is There Hope? Clin. Interv. Aging 2016, 11, 1601–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borzabadi, S.; Oryan, S.; Eidi, A.; Aghadavod, E.; Daneshvar Kakhaki, R.; Tamtaji, O.R.; Taghizadeh, M.; Asemi, Z. The Effects of Probiotic Supplementation on Gene Expression Related to Inflammation, Insulin and Lipid in Patients with Parkinson’s Disease: A Randomized, Double-Blind, PlaceboControlled Trial. Arch. Iran Med. 2018, 21, 289–295. [Google Scholar]
- Tamtaji, O.R.; Taghizadeh, M.; Daneshvar Kakhaki, R.; Kouchaki, E.; Bahmani, F.; Borzabadi, S.; Oryan, S.; Mafi, A.; Asemi, Z. Clinical and Metabolic Response to Probiotic Administration in People with Parkinson’s Disease: A Randomized, Double-Blind, Placebo-Controlled Trial. Clin. Nutr. 2019, 38, 1031–1035. [Google Scholar] [CrossRef]
- Ibrahim, A.; Ali, R.A.R.; Manaf, M.R.A.; Ahmad, N.; Tajurruddin, F.W.; Qin, W.Z.; Desa, S.H.M.; Ibrahim, N.M. Multi-Strain Probiotics (Hexbio) Containing MCP BCMC Strains Improved Constipation and Gut Motility in Parkinson’s Disease: A Randomised Controlled Trial. PLoS ONE 2020, 15, e0244680. [Google Scholar] [CrossRef]
- Tan, A.H.; Lim, S.-Y.; Chong, K.K.; Azhan A Manap, M.A.; Hor, J.W.; Lim, J.L.; Low, S.C.; Chong, C.W.; Mahadeva, S.; Lang, A.E. Probiotics for Constipation in Parkinson’s Disease: A Randomized Placebo-Controlled Study. Neurology 2020, 96, e772–e782. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Zhao, F.; Liu, Y.; Ma, T.; Jin, H.; Quan, K.; Leng, B.; Zhao, J.; Yuan, X.; Li, Z.; et al. Probiotics Synergized with Conventional Regimen in Managing Parkinson’s Disease. NPJ Park. Dis. 2022, 8. [Google Scholar] [CrossRef]
- Surwase, S.N.; Jadhav, J.P. Bioconversion of L-Tyrosine to l-DOPA by a Novel Bacterium Bacillus sp. JPJ. Amino Acids 2011, 41, 495–506. [Google Scholar] [CrossRef]
- Goya, M.E.; Xue, F.; Sampedro-Torres-Quevedo, C.; Arnaouteli, S.; Riquelme-Dominguez, L.; Romanowski, A.; Brydon, J.; Ball, K.L.; Stanley-Wall, N.R.; Doitsidou, M. Probiotic Bacillus Subtilis Protects against α-Synuclein Aggregation in C. Elegans. Cell Rep. 2020, 30, 367–380.e7. [Google Scholar] [CrossRef] [PubMed]
- Perez Visñuk, D.; Savoy de Giori, G.; LeBlanc, J.G.; de Moreno de LeBlanc, A. Neuroprotective Effects Associated with Immune Modulation by Selected Lactic Acid Bacteria in a Parkinson’s Disease Model. Nutrition 2020, 79–80, 110995. [Google Scholar] [CrossRef] [PubMed]
- Tsao, S.-P.; Nurrahma, B.A.; Kumar, R.; Wu, C.-H.; Yeh, T.-H.; Chiu, C.-C.; Lee, Y.-P.; Liao, Y.-C.; Huang, C.-H.; Yeh, Y.-T.; et al. Probiotic Enhancement of Antioxidant Capacity and Alterations of Gut Microbiota Composition in 6-Hydroxydopamin-Induced Parkinson’s Disease Rats. Antioxidants 2021, 10, 1823. [Google Scholar] [CrossRef] [PubMed]
- Alipour Nosrani, E.; Tamtaji, O.R.; Alibolandi, Z.; Sarkar, P.; Ghazanfari, M.; Azami Tameh, A.; Taghizadeh, M.; Banikazemi, Z.; Hadavi, R.; Naderi Taheri, M. Neuroprotective Effects of Probiotics Bacteria on Animal Model of Parkinson’s Disease Induced by 6-Hydroxydopamine: A Behavioral, Biochemical, and Histological Study. J. Immunoass. Immunochem. 2021, 42, 106–120. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Prasad, A.A. Probiotics Treatment Improves Hippocampal Dependent Cognition in a Rodent Model of Parkinson’s Disease. Microorganisms 2020, 8, 1661. [Google Scholar] [CrossRef]
- Fang, X.; Tian, P.; Zhao, X.; Jiang, C.; Chen, T. Neuroprotective Effects of an Engineered Commensal Bacterium in the 1-methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine Parkinson Disease Mouse Model via Producing Glucagon-like Peptide-1. J. Neurochem. 2019, 150, 441–452. [Google Scholar] [CrossRef]
- Fang, X.; Zhou, X.; Miao, Y.; Han, Y.; Wei, J.; Chen, T. Therapeutic Effect of GLP-1 Engineered Strain on Mice Model of Alzheimer’s Disease and Parkinson’s Disease. AMB Express 2020, 10, 13–80. [Google Scholar] [CrossRef]
- Tan, A.H.; Hor, J.W.; Chong, C.W.; Lim, S. Probiotics for Parkinson’s Disease: Current Evidence and Future Directions. JGH Open 2021, 5, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Xie, M.; Xue, J.; Xiang, L.; Li, Y.; Xiao, J.; Xiao, G.; Wang, H. EGCG Ameliorates Neuronal and Behavioral Defects by Remodeling Gut Microbiota and TotM Expression in Drosophila Models of Parkinson’s Disease. FASEB J. 2020, 34, 5931–5950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dwyer, Z.; Chaiquin, M.; Landrigan, J.; Ayoub, K.; Shail, P.; Rocha, J.; Childers, C.L.; Storey, K.B.; Philpott, D.J.; Sun, H.; et al. The Impact of Dextran Sodium Sulphate and Probiotic Pre-Treatment in a Murine Model of Parkinson’s Disease. J. Neuroinflamm. 2021, 18, 20. [Google Scholar] [CrossRef]
- Lu, C.-S.; Chang, H.-C.; Weng, Y.-H.; Chen, C.-C.; Kuo, Y.-S.; Tsai, Y.-C. The Add-On Effect of Lactobacillus Plantarum PS128 in Patients With Parkinson’s Disease: A Pilot Study. Front. Nutr. 2021, 8, 5931–5950. [Google Scholar] [CrossRef] [PubMed]
- Yahfoufi, N.; Matar, C.; Ismail, N. Adolescence and Aging: Impact of Adolescence Inflammatory Stress and Microbiota Alterations on Brain Development, Aging, and Neurodegeneration. J. Gerontol. Ser. A 2020, 75, 1251–1257. [Google Scholar] [CrossRef] [Green Version]
- Enck, K.; Banks, S.; Yadav, H.; Welker, M.E.; Opara, E.C. Development of a Novel Oral Delivery Vehicle for Probiotics. Curr. Pharm. Des. 2020, 26, 3134–3140. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Sun, T.; Cui, M.; Ma, N.; Yang, C.; Liu, J.; Pang, G.; Liu, B.; Li, L.; Zhang, X.; et al. Light-Sensitive Lactococcus Lactis for Microbe–Gut–Brain Axis Regulating via Upconversion Optogenetic Micro-Nano System. ACS Nano 2022, 16, 6049–6063. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert Consensus Document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Pandey, K.R.; Naik, S.R.; Vakil, B.V. Probiotics, Prebiotics and Synbiotics- a Review. J. Food Sci. Technol. 2015, 52, 7577–7587. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Luo, Y.; Ray Chaudhuri, K.; Reynolds, R.; Tan, E.-K.; Pettersson, S. The Role of Gut Dysbiosis in Parkinson’s Disease: Mechanistic Insights and Therapeutic Options. Brain 2021, 144, 2571–2593. [Google Scholar] [CrossRef]
- Wiciński, M.; Gębalski, J.; Mazurek, E.; Podhorecka, M.; Śniegocki, M.; Szychta, P.; Sawicka, E.; Malinowski, B. The Influence of Polyphenol Compounds on Human Gastrointestinal Tract Microbiota. Nutrients 2020, 12, 350. [Google Scholar] [CrossRef] [Green Version]
- de Vrese, M.; Schrezenmeir, J. Probiotics, Prebiotics, and Synbiotics. Adv. Biochem. Eng. Biotechnol. 2008, 111, 1–66. [Google Scholar] [CrossRef] [PubMed]
- Manzoor, S.; Wani, S.M.; Ahmad Mir, S.; Rizwan, D. Role of Probiotics and Prebiotics in Mitigation of Different Diseases. Nutrition 2022, 96, 111602. [Google Scholar] [CrossRef] [PubMed]
- Raval, U.; Harary, J.M.; Zeng, E.; Pasinetti, G.M. The Dichotomous Role of the Gut Microbiome in Exacerbating and Ameliorating Neurodegenerative Disorders. Expert Rev. Neurother. 2020, 20, 673–686. [Google Scholar] [CrossRef] [PubMed]
- Cantu-Jungles, T.M.; Rasmussen, H.E.; Hamaker, B.R. Potential of Prebiotic Butyrogenic Fibers in Parkinson’s Disease. Front. Neurol. 2019, 10, 663. [Google Scholar] [CrossRef] [Green Version]
- Guo, T.; Chen, L. Gut Microbiota and Inflammation in Parkinson’s Disease: Pathogenetic and Therapeutic Insights. Eur. J. Inflamm. 2022, 20. [Google Scholar] [CrossRef]
- Yang, H.; Liu, Y.; Cai, R.; Li, Y.; Gu, B. A Narrative Review of Relationship between Gut Microbiota and Neuropsychiatric Disorders: Mechanisms and Clinical Application of Probiotics and Prebiotics. Ann. Palliat. Med. 2021, 10, 2304–2313. [Google Scholar] [CrossRef]
- Lee, Y.-S.; Lai, D.-M.; Huang, H.-J.; Lee-Chen, G.-J.; Chang, C.-H.; Hsieh-Li, H.M.; Lee, G.-C. Prebiotic Lactulose Ameliorates the Cognitive Deficit in Alzheimer’s Disease Mouse Model through Macroautophagy and Chaperone-Mediated Autophagy Pathways. J. Agric. Food Chem. 2021, 69, 2422–2437. [Google Scholar] [CrossRef]
- Grimaldi, R.; Gibson, G.R.; Vulevic, J.; Giallourou, N.; Castro-Mejía, J.L.; Hansen, L.H.; Leigh Gibson, E.; Nielsen, D.S.; Costabile, A. A Prebiotic Intervention Study in Children with Autism Spectrum Disorders (ASDs). Microbiome 2018, 6, 133. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Yang, X.; Yang, J.; Lai, G.; Yong, T.; Tang, X.; Shuai, O.; Zhou, G.; Xie, Y.; Wu, Q. Prebiotic Effect of Fructooligosaccharides from Morinda Officinalis on Alzheimer’s Disease in Rodent Models by Targeting the Microbiota-Gut-Brain Axis. Front. Aging Neurosci. 2017, 9, 403. [Google Scholar] [CrossRef] [Green Version]
- Xin, Y.; Diling, C.; Jian, Y.; Ting, L.; Guoyan, H.; Hualun, L.; Xiaocui, T.; Guoxiao, L.; Ou, S.; Chaoqun, Z.; et al. Effects of Oligosaccharides From Morinda Officinalis on Gut Microbiota and Metabolome of APP/PS1 Transgenic Mice. Front. Neurol. 2018, 9, 412. [Google Scholar] [CrossRef] [PubMed]
- Perez-Pardo, P.; de Jong, E.M.; Broersen, L.M.; van Wijk, N.; Attali, A.; Garssen, J.; Kraneveld, A.D. Promising Effects of Neurorestorative Diets on Motor, Cognitive, and Gastrointestinal Dysfunction after Symptom Development in a Mouse Model of Parkinson’s Disease. Front. Aging Neurosci. 2017, 9, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Du, Z.R.; Wang, X.; Sun, X.R.; Zhao, Q.; Zhao, F.; Wong, W.T.; Wong, K.H.; Dong, X.-L. Polymannuronic Acid Prebiotic plus Lacticaseibacillus Rhamnosus GG Probiotic as a Novel Synbiotic Promoted Their Separate Neuroprotection against Parkinson’s Disease. Food Res. Int. 2022, 155, 111067. [Google Scholar] [CrossRef] [PubMed]
- Astarloa, R.; Mena, M.A.; Sánchez, V.; de la Vega, L.; de Yébenes, J.G. Clinical and Pharmacokinetic Effects of a Diet Rich in Insoluble Fiber on Parkinson Disease. Clin. Neuropharmacol. 1992, 15, 375–380. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, W.; Pfeiffer, R.F.; Park, F.; Lof, J.; Quigley, E.M.M. Constipation in Parkinson’s Disease: Objective Assessment and Response to Psyllium. Mov. Disord. 1997, 12, 946–951. [Google Scholar] [CrossRef] [PubMed]
- Nurrahma, B.A.; Tsao, S.-P.; Wu, C.-H.; Yeh, T.-H.; Hsieh, P.-S.; Panunggal, B.; Huang, H.-Y. Probiotic Supplementation Facilitates Recovery of 6-OHDA-Induced Motor Deficit via Improving Mitochondrial Function and Energy Metabolism. Front. Aging Neurosci. 2021, 13, 668775. [Google Scholar] [CrossRef]
- Savignac, H.M.; Corona, G.; Mills, H.; Chen, L.; Spencer, J.P.E.; Tzortzis, G.; Burnet, P.W.J. Prebiotic Feeding Elevates Central Brain Derived Neurotrophic Factor, N-Methyl-d-Aspartate Receptor Subunits and d-Serine. Neurochem. Int. 2013, 63, 756–764. [Google Scholar] [CrossRef] [Green Version]
- Bathina, S.; Das, U.N. Brain-Derived Neurotrophic Factor and Its Clinical Implications. Arch. Med. Sci. 2015, 6, 1164–1178. [Google Scholar] [CrossRef]
- St. Laurent, R.; O’Brien, L.M.; Ahmad, S.T. Sodium Butyrate Improves Locomotor Impairment and Early Mortality in a Rotenone-Induced Drosophila Model of Parkinson’s Disease. Neuroscience 2013, 246, 382–390. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Bercury, K.; Cummiskey, J.; Luong, N.; Lebin, J.; Freed, C.R. Phenylbutyrate Up-Regulates the DJ-1 Protein and Protects Neurons in Cell Culture and in Animal Models of Parkinson Disease. J. Biol. Chem. 2011, 286, 14941–14951. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, V.E.; Herrera, P.F.; Laura, R. Effect of Nutrition on Neurodegenerative Diseases. A Systematic Review. Nutr. Neurosci. 2021, 24, 810–834. [Google Scholar] [CrossRef]
- Solfrizzi, V.; Custodero, C.; Lozupone, M.; Imbimbo, B.P.; Valiani, V.; Agosti, P.; Schilardi, A.; D’Introno, A.; la Montagna, M.; Calvani, M.; et al. Relationships of Dietary Patterns, Foods, and Micro- and Macronutrients with Alzheimer’s Disease and Late-Life Cognitive Disorders: A Systematic Review. J. Alzheimer’s Dis. 2017, 59, 815–849. [Google Scholar] [CrossRef] [Green Version]
- Metcalfe-Roach, A.; Yu, A.C.; Golz, E.; Cirstea, M.; Sundvick, K.; Kliger, D.; Foulger, L.H.; Mackenzie, M.; Finlay, B.B.; Appel-Cresswell, S. MIND and Mediterranean Diets Associated with Later Onset of Parkinson’s Disease. Mov. Disord. 2021, 36, 977–984. [Google Scholar] [CrossRef] [PubMed]
- Paknahad, Z.; Sheklabadi, E.; Derakhshan, Y.; Bagherniya, M.; Chitsaz, A. The Effect of the Mediterranean Diet on Cognitive Function in Patients with Parkinson’s Disease: A Randomized Clinical Controlled Trial. Complement. Ther. Med. 2020, 50, 102366. [Google Scholar] [CrossRef] [PubMed]
- Rusch, C.; Beke, M.; Tucciarone, L.; Nieves, C.; Ukhanova, M.; Tagliamonte, M.S.; Mai, V.; Suh, J.H.; Wang, Y.; Chiu, S.; et al. Mediterranean Diet Adherence in People With Parkinson’s Disease Reduces Constipation Symptoms and Changes Fecal Microbiota After a 5-Week Single-Arm Pilot Study. Front. Neurol. 2021, 12, 794640. [Google Scholar] [CrossRef]
- Cassani, E.; Barichella, M.; Ferri, V.; Pinelli, G.; Iorio, L.; Bolliri, C.; Caronni, S.; Faierman, S.A.; Mottolese, A.; Pusani, C.; et al. Dietary Habits in Parkinson’s Disease: Adherence to Mediterranean Diet. Park. Relat. Disord. 2017, 42, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Maraki, M.I.; Yannakoulia, M.; Stamelou, M.; Stefanis, L.; Xiromerisiou, G.; Kosmidis, M.H.; Dardiotis, E.; Hadjigeorgiou, G.M.; Sakka, P.; Anastasiou, C.A.; et al. Mediterranean Diet Adherence Is Related to Reduced Probability of Prodromal Parkinson’s Disease. Mov. Disord. 2019, 34, 48–57. [Google Scholar] [CrossRef]
- Paknahad, Z.; Sheklabadi, E.; Moravejolahkami, A.R.; Chitsaz, A.; Hassanzadeh, A. The Effects of Mediterranean Diet on Severity of Disease and Serum Total Antioxidant Capacity (TAC) in Patients with Parkinson’s Disease: A Single Center, Randomized Controlled Trial. Nutr. Neurosci. 2022, 25, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Alcalay, R.N.; Gu, Y.; Mejia-Santana, H.; Cote, L.; Marder, K.S.; Scarmeas, N. The Association between Mediterranean Diet Adherence and Parkinson’s Disease. Mov. Disord. 2012, 27, 771–774. [Google Scholar] [CrossRef] [Green Version]
- Strikwerda, A.J.; Dommershuijsen, L.J.; Ikram, M.K.; Voortman, T. Diet Quality and Risk of Parkinson’s Disease: The Rotterdam Study. Nutrients 2021, 13, 3970. [Google Scholar] [CrossRef]
- Yin, W.; Löf, M.; Pedersen, N.L.; Sandin, S.; Fang, F. Mediterranean Dietary Pattern at Middle Age and Risk of Parkinson’s Disease: A Swedish Cohort Study. Mov. Disord. 2021, 36, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, P.; Wang, Y.; Buchman, A.S.; Holland, T.M.; Bennett, D.A.; Morris, M.C. MIND Diet Associated with Reduced Incidence and Delayed Progression of Parkinsonism in Old Age. J. Nutr. Health Aging 2018, 22, 1211–1215. [Google Scholar] [CrossRef] [PubMed]
- Lawrie, S.; Coe, S.; Mansoubi, M.; Welch, J.; Razzaque, J.; Hu, M.T.; Dawes, H. Dietary Patterns and Nonmotor Symptoms in Parkinson’s Disease: A Cross-Sectional Analysis. J. Am. Nutr. Assoc. 2022, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Koyuncu, H.; Fidan, V.; Toktas, H.; Binay, O.; Celik, H. Effect of Ketogenic Diet versus Regular Diet on Voice Quality of Patients with Parkinson’s Disease. Acta Neurol. Belg. 2021, 121, 1729–1732. [Google Scholar] [CrossRef]
- VanItallie, T.B.; Nonas, C.; di Rocco, A.; Boyar, K.; Hyams, K.; Heymsfield, S.B. Treatment of Parkinson Disease with Diet-Induced Hyperketonemia: A Feasibility Study. Neurology 2005, 64, 728–730. [Google Scholar] [CrossRef]
- Phillips, M.C.L.; Murtagh, D.K.J.; Gilbertson, L.J.; Asztely, F.J.S.; Lynch, C.D.P. Low-Fat versus Ketogenic Diet in Parkinson’s Disease: A Pilot Randomized Controlled Trial. Mov. Disord. 2018, 33, 1306–1314. [Google Scholar] [CrossRef] [Green Version]
- Tidman, M. Effects of a Ketogenic Diet on Symptoms, Biomarkers, Depression, and Anxiety in Parkinson’s Disease: A Case Study. Cureus 2022, 14, e23684. [Google Scholar] [CrossRef]
- Mischley, L.K.; Lau, R.C.; Bennett, R.D. Role of Diet and Nutritional Supplements in Parkinson’s Disease Progression. Oxid Med. Cell Longev. 2017, 2017, 6405278. [Google Scholar] [CrossRef] [Green Version]
- Kalampokini, S.; Becker, A.; Fassbender, K.; Lyros, E.; Unger, M.M. Nonpharmacological Modulation of Chronic Inflammation in Parkinson’s Disease: Role of Diet Interventions. Park. Dis. 2019, 2019, 7535472. [Google Scholar] [CrossRef] [Green Version]
- Jackson, A.; Forsyth, C.B.; Shaikh, M.; Voigt, R.M.; Engen, P.A.; Ramirez, V.; Keshavarzian, A. Diet in Parkinson’s Disease: Critical Role for the Microbiome. Front. Neurol. 2019, 10, 1245. [Google Scholar] [CrossRef]
- Sofi, F.; Cesari, F.; Abbate, R.; Gensini, G.F.; Casini, A. Adherence to Mediterranean Diet and Health Status: Meta-Analysis. BMJ 2008, 337, a1344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solch, R.J.; Aigbogun, J.O.; Voyiadjis, A.G.; Talkington, G.M.; Darensbourg, R.M.; O’Connell, S.; Pickett, K.M.; Perez, S.R.; Maraganore, D.M. Mediterranean Diet Adherence, Gut Microbiota, and Alzheimer’s or Parkinson’s Disease Risk: A Systematic Review. J. Neurol. Sci. 2022, 434, 120166. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.; Bryan, J.; Hodgson, J.; Murphy, K. Definition of the Mediterranean Diet; A Literature Review. Nutrients 2015, 7, 9139–9153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, W.; Nguyen, L.H.; Song, M.; Wang, D.D.; Franzosa, E.A.; Cao, Y.; Joshi, A.; Drew, D.A.; Mehta, R.; Ivey, K.L.; et al. Dietary Fiber Intake, the Gut Microbiome, and Chronic Systemic Inflammation in a Cohort of Adult Men. Genome Med. 2021, 13, 102. [Google Scholar] [CrossRef]
- Koh, A.; de Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Mantrana, I.; Selma-Royo, M.; Alcantara, C.; Collado, M.C. Shifts on Gut Microbiota Associated to Mediterranean Diet Adherence and Specific Dietary Intakes on General Adult Population. Front. Microbiol. 2018, 9, 890. [Google Scholar] [CrossRef] [Green Version]
- Baert, F.; Matthys, C.; Mellaerts, R.; Lemaître, D.; Vlaemynck, G.; Foulon, V. Dietary Intake of Parkinson’s Disease Patients. Front. Nutr. 2020, 7, 105. [Google Scholar] [CrossRef]
- Yang, X.; Feng, P.; Ji, R.; Ren, Y.; Wei, W.; Hölscher, C. Therapeutic Application of GLP-1 and GIP Receptor Agonists in Parkinson’s Disease. Expert Opin. Ther. Targets 2022, 26, 445–460. [Google Scholar] [CrossRef]
- Palasz, E.; Wysocka, A.; Gasiorowska, A.; Chalimoniuk, M.; Niewiadomski, W.; Niewiadomska, G. BDNF as a Promising Therapeutic Agent in Parkinson’s Disease. Int. J. Mol. Sci. 2020, 21, 1170. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Chen, L.; Gutin, B.; Zhu, H. Total, Insoluble, and Soluble Dietary Fiber Intake and Insulin Resistance and Blood Pressure in Adolescents. Eur. J. Clin. Nutr. 2019, 73, 1172–1178. [Google Scholar] [CrossRef]
- Namsi, A.; Nury, T.; Hamdouni, H.; Yammine, A.; Vejux, A.; Vervandier-Fasseur, D.; Latruffe, N.; Masmoudi-Kouki, O.; Lizard, G. Induction of Neuronal Differentiation of Murine N2a Cells by Two Polyphenols Present in the Mediterranean Diet Mimicking Neurotrophins Activities: Resveratrol and Apigenin. Diseases 2018, 6, 67. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Munishkina, L.A.; Fink, A.L.; Uversky, V.N. Molecular Mechanisms Underlying the Flavonoid-Induced Inhibition of α-Synuclein Fibrillation. Biochemistry 2009, 48, 8206–8224. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Cassidy, A.; Schwarzschild, M.A.; Rimm, E.B.; Ascherio, A. Habitual Intake of Dietary Flavonoids and Risk of Parkinson Disease. Neurology 2012, 78, 1138–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, U.J.; Kim, S.R. Beneficial Effects of Flavonoids Against Parkinson’s Disease. J. Med. Food 2018, 21, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Giuliano, C.; Cerri, S.; Blandini, F. Potential Therapeutic Effects of Polyphenols in Parkinson’s Disease: In Vivo and in Vitro Pre-Clinical Studies. Neural Regen Res. 2021, 16, 234. [Google Scholar] [CrossRef] [PubMed]
- di Rosa, G.; Brunetti, G.; Scuto, M.; Trovato Salinaro, A.; Calabrese, E.J.; Crea, R.; Schmitz-Linneweber, C.; Calabrese, V.; Saul, N. Healthspan Enhancement by Olive Polyphenols in C. Elegans Wild Type and Parkinson’s Models. Int. J. Mol. Sci. 2020, 21, 3893. [Google Scholar] [CrossRef]
- Gallardo-Fernández, M.; Hornedo-Ortega, R.; Alonso-Bellido, I.M.; Rodríguez-Gómez, J.A.; Troncoso, A.M.; García-Parrilla, M.C.; Venero, J.L.; Espinosa-Oliva, A.M.; de Pablos, R.M. Hydroxytyrosol Decreases LPS- and α-Synuclein-Induced Microglial Activation In Vitro. Antioxidants 2019, 9, 36. [Google Scholar] [CrossRef] [Green Version]
- Brunetti, G.; di Rosa, G.; Scuto, M.; Leri, M.; Stefani, M.; Schmitz-Linneweber, C.; Calabrese, V.; Saul, N. Healthspan Maintenance and Prevention of Parkinson’s-like Phenotypes with Hydroxytyrosol and Oleuropein Aglycone in C. Elegans. Int. J. Mol. Sci. 2020, 21, 2588. [Google Scholar] [CrossRef] [Green Version]
- Godos, J.; Tieri, M.; Ghelfi, F.; Titta, L.; Marventano, S.; Lafranconi, A.; Gambera, A.; Alonzo, E.; Sciacca, S.; Buscemi, S.; et al. Dairy Foods and Health: An Umbrella Review of Observational Studies. Int. J. Food Sci. Nutr. 2020, 71, 138–151. [Google Scholar] [CrossRef] [PubMed]
- Comerford, K.; Pasin, G. Gene–Dairy Food Interactions and Health Outcomes: A Review of Nutrigenetic Studies. Nutrients 2017, 9, 710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paganoni, S.; Schwarzschild, M.A. Urate as a Marker of Risk and Progression of Neurodegenerative Disease. Neurotherapeutics 2017, 14, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Dardiotis, E.; Xiromerisiou, G.; Hadjichristodoulou, C.; Tsatsakis, A.M.; Wilks, M.F.; Hadjigeorgiou, G.M. The Interplay between Environmental and Genetic Factors in Parkinson’s Disease Susceptibility: The Evidence for Pesticides. Toxicology 2013, 307, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.C.; Tangney, C.C.; Wang, Y.; Sacks, F.M.; Barnes, L.L.; Bennett, D.A.; Aggarwal, N.T. MIND Diet Slows Cognitive Decline with Aging. Alzheimer’s Dement. 2015, 11, 1015–1022. [Google Scholar] [CrossRef] [Green Version]
- Pietrzak, D.; Kasperek, K.; Rękawek, P.; Piątkowska-Chmiel, I. The Therapeutic Role of Ketogenic Diet in Neurological Disorders. Nutrients 2022, 14, 1952. [Google Scholar] [CrossRef]
- Włodarek, D. Role of Ketogenic Diets in Neurodegenerative Diseases (Alzheimer’s Disease and Parkinson’s Disease). Nutrients 2019, 11, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartman, A.L.; Vining, E.P.G. Clinical Aspects of the Ketogenic Diet. Epilepsia 2007, 48, 31–42. [Google Scholar] [CrossRef]
- Cheng, B.; Yang, X.; An, L.; Gao, B.; Liu, X.; Liu, S. Ketogenic Diet Protects Dopaminergic Neurons against 6-OHDA Neurotoxicity via up-Regulating Glutathione in a Rat Model of Parkinson’s Disease. Brain Res. 2009, 1286, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Cheng, B. Neuroprotective and Anti-Inflammatory Activities of Ketogenic Diet on MPTP-Induced Neurotoxicity. J. Mol. Neurosci. 2010, 42, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Shaafi, S.; Najmi, S.; Aliasgharpour, H.; Mahmoudi, J.; Sadigh-Etemad, S.; Farhoudi, M.; Baniasadi, N. The Efficacy of the Ketogenic Diet on Motor Functions in Parkinson’s Disease: A Rat Model. Iran J. Neurol. 2016, 15, 63–69. [Google Scholar]
- Tieu, K.; Perier, C.; Caspersen, C.; Teismann, P.; Wu, D.-C.; Yan, S.-D.; Naini, A.; Vila, M.; Jackson-Lewis, V.; Ramasamy, R.; et al. D-β-Hydroxybutyrate Rescues Mitochondrial Respiration and Mitigates Features of Parkinson Disease. J. Clin. Investig. 2003, 112, 892–901. [Google Scholar] [CrossRef] [Green Version]
- Cornille, E.; Abou-Hamdan, M.; Khrestchatisky, M.; Nieoullon, A.; de Reggi, M.; Gharib, B. Enhancement of L-3-Hydroxybutyryl-CoA Dehydrogenase Activity and Circulating Ketone Body Levels by Pantethine. Relevance to Dopaminergic Injury. BMC Neurosci. 2010, 11, 51. [Google Scholar] [CrossRef]
- Pinto, A.; Bonucci, A.; Maggi, E.; Corsi, M.; Businaro, R. Anti-Oxidant and Anti-Inflammatory Activity of Ketogenic Diet: New Perspectives for Neuroprotection in Alzheimer’s Disease. Antioxidants 2018, 7, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, N.J.; Wodschow, H.Z.; Nilsson, M.; Rungby, J. Effects of Ketone Bodies on Brain Metabolism and Function in Neurodegenerative Diseases. Int. J. Mol. Sci. 2020, 21, 8767. [Google Scholar] [CrossRef]
- Huang, C.; Wang, P.; Xu, X.; Zhang, Y.; Gong, Y.; Hu, W.; Gao, M.; Wu, Y.; Ling, Y.; Zhao, X.; et al. The Ketone Body Metabolite β-Hydroxybutyrate Induces an Antidepression-Associated Ramification of Microglia via HDACs Inhibition-Triggered Akt-Small RhoGTPase Activation. Glia 2018, 66, 256–278. [Google Scholar] [CrossRef] [PubMed]
- Youm, Y.-H.; Nguyen, K.Y.; Grant, R.W.; Goldberg, E.L.; Bodogai, M.; Kim, D.; D’Agostino, D.; Planavsky, N.; Lupfer, C.; Kanneganti, T.D.; et al. The Ketone Metabolite β-Hydroxybutyrate Blocks NLRP3 Inflammasome–Mediated Inflammatory Disease. Nat. Med. 2015, 21, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Shimazu, T.; Hirschey, M.D.; Newman, J.; He, W.; Shirakawa, K.; le Moan, N.; Grueter, C.A.; Lim, H.; Saunders, L.R.; Stevens, R.D.; et al. Suppression of Oxidative Stress by β-Hydroxybutyrate, an Endogenous Histone Deacetylase Inhibitor. Science (1979) 2013, 339, 211–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, S.-P.; Wang, J.-F.; Xue, W.-J.; Liu, H.-M.; Liu, B.; Zeng, Y.-L.; Li, S.-N.; Huang, B.-X.; Lv, Q.-K.; Wang, W.; et al. Anti-Inflammatory Effects of BHBA in Both in Vivo and in Vitro Parkinson’s Disease Models Are Mediated by GPR109A-Dependent Mechanisms. J. Neuroinflamm. 2015, 12, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, G.; Lv, T.; Zhang, M.; Chen, P.; Sun, Q.; Zhang, J.; Li, Q. β-Hydroxybutyrate (β-HB) Exerts Anti-Inflammatory and Antioxidant Effects in Lipopolysaccharide (LPS)-Stimulated Macrophages in Liza Haematocheila. Fish Shellfish. Immunol. 2020, 107, 444–451. [Google Scholar] [CrossRef]
- Elbarbry, F.; Nguyen, V.; Mirka, A.; Zwickey, H.; Rosenbaum, R. A New Validated HPLC Method for the Determination of Levodopa: Application to Study the Impact of Ketogenic Diet on the Pharmacokinetics of Levodopa in Parkinson’s Participants. Biomed. Chromatogr. 2019, 33, e4382. [Google Scholar] [CrossRef] [Green Version]
- Calder, P.C. Omega-3 Fatty Acids and Inflammatory Processes. Nutrients 2010, 2, 355–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avallone, R.; Vitale, G.; Bertolotti, M. Omega-3 Fatty Acids and Neurodegenerative Diseases: New Evidence in Clinical Trials. Int. J. Mol. Sci. 2019, 20, 4256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Lau, L.M.L.; Bornebroek, M.; Witteman, J.C.M.; Hofman, A.; Koudstaal, P.J.; Breteler, M.M.B. Dietary Fatty Acids and the Risk of Parkinson Disease. Neurology 2005, 64, 2040–2045. [Google Scholar] [CrossRef] [PubMed]
- Abbott, R.D.; Webster Ross, G.; White, L.R.; Sanderson, W.T.; Burchfiel, C.M.; Kashon, M.; Sharp, D.S.; Masaki, K.H.; Curb, J.D.; Petrovitch, H. Environmental, Life-Style, and Physical Precursors of Clinical Parkinson?S Disease: Recent Findings from the Honolulu-Asia Aging Study. J. Neurol. 2003, 250, iii30–iii39. [Google Scholar] [CrossRef] [PubMed]
- Pantzaris, M.; Loukaides, G.; Paraskevis, D.; Kostaki, E.-G.; Patrikios, I. Neuroaspis PLP10TM, a Nutritional Formula Rich in Omega-3 and Omega-6 Fatty Acids with Antioxidant Vitamins Including Gamma-Tocopherol in Early Parkinson’s Disease: A Randomized, Double-Blind, Placebo-Controlled Trial. Clin. Neurol. Neurosurg. 2021, 210, 106954. [Google Scholar] [CrossRef] [PubMed]
- Reijnders, J.S.A.M.; Ehrt, U.; Weber, W.E.J.; Aarsland, D.; Leentjens, A.F.G. A Systematic Review of Prevalence Studies of Depression in Parkinson’s Disease. Mov. Disord. 2008, 23, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Murakami, K.; Sasaki, S. Dietary Intake and Depressive Symptoms: A Systematic Review of Observational Studies. Mol. Nutr. Food Res. 2010, 54, 471–488. [Google Scholar] [CrossRef] [PubMed]
- Grosso, G.; Micek, A.; Marventano, S.; Castellano, S.; Mistretta, A.; Pajak, A.; Galvano, F. Dietary N-3 PUFA, Fish Consumption and Depression: A Systematic Review and Meta-Analysis of Observational Studies. J. Affect Disord. 2016, 205, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Assies, J.; Lok, A.; Bockting, C.L.; Weverling, G.J.; Lieverse, R.; Visser, I.; Abeling, N.G.G.M.; Duran, M.; Schene, A.H. Fatty Acids and Homocysteine Levels in Patients with Recurrent Depression: An Explorative Pilot Study. Prostaglandins Leukot Essent Fat. Acids 2004, 70, 349–356. [Google Scholar] [CrossRef]
- Sánchez-Villegas, A.; Álvarez-Pérez, J.; Toledo, E.; Salas-Salvadó, J.; Ortega-Azorín, C.; Zomeño, M.; Vioque, J.; Martínez, J.; Romaguera, D.; Pérez-López, J.; et al. Seafood Consumption, Omega-3 Fatty Acids Intake, and Life-Time Prevalence of Depression in the PREDIMED-Plus Trial. Nutrients 2018, 10, 2000. [Google Scholar] [CrossRef] [Green Version]
- da Silva, T.M.; Munhoz, R.P.; Alvarez, C.; Naliwaiko, K.; Kiss, Á.; Andreatini, R.; Ferraz, A.C. Depression in Parkinson’s Disease: A Double-Blind, Randomized, Placebo-Controlled Pilot Study of Omega-3 Fatty-Acid Supplementation. J. Affect Disord. 2008, 111, 351–359. [Google Scholar] [CrossRef]
- Li, P.; Song, C. Potential Treatment of Parkinson’s Disease with Omega-3 Polyunsaturated Fatty Acids. Nutr. Neurosci. 2022, 25, 180–191. [Google Scholar] [CrossRef]
- Kujawska, M.; Domanskyi, A.; Kreiner, G. Editorial: Common Pathways Linking Neurodegenerative Diseases—The Role of Inflammation. Front. Cell. Neurosci. 2021, 15, 754051. [Google Scholar] [CrossRef] [PubMed]
- Subramaniam, S.R.; Federoff, H.J. Targeting Microglial Activation States as a Therapeutic Avenue in Parkinson’s Disease. Front. Aging Neurosci. 2017, 9, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijeyekoon, R.S.; Moore, S.F.; Farrell, K.; Breen, D.P.; Barker, R.A.; Williams-Gray, C.H. Cerebrospinal Fluid Cytokines and Neurodegeneration-Associated Proteins in Parkinson’s Disease. Mov. Disord. 2020, 35, 1062–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tansey, M.G.; Goldberg, M.S. Neuroinflammation in Parkinson’s Disease: Its Role in Neuronal Death and Implications for Therapeutic Intervention. Neurobiol. Dis. 2010, 37, 510–518. [Google Scholar] [CrossRef]
- Hirsch, E.C.; Hunot, S. Neuroinflammation in Parkinson’s Disease: A Target for Neuroprotection? Lancet Neurol. 2009, 8, 382–397. [Google Scholar] [CrossRef]
- Teleanu, D.M.; Niculescu, A.-G.; Lungu, I.I.; Radu, C.I.; Vladâcenco, O.; Roza, E.; Costăchescu, B.; Grumezescu, A.M.; Teleanu, R.I. An Overview of Oxidative Stress, Neuroinflammation, and Neurodegenerative Diseases. Int. J. Mol. Sci. 2022, 23, 5938. [Google Scholar] [CrossRef]
- Taghizadeh, M.; Tamtaji, O.R.; Dadgostar, E.; Daneshvar Kakhaki, R.; Bahmani, F.; Abolhassani, J.; Aarabi, M.H.; Kouchaki, E.; Memarzadeh, M.R.; Asemi, Z. The Effects of Omega-3 Fatty Acids and Vitamin E Co-Supplementation on Clinical and Metabolic Status in Patients with Parkinson’s Disease: A Randomized, Double-Blind, Placebo-Controlled Trial. Neurochem. Int. 2017, 108, 183–189. [Google Scholar] [CrossRef]
- Cardoso, H.D.; dos Santos Junior, E.F.; de Santana, D.F.; Gonçalves-Pimentel, C.; Angelim, M.K.; Isaac, A.R.; Lagranha, C.J.; Guedes, R.C.A.; Beltrão, E.I.; Morya, E.; et al. Omega-3 Deficiency and Neurodegeneration in the Substantia Nigra: Involvement of Increased Nitric Oxide Production and Reduced BDNF Expression. Biochim. Et Biophys. Acta (BBA)-Gen. Subj. 2014, 1840, 1902–1912. [Google Scholar] [CrossRef]
- Mori, M.A.; Delattre, A.M.; Carabelli, B.; Pudell, C.; Bortolanza, M.; Staziaki, P.V.; Visentainer, J.V.; Montanher, P.F.; del Bel, E.A.; Ferraz, A.C. Neuroprotective Effect of Omega-3 Polyunsaturated Fatty Acids in the 6-OHDA Model of Parkinson’s Disease Is Mediated by a Reduction of Inducible Nitric Oxide Synthase. Nutr. Neurosci. 2018, 21, 341–351. [Google Scholar] [CrossRef]
- Healy-Stoffel, M.; Levant, B. N-3 (Omega-3) Fatty Acids: Effects on Brain Dopamine Systems and Potential Role in the Etiology and Treatment of Neuropsychiatric Disorders. CNS Neurol. Disord. Drug Targets 2018, 17, 216–232. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Khanna, S. Fecal Microbiota Transplantation. JAMA 2017, 318, 102. [Google Scholar] [CrossRef] [PubMed]
- Tan, P.; Li, X.; Shen, J.; Feng, Q. Fecal Microbiota Transplantation for the Treatment of Inflammatory Bowel Disease: An Update. Front. Pharmacol. 2020, 11, 574533. [Google Scholar] [CrossRef] [PubMed]
- Ramai, D. Fecal Microbiota Transplantation: Donor Relation, Fresh or Frozen, Delivery Methods, Cost-Effectiveness. Ann. Gastroenterol. 2018, 32, 30–38. [Google Scholar] [CrossRef]
- Vendrik, K.E.W.; Ooijevaar, R.E.; de Jong, P.R.C.; Laman, J.D.; van Oosten, B.W.; van Hilten, J.J.; Ducarmon, Q.R.; Keller, J.J.; Kuijper, E.J.; Contarino, M.F. Fecal Microbiota Transplantation in Neurological Disorders. Front. Cell. Infect Microbiol. 2020, 10, 98. [Google Scholar] [CrossRef]
- Kang, D.-W.; Adams, J.B.; Gregory, A.C.; Borody, T.; Chittick, L.; Fasano, A.; Khoruts, A.; Geis, E.; Maldonado, J.; McDonough-Means, S.; et al. Microbiota Transfer Therapy Alters Gut Ecosystem and Improves Gastrointestinal and Autism Symptoms: An Open-Label Study. Microbiome 2017, 5, 10. [Google Scholar] [CrossRef]
- Evrensel, A.; Ceylan, M.E. Fecal Microbiota Transplantation and Its Usage in Neuropsychiatric Disorders. Clin. Psychopharmacol. Neurosci. 2016, 14, 231–237. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.-Q. Fecal Microbiota Transplantation Broadening Its Application beyond Intestinal Disorders. World J. Gastroenterol. 2015, 21, 102. [Google Scholar] [CrossRef]
- Sun, M.-F.; Zhu, Y.-L.; Zhou, Z.-L.; Jia, X.-B.; Xu, Y.-D.; Yang, Q.; Cui, C.; Shen, Y.-Q. Neuroprotective Effects of Fecal Microbiota Transplantation on MPTP-Induced Parkinson’s Disease Mice: Gut Microbiota, Glial Reaction and TLR4/TNF-α Signaling Pathway. Brain Behav. Immun. 2018, 70, 48–60. [Google Scholar] [CrossRef]
- Zhao, Z.; Ning, J.; Bao, X.; Shang, M.; Ma, J.; Li, G.; Zhang, D. Fecal Microbiota Transplantation Protects Rotenone-Induced Parkinson’s Disease Mice via Suppressing Inflammation Mediated by the Lipopolysaccharide-TLR4 Signaling Pathway through the Microbiota-Gut-Brain Axis. Microbiome 2021, 9, 226. [Google Scholar] [CrossRef]
- Zhong, Z.; Chen, W.; Gao, H.; Che, N.; Xu, M.; Yang, L.; Zhang, Y.; Ye, M. Fecal Microbiota Transplantation Exerts a Protective Role in MPTP-Induced Parkinson’s Disease via the TLR4/PI3K/AKT/NF-ΚB Pathway Stimulated by α-Synuclein. Neurochem. Res. 2021, 46, 3050–3058. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Wang, T.; Chen, X.; Zhao, Z.; Chen, Z. Gut Microbiota Relieves Inflammation in the Substantia Nigra of Chronic Parkinson’s Disease by Protecting the Function of Dopamine Neurons. Exp. Ther. Med. 2021, 23, 52. [Google Scholar] [CrossRef]
- Zhou, Z.-L.; Jia, X.-B.; Sun, M.-F.; Zhu, Y.-L.; Qiao, C.-M.; Zhang, B.-P.; Zhao, L.-P.; Yang, Q.; Cui, C.; Chen, X.; et al. Neuroprotection of Fasting Mimicking Diet on MPTP-Induced Parkinson’s Disease Mice via Gut Microbiota and Metabolites. Neurotherapeutics 2019, 16, 741–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Xu, H.; Luo, Q.; He, J.; Li, M.; Chen, H.; Tang, W.; Nie, Y.; Zhou, Y. Fecal Microbiota Transplantation to Treat Parkinson’s Disease with Constipation. Medicine 2019, 98, e16163. [Google Scholar] [CrossRef] [PubMed]
- Kuai, X.; Yao, X.; Xu, L.; Zhou, Y.; Zhang, L.; Liu, Y.; Pei, S.; Zhou, C. Evaluation of Fecal Microbiota Transplantation in Parkinson’s Disease Patients with Constipation. Microb. Cell. Fact 2021, 20, 98. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.-J.; Yang, X.-Z.; Tong, Q.; Shen, P.; Ma, S.-J.; Wu, S.-N.; Zheng, J.-L.; Wang, H.-G. Fecal Microbiota Transplantation Therapy for Parkinson’s Disease. Medicine 2020, 99, e22035. [Google Scholar] [CrossRef] [PubMed]
- Segal, A.; Zlotnik, Y.; Moyal-Atias, K.; Abuhasira, R.; Ifergane, G. Fecal Microbiota Transplant as a Potential Treatment for Parkinson’s Disease—A Case Series. Clin. Neurol. Neurosurg. 2021, 207, 106791. [Google Scholar] [CrossRef]
- Knott, C.; Stern, G.; Wilkin, G.P. Inflammatory Regulators in Parkinson’s Disease: INOS, Lipocortin-1, and Cyclooxygenases-1 and -2. Mol. Cell. Neurosci. 2000, 16, 724–739. [Google Scholar] [CrossRef]
- Li, J.; Ma, S.; Chen, J.; Hu, K.; Li, Y.; Zhang, Z.; Su, Z.; Woodgett, J.R.; Li, M.; Huang, Q. GSK-3β Contributes to Parkinsonian Dopaminergic Neuron Death: Evidence From Conditional Knockout Mice and Tideglusib. Front. Mol. Neurosci. 2020, 13, 81. [Google Scholar] [CrossRef]
- Chen, X.; Hu, Y.; Cao, Z.; Liu, Q.; Cheng, Y. Cerebrospinal Fluid Inflammatory Cytokine Aberrations in Alzheimer’s Disease, Parkinson’s Disease and Amyotrophic Lateral Sclerosis: A Systematic Review and Meta-Analysis. Front. Immunol. 2018, 9, 2122. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.H.; Cho, Y.-S. Fecal Microbiota Transplantation: Current Applications, Effectiveness, and Future Perspectives. Clin. Endosc. 2016, 49, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Schmulson, M.; Bashashati, M. Fecal Microbiota Transfer for Bowel Disorders: Efficacy or Hype? Curr. Opin. Pharmacol. 2018, 43, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Shanahan, F.; Quigley, E.M.M. Manipulation of the Microbiota for Treatment of IBS and IBD—Challenges and Controversies. Gastroenterology 2014, 146, 1554–1563. [Google Scholar] [CrossRef] [PubMed]
- Lopetuso, L.R.; Ianiro, G.; Allegretti, J.R.; Bibbò, S.; Gasbarrini, A.; Scaldaferri, F.; Cammarota, G. Fecal Transplantation for Ulcerative Colitis: Current Evidence and Future Applications. Expert Opin. Biol. Ther. 2020, 20, 343–351. [Google Scholar] [CrossRef]
- Wang, J.-W.; Kuo, C.-H.; Kuo, F.-C.; Wang, Y.-K.; Hsu, W.-H.; Yu, F.-J.; Hu, H.-M.; Hsu, P.-I.; Wang, J.-Y.; Wu, D.-C. Fecal Microbiota Transplantation: Review and Update. J. Formos. Med. Assoc. 2019, 118, S23–S31. [Google Scholar] [CrossRef]
- Aroniadis, O.C.; Brandt, L.J. Fecal Microbiota Transplantation. Curr. Opin. Gastroenterol. 2013, 29, 79–84. [Google Scholar] [CrossRef]
- Singh, H.; Torralba, M.G.; Moncera, K.J.; DiLello, L.; Petrini, J.; Nelson, K.E.; Pieper, R. Gastro-Intestinal and Oral Microbiome Signatures Associated with Healthy Aging. Geroscience 2019, 41, 907–921. [Google Scholar] [CrossRef]
- Irfan, M.; Delgado, R.Z.R.; Frias-Lopez, J. The Oral Microbiome and Cancer. Front. Immunol. 2020, 11, 591088. [Google Scholar] [CrossRef]
- Hayes, R.B.; Ahn, J.; Fan, X.; Peters, B.A.; Ma, Y.; Yang, L.; Agalliu, I.; Burk, R.D.; Ganly, I.; Purdue, M.P.; et al. Association of Oral Microbiome With Risk for Incident Head and Neck Squamous Cell Cancer. JAMA Oncol. 2018, 4, 358. [Google Scholar] [CrossRef]
- Pathak, J.L.; Yan, Y.; Zhang, Q.; Wang, L.; Ge, L. The Role of Oral Microbiome in Respiratory Health and Diseases. Respir. Med. 2021, 185, 106475. [Google Scholar] [CrossRef]
- Sureda, A.; Daglia, M.; Argüelles Castilla, S.; Sanadgol, N.; Fazel Nabavi, S.; Khan, H.; Belwal, T.; Jeandet, P.; Marchese, A.; Pistollato, F.; et al. Oral Microbiota and Alzheimer’s Disease: Do All Roads Lead to Rome? Pharmacol. Res. 2020, 151, 104582. [Google Scholar] [CrossRef] [PubMed]
- Shoemark, D.K.; Allen, S.J. The Microbiome and Disease: Reviewing the Links between the Oral Microbiome, Aging, and Alzheimer’s Disease. J. Alzheimer’s Dis. 2014, 43, 725–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, I.; Arthur, R.A.; Zhao, L.; Clark, J.; Hu, Y.; Corwin, E.J.; Lah, J. The Oral Microbiome and Inflammation in Mild Cognitive Impairment. Exp. Gerontol. 2021, 147, 111273. [Google Scholar] [CrossRef] [PubMed]
- Yussof, A.; Yoon, P.; Krkljes, C.; Schweinberg, S.; Cottrell, J.; Chu, T.; Chang, S.L. A Meta-Analysis of the Effect of Binge Drinking on the Oral Microbiome and Its Relation to Alzheimer’s Disease. Sci. Rep. 2020, 10, 19872. [Google Scholar] [CrossRef] [PubMed]
- Cirstea, M.S.; Kliger, D.; MacLellan, A.D.; Yu, A.C.; Langlois, J.; Fan, M.; Boroomand, S.; Kharazyan, F.; Hsiung, R.G.Y.; MacVicar, B.A.; et al. The Oral and Fecal Microbiota in a Canadian Cohort of Alzheimer’s Disease. J. Alzheimer’s Dis. 2022, 87, 247–258. [Google Scholar] [CrossRef]
- Rozas, N.S.; Tribble, G.D.; Jeter, C.B. Oral Factors That Impact the Oral Microbiota in Parkinson’s Disease. Microorganisms 2021, 9, 1616. [Google Scholar] [CrossRef] [PubMed]
- Mihaila, D.; Donegan, J.; Barns, S.; LaRocca, D.; Du, Q.; Zheng, D.; Vidal, M.; Neville, C.; Uhlig, R.; Middleton, F.A. The Oral Microbiome of Early Stage Parkinson’s Disease and Its Relationship with Functional Measures of Motor and Non-Motor Function. PLoS ONE 2019, 14, e0218252. [Google Scholar] [CrossRef]
- Jo, S.; Kang, W.; Hwang, Y.S.; Lee, S.H.; Park, K.W.; Kim, M.S.; Lee, H.; Yoon, H.J.; Park, Y.K.; Chalita, M.; et al. Oral and Gut Dysbiosis Leads to Functional Alterations in Parkinson’s Disease. NPJ Park. Dis. 2022, 8, 87. [Google Scholar] [CrossRef] [PubMed]
- Fleury, V.; Zekeridou, A.; Lazarevic, V.; Gaïa, N.; Giannopoulou, C.; Genton, L.; Cancela, J.; Girard, M.; Goldstein, R.; Bally, J.F.; et al. Oral Dysbiosis and Inflammation in Parkinson’s Disease. J. Park. Dis. 2021, 11, 619–631. [Google Scholar] [CrossRef]
- Zapała, B.; Stefura, T.; Milewicz, T.; Wątor, J.; Piwowar, M.; Wójcik-Pędziwiatr, M.; Doręgowska, M.; Dudek, A.; Jania, Z.; Rudzińska-Bar, M. The Role of the Western Diet and Oral Microbiota in Parkinson’s Disease. Nutrients 2022, 14, 355. [Google Scholar] [CrossRef]
- Lamont, R.J.; Koo, H.; Hajishengallis, G. The Oral Microbiota: Dynamic Communities and Host Interactions. Nat. Rev. Microbiol. 2018, 16, 745–759. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.-K.; Wu, Q.-L.; Peng, Y.-W.; Liang, F.-Y.; You, H.-J.; Feng, Y.-W.; Li, G.; Li, X.-J.; Liu, S.-H.; Li, Y.-C.; et al. Oral P. Gingivalis Impairs Gut Permeability and Mediates Immune Responses Associated with Neurodegeneration in LRRK2 R1441G Mice. J. Neuroinflamm. 2020, 17, 347. [Google Scholar] [CrossRef]
- Pal, G.; Ramirez, V.; Engen, P.A.; Naqib, A.; Forsyth, C.B.; Green, S.J.; Mahdavinia, M.; Batra, P.S.; Tajudeen, B.A.; Keshavarzian, A. Deep Nasal Sinus Cavity Microbiota Dysbiosis in Parkinson’s Disease. NPJ Park. Dis. 2021, 7, 111. [Google Scholar] [CrossRef]
- Xie, J.; Tian, S.; Liu, J.; Cao, R.; Yue, P.; Cai, X.; Shang, Q.; Yang, M.; Han, L.; Zhang, D. Dual Role of the Nasal Microbiota in Neurological Diseases—An Unignorable Risk Factor or a Potential Therapy Carrier. Pharmacol. Res. 2022, 179, 106189. [Google Scholar] [CrossRef]
- Wagner Mackenzie, B.; Chang, K.; Zoing, M.; Jain, R.; Hoggard, M.; Biswas, K.; Douglas, R.G.; Taylor, M.W. Longitudinal Study of the Bacterial and Fungal Microbiota in the Human Sinuses Reveals Seasonal and Annual Changes in Diversity. Sci. Rep. 2019, 9, 17416. [Google Scholar] [CrossRef] [PubMed]
- Shen, L. Gut, Oral and Nasal Microbiota and Parkinson’s Disease. Microb. Cell. Fact 2020, 19, 50. [Google Scholar] [CrossRef] [Green Version]
- Boertien, J.M.; Pereira, P.A.B.; Aho, V.T.E.; Scheperjans, F. Increasing Comparability and Utility of Gut Microbiome Studies in Parkinson’s Disease: A Systematic Review. J. Park. Dis. 2019, 9, S297–S312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conlon, M.; Bird, A. The Impact of Diet and Lifestyle on Gut Microbiota and Human Health. Nutrients 2014, 7, 17–44. [Google Scholar] [CrossRef] [PubMed]
- Aslam, H.; Marx, W.; Rocks, T.; Loughman, A.; Chandrasekaran, V.; Ruusunen, A.; Dawson, S.L.; West, M.; Mullarkey, E.; Pasco, J.A.; et al. The Effects of Dairy and Dairy Derivatives on the Gut Microbiota: A Systematic Literature Review. Gut Microbes 2020, 12, 1799533. [Google Scholar] [CrossRef] [PubMed]
- Cortes-Ortiz, L.; Amato, K.R. Host Genetics Influence the Gut Microbiome. Science (1979) 2021, 373, 159–160. [Google Scholar] [CrossRef] [PubMed]
- Jain, N. The Need for Personalized Approaches to Microbiome Modulation. Front. Public Health 2020, 8, 144. [Google Scholar] [CrossRef] [PubMed]
- Vandeputte, D. Personalized Nutrition Through The Gut Microbiota: Current Insights And Future Perspectives. Nutr. Rev. 2020, 78, 66–74. [Google Scholar] [CrossRef]
- Molsberry, S.; Bjornevik, K.; Hughes, K.C.; Healy, B.; Schwarzschild, M.; Ascherio, A. Diet Pattern and Prodromal Features of Parkinson Disease. Neurology 2020, 95, e2095–e2108. [Google Scholar] [CrossRef]



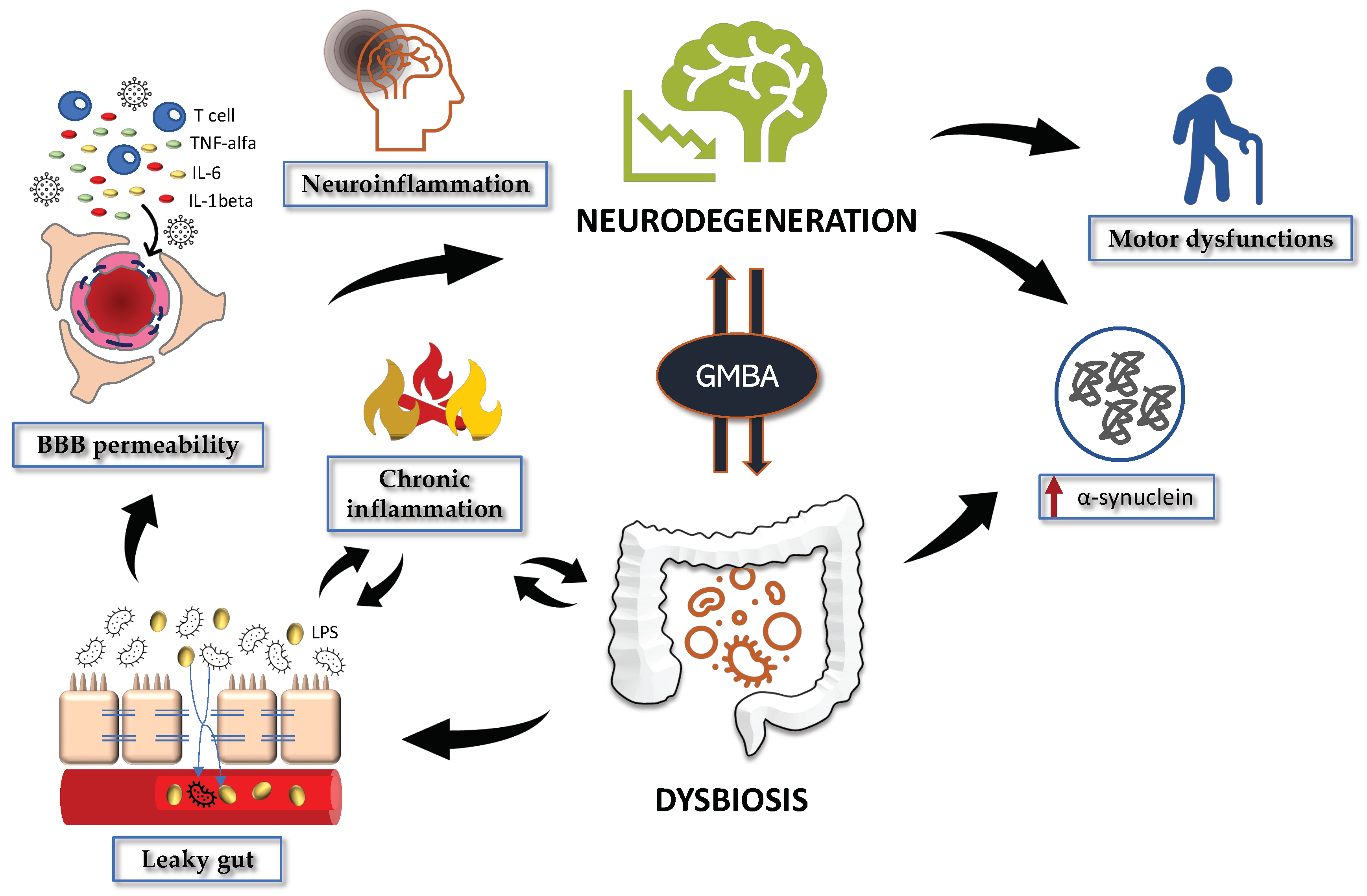{kind=link}
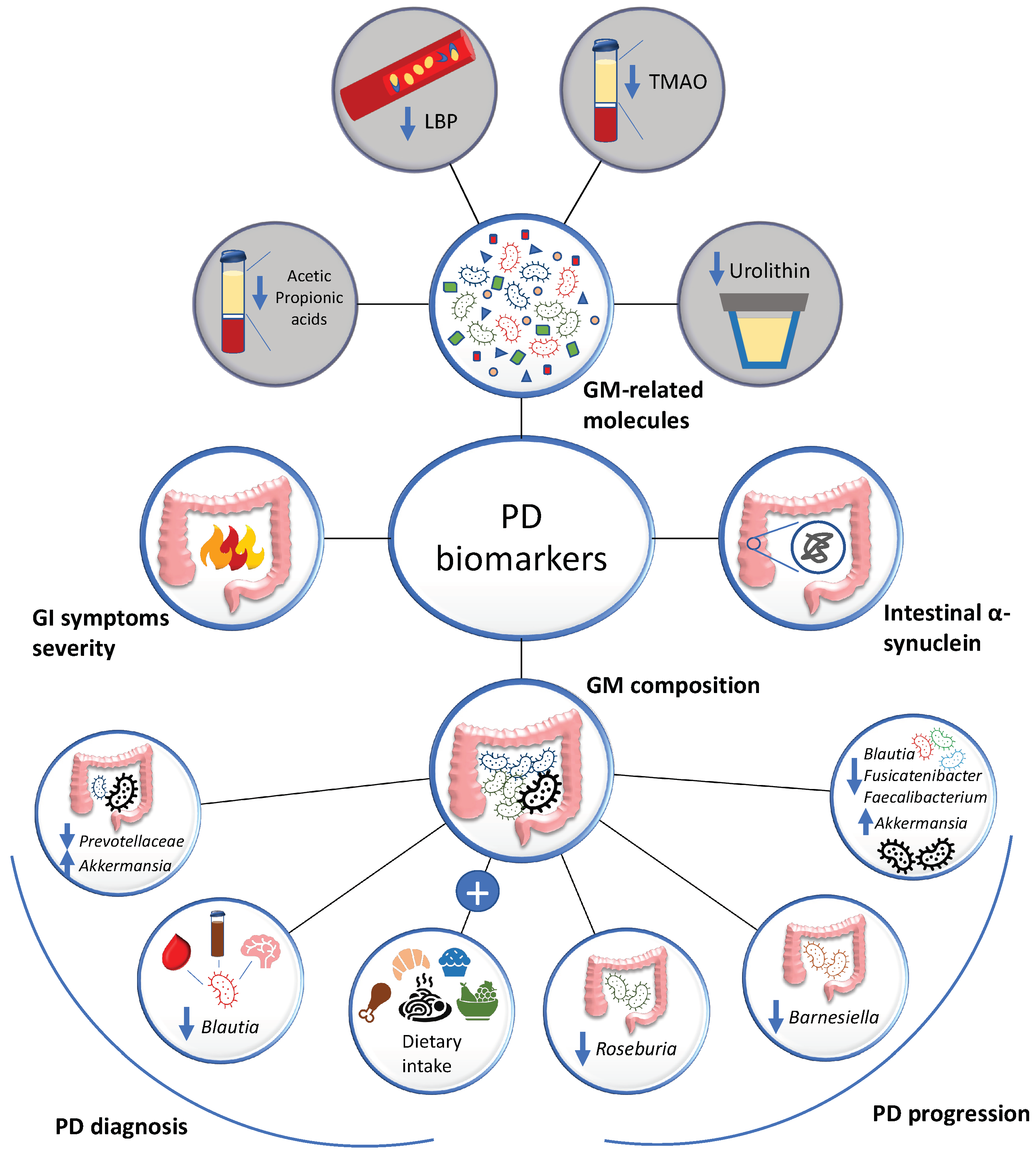{kind=link}
| Probiotic | Experimental Model | Treatment Duration | Treatment Effects | Reference |
|---|---|---|---|---|
| Lactobacillus salivarius, L. plantarum, L. acidophilus, L. rhamnosus, Bifidobacterium animalis subsp. lactis, B. breve | In vitro. PBMCs from 40 PD patients | 24 h | Increase of anti-inflammatory cytokines (IL-4, IL-10) Decrease of pro-inflammatory cytokines (TNF-α, IL-6, IL-17A) Reduction of ROS production | (Magistrelli et al., 2019) [328] |
| In vitro. Caco-2 cells | 2 h | Protection of epithelium from gut permeability | ||
| In vitro. Escherichia coli and Klebsiella pneumoniae inoculation | 48 h | Inhibition of pathogen bacteria proliferation | ||
| Symprove (L. acidophilus, L. plantarum, L. rhamnosus, Enterococcus faecium) | In vitro. Caco-2/THP1 cells | 24 h | Improvement in epithelial tight-junction integrity, and in wound healing Increase of anti-inflammatory cytokines (IL-6, IL-10) Decrease of pro-inflammatory chemokine IL-8 | (Ghyselinck et al., 2021) [329] |
| In vitro. Stool samples from 3 PD patients | 48 h | Modulation of GM composition (↑Firmicutes, ↓Bacteroidetes) Production of SCFAs and lactate | ||
| L. plantarum 200655 | In vitro. H2O2-treated SH-SY5Y cells | 4 h | Increase of BDNF and TH mRNA expression Attenuation of apoptosis (↓apoptosis-related Bax/Bcl-2 ratio, ↓caspase-3 activity) | (Cheon et al., 2021) [330] |
| SLAB51 (Streptococcus thermophilus, B. longum, B. breve, B. infantis, L. acidophilus, L. plantarum, L. paracasei, L. delbrueckii subsp. bulgaricus, L. brevis) | In vitro. 6-OHDA-treated SH-SY5Y cells | 2 h | Reduction in dopaminergic neuronal loss, by increasing the neuronal survival BDNF pathway (mBDNF, p-TrkB, p-ERK5, p-CREB, PI3K/Akt) and decreasing neuronal death pathway (pro-BDNF, p-JNK, p-ERK1,2, P75) Protection against OS (↓4-HNE). | (Castelli et al., 2020) [331] |
| In vivo. 6-OHDA-treated mice | 5 weeks | Improvement in motor impairment. Reduction in neuroinflammation (↓Iba1, ↓GFAP), and in OS, by restoring Nrf2/HO-1 activity and inhibiting NF-κB. | ||
| L. rhamnosus GG, B. animalis lactis, and L. acidophilus | In vivo. MPTP- and rotenone-induced mouse model | 4 weeks | Prevention of dopaminergic neurons loss by upregulation of BDNF and GDNF, and inhibition of MAO-B expression Amelioration of behavioural impairments. Raise in butyrate levels. | (Srivastav et al., 2019) [325] |
| L. plantarum PS128 | In vivo. MPTP-induced mouse model | 28 days | Alleviation of neuroinflammation (↓TNF-α, ↓IL-1β, ↓IL-6) and OS (↑SOD, ↑GSH, ↑CAT, ↑GPx). Improvement in motor deficits and dopaminergic neuronal cell death. Reduction of glial reactivity. Increase in BDNF expression and striatal dopamine level. | (Liao et al., 2020) [332] |
| B. bifidum, B. longum, L. rhamnosis, L. rhamnosus GG, L. plantarum LP28, and Lactococcus lactis subsp. Lactis | In vivo. MitoPark mouse model | 16 weeks | Improvement in motor impairment, balance function, and motor coordination. Reduction in dopaminergic neuronal loss. | (Hsieh et al., 2020) [333] |
| Clostridum butyricum | In vivo. MPTP-induced mouse model | 4 weeks | Reduction in motor impairment, dopaminergic neuronal loss (↑TH) and synaptic dysfunction (↑synapsin I). Inhibition of excessive microglia activation, via reversing GM dysbiosis. Increase in GLP-1 and GLP-1 receptor levels in the brain. | (J. Sun et al., 2021) [334] |
| Lactobacillus plantarum DP189 | In vivo. MPTP-induced mouse model | 14 days | Modulation of OS (↑SOD, ↑GSH-Px ↓MDA, ↓ROS), inflammation (↑IL-10, ↓TNF-α, ↓IL-6 and ↓IL-1β) and GM dysbiosis (↓Proteobacteria, ↓Actinobacteria, ↑Lactobacillus, ↑Prevotella). Reduction in α-SYN accumulation in the substantia nigra. Activation of Nrf2/ARE and PGC-1α pathways. Suppression of NLRP3 inflammasome. | (L. Wang et al., 2022) [335] |
| Lactobacillus casei Shirota | 40 PD patients | 5 weeks | Improvement in stool consistency. Reduction of bloating and abdominal pain. | (Cassani et al., 2011) [336] |
| L. acidophilus and B. infantis | 40 PD patients | 12 weeks | Reduction of bloating and abdominal pain. | (Georgescu et al., 2016) [337] |
| L. casei, L. fermentum, L. acidophilus, B. bifidum | 50 PD patients | 12 weeks | Reduction of IL-1, IL-8 and TNF-α gene expression. Increase in TGF-β and PPAR-γ. | (Borzabadi et al., 2018) [338] |
| L. acidophilus, L. reuteri, L. fermentum, and Bifidobacterium bifidum | 60 PD patients | 12 weeks | Reduction of UPDRS motor scores. Decrease of hs-CRP, MDA, insulin levels, and insulin resistance. Enhancement in GSH levels and insulin sensitivity. | (Tamtaji et al., 2019) [339] |
| Xexbio (L. acidophilus, L. casei, L. lactis, B. infantis and B. longum) | 48 PD patients | 8 weeks | Improvement of constipation and gut motility. | (Ibrahim et al., 2020) [340] |
| L. acidophilus, L. reuteri, L. gasseri, L. rhamnosus, B. bifidum, B. longum, Enterococcus faecalis, E. faecium | 72 PD patients | 4 weeks | Improvement of constipation, stool consistency and quality of life. | (Tan et al., 2020) [341] |
| Probio-M8 (B. animalis subsp. lactis Probio M-8) | 82 PD patients | 12 weeks | Amelioration in sleep quality, cognitive dysfunction, and defecation. Attenuation of anxiety, depression, and gastrointestinal symptoms. Modulation of gut microbiome, lipid metabolism, SCFAs and neurotransmitters serum levels. | (H. Sun et al., 2022) [342] |
| Prebiotic | Experimental Model | Treatment Duration | Treatment Effects | Reference |
|---|---|---|---|---|
| GOS, lcFOS, scFOS, nutriose | Rotenone-induced mice model | 10 weeks | Improvement of motor symptoms, gastrointestinal dysfunction, and inflammation (↓GFAP, ↓T-cells infiltration). Restoration of DAT expression. Reduction of α-synuclein levels. | (Perez-Pardo et al., 2017) [372] |
| Polymannuronic acid | MPTP-induced model mice | 5 weeks | Abolition of the apoptotic process (↓Bax, ↓Bax/Bcl-2 ratio). Prevention of dopaminergic neuronal loss (↑TH gene and protein expression in the striatum). Increase of faecal acetate, butyrate, and total SCFAs levels. Inhibition of striatal inflammation (↓TNF-α mRNA levels). | (Liu et al., 2022) [373] |
| Prebiotic residual medium obtained from L. salivarius subsp. salicinium AP-32 culture medium | 6-OHDA-induced rat model | 8 weeks | Reduction of dopaminergic neuronal loss, motor dysfunctions, and muscle atrophy. Increase of GPx and faecal SCFAs (propionate, butyrate). Restoration of mitochondrial function and energy metabolism. | (Nurrahma et al., 2021) [376] |
| Prebiotic residual medium obtained from L. salivarius subsp. salicinium AP-32 culture medium | 6-OHDA-induced rat model | 8 weeks | Amelioration of motor symptoms. Reduction of inflammation (↓TNF-α) and OS (↓ROS, ↑SOD, ↑GPx). Increase of SCFAs production (propionate, butyrate). Modulation of GM composition (↑Ruminococcaceae, ↑Bifidobacterium, ↑Faecalibacterium, ↓Propionibacterium, ↓Clostridium, ↓Cylindriodes, ↓Ruminantium). | (Tsao et al., 2021) [346] |
| Dietetic fiber supplements (wheat, pectin, dimethylpolyoxyhexane-900) | 19 PD patients | 8 weeks | Improvement in constipation and in motor function. Increase of total plasma levodopa levels. | (Astarloa et al., 1992) [374] |
| Psyllium | 7 PD patients | 8 weeks | Increase in stool frequency and weight. | (Ashraf et al., 1997) [375] |
| Resistant starch | 57 PD patients | 8 weeks | Improvement of non-motor symptoms. Reduction of calprotectin levels. Increase in butyrate concentration. | (Becker et al., 2021) [316] |
| Reference | Type of Study | Type of Dietary Intervention | Aim | Outcomes |
|---|---|---|---|---|
| Metcalfe-Roach et al., 2021 [383] | CrS | MIND or Medi | MIND/Medi vs. PD onset | Both diets delay PD onset; MIND slightly superior in the female subgroup. |
| Paknahad et al., 2020 [384] | CT | Medi | Medi vs. cognitive function | Improvement in executive function, language, attention, concentration, active memory and in the total score of cognitive assessment. |
| Rusch et al., 2021 [385] | CT | Medi | Medi vs. GI function | Correlation with weight loss, improved constipation, and modified gut microbiota in PD patients. |
| Cassani et al., 2017 [386] | Medi | Medi vs. PD progression | No significant correlation. | |
| Maraki et al., 2019 [387] | CS | Medi | Medi vs. PD onset | Correlation with lower probability of prodromal PD in older people. |
| Zamzam Paknahad et al., 2022 [388] | CT | Medi | Medi vs. total antioxidant capacity (TAC) and PD severity | Improvements in TAC and PD severity. |
| Alcalay et al., 2012 [389] | CCS | Medi | Medi vs. PD status | Medi adherence is associated with PD age at onset. |
| Strikwerda et al., 2021 [390] | CS | Medi, Dutch diets | Medi, Dutch diets vs. PD risk | Protective effect. |
| Yin et al., 2021 [391] | CS | Medi | Medi vs. PD risk | Protective effect. |
| Agarwal et al., 2018 [392] | LS | MIND | MIND vs. PD development and progression | Decreased risk and slower progression of PD in older adults. |
| Lawrie et al., 2022 [393] | CrS | MIND | MIND vs. PD severity | Decreased fatigue and depression. |
| Koyuncu et al., 2021 [394] | KD | KD vs. PD patients voice quality | VHI * score improvement | |
| VanItallie et al., 2005 [395] | Feasibility study | KD | KD vs. PD progression | UPDRS scores improvement. |
| Phillips et al., 2018 [396] | CT | KD vs a low-fat, high-carbohydrate diet | KD vs. PD progression | Motor and nonmotor symptoms improvement. |
| Tidman et al., 2022 [397] | CT | KD | KD vs. PD progression | UPDRS scores improvement. |
| Ref. | Study Cohort | Study Groups | Donor | Recipient | Experimental Procedure | Results | Adverse Events |
|---|---|---|---|---|---|---|---|
| Zhao et al., 2021 [470] | Mice | Controls (n = 15), rotenone (n = 15) and rotenone + FMT (n = 15) | Control mice | Rotenone-induced PD mice | Oral gavage (100 μL bacterial suspension) daily for 2 weeks | ↓ Dysbiosis ↓ Motor symptoms ↑ Intestinal barrier and BBB integrity ↓ Systemic inflammation ↓ Neuroinflammation (SN) ↓ LPS (serum, colon and SN) ↓ TLR4/NF-κB pathway (colon and SN) | N.A. |
| Sun et al., 2018 [469] | Mice | Controls (n = 15), MPTP + PBS (n = 15) and MPTP + FMT (n = 15) | Control mice | MPTP-induced PD mice | Gavage (200 μL bacterial suspension containing 108 CFU/mL) daily for 7 days | ↓ Dysbiosis ↓ Fecal SCFAs ↓ Motor symptoms ↑ DA and 5-HT (striatum) ↓ Microglia and astrocyte activation (SN) ↓ TLR4/TNF-α pathway (gut and brain) | N.A. |
| Zhong et al., 2021 [471] | Mice | Controls + PBS (n = 10), controls + FMT (n = 10), MPTP + PBS (n = 10), MPTP + FMT (n = 10) | Control mice | Controls or MPTP-induced PD mice | Gavage (200 μL bacterial suspension containing 108 CFU/mL) daily for 7 days | ↓ Motor symptoms ↓ Fecal SCFAs ↓ α-syn (SN) ↓ Microglia activation (SN) ↓ TLR4/NF-κB pathway (striatum and SN) | N.A. |
| Zhang et al., 2021 [472] | Mice | Controls (n = 3), MPTP (n = 3) and MPTP + FMT (n = 3) | Control mice | MPTP-induced PD mice | Transplantation with 200 μL bacterial suspension (containing 108 CFU/mL) daily for 2 weeks | ↓ Neuroinflammation (SN) ↓ Motor symptoms ↑ Blautia ↓ Anaerostipes, ASF356, Ruminococcus and Bifidobacterium ↓ Microglia and astrocyte activation (SN) ↓ IL-1β, iNOS, GSK3β and p-PTEN (SN) | N.A. |
| Zhou et al., 2019 [473] | Mice | Mice pre-treated with MPTP and antibiotics, divided in PD-PBS (n = 8), PD-NA (n = 8), PD-NF (n = 8) and PD-NF/HK (n = 8) | Control mice or control mice undergoing FMT | MPTP-induced PD mice pre-treated with antibiotics | Gastric gavage (200 μL bacterial suspension containing 108 CFU/mL) daily for 7 days | ↑ DA and 5-HT (striatum) in PD-NF mice ↑ Neuroprotection in PD-NF mice | N.A. |
| Sampson et al., 2016 [40] | Mice | GF + FMT from SPF control mice, GF + FMT from PD patients, GF + FMT from healthy patients | Human PD patients (n = 6), human healthy controls (n = 6) or SPF control mice (n = 3) | α-syn-overexpressing mice | Oral gavage | In GF + PD-FMT mice: ↑ Physical impairment ↑ Proteus, Bilophila and Roseburia ↓ Lachnospiraceae, Rikenellaceae, Peptostreptococcaceae and Butyricicoccus ↓ Acetate ↑ Proprionate and butyrate | N.A. |
| Huang et al., 2019 [474] | Human (case report) | PD patient presenting tremor for 7 years and constipation (>3 years) | 26 y.o. healthy male | 71 y.o. male PD patients | Colonoscopy (200 mL of fecal microbiota suspension) daily for 3 days | ↓ Tremor (no tremor for 2 months) ↓ Constipation ↑ α-diversity ↓ UPDRS score 1 week after FMT | No |
| Kuai et al., 2021 [475] | Humans (prospective single study) | PD patients | Frozen fecal microbiota from the China fmtBank | PD patients (n = 11) | Intra-intestine transplantation of 40–50 mL of frozen fecal microbiota resuspended in 200 mL saline solution | ↑ Blautia and Prevotella ↓ Bacteroidetes ↓ H-Y, UPDRS and NMSS scores ↓ Wexner constipation and PAC-QOL scores | No |
| Xue et al., 2020 [476] | Humans | PD patients + FMT (via colonoscopy, n = 10; nasointestinally, n = 5) | 5 Healthy donors (mean 22 y.o., 3 males and 2 females) | PD patients | Colonoscopy or nasointestinal administration | ↓ PSQI, HAMA, PDQ-39, HAMD, UPDRS-III and NMSQ | 5 cases: diarrhea (n = 2), abdominal pain (n = 2) and flatulence (n = 1) |
| Segal et al., 2021 [477] | Humans (uncontrolled case series) | 6 PD patients with symptoms for 5 years (mean). | 2 healthy donors (males, 38 and 50 y.o.) | 6 PD patients with constipation (mean 52 y.o.), 3 males and 3 females | Colonoscopy (300 mL of fecal suspension) | ↓ Motor and non-motor symptoms ↓ Constipation | 1 case requiring hospitalization for observation |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varesi, A.; Campagnoli, L.I.M.; Fahmideh, F.; Pierella, E.; Romeo, M.; Ricevuti, G.; Nicoletta, M.; Chirumbolo, S.; Pascale, A. The Interplay between Gut Microbiota and Parkinson’s Disease: Implications on Diagnosis and Treatment. Int. J. Mol. Sci. 2022, 23, 12289. https://doi.org/10.3390/ijms232012289
Varesi A, Campagnoli LIM, Fahmideh F, Pierella E, Romeo M, Ricevuti G, Nicoletta M, Chirumbolo S, Pascale A. The Interplay between Gut Microbiota and Parkinson’s Disease: Implications on Diagnosis and Treatment. International Journal of Molecular Sciences. 2022; 23(20):12289. https://doi.org/10.3390/ijms232012289
Chicago/Turabian StyleVaresi, Angelica, Lucrezia Irene Maria Campagnoli, Foroogh Fahmideh, Elisa Pierella, Marcello Romeo, Giovanni Ricevuti, Marchesi Nicoletta, Salvatore Chirumbolo, and Alessia Pascale. 2022. "The Interplay between Gut Microbiota and Parkinson’s Disease: Implications on Diagnosis and Treatment" International Journal of Molecular Sciences 23, no. 20: 12289. https://doi.org/10.3390/ijms232012289