
Non-Cytokine Protein Profile of the Mesenchymal Stem Cell Secretome That Regulates the Androgen Production Pathway

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
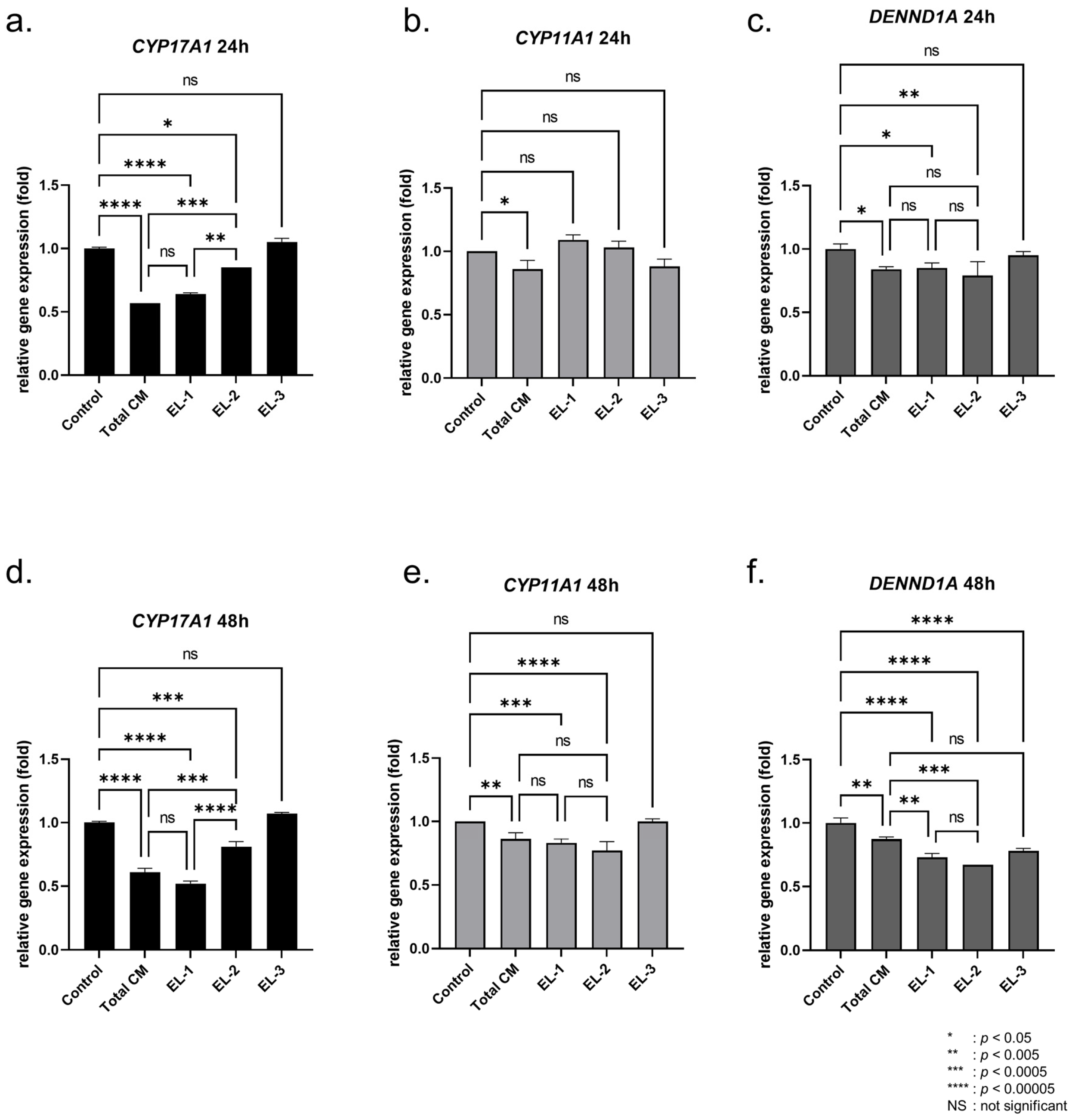
2.1. Effect of Fractionated Sample on Androgen Production
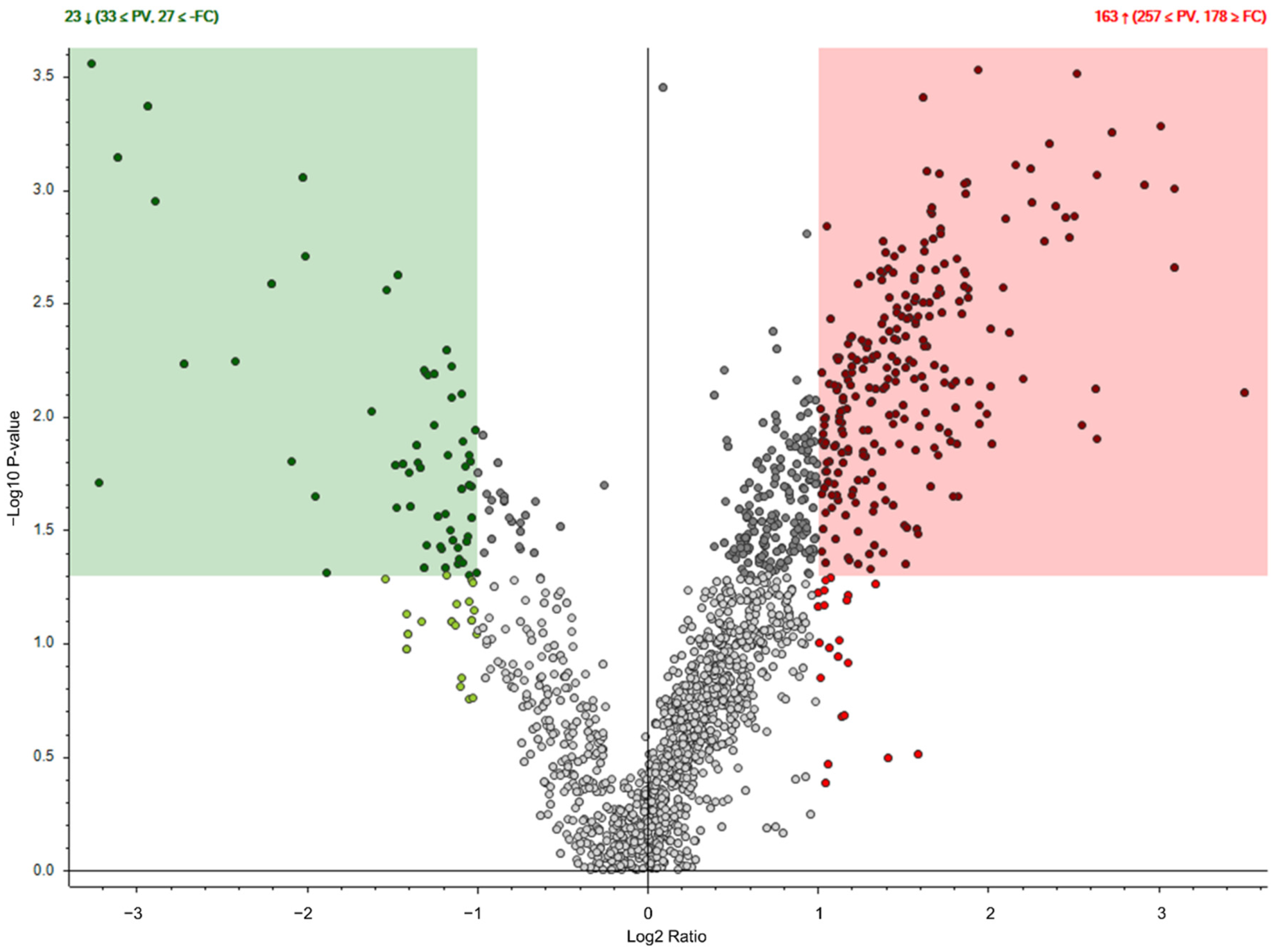
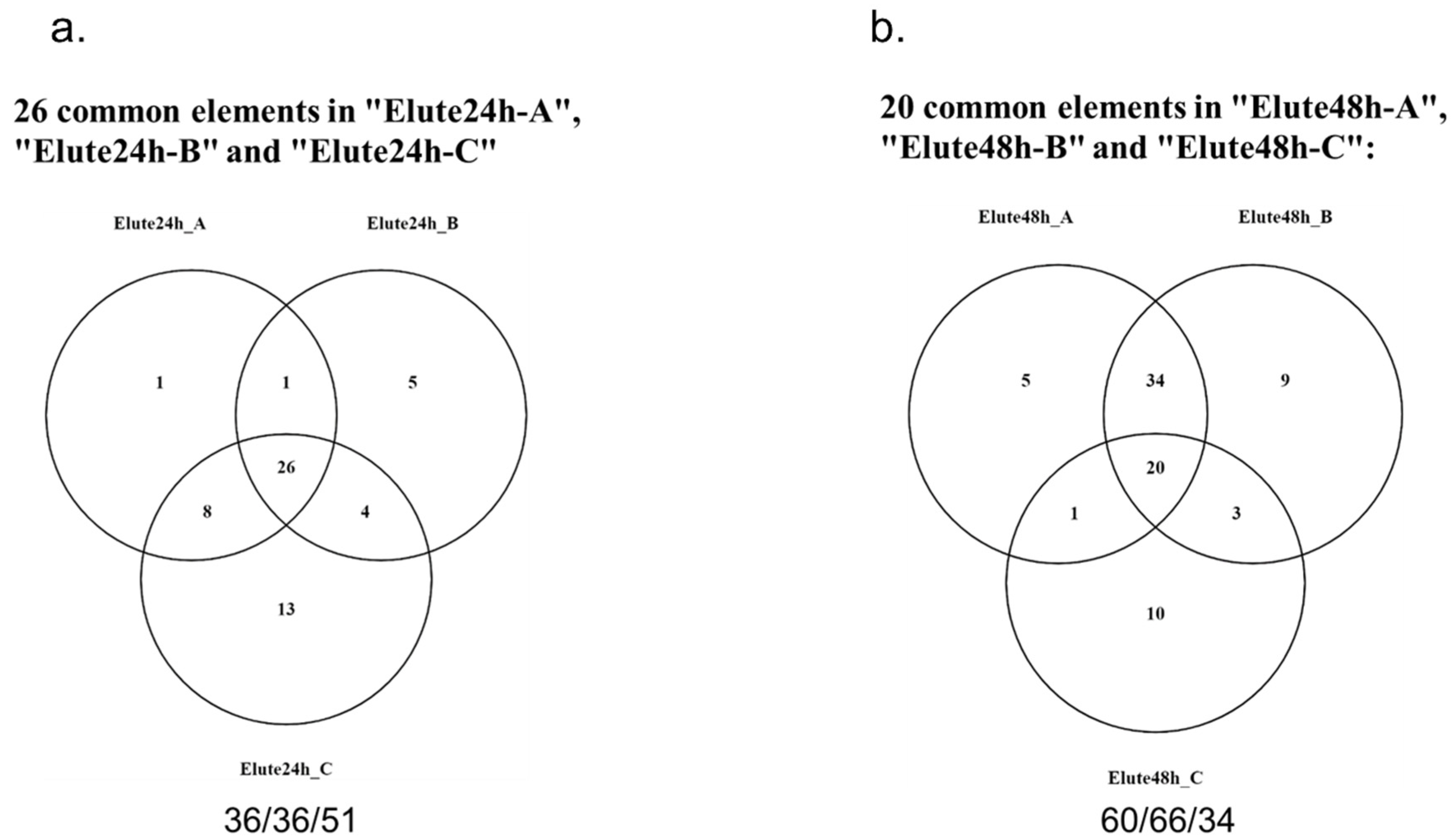
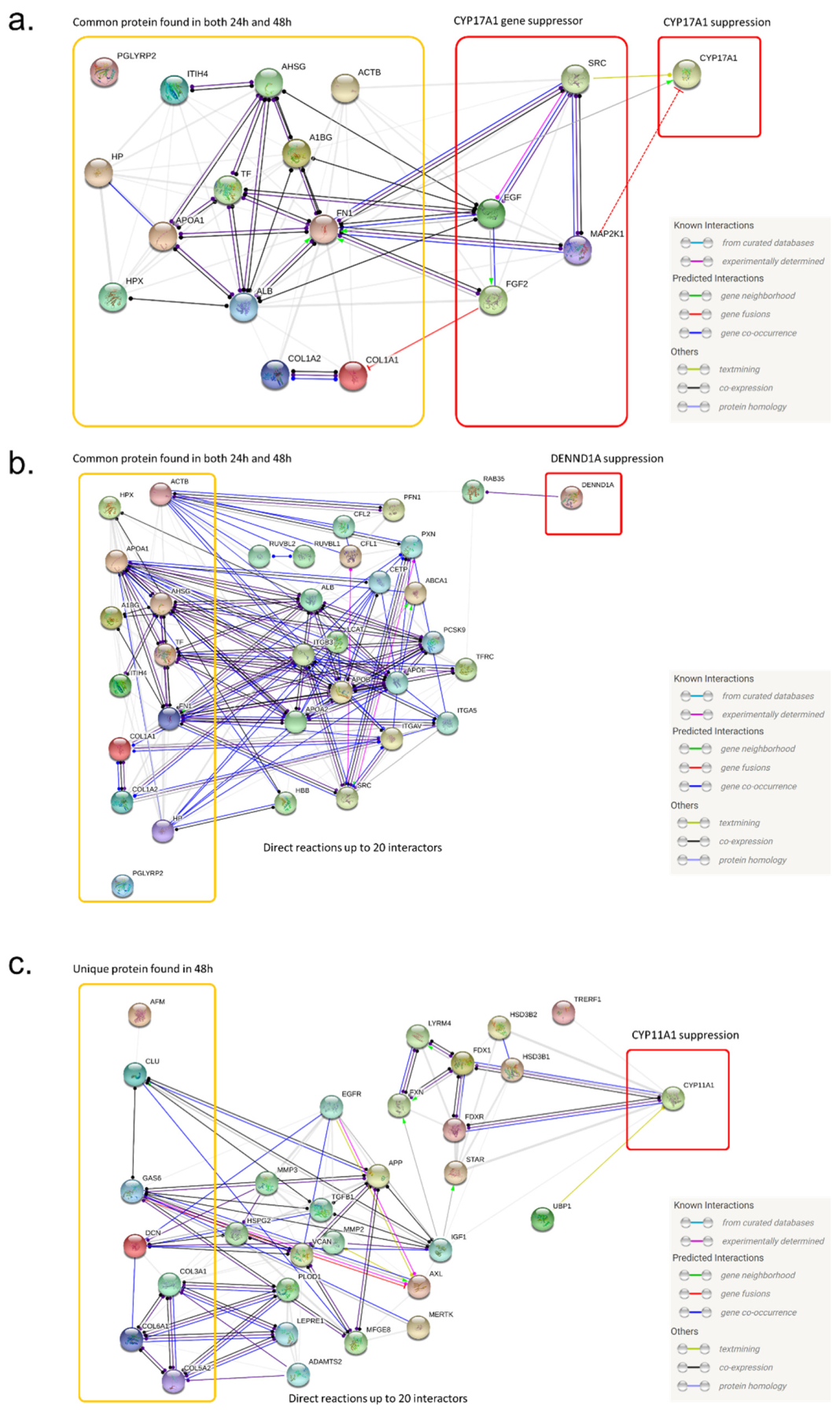
2.2. Identification of MSC Proteins That Regulate Androgen Synthesis
2.3. Regulation of Androgen Synthesis Genes by MSC-Derived Proteins
3. Discussion
4. Materials and Methods
4.1. Human Mesenchymal Stem Cell Culture
4.2. Collection of the Secretome from MSCs
4.3. Human Fibroblast Culture and Secretome Collection
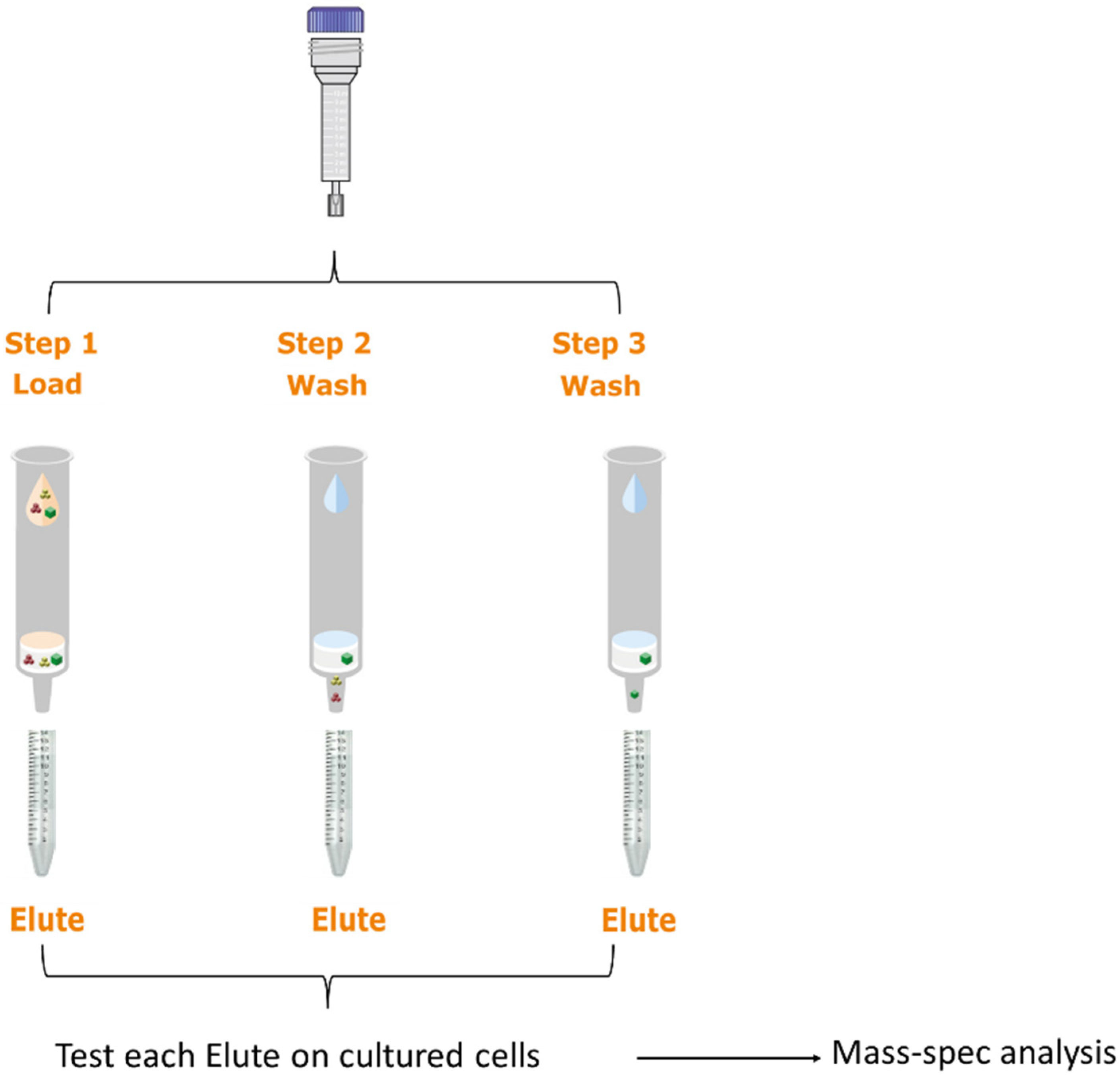
4.4. Fractionation of hBM-MSC-CM
4.5. Activity-Based Analysis of hBM-MSC-CM Fractions
4.6. Quantitative RT-qPCR
4.7. Mass Spectrometry Analysis of MSC-CM
4.8. Protein–Protein Interaction Analysis
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Rosenfield, R.L.; Ehrmann, D.A. The Pathogenesis of Polycystic Ovary Syndrome (PCOS): The Hypothesis of PCOS as Functional Ovarian Hyperandrogenism Revisited. Endocr. Rev. 2016, 37, 467–520. [Google Scholar] [CrossRef] [PubMed]
- González, F. Inflammation in Polycystic Ovary Syndrome: Underpinning of insulin resistance and ovarian dysfunction. Steroids 2012, 77, 300–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dennett, C.C.; Simon, J. The role of polycystic ovary syndrome in reproductive and metabolic health: Overview and approaches for treatment. Diabetes Spectr. 2015, 28, 116–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Luca, C.; Olefsky, J.M. Inflammation and insulin resistance. FEBS Lett. 2008, 582, 97–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbould, A. Effects of androgens on insulin action in women: Is androgen excess a component of female metabolic syndrome? Diabetes Metab. Res. Rev. 2008, 24, 520–532. [Google Scholar] [CrossRef]
- Fox, C.W.; Zhang, L.; Sohni, A.; Doblado, M.; Wilkinson, M.F.; Chang, R.J.; Duleba, A.J. Inflammatory Stimuli Trigger Increased Androgen Production and Shifts in Gene Expression in Theca-Interstitial Cells. Endocrinology 2019, 160, 2946–2958. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.y.; Zhu, F.f.; Zhu, Y.j.; Hu, Y.j.; Chen, X. Effects of IL-18 on the proliferation and steroidogenesis of bovine theca cells: Possible roles in the pathogenesis of polycystic ovary syndrome. J. Cell. Mol. Med. 2021, 25, 1128–1139. [Google Scholar] [CrossRef]
- Rojas, J.; Chávez, M.; Olivar, L.; Rojas, M.; Morillo, J.; Mejías, J.; Calvo, M.; Bermúdez, V. Polycystic Ovary Syndrome, Insulin Resistance, and Obesity: Navigating the Pathophysiologic Labyrinth. Int. J. Reprod. Med. 2014, 2014, 719050. [Google Scholar] [CrossRef]
- Gambineri, A.; Patton, L.; Altieri, P.; Pagotto, U.; Pizzi, C.; Manzoli, L.; Pasquali, R. Polycystic ovary syndrome is a risk factor for type 2 diabetes: Results from a long-term prospective study. Diabetes 2012, 61, 2369–2374. [Google Scholar] [CrossRef] [Green Version]
- Scicchitano, P.; Dentamaro, I.; Carbonara, R.; Bulzis, G.; Dachille, A.; Caputo, P.; Riccardi, R.; Locorotondo, M.; Mandurino, C.; Matteo Ciccone, M. Cardiovascular Risk in Women With PCOS. Int. J. Endocrinol. Metab. 2012, 10, 611–618. [Google Scholar] [CrossRef] [Green Version]
- Ding, D.C.; Chen, W.; Wang, J.H.; Lin, S.Z. Association between polycystic ovarian syndrome and endometrial, ovarian, and breast cancer: A population-based cohort study in Taiwan. Medicine 2018, 97, e12608. [Google Scholar] [CrossRef] [PubMed]
- Calcaterra, V.; Verduci, E.; Cena, H.; Magenes, V.C.; Todisco, C.F.; Tenuta, E.; Gregorio, C.; De Giuseppe, R.; Bosetti, A.; Di Profio, E.; et al. Polycystic Ovary Syndrome in Insulin-Resistant Adolescents with Obesity: The Role of Nutrition Therapy and Food Supplements as a Strategy to Protect Fertility. Nutrients 2021, 13, 1848. [Google Scholar] [CrossRef] [PubMed]
- Salek, M.; Clark, C.C.T.; Taghizadeh, M.; Jafarnejad, S. N-3 fatty acids as preventive and therapeutic agents in attenuating PCOS complications. EXCLI J. 2019, 18, 558–575. [Google Scholar] [CrossRef] [PubMed]
- Wawrzkiewicz-Jalowiecka, A.; Kowalczyk, K.; Trybek, P.; Jarosz, T.; Radosz, P.; Setlak, M.; Madej, P. In Search of New Therapeutics-Molecular Aspects of the PCOS Pathophysiology: Genetics, Hormones, Metabolism and Beyond. Int. J. Mol. Sci. 2020, 21, 7054. [Google Scholar] [CrossRef] [PubMed]
- Kalhori, Z.; Azadbakht, M.; Soleimani Mehranjani, M.; Shariatzadeh, M.A. Improvement of the folliculogenesis by transplantation of bone marrow mesenchymal stromal cells in mice with induced polycystic ovary syndrome. Cytotherapy 2018, 20, 1445–1458. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Xiong, X.; Xiao, N.; He, K.; Chen, M.; Peng, J.; Su, X.; Mei, H.; Dai, Y.; Wei, D.; et al. Mesenchymal Stem Cells Alleviate DHEA-Induced Polycystic Ovary Syndrome (PCOS) by Inhibiting Inflammation in Mice. Stem Cells Int. 2019, 2019, 9782373. [Google Scholar] [CrossRef] [Green Version]
- Driscoll, J.; Patel, T. The mesenchymal stem cell secretome as an acellular regenerative therapy for liver disease. J. Gastroenterol. 2019, 54, 763–773. [Google Scholar] [CrossRef] [Green Version]
- Jeong, H.; Yim, H.W.; Park, H.J.; Cho, Y.; Hong, H.; Kim, N.J.; Oh, I.H. Mesenchymal Stem Cell Therapy for Ischemic Heart Disease: Systematic Review and Meta-analysis. Int. J. Stem Cells 2018, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Momin, E.N.; Mohyeldin, A.; Zaidi, H.A.; Vela, G.; Quiñones-Hinojosa, A. Mesenchymal stem cells: New approaches for the treatment of neurological diseases. Curr. Stem Cell Res. Ther. 2010, 5, 326–344. [Google Scholar] [CrossRef] [Green Version]
- Moreira, A.; Kahlenberg, S.; Hornsby, P. Therapeutic potential of mesenchymal stem cells for diabetes. J. Mol. Endocrinol. 2017, 59, R109–R120. [Google Scholar] [CrossRef]
- Chapel, A.; Bertho, J.M.; Bensidhoum, M.; Fouillard, L.; Young, R.G.; Frick, J.; Demarquay, C.; Cuvelier, F.; Mathieu, E.; Trompier, F.; et al. Mesenchymal stem cells home to injured tissues when co-infused with hematopoietic cells to treat a radiation-induced multi-organ failure syndrome. J. Gene Med. 2003, 5, 1028–1038. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I. Why are MSCs therapeutic? New data: New insight. J. Pathol. 2009, 217, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Ullah, M.; Liu, D.D.; Thakor, A.S. Mesenchymal Stromal Cell Homing: Mechanisms and Strategies for Improvement. iScience 2019, 15, 421–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, X.L.; Zhang, Y.; Li, X.; Fu, Q.L. Mechanisms underlying the protective effects of mesenchymal stem cell-based therapy. Cell. Mol. Life Sci. 2020, 77, 2771–2794. [Google Scholar] [CrossRef] [Green Version]
- Salgado, A.J.; Reis, R.L.; Sousa, N.J.; Gimble, J.M. Adipose tissue derived stem cells secretome: Soluble factors and their roles in regenerative medicine. Curr. Stem Cell Res. Ther. 2010, 5, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.Z.; Abelson, B.; Babbar, P.; Damaser, M.S. Harnessing the mesenchymal stem cell secretome for regenerative urology. Nat. Rev. Urol. 2019, 16, 363–375. [Google Scholar] [CrossRef]
- Hmadcha, A.; Martin-Montalvo, A.; Gauthier, B.R.; Soria, B.; Capilla-Gonzalez, V. Therapeutic Potential of Mesenchymal Stem Cells for Cancer Therapy. Front. Bioeng. Biotechnol. 2020, 8, 43. [Google Scholar] [CrossRef]
- Harman, R.M.; Marx, C.; Van de Walle, G.R. Translational Animal Models Provide Insight Into Mesenchymal Stromal Cell (MSC) Secretome Therapy. Front. Cell Dev. Biol. 2021, 9, 654885. [Google Scholar] [CrossRef]
- Munshi, A.; Mehic, J.; Creskey, M.; Gobin, J.; Gao, J.; Rigg, E.; Muradia, G.; Luebbert, C.C.; Westwood, C.; Stalker, A.; et al. A comprehensive proteomics profiling identifies NRP1 as a novel identity marker of human bone marrow mesenchymal stromal cell-derived small extracellular vesicles. Stem Cell Res. Ther. 2019, 10, 401. [Google Scholar] [CrossRef] [Green Version]
- Rolandsson Enes, S.; Ahrman, E.; Palani, A.; Hallgren, O.; Bjermer, L.; Malmstrom, A.; Scheding, S.; Malmstrom, J.; Westergren-Thorsson, G. Quantitative proteomic characterization of lung-MSC and bone marrow-MSC using DIA-mass spectrometry. Sci. Rep. 2017, 7, 9316. [Google Scholar] [CrossRef] [Green Version]
- McAllister, J.M.; Han, A.X.; Modi, B.P.; Teves, M.E.; Mavodza, G.R.; Anderson, Z.L.; Shen, T.; Christenson, L.K.; Archer, K.J.; Strauss, J.F. miRNA Profiling Reveals miRNA-130b-3p Mediates DENND1A Variant 2 Expression and Androgen Biosynthesis. Endocrinology 2019, 160, 1964–1981. [Google Scholar] [CrossRef] [PubMed]
- Hecker, M.; Newsted, J.L.; Murphy, M.B.; Higley, E.B.; Jones, P.D.; Wu, R.; Giesy, J.P. Human adrenocarcinoma (H295R) cells for rapid in vitro determination of effects on steroidogenesis: Hormone production. Toxicol. Appl. Pharmacol. 2006, 217, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Marti, N.; Bouchoucha, N.; Sauter, K.S.; Fluck, C.E. Resveratrol inhibits androgen production of human adrenocortical H295R cells by lowering CYP17 and CYP21 expression and activities. PLoS ONE 2017, 12, e0174224. [Google Scholar] [CrossRef] [PubMed]
- Mikhaylova, I.V.; Kuulasmaa, T.; Jaaskelainen, J.; Voutilainen, R. Tumor necrosis factor-alpha regulates steroidogenesis, apoptosis, and cell viability in the human adrenocortical cell line NCI-H295R. Endocrinology 2007, 148, 386–392. [Google Scholar] [CrossRef]
- Nelson, V.L.; Qin, K.N.; Rosenfield, R.L.; Wood, J.R.; Penning, T.M.; Legro, R.S.; Strauss, J.F., 3rd; McAllister, J.M. The biochemical basis for increased testosterone production in theca cells propagated from patients with polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2001, 86, 5925–5933. [Google Scholar] [CrossRef]
- Wickenheisser, J.K.; Quinn, P.G.; Nelson, V.L.; Legro, R.S.; Strauss, J.F., III; McAllister, J.M. Differential Activity of the Cytochrome P450 17α-Hydroxylase and Steroidogenic Acute Regulatory Protein Gene Promoters in Normal and Polycystic Ovary Syndrome Theca Cells1. J. Clin. Endocrinol. Metab. 2000, 85, 2304–2311. [Google Scholar] [CrossRef] [Green Version]
- Chugh, R.M.; Park, H.S.; Esfandyari, S.; Elsharoud, A.; Ulin, M.; Al-Hendy, A. Mesenchymal Stem Cell-Conditioned Media Regulate Steroidogenesis and Inhibit Androgen Secretion in a PCOS Cell Model via BMP-2. Int. J. Mol. Sci. 2021, 22, 9184. [Google Scholar] [CrossRef]
- Chugh, R.M.; Park, H.S.; El Andaloussi, A.; Elsharoud, A.; Esfandyari, S.; Ulin, M.; Bakir, L.; Aboalsoud, A.; Ali, M.; Ashour, D.; et al. Mesenchymal stem cell therapy ameliorates metabolic dysfunction and restores fertility in a PCOS mouse model through interleukin-10. Stem Cell Res. Ther. 2021, 12, 388. [Google Scholar] [CrossRef]
- Cook, H.V.; Doncheva, N.T.; Szklarczyk, D.; von Mering, C.; Jensen, L.J. Viruses.STRING: A Virus-Host Protein-Protein Interaction Database. Viruses 2018, 10, 519. [Google Scholar] [CrossRef] [Green Version]
- Mei, S. In Silico Enhancing M. tuberculosis Protein Interaction Networks in STRING To Predict Drug-Resistance Pathways and Pharmacological Risks. J. Proteome Res. 2018, 17, 1749–1760. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING v10: Protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Jin, J.; Shen, S.; Xia, Y.; Xu, P.; Zou, X.; Wang, H.; Yi, L.; Wang, Y.; Gao, Q. Modulation of expression of 17-Hydroxylase/17,20 lyase (CYP17) and P450 aromatase (CYP19) by inhibition of MEK1 in a human ovarian granulosa-like tumor cell line. Gynecol. Endocrinol. 2016, 32, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Kempna, P.; Hofer, G.; Mullis, P.E.; Fluck, C.E. Pioglitazone inhibits androgen production in NCI-H295R cells by regulating gene expression of CYP17 and HSD3B2. Mol. Pharmacol. 2007, 71, 787–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sewer, M.B.; Waterman, M.R. Adrenocorticotropin/cyclic adenosine 3′,5′-monophosphate-mediated transcription of the human CYP17 gene in the adrenal cortex is dependent on phosphatase activity. Endocrinology 2002, 143, 1769–1777. [Google Scholar] [CrossRef]
- Dilogo, I.H.; Fiolin, J.; Aprianto, P. Osteogenic Potency of Secretome Bone Marrow Derived Mesenchymal Stem Cells: A Literature Review. Adv. Sci. Lett. 2018, 24, 6206–6208. [Google Scholar] [CrossRef]
- Polacek, M.; Bruun, J.A.; Elvenes, J.; Figenschau, Y.; Martinez, I. The secretory profiles of cultured human articular chondrocytes and mesenchymal stem cells: Implications for autologous cell transplantation strategies. Cell Transpl. 2011, 20, 1381–1393. [Google Scholar] [CrossRef]
- Nourian Dehkordi, A.; Mirahmadi Babaheydari, F.; Chehelgerdi, M.; Raeisi Dehkordi, S. Skin tissue engineering: Wound healing based on stem-cell-based therapeutic strategies. Stem Cell Res. Ther. 2019, 10, 111. [Google Scholar] [CrossRef] [Green Version]
- Soleymaninejadian, E.; Pramanik, K.; Samadian, E. Immunomodulatory properties of mesenchymal stem cells: Cytokines and factors. Am. J. Reprod. Immunol. 2012, 67, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Yang, R.; Biswas, S.; Qin, X.; Zhang, M.; Deng, W. Mesenchymal stem cells and induced pluripotent stem cells as therapies for multiple sclerosis. Int. J. Mol. Sci. 2015, 16, 9283–9302. [Google Scholar] [CrossRef] [Green Version]
- Schaefer, C.F.; Anthony, K.; Krupa, S.; Buchoff, J.; Day, M.; Hannay, T.; Buetow, K.H. PID: The Pathway Interaction Database. Nucleic Acids Res. 2009, 37, D674–D679. [Google Scholar] [CrossRef]
- van Bilsen, J.H.M.; van den Brink, W.; van den Hoek, A.M.; Dulos, R.; Caspers, M.P.M.; Kleemann, R.; Wopereis, S.; Verschuren, L. Mechanism-Based Biomarker Prediction for Low-Grade Inflammation in Liver and Adipose Tissue. Front. Physiol. 2021, 12, 703370. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.H.; Lee, W.S.; Won, M.; Park, M.; Park, H.O.; Kim, E.; Lee, K.A.; Bae, J. The apolipoprotein A-I level is downregulated in the granulosa cells of patients with polycystic ovary syndrome and affects steroidogenesis. J. Proteome Res. 2010, 9, 4329–4336. [Google Scholar] [CrossRef] [PubMed]
- Hovingh, G.K.; Brownlie, A.; Bisoendial, R.J.; Dube, M.P.; Levels, J.H.; Petersen, W.; Dullaart, R.P.; Stroes, E.S.; Zwinderman, A.H.; de Groot, E.; et al. A novel apoA-I mutation (L178P) leads to endothelial dysfunction, increased arterial wall thickness, and premature coronary artery disease. J. Am. Coll. Cardiol. 2004, 44, 1429–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tobin, M.K.; Stephen, T.K.L.; Lopez, K.L.; Pergande, M.R.; Bartholomew, A.M.; Cologna, S.M.; Lazarov, O. Activated Mesenchymal Stem Cells Induce Recovery Following Stroke Via Regulation of Inflammation and Oligodendrogenesis. J. Am. Heart Assoc. 2020, 9, e013583. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | |
|---|---|
| 1 | Actin, cytoplasmic 1 OS = Homo sapiens OX = 9606 GN = ACTB PE = 1 SV = 1 |
| 2 | Alpha-1B-glycoprotein OS = Homo sapiens OX = 9606 GN = A1BG PE = 1 SV = 4 |
| 3 | Alpha-2-HS-glycoprotein OS = Homo sapiens OX = 9606 GN = AHSG PE = 1 SV = 2 |
| 4 | Apolipoprotein A-I OS = Homo sapiens OX = 9606 GN = APOA1 PE = 1 SV = 1 |
| 5 | Beta-2-glycoprotein 1 OS = Homo sapiens OX = 9606 GN = APOH PE = 1 SV = 3 |
| 6 | Collagen alpha-1(I) chain OS = Homo sapiens OX = 9606 GN = COL1A1 PE = 1 SV = 5 |
| 7 | Collagen alpha-2(I) chain OS = Homo sapiens OX = 9606 GN = COL1A2 PE = 1 SV = 7 |
| 8 | Fibrillin-2 OS = Homo sapiens OX = 9606 GN = FBN2 PE = 1 SV = 3 |
| 9 | Fibronectin OS = Homo sapiens OX = 9606 GN = FN1 PE = 1 SV = 4 |
| 10 | Fibulin-1 OS = Homo sapiens OX = 9606 GN = FBLN1 PE = 1 SV = 4 |
| 11 | Fibulin-5 OS = Homo sapiens OX = 9606 GN = FBLN5 PE = 1 SV = 1 |
| 12 | Haptoglobin OS = Homo sapiens OX = 9606 GN = HP PE = 1 SV = 1 |
| 13 | Haptoglobin-related protein OS = Homo sapiens OX = 9606 GN = HPR PE = 2 SV = 2 |
| 14 | Hemoglobin subunit alpha OS = Homo sapiens OX = 9606 GN = HBA1 PE = 1 SV = 2 |
| 15 | Hemopexin OS = Homo sapiens OX = 9606 GN = HPX PE = 1 SV = 2 |
| 16 | Immunoglobulin heavy constant gamma 3 OS = Homo sapiens OX = 9606 GN = IGHG3 PE = 1 SV = 2 |
| 17 | Insulin-like growth factor-binding protein 4 OS = Homo sapiens OX = 9606 GN = IGFBP4 PE = 1 SV = 2 |
| 18 | Insulin-like growth factor-binding protein 6 OS = Homo sapiens OX = 9606 GN = IGFBP6 PE = 1 SV = 1 |
| 19 | Inter-alpha-trypsin inhibitor heavy chain H4 OS = Homo sapiens OX = 9606 GN = ITIH4 PE = 1 SV = 4 |
| 20 | Isoform LMW of Kininogen-1 OS = Homo sapiens OX = 9606 GN = KNG1 |
| 21 | Latent-transforming growth factor beta-binding protein 2 OS = Homo sapiens OX = 9606 GN = LTBP2 PE = 1 SV = 3 |
| 22 | Metalloproteinase inhibitor 1 OS = Homo sapiens OX = 9606 GN = TIMP1 PE = 1 SV = 1 |
| 23 | N-acetylmuramoyl-L-alanine amidase OS = Homo sapiens OX = 9606 GN = PGLYRP2 PE = 1 SV = 1 |
| 24 | Serotransferrin OS = Homo sapiens OX = 9606 GN = TF PE = 1 SV = 3 |
| 25 | Serum albumin OS = Homo sapiens OX = 9606 GN = ALB PE = 1 SV = 2 |
| 26 | Thrombospondin-2 OS = Homo sapiens OX = 9606 GN = THBS2 PE = 1 SV = 2 |
| Description | |
|---|---|
| 1 | Actin, cytoplasmic 1 OS = Homo sapiens OX = 9606 GN = ACTB PE = 1 SV = 1 |
| 2 | Afamin OS = Homo sapiens OX = 9606 GN = AFM PE = 1 SV = 1 |
| 3 | Alpha-1B-glycoprotein OS = Homo sapiens OX = 9606 GN = A1BG PE = 1 SV = 4 |
| 4 | Alpha-2-HS-glycoprotein OS = Homo sapiens OX = 9606 GN = AHSG PE = 1 SV = 2 |
| 5 | Apolipoprotein A-I OS = Homo sapiens OX = 9606 GN = APOA1 PE = 1 SV = 1 |
| 6 | Clusterin OS = Homo sapiens OX = 9606 GN = CLU PE = 1 SV = 1 |
| 7 | Collagen alpha-1(I) chain OS = Homo sapiens OX = 9606 GN = COL1A1 PE = 1 SV = 5 |
| 8 | Collagen alpha-1(III) chain OS = Homo sapiens OX = 9606 GN = COL3A1 PE = 1 SV = 4 |
| 9 | Collagen alpha-1(VI) chain OS = Homo sapiens OX = 9606 GN = COL6A1 PE = 1 SV = 3 |
| 10 | Collagen alpha-2(I) chain OS = Homo sapiens OX = 9606 GN = COL1A2 PE = 1 SV = 7 |
| 11 | Collagen alpha-2(V) chain OS = Homo sapiens OX = 9606 GN = COL5A2 PE = 1 SV = 3 |
| 12 | Decorin OS = Homo sapiens OX = 9606 GN = DCN PE = 1 SV = 1 |
| 13 | Fibronectin OS = Homo sapiens OX = 9606 GN = FN1 PE = 1 SV = 4 |
| 14 | Growth arrest-specific protein 6 OS = Homo sapiens OX = 9606 GN = GAS6 PE = 1 SV = 3 |
| 15 | Haptoglobin OS = Homo sapiens OX = 9606 GN = HP PE = 1 SV = 1 |
| 16 | Hemopexin OS = Homo sapiens OX = 9606 GN = HPX PE = 1 SV = 2 |
| 17 | Inter-alpha-trypsin inhibitor heavy chain H4 OS = Homo sapiens OX = 9606 GN = ITIH4 PE = 1 SV = 4 |
| 18 | N-acetylmuramoyl-L-alanine amidase OS = Homo sapiens OX = 9606 GN = PGLYRP2 PE = 1 SV = 1 |
| 19 | Serotransferrin OS = Homo sapiens OX = 9606 GN = TF PE = 1 SV = 3 |
| 20 | Serum albumin OS = Homo sapiens OX = 9606 GN = ALB PE = 1 SV = 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, H.-S.; Chugh, R.M.; Pergande, M.R.; Cetin, E.; Siblini, H.; Esfandyari, S.; Cologna, S.M.; Al-Hendy, A. Non-Cytokine Protein Profile of the Mesenchymal Stem Cell Secretome That Regulates the Androgen Production Pathway. Int. J. Mol. Sci. 2022, 23, 4633. https://doi.org/10.3390/ijms23094633
Park H-S, Chugh RM, Pergande MR, Cetin E, Siblini H, Esfandyari S, Cologna SM, Al-Hendy A. Non-Cytokine Protein Profile of the Mesenchymal Stem Cell Secretome That Regulates the Androgen Production Pathway. International Journal of Molecular Sciences. 2022; 23(9):4633. https://doi.org/10.3390/ijms23094633
Chicago/Turabian StylePark, Hang-Soo, Rishi Man Chugh, Melissa R. Pergande, Esra Cetin, Hiba Siblini, Sahar Esfandyari, Stephanie M. Cologna, and Ayman Al-Hendy. 2022. "Non-Cytokine Protein Profile of the Mesenchymal Stem Cell Secretome That Regulates the Androgen Production Pathway" International Journal of Molecular Sciences 23, no. 9: 4633. https://doi.org/10.3390/ijms23094633