Identifying the Therapeutic Significance of Mesenchymal Stem Cells
by
 ,
,
Vineet Kumar Mishra
1,†, 
Hui-Hsuan Shih
2,3,†,
Farzana Parveen
1,
David Lenzen
1,
Etsuro Ito
4,5,6 ,
,
Te-Fu Chan
4,7,* and
Liang-Yin Ke
1,2,4,* 1
Department of Medical Laboratory Science and Biotechnology, College of Health Sciences, Kaohsiung Medical University, Kaohsiung 807378, Taiwan
2
Center for Lipid Biosciences, Kaohsiung Medical University Hospital & Lipid Science and Aging Research Center, Kaohsiung Medical University, Kaohsiung 807377, Taiwan
3
Division of Chest Medicine, Department of International Medicine, MacKay Memorial Hospital, Taipei 104217, Taiwan
4
Graduate Institute of Medicine, College of Medicine & Drug Development and Value Creation Research Center, Kaohsiung Medical University, Kaohsiung 807378, Taiwan
5
Department of Biology, Waseda University, Tokyo 162-8480, Japan
6
Waseda Research Institute for Science and Engineering, Waseda University, Tokyo 162-8480, Japan
7
Department of Obstetrics and Gynecology, Kaohsiung Medical University Hospital, Kaohsiung 807377, Taiwan
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Cells 2020, 9(5), 1145; https://doi.org/10.3390/cells9051145
Submission received: 13 March 2020
/
Revised: 4 May 2020
/
Accepted: 5 May 2020
/
Published: 6 May 2020
(This article belongs to the Collection Stem Cell Therapies for Treating Diabetes)
Abstract
:The pleiotropic behavior of mesenchymal stem cells (MSCs) has gained global attention due to their immense potential for immunosuppression and their therapeutic role in immune disorders. MSCs migrate towards inflamed microenvironments, produce anti-inflammatory cytokines and conceal themselves from the innate immune system. These signatures are the reason for the uprising in the sciences of cellular therapy in the last decades. Irrespective of their therapeutic role in immune disorders, some factors limit beneficial effects such as inconsistency of cell characteristics, erratic protocols, deviating dosages, and diverse transfusion patterns. Conclusive protocols for cell culture, differentiation, expansion, and cryopreservation of MSCs are of the utmost importance for a better understanding of MSCs in therapeutic applications. In this review, we address the immunomodulatory properties and immunosuppressive actions of MSCs. Also, we sum up the results of the enhancement, utilization, and therapeutic responses of MSCs in treating inflammatory diseases, metabolic disorders and diabetes.
1. Introduction
MSCs are Mesenchymal Stem Cells (MSCs), which can be defined as non-hematopoietic multipotent stem cells with the ability to differentiate into mesodermal lineage (adipocytes, osteocytes and chondrocytes), ectodermal lineage (neurocytes) and endodermal lineage (hepatocytes) [1,2]. In 1966, Friedenstein and his team discovered the multipotent behavior of MSCs for the first time [3]. After many years, the term MSCs was coined by Caplan et al. in 1991 [4]. Since then, MSCs have become a well-known and outstanding cell source alluring for clinical applications. They have an excellent capacity for self-renewal in vitro, lasting for more than four months [5].
Earlier, there was a debate among scientists about the stemness and nomenclature of MSCs. Some articles preferred mesenchymal “stromal” cells instead of stem cells [6]. Some researchers attempted to change the name of MSCs to medicinal signaling cells because of their secretory role in the locations of diseases, injuries, and inflammations [7,8]. However, later reports showed that prostaglandin E2 (PGE2) secreted by MSCs is responsible for maintaining the self-renewal ability and PGE2 is also involved in the immunomodulation of MSCs, creating a cascade of events, which proves the stemness of MSCs [9]. Hence, the term Mesenchymal Stem Cells is justified.
There are various sources of isolation of MSCs such as adipose tissue, bone marrow, peripheral blood and neonatal tissues (umbilical cord, placenta, amniotic fluid, and amniotic membrane) [2,10,11,12]. The source of isolation of MSCs greatly affects the yield, the expressed variety of surface markers and cytokine profile [13,14,15]. However, the basic identification markers of MSCs are CD73 (cluster of differentiation 73), CD105, CD90 but they should be CD34-, CD14-, CD45-, CD11b-, CD19- and lack HLAII (Human Leukocyte Antigen complex 2) [16]. Besides that, MSCs must express transcription factors such as octamer-binding transcription factor 4 (OCT-4) and homeobox protein NANOG [17].
Currently, there is a lack of standardized methods for the isolation and culture of MSCs [18]. Comparative studies are challenging due to MSCs showing different features depending on their source and microenvironment from which they are isolated [19,20,21]. Several researchers focused on optimizing the isolation of MSCs. Talwadekar et al. found out that the clonogenicity and function of placenta MSCs (P-MSCs) were superior to cord MSCs (C-MSCs) [22]. Similarly, Nagaishi et al. showed that MSCs from umbilical cord significantly improve diabetic abnormalities and diabetic nephropathy in comparison to the bone marrow-derived MSCs [23]. Therefore, the source of MSC isolation is critical in context with its potential and efficacy towards its properties [2,23].
In this review, we aim to discuss the immunomodulatory properties and immunosuppressive actions of MSCs. Besides that, we sum up the results of the enhancement, utilization, and therapeutic responses of MSCs in treating human diseases, and particularly their potential towards diabetes and adipose tissue dysfunction.
2. The Immunological Functions of Mesenchymal Stem Cells
The primary functions of MSCs are: (1) immunomodulation, (2) autocrine and paracrine activities, and (3) evasion of innate immunity. Below, we discuss every aspect in detail.
2.1. Immunomodulation
The mode of immunomodulation is mediated by cell-cell interactions, cytokines and soluble factors. Note that, depending on the levels of stimulation, MSCs play either pro-inflammatory or anti-inflammatory roles in its microenvironment (Figure 1) [13,15,24,25].
2.1.1. Pro-Inflammation
At low levels of interferon-gamma (IFN-γ) and tumor necrosis factor-α (TNF-α), MSCs show the pro-inflammatory phenotype (Figure 1, red arrows). MSCs produce chemokines such as macrophage inflammatory protein-1α/β (MIP-1α/β), RANTES, chemokine (C-X-C motif) ligand 9 (CXCL9), and CXCL10 within its microenvironment to activate the T cells [26]. MSCs secreting PGE-2 severely hamper dendritic cell (DC) precursors during the process of differentiation and maturation [27,28]. In the absence of interleukin-6 (IL-6), MSCs promote proliferation and activation of M1 macrophage [29]. The transition from M0 to pro-inflammatory M1 macrophage is induced by secretion of interferon-γ (IFN-γ) and IL-1, along with surface protein expression of CD40 ligand (CD40L). Further, these M1 macrophages express IFN-γ and tumor necrosis factor-α (TNF-α) within the microenvironment along with the co-stimulatory surface molecules to respond to T cell activation [29,30]. In contrast, in an anti-inflammatory microenvironment, this phenomenon can be inhibited by TNF-stimulated gene 6 protein (TSG-6) [31]. The feedback mechanism studied under in-vitro conditions of pro-inflammatory cytokines make MSCs enhance the immune response by producing chemokines CXCL-9, CXCL-10, and CXCL-11. This leads to more neutrophils moving towards the site of inflammation where they act mainly by phagocytosis [26,32]. The point to be noticed here is that human and mice derived MSCs have these effects only when exposed to low or insufficient levels of pro-inflammatory cytokines such as IFN-γ and TNF-α.
2.1.2. Anti-Inflammation
In the anti-inflammatory condition, MSCs suppress the immune response in inflammatory cytokine-rich microenvironments, such as wounds, infections, or organ transplantations. These immunosuppressive indications were observed in humans [33,34,35,36,37], baboons [37], and murine [38], where MSCs successfully inhibited T lymphocyte activation and proliferation (Figure 1, green arrows). This particular behavior of MSCs in the presence and absence of inflammatory molecules is called MSC polarization [29]. Under high levels of IFN-γ and TNF-α, MSCs produce cytokines such as transforming growth factor-β (TGF-β), hepatocyte growth factor (HGF) [39], and secrete soluble factors such as indoleamine 2,3-dioxygenase (IDO), PGE2 and nitric oxide (NO) [40]. These factors directly promote the activation of regulatory T cells (Tregs) (CD4+, CD25+, forkhead box P3 (FOXP3+)). Besides, in responding to IL-6 stimulation, MSCs secrete TGF-β and PGE2 again to induce Treg cell activation.
MSCs can also promote the activation of T-reg cells indirectly. Recently Heo et al. described the M2 macrophage stimulation and phenotype changes by the exosomes derived from AD-MSCs in an in- vitro study [41]. Furthermore, these M2 macrophages (MSC stimulated) express CCL-18 and activate Tregs cells [42]. The process of immunosuppression is also dependent on factors released by MSCs or in the microenvironment. MSCs trigger the expression of cyclooxygenase 2 (COX2) and IDO and further promote homeostatic response towards macrophage polarization [15,43]. In that response, M2 macrophage expresses CD206 and CD163 co-stimulatory molecules, along with enriching the microenvironment by IL-6 and IL-10 expression [44]. The excessive IL-10 production by DCs and M2 upon MSCs co-culture further boosts the immunosuppression by suppressing effector T cells [45,46].
MSCs trigger the proliferation, activation and immunoglobulin G (IgG) secretion of B cells by IDO [47,48,49]. These again inhibit other T-effector cells to support anti-inflammation (Figure 1) [29,44,50]. Human- and mouse-derived BM-MSCs release specific chemokine ligands such as C-C motif chemokine ligand 2 (CCL2), CCL3 and CCL12 which help them to work with monocytes and macrophages in tissue homeostasis and wound healing [51].
2.2. Autocrine and Paracrine Role of MSC Secreted Molecules
MSCs secrete biomolecules such as growth factors, cytokines and chemokines which help their biological activities in the autocrine or paracrine manner in consonance to the encompassing microenvironment [52,53].
A typical autocrine manner of MSCs for maintaining self-renewal capacity is through PGE2 signaling [9] (Figure 2). PGE2 belongs to the prostaglandin family. It plays a role in lipid mediation and other physiological effects. For synthesizing PGE2, the conversion of arachidonic acid to prostaglandin H2 (PGH2) is regulated by COX-2, a prostaglandin-endoperoxidase synthase; and the isomerization of PGH2 to PGE2 is induced by PGE2 synthase [54]. COX-2 is the rate-limiting enzyme to mediate inflammatory cytokines, growth factors and tumor promoters [55,56]. MSCs secrete PGE2 to the extracellular environment by multidrug-resistant protein 4 (MRP4), later it binds to the PGE2 receptors on target cells [57]. PGE2 receptor 2 (EP2) augments cell proliferation and neovascularization by advancing the secretion of vascular endothelial growth factors (VEGF) [55,58,59]. By reviewing considerable amounts of in vitro and in vivo studies of immune disorders, we can say that COX-2 mediated PGE2 expression by MSCs are pivotal factors for the immunomodulatory ability of MSCs [60,61,62,63,64]. The loop between COX-2 and PGE2 maintains an axis that influences cell-cycle, cell-proliferation and cell viability by invigoration of one or more EP receptors [65].
MSCs are very competent in suppressing and tolerating the immune system via cell-cell contacts and soluble factors production. The migratory ability of MSCs toward inflammatory signals and damaged tissues makes them an excellent vector for therapeutics. Gnecchi et al. characterized MSCs and demonstrated the paracrine mechanism by genetically engineered protein kinase B (PKB, also known as Akt)-MSCs [66,67] (Figure 2). In this in vitro experiment, Akt-MSCs-derived conditioned medium collected from hypoxic Akt-MSCs was found to be efficiently protective to cardiomyocytes by inhibiting the apoptosis. The paracrine effect of MSCs became clearer when they found the effect of other valuable factors such as VEGF, bFGF (basic fibroblast growth factors), HGF and thymosin β-4 (TB4).
The paracrine mechanism plays a major role in immunomodulation, where MSCs intensify their action in localized lymphocytes by the factors involving TGF-β, HGF, PGE2, human leukocyte antigen-G5 (HLAG-5), IL-6, CCL-2, CCL-5 and some other chemokines [68,69,70,71]. In the mammalian immune system, DCs are considered to be one of the most important antigen-presenting cells (APCs). However, due to the effects of MSCs, immature DC precursors (CD34+ precursors) ceased at the differentiation [72,73,74]. PGE2 from MSCs has been reported for its tolerogenic features, such as IL-10 release, IDO1 expression, also when merged with TNF-α, IL-1β, and IL6, it intensifies the immunogenicity with the co-stimulatory molecules [75,76,77]. In the vicinity of microenvironments treated with MSCs, mature DCs (CD83+ DCs with CD80 and CD86 costimulatory molecules) get obstructed in their efficiency for T cell activation [74,78]. Together, all these factors and chemokines generate tolerance against the immune cells and help MSCs to facilitate favorable therapeutic roles.
2.3. Escape Mechanism of MSCs From the Innate Immune System
MSCs escape from the immune system by the cell-cell interactions through the production of immune regulatory molecules such as IFN-γ, COX 2, PGE2 and IDO [26,79,80]. Among these factors, IFN-γ plays a crucial role in inducing IDO expression and tryptophan depletion for the escape mechanism [79]. In addition, HGF and TGF-β support MSCs to develop resistance against immune cells [81]. Meisel et al. provided comprehensive in vitro results, where the T cell proliferation was partly restored after adding monoclonal antibodies against TGF-β or HGF to show that both cytokines are participating in this interplay [82].
Previously, both autologous and allogeneic MSCs were killed by activated natural killer cells (NK cells) due to surface receptors such as MHC class I-related chain A (MIC-A), UL16-binding protein (ULBPs), poliovirus receptor and nectin-2 [83,84]. However, IFN-γ treated MSCs are less likely to be killed by NK cells [84]. MSCs inhibit their cytokines production, cytotoxicity, and cell proliferation through PGE2 and IDO [85,86]. In a nutshell, there are three large and extensive laps of the immune response: The first is antigen recognition and presentation, the second is T cell activation, proliferation and differentiation and the third is the effective stage. The escape mechanism of MSCs can be seen throughout all the three stages of immune responses.
2.4. Complement System and MSCs
The complement system of a body is the first line of defense against pathogenic foreign invaders [87]. There are more than 30 proteins involved in the interplay of human complement system during an inflammatory response [88]. The three humanistic pathways by which the complement system activates are the classical, lectin and alternative pathway. The classical pathway involves complement component 1q (C1q) which is activated by antibodies. The lectin pathway becomes activated by carbohydrate moieties, and the alternative pathway is stimulated by hydrolysis of complement component 3 (C3). The ultimate result of this activation is to form a membrane attack complex (MAC) for the initiation of cell lysis [89].
2.4.1. Complement System Attack on MSCs
It is well documented that during an infusion of MSCs to study clinical approaches, MSCs are prone to attract the complements in the blood and suffer damage caused by MACs. Injured MSCs are more vulnerable to further immune responses and also get compromised in their potent functionality [90]. Receptors present on monocytes such as CR3 (complement receptor 3) assist in phagocytosis of complement opsonized MSCs [91]. In some MSC-infusions in vivo experiments, a particular suppressive cell population has emerged along with M2 monocytes, observed to decrease the MSC population [15,92,93,94]. The interactions of MSCs with blood plasma require more extensive research to find the reason behind MSC depletion after infusion. Recently, Gavin et al. demonstrated the C3 mediated complement phagocytosis of MSCs by monocytes with markers CD14+CD16- excluding the involvement of C5 complex [95]. Such examples may answer the question of depletion in the population of MSC after infusion.
2.4.2. MSCs’ Ability to Counteract the Complement System
The MSC has surface receptors C3aR (C3a receptor) and C5aR (C5a receptor). During a surrounding inflammatory action in the presence of C3a and C5a, MSCs express them on their surface and bind to generate resistance against oxidative stress and apoptosis-inducing mechanisms [96]. Additionally, MSCs have surface expression of CD46, CD55 and CD59, which protect MSCs against the complement action and consequently prevent cell death [96,97]. Although, if the process of cell lysis is initiated somehow on an MSC by the complement system, then it is almost impossible to stop the killing [90].
Multiple external factors have been studied to improve the allogenic survival of MSCs and counteraction against the complement system. Scientists have used anti-C5 antibodies treatment to MSCs before the infusion, as well as the transfection of recombinant adenovirus for the specific overexpression of CD55 inhibitor [90]. Li et al. recognized MSCs propagated antigen in vitro, which is a naturally occurring antibody and has the potential to activate the host complement response. In addition, they demonstrated a simple and economical method to generate heparin-coated MSCs to inhibit the complement system within microenvironments [98]. Besides that, the combined pretreatment of MSCs with IFN-γ and TNF-α has proven to be the best method to inhibit the complement system. This treatment resulted in the secretion of factor H, which is the primary complement inhibitor produced by MSCs [97]. We will discuss this pretreatment application in detail later in this review.
3. Therapeutic Role of Modified MSCs
3.1. Unveiled the Role of MSCs in Therapeutics
MSCs have become an alluring topic of research due to their role in modern-day therapeutics. Recently, a new therapeutic paradigm has emerged using MSC-derived exosomes and modified MSCs. The exosomes (extracellular vesicles) are rich in lipids, proteins, mRNAs, tRNAs, long non-coding RNAs, micro-RNAs as well as mitochondrial DNA, which is transferred between cells in both near and distant vicinities [99,100]. Research evidence has shown that MSC-derived exosomes exert helpful effects on different disease models such as myocardial infarction [101,102,103,104], hepatic fibrosis [105,106,107] and cancer [108] [107,109,110,111,112]. Additionally, the modified MSCs strive in their effects with the inflammatory factors and the microenvironment of MSCs, which are responsible for their phenotypic effects being exerted on the immune system [113].
Inflammation makes MSCs secrete molecules such as IL-10, galectins, IDO, and PGE2, heme oxygenase-1 (HO-1), TNF-inducible gene-6 protein (TSG6), chemokine ligand-2 (CCL2) and NO which are responsible for tissue homeostasis [114]. In addition to that, HLAG-5 secreted by MSCs has the ability to interact with allo-stimulated T cells which assists them to suppress T-lymphocytes, NK cells and regulatory T cells (CD4+CD25highFOXP3+ T cells) [115]. These effects were analyzed by comparing the secretion patterns among molecules of MSCs from murine and humans. Their results showed that IDO and HLA-G are the key effectors from human, whereas NO was from murine MSCs. Also, for inducing the immune response, it was IFN-γ in humans for induction of IDO, but both IFN-γ and TNF-α contributed to iNOS (inducible NO Synthase)/NO induction in murine [29].
Chinnadurai et al. studied this immunosuppression by analyzing basal and inducible MSCs secretory molecular patterns, which are responsible for the suppression of T cell proliferation. They identified IDO as a definitive enzyme, which plays a dominant role in MSC-mediated inhibition of T cell proliferation. Furthermore, the blocking of VEGF, granulocyte-colony stimulating factor (GCSF), CXCL9, CXCL10, IFN-α, CCL2 and IL-7 failed to inhibit MSC’s effect on blocking the T cell proliferation. However, this correlation was lost in frozen-thawed MSCs [116]. The potency of MSCs is only possible after some modifications prior to them being used therapeutically. This was shown by Kim and Jang et al. who experimented with IFN-γ primed MSCs to study their influence on IDO activity for progressive inhibition of cell-mediated immunity in graft-versus-host disease (GvHD) [116,117].
3.2. MSCs Priming and Treatment
Modified or treated MSCs found to be enhanced in their immunomodulatory effects on the immune system. IFN-γ is a key regulatory cytokine due to which MSCs are privileged for immunosuppressive functions [118]. Further modifications of MSCs such as homing at targeted sites can promote not only the escape, but also help in the migration of MSCs towards secondary lymphoid organs. The expression of C-C chemokine receptor type 7 (CCR7) gene after bioengineering has shown enhanced immunomodulation and tolerance [119]. The expression of GCSF, CXCL9, IL-7, and CCL2 by MSCs during the interaction with activated peripheral blood mononuclear cells (PBMCs) explains the mechanism of immunosuppression of MSCs. Some reports also showed that the secretion of VEGF is only possible with fresh MSCs, otherwise, there was a complete loss of VEGF production and T cell suppression activity. Such a case was studied with a comparison between fresh and frozen-thawed MSCs [116].
Previously, glucocorticoids, budesonide or dexamethasone-treated MSCs have shown IFN-γ stimulatory effects. The treatments lead to an enhancement of therapeutic potential of MSCs by inhibiting active inflammatory cytokines and raising tolerance towards GvHD and Crohn’s disease [120,121,122,123,124]. In 2010, Dey et al. found beneficial effects by treating mice with genetically engineered MSCs in cases of Huntington disease. MSCs from brain-derived neurotrophic factor (BDNF) or nerve growth factor (NGF) transgenic mice could create microenvironments in the striatum, which ultimately slowed the neurodegenerative process [125]. In 2013, Kwon showed TNF-α-priming MSCs may manifest the inhibition of tissue necrosis along with the promotion of endothelial progenitor cells homing and angiogenesis in the ischemic hind limb animal models [126]. Recently, Kim et al. showed that IFN-γ-priming of human MSCs resulted in the enhancement of immunosuppressive properties. In contrast, treatment with anti-IFN-γ antibody impairs the properties [117]. Hence, the priming of MSCs with different factors opens a wide spectrum for its therapeutic applicability. A brief update for the sites and sources for mesenchymal stem cells used as a therapeutic tool in various models is given in Table 1.
3.3. Outcome of Modified MSCs: Positive and Negative Aspects
The immunosuppressive and immunomodulatory action potential of MSCs has made them a double-edged sword, which can act favorable as well as against the therapeutics. In 2018, Wang et al. showed that IL-35 gene-modified MSCs exhibited better protective effects on Concanavalin A (Con A)-induced autoimmune hepatitis. IL-35 is required for the regulatory and suppressive functions of Tregs. By a gene-delivery vehicle, IL-35-expressing MSCs decrease IFN-γ and Fas ligand (FasL) levels in mononuclear cells through the Janus kinase 1 (JAK1)-signal transducer and activator of transcription 1 (STAT1)/STAT4 signal pathway and eventually inhibit the hepatocyte apoptosis [142].
The behavior of MSCs towards and within its microenvironment is highly complex and needs further research. One of many versatilities was reported by Galland et al., when the group correlated the immunomodulatory effect on NK cells between intra-tumor (T) and adjacent non-tumor tissue (N)-extracted MSCs. The results were astonishing, as they found out that tonsil-derived mesenchymal stem cells (T-MSCs) were more potent immunosuppressive agents as compared to N-MSCs. T-MSCs showed dominance in affecting the NK cells function and phenotype as confirmed by CD56 expression. Upon detailed insight observation, they concluded that tumor-derived MSCs have a definite mechanistic pathway to block the activity of NK cell subsets [145]. The microenvironment of MSCs treatment affects NK cells very deeply and easily. This may be the reason why these cells act favorably depending upon the microenvironment.
In contradiction with this property, Fregni et al. reported that the tumor microenvironment of MSCs implies some of the selected genes to overexpress and promotes metastasis in the case of lung cancer-derived MSCs. Here, the overexpressed genes were found out to be tumor-initiating markers and progressive towards metastasis [146]. MSCs can be miscreant due to their immune-microenvironment modulatory property, one recent example was the study of MSCs residing in the tumor microenvironment, where they developed therapy resistance in tumor cells [147]. However, these effects were influenced by gemcitabine which made them secrete CXCL10, consequently activating the CXCL10-CXCR3 axis in tumor-initiating cells. Hence, MSCs should not be held responsible for resistance in the chemotherapy of pancreatic adenocarcinomas [147] as the results discussed here are influenced by many factors. Therefore, we believe that MSCs could work against the favorable therapeutics, if not applied with prior extensive research.
4. Role of MSCs in Adipocyte Vicinity
It is well known that high-fat diets can induce obesity which represents the risk factors for the development of insulin resistance (IR) and type 2 diabetes (T2DM) [148,149]. Obesity-related diseases such as T2DM induce diabetic wounds and are also associated with rapid cartilage loss and osteoarthritis [150,151,152]. Among the sources of MSCs, adipose tissue is the preferred provider of adipose-derived mesenchymal stem cells (AD-MSCs) [153]. There has been an increase in interest for its therapy potential toward wound healing, tissue engineering and hepatocellular carcinoma [154,155].
4.1. Endocrine Function of Adipose Tissue and MSCs Within
Adipose tissue plays an important role in continuing optimal lipid and glucose homeostasis [156]. The adipocytes possess an endocrine system that helps them to alter metabolism known as adipokines [157]. Before going into the details, we would like to shed some light on the role of TGF-β/BMP (bone morphogenic protein) signaling towards the adipogenic and osteogenic differentiation. Both TGFβ and BMPs are recognized to possess dual differentiation function with MSCs and it has also been observed that upon TGF-β/BMP stimulation, the overall expression of runt-related gene 2 (Runx2/Cbfa1) and peroxisome proliferator-activated receptor-γ (PPAR-γ) can be regulated [158]. Recently, studies have shown BMP2 to be forcing MSCs towards both adipogenic or osteogenic differentiation, while TGF-β was found to be inhibiting this development [159,160].
In Figure 3, we characterized the important contributions of MSCs in adipocyte vicinity by undergoing the process of adipogenic differentiation. Previously, research revealed that BMP2- and BMP4-expressing cells in cultured fibroblasts are committed towards adipogenesis [161,162,163]. PPAR-γ plays an imperative role in adipocyte differentiation during adipogenesis in vitro [164] as well as in vivo [165,166,167]. During adipogenesis, MSCs start expressing transcription factor ZFP423 (Zinc finger protein 423) for committing to pre-adipocyte lineage and the produce regulators PPAR-γ and C/EBP-α/β (transcription co-activators CCAAT/enhancer-binding protein α and β) for differentiation and maturation [168,169,170]. Hence, MSCs residing in the adipose tissue contribute towards healthy mass storage by their differentiation into adipocytes.
4.2. Loss of Potency of MSCs
There are suggestions from many teams that dysfunctional adipocytes in obese or diabetic patients are due to pro-inflammatory cytokines [171,172,173,174]. In our previous work, understanding the mechanism of electronegative low-density lipoprotein (L5 LDL) involved adipose tissue inflammation; we observed that atherogenic lipid deposition and excessive hypertrophy resulted in macrophage infiltration and adipose tissue dysfunction [175]. Figure 4 highlights the hallmarks of adipose tissue dysfunction. In the case of overnutrition, the storage of extra calories in adipose tissue is needed, and to accommodate extra calories, either adipose tissue needs to expand (hypertrophy) or increase its number (hyperplasia). The excessive fat overload leads to an increase in serum LDL and VLDL, which further promotes atherogenic lectin-like oxidized low-density lipoprotein receptor-1 (LOX-1) overexpression, these interactions result in variable postliminary processes depending upon microenvironment conditions and type of cells [176]. The proposed hypothesis in Figure 4 suggests the possible links of a feedback loop mechanism of adipocyte dysfunction which promotes adipocyte hypertrophy and dysfunction by inhibiting hyperplasia.
Adipocyte hyperplasia or hypertrophy occurs in an operative state by a distinct secretory pattern of adipokines of these cells [177,178]. An impaired multipotency of MSCs was found in T2DM cases, where oxidative stress impaired the blood flux recovery [179]. Adipogenic MSCs from the diabetic patients failed to differentiate into fully functional adipocytes. Hence, insulin resistance might advance via hypertrophy of existing mature adipocytes [180]. The mechanistic view behind this impairment is hyperinsulinemia induced Nox4 (NADPH oxidase 4) triggering oxidative stress leading to restrict multipotency and increases adipogenic predictions of diabetic mice [181]. Additionally, insulin treatment of WT-MSCs elevated the expression of Nox4 and ultimately increased the rate of differentiation into adipocytes [181,182,183].
4.3. Need for Therapeutic Targets
Yan et al. unveiled that MSCs derived from T2DM are compromised in their multipotency which made them boost post-ischemic neovascularization in diabetic mice [181]. In 2016, Zoelen et al. showed that TGF-β promotes osteogenic differentiation of hMSCs while at the same time inhibiting adipogenic differentiation by lowering the expression of PPARγ, ADAMTS5, and AKR1B10. They concluded that these findings might support the therapeutic capabilities for preventing osteoporosis and obesity [160]. However, antidiabetic TZDs to target PPARγ, exaggerate the expression of oxidized LDL receptor 1 (OLR1) in adipocytes. Hence, targeting PPARγ alone may worsen the obesity seen in other metabolic diseases [156]. The mechanism of fat mass expansion is poorly understood. Earlier, scientists concluded that adipocyte hypertrophy comes down to fat mass expansion [184,185,186] and they justified that this condition is due to a fixed number of pre-adipocytes in adults [187,188,189,190]. Later, scientists proved this by tailoring the isotopic methodology to track down the process of adipogenesis with rare stable isotopes. Some groups were successful in identifying the adipocyte hypertrophy as the dominant mechanism of adult fat mass expansion by using mass spectrometry [191,192].
5. MSCs Response and Potential Towards Diabetes
Diabetes affect millions of people and is considered as a worldwide epidemic [193]. T2DM covers 85-95% of overall diabetes cases, having insulin resistance or problem with the insulin secretion by pancreatic β-cells [194]. MSCs have served to generate insulin-secretory cells, increase in islet engraftment and survival, and also be useful in treating diabetic ulcers and limb ischemia [195,196,197,198]. They also facilitate a micro-environmental niche by the secretion of some paracrine factors and deposition of extracellular matrix [51,199,200,201,202,203]. Therefore, MSCs have huge potential for diabetic therapeutics.
5.1. Direct therapeutic use of MSCs
There are many reports on animal models for dose-dependent therapeutics of MSCs infusion (mixed MSCs from different sites). Due to a lack of standardization, there are great variations within clinical trials regarding MSCs dosage and the frequency [120,121,122,204,205,206]. However, the factors influencing MSCs therapy are the type of MSCs (isolation and source dependent), administration of delivery, viability and purity of MSCs. Besides these factors, the most important issues are the stages, types and conditions of diabetic patients [207]. This was supported by a study in which researchers administered a direct injection of the MSCs for both the single as well as multiple times to diabetic rats that improved hyperglycemia in 4 weeks [200]. Similarly, in some other studies for the treatment of diabetic patients, scientists considered a single injection to not be enough, therefore, they administered mainly 2-4 times a day for 2 to 12- weeks, multiple injections which resulted in improvement of the patients [208,209,210,211]. In 2014, Bhansali et al. demonstrated 9 out of 11 diabetic patients reached the endpoint, and insulin requirement was lowered down by 66.7%. Also, the same group found that 7 out of 10 patients ended with 75% lower insulin requirements. Additionally, three out of them were able to discontinue the insulin completely after a single BM-MSC autologous transplantation. However, they did not mention the effective duration in their reports [208,212]. These reports still need some clarification regarding how they understood the mechanism of improvement in diabetes through MSCs therapeutics (for quality assurance).
5.2. MSCs and Their Exertion in Diabetes
It was Chen et al. who first made the incompletely differentiated MSCs into insulin-producing cells (IPCs) derived from rat, which successfully expressed insulin and nestin [213]. Later, MSCs were found to be successfully promoted islet of beta cells even after hypoxia and oxidative stress [214,215]. Until now, more than 96 phase I/II clinical trials have been attempted for the treatment of diabetes (https://clinicaltrials.gov/ct2/results?cond=Diabetes+Mellitus&term=MSC&cntry=&state=&city=&dist=), yet only few of them (Table 2) were considered promising for the effect of MSC treatment in the management of T2DM [216].
5.2.1. MSCs Clinical Trials and Combination Therapy
The randomized trials of phase 1 were first studied by a combined infusion therapy. Bone marrow derived mononuclear cells (BM-MNCs) and hyperbaric oxygen treatment combination demonstrated that infusion formula between two or more is beneficial, although both failed to synergize [217,218]. The most important scrutiny in clinical trials is the ratio of patient safety to risk. In that concern, there must be an evaluation of MSCs treatment in T2DM cases for analysis of detrimental events. In many studies, no acute and immunological events were noted, as the hypothetical risks involved here, pulmonary and upper respiratory destructions by intravenous injection which may be followed for interspersing cell fleeting through lungs, bruises caused by perforation, all these studies noticed no such developments [208,209,210,212,217,218,219,220,221,222,223,224,225]. However, mild nausea, headache, vomiting and abdominal pain were noted after MSC transplantation [212,219,222]. Therefore, the response of MSC towards diabetes is very optimistic and can be considered as a therapeutic option.
Bhansali et al. performed an infused transplant of autologous BM-MSCs and BM-MNCs in T2DM cases and found improvement in insulin sensitivity in 67% of patients. Besides this, they also concluded that MSCs mediated the IRS-1 gene expression for the betterment of insulin sensitivity, while MNCs boosted the C-peptide response [226]. Similar infusion results were obtained by Wang et al. [224]. However, the withdrawal technique of BM-MSCs and BM-MNCs from the femur or iliac crest is painful and could cause infection [227].
The increase in C-peptide and decrease in HbA1c are considered to be two major positive outcomes of MSC therapy. Guan et al. explored such improvements with WJ-MSCs therapy treatment twice in diabetes patients [228]. Taking a message from these results, the number of T-reg cells was also increased after infusion of WJ-MSCs with UC-MSCs, followed by reduced insulin dose [210]. Besides these trials, the motive of stem cell therapy was to reverse the insulin resistance and improvement in immune dysfunction. To some extent Zhao et al. found some positive results of reversing the immune dysfunction. They observed metabolic improvement and balancing between Th1/Th2/Th3 cytokine secretion using CB-MSCs, but the reversal in insulin resistance was not achieved [225].
5.2.2. Mechanistic Details of MSCs with Clinical Trials
Along with immunomodulatory and immunosuppressive roles, the regenerating capability of MSCs make them unique and a very suitable candidate for cell-based therapy in autoimmune and inflammatory disorders [229,230,231]. The differentiation potential of MSCs into IPCs is the most striking of their features, which can be used to ameliorate hyperglycemia. The differentiation of endocrine portions within the pancreas is strictly regulated by transcription factors Pdx-1, Ngn-3, NeuroD1, Pax4 and Pax6 [232]. For the equitable reprogramming of these cells, it becomes mandatory for the MSCs to differentiate into IPCs. In 2005, human BM-MSCs were successfully differentiated into IPCs with the use of adenoviral vectors encoding for murine Pdx-1. Later, Xie et al. studied the differentiation of human BM-MSCs into IPCs by a three-step process and resulted in them becoming insulin-secreting cells in a glucose dose-dependent manner [233,234]. Similarly, Nam et al. performed in vitro differentiation of human eyelid AD-MSCs into IPCs and transplanted them into T2DM mice model [235]. Out of the two groups; T2DM and control, T2DM mice group experienced IPCs mediated improvement, high IL-6 and also an increase in circulating insulin level promoting the metabolic improvements.
5.3. Possible Approaches of MSCs prior to Clinical Trials
Functions of stem cells are highly dependent on the microenvironment, which is strictly regulated by its ECM (extracellular matrix), growth factors and immune cells [236]. ECM of MSCs has a great potential for the quality improvement of MSCs such as adhesion and proliferation. Disturbed metabolic conditions and hyperglycemia often leads to organ damage and also hampers the quality of MSCs, which is a major hurdle of autologous clinical application. Therefore, we highly recommend a particular kind of 3D (three dimensional) ECM culture, which may be used to enhance such functions of MSCs [237]. For instance, Block et al. used 3D-ECM culture for a phenotype of interest on the basis of cell size and stage-specific embryonic antigen-4 levels [238]. These kinds of findings may assure the autologous use of MSCs, with high quality and genetic modifications in order to improve the therapeutic potential of MSCs in several diseases and disorders [239].
The survival rate of MSCs is an essential quality factor for clinical trials. We have discussed the loss of potency in frozen-thawed MSCs [240], but in an experiment of allergic asthma, frozen-thawed MSCs were as efficient as the fresh MSCs [241]. Hence, we need more light to explore the features of frozen-thawed MSCs as compared to fresh MSCs. Apart from that, some scientists have explored the challenges of MSCs to sublethal risks of cellular stress in vitro, i.e., hypoxia, heat or compromised nutrition. With preconditioning incubation, MSCs demonstrated a reduction of cell apoptosis in vivo while maintaining their biological functions [242,243]. When comparing hypoxia and normoxia exposure to cells in vitro, hypoxia-treated MSCs emerged to show an increase in population multiplying rate [244,245]. This phenomenon can be explained by understanding the role of stabilizing factor HIF-1α. During hypoxia, the expression of HIF-1α rises by 3.4 fold in MSCs that in turn to reduce the ROS (reactive oxygen species), block the oxidative phosphorylation and promote glycolysis [242,245]. Besides that, HIF-1α also activates NF-κB to downregulate the level of Bcl (B cell lymphoma) and caspase 3, and activation of PrPc (cellular prion protein) enhances the superoxide dismutase (SOD) and catalase to attain complete protection from oxidative stress [242,246].
Enduring oxidative stress in hypertension and diabetic conditions, cells are influenced by ROS [247]. High-calorie diet induces ROS accumulation in the adipose tissue of the T2DM mice model and promotes senescence of MSCs by expressing β-galactosidase and p53 [248]. Besides, the viability of MSCs is severely hampered by oxidative stress [249,250]. Thus, the direct use of MSCs for treatment may not be practical. In 2016, Cheng et al. observed the upregulation of sex-determining region Y-box 2, (SOX2), Oct-4, and NANOG in high glucose-treated MSCs by the influence of intracellular ROS formation, signifying the improvement in the stemness of MSCs [251]. These pieces of evidence show preconditioning of MSCs may be of help in maintaining function and enhancing survival rates in clinical trials.
6. Perspective
MSC therapy exemplifies a materializing style of modern alternative treatment with the retention to hold site-specific immune regulation that controls T cells in autoimmune diseases and allograft rejection. The current review article on immunosuppressive properties holds great confidence for treating immune-mediated diseases, obesity, CVD and diabetes. However, it is also mandatory not to augment the therapeutic potential as many unanswered questions need elucidation before making promises. MSCs are known to bear effects in vascular repair by enactments into blood vessels, differentiation into endothelial cells, pericytes and other vascular phenotypes [252,253,254], which is further supported by autocrine and paracrine properties by producing vascular growth factors and proangiogenic cytokines [255,256]. As far as the endocrine properties of MSCs are concerned, it is credible that the classification of admissible effector molecules could lead to novel treatment and rehabilitation of cellular therapy with MSCs [29].
There is very limiting but reassuring data available for ameliorating glycemic controlled MSCs therapy in T2DM cases. We have highlighted many experiments to overcome hyperglycemia such as differentiating MSCs into IPCs (insulin-producing cells), mitigating insulin resistance, conversion of alpha cells to beta cells and remodeling pancreatic regeneration. However, animal models used for the aforementioned analysis did not equal human T2DM patients and hence, the underlying scheme involved here must be cross-examined in detail.
Coordinators for upcoming research must incorporate a definite identification of cell markers such as the marker explored by Hudak et al. [257]. Standardized and validated isolation and culture protocols with lineage differentiation and stimulation methods, which may ease the animal and clinical studies. Additionally, cell modification, injection frequency and dosages are required to be studied in detail to further guide the therapeutic potential of MSCs. An exceptional insight of this compelling cell population might be apprehensive of a contemporary therapeutic scheme to recover the immune response in an array of immune-mediated diseases, obesity, CVD and diabetes.
Author Contributions
Writing—original draft preparation, V.K.M. and F.P.; writing—review and editing, H.-H.S., and D.L.; visualization, F.P. and H.-H.S.; supervision, T.-F.C., E.I., L.-Y.K.; project administration, T.-F.C. and L.-Y.K.; funding acquisition, T.-F.C., L.-Y.K. and H.-H.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported in part by grants from the Kaohsiung Medical University grant (KMU-DK108004, KMU-TC108A03-0) and Taiwan Ministry of Science and Technology (107-2314-B-037-114-MY3, 107-2314-B-037-064-MY3, 106-2628-B-037-001-MY3) to L.-Y.K., T.-F.C. and H.-C.C. respectively.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| ADAMTS5 | A disintegrin and metalloproteinase with thrombospondin motifs 5 |
| AKR1B10 | Aldo-keto reductase Family 1 Member B10 |
| Akt-MSC | Protein kinase B recombinant MSCs |
| APCs | Antigen presenting cells |
| BCL | B Cell Lymphoma |
| BDNF | Brain-derived neurotropic factor |
| BMP | Bone morphogenic protein |
| C/EBP | CCAAT-enhancer binding proteins |
| Cbfa1 | Core-binding factor alpha 1 |
| CCL2 | Chemokine ligand-2 |
| CD | Cluster of differentiation |
| C-MSC | Cord tissue derived MSCs |
| COX-2 | Cyclooxygenase 2 |
| CR3 | Complement Receptor 3 |
| CVD | Cardiovascular disease |
| CXCL | Chemokine Ligand |
| CXCR | Chemokine receptor |
| ECM | Extracellular Matrix |
| Foxp3+ | Fork-head box P3 |
| GCSF | Granulocyte-colony stimulating factor |
| GDM | Gestational diabetes mellitus |
| HGF | Hepatocyte growth factor |
| HIF-1α | Hypoxia inducible factor-1α |
| HLA-G | Human leukocyte antigen-G |
| HO | Heme-oxygenase |
| IBMX | 3-isobutyl-1-methylxanthine |
| IDO | Indolamine 2:3-dioxygenase |
| IL | Interleukin |
| IR | Insulin resistance |
| IPC | Insulin producing cell |
| iNOS | Inducible NO synthase |
| MAC | Membrane attack complex |
| MMP-9 | Metalloproteinase 9 |
| MIP α/β | Macrophage inflammatory protein |
| MNC | Mononuclear cell |
| MSC | Mesenchymal stem cell |
| N-MSC | Non-tumor MSCs |
| Ngn 3 | Neurogenin 3 |
| NO | Nitric oxide |
| Nox4 | NADPH oxidase 4 |
| 04-Oct | Octamer-binding transcription factor 4 |
| OLR1 | Oxidized LDL receptor 1 |
| P-MSC | Placenta tissue derived MSCs |
| Pax 4 | Pairedbox 4 |
| PBMC | Peripheral blood mononuclear cell |
| Pdx-1 | Pancreatic and duodenal homeobox-1 |
| PGE2 | Prostaglandin E2 |
| PGH2 | Prostaglandin H2 |
| PPAR-γ | Peroxisome proliferator-activated receptor-γ |
| RANTES | Regulated on activation, Normal T cell expressed and secreted |
| ROS | Reactive oxygen species |
| Runx2 | Runt-related gene 2 |
| SOD | Superoxide dismutase |
| SOX2 | Sex-determining region Y-box 2, (SOX2) |
| T1DM | Type 1 diabetes mellitus |
| T2DM | Type 2 diabetes mellitus |
| TB4 | Thymosin β-4 |
| TNF-α | Tissue necrosis factor-α |
| Treg | Immunoregulatory T cell |
| T-MSC | Tumor derived MSCs |
| TGF-β | Transforming growth factor-β |
| TSG6 | TNF-inducible gene -6 protein |
| TZD | Thiazolidinedione |
| VEGF | Vascular endothelial growth factors |
| ULBP | UL16 binding protein |
| ZFP423 | Zinc finger protein 423 |
References
- Wang, Y.; Han, Z.B.; Ma, J.; Zuo, C.; Geng, J.; Gong, W.; Sun, Y.; Li, H.; Wang, B.; Zhang, L.; et al. A toxicity study of multiple-administration human umbilical cord mesenchymal stem cells in cynomolgus monkeys. Stem Cells Dev. 2012, 21, 1401–1408. [Google Scholar] [CrossRef]
- Ullah, I.; Subbarao, R.B.; Rho, G.J. Human mesenchymal stem cells—Current trends and future prospective. Biosci. Rep. 2015, 35. [Google Scholar] [CrossRef] [PubMed]
- Friedenstein, A.J.; Piatetzky, S.I.I.; Petrakova, K.V. Osteogenesis in transplants of bone marrow cells. J. Embryol. Exp. Morphol. 1966, 16, 381–390. [Google Scholar] [PubMed]
- Caplan, A.I. Mesenchymal stem cells. J. Orthop. Res. Off. Publ. Orthop. Res. Soc. 1991, 9, 641–650. [Google Scholar] [CrossRef]
- Qiao, C.; Xu, W.; Zhu, W.; Hu, J.; Qian, H.; Yin, Q.; Jiang, R.; Yan, Y.; Mao, F.; Yang, H.; et al. Human mesenchymal stem cells isolated from the umbilical cord. Cell Biol. Int. 2008, 32, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, E.M.; Le Blanc, K.; Dominici, M.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Deans, R.J.; Krause, D.S.; Keating, A.; The International Society for Cellular Therapy. Clarification of the nomenclature for MSC: The International Society for Cellular Therapy position statement. Cytotherapy 2005, 7, 393–395. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I. What’s in a name? Tissue Eng. Part A 2010, 16, 2415–2417. [Google Scholar] [CrossRef] [Green Version]
- Caplan, A.I. Mesenchymal Stem Cells: Time to Change the Name! Stem Cells Transl. Med. 2017, 6, 1445–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.C.; Kim, H.S.; Shin, T.H.; Kang, I.; Lee, J.Y.; Kim, J.J.; Kang, H.K.; Seo, Y.; Lee, S.; Yu, K.R.; et al. PGE2 maintains self-renewal of human adult stem cells via EP2-mediated autocrine signaling and its production is regulated by cell-to-cell contact. Sci. Rep. 2016, 6, 26298. [Google Scholar] [CrossRef] [Green Version]
- Mareschi, K.; Ferrero, I.; Rustichelli, D.; Aschero, S.; Gammaitoni, L.; Aglietta, M.; Madon, E.; Fagioli, F. Expansion of mesenchymal stem cells isolated from pediatric and adult donor bone marrow. J. Cell Biochem. 2006, 97, 744–754. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Wehner, R.; Bornhauser, M.; Wassmuth, R.; Bachmann, M.; Schmitz, M. Immunomodulatory properties of mesenchymal stromal cells and their therapeutic consequences for immune-mediated disorders. Stem Cells Dev. 2010, 19, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Hua, J.; Gong, J.; Meng, H.; Xu, B.; Yao, L.; Qian, M.; He, Z.; Zou, S.; Zhou, B.; Song, Z. Comparison of different methods for the isolation of mesenchymal stem cells from umbilical cord matrix: Proliferation and multilineage differentiation as compared to mesenchymal stem cells from umbilical cord blood and bone marrow. Cell Biol. Int. 2013. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Hoogduijn, M.J.; Baan, C.C.; Korevaar, S.S.; de Kuiper, R.; Yan, L.; Wang, L.; van Besouw, N.M. Adipose Tissue-Derived Mesenchymal Stem Cells Have a Heterogenic Cytokine Secretion Profile. Stem Cells Int. 2017, 2017, 4960831. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Yu, X.; Wang, Z.; Wang, F.; Wang, L.; Gao, H.; Chen, Y.; Zhao, W.; Jia, Z.; Yan, S.; et al. Long term effects of the implantation of Wharton’s jelly-derived mesenchymal stem cells from the umbilical cord for newly-onset type 1 diabetes mellitus. Endocr. J. 2013, 60, 347–357. [Google Scholar] [CrossRef] [Green Version]
- Nemeth, K.; Leelahavanichkul, A.; Yuen, P.S.; Mayer, B.; Parmelee, A.; Doi, K.; Robey, P.G.; Leelahavanichkul, K.; Koller, B.H.; Brown, J.M.; et al. Bone marrow stromal cells attenuate sepsis via prostaglandin E(2)-dependent reprogramming of host macrophages to increase their interleukin-10 production. Nat. Med. 2009, 15, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Sabapathy, V.; Ravi, S.; Srivastava, V.; Srivastava, A.; Kumar, S. Long-term cultured human term placenta-derived mesenchymal stem cells of maternal origin displays plasticity. Stem Cells Int. 2012, 2012, 174328. [Google Scholar] [CrossRef]
- Yin, J.Q.; Zhu, J.; Ankrum, J.A. Manufacturing of primed mesenchymal stromal cells for therapy. Nat. Biomed. Eng. 2019, 3, 90–104. [Google Scholar] [CrossRef]
- Elahi, K.C.; Klein, G.; Avci-Adali, M.; Sievert, K.D.; MacNeil, S.; Aicher, W.K. Human Mesenchymal Stromal Cells from Different Sources Diverge in Their Expression of Cell Surface Proteins and Display Distinct Differentiation Patterns. Stem Cells Int. 2016, 2016, 5646384. [Google Scholar] [CrossRef] [Green Version]
- Hass, R.; Kasper, C.; Bohm, S.; Jacobs, R. Different populations and sources of human mesenchymal stem cells (MSC): A comparison of adult and neonatal tissue-derived MSC. Cell Commun. Signal CCS 2011, 9, 12. [Google Scholar] [CrossRef] [Green Version]
- Da Silva Meirelles, L.; Chagastelles, P.C.; Nardi, N.B. Mesenchymal stem cells reside in virtually all post-natal organs and tissues. J. Cell Sci. 2006, 119, 2204–2213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talwadekar, M.D.; Kale, V.P.; Limaye, L.S. Placenta-derived mesenchymal stem cells possess better immunoregulatory properties compared to their cord-derived counterparts-a paired sample study. Sci. Rep. 2015, 5, 15784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagaishi, K.; Mizue, Y.; Chikenji, T.; Otani, M.; Nakano, M.; Saijo, Y.; Tsuchida, H.; Ishioka, S.; Nishikawa, A.; Saito, T.; et al. Umbilical cord extracts improve diabetic abnormalities in bone marrow-derived mesenchymal stem cells and increase their therapeutic effects on diabetic nephropathy. Sci. Rep. 2017, 7, 8484. [Google Scholar] [CrossRef] [Green Version]
- Luk, F.; Carreras-Planella, L.; Korevaar, S.S.; de Witte, S.F.H.; Borras, F.E.; Betjes, M.G.H.; Baan, C.C.; Hoogduijn, M.J.; Franquesa, M. Inflammatory Conditions Dictate the Effect of Mesenchymal Stem or Stromal Cells on B Cell Function. Front. Immunol. 2017, 8, 1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Chen, X.; Cao, W.; Shi, Y. Plasticity of mesenchymal stem cells in immunomodulation: Pathological and therapeutic implications. Nat. Immunol. 2014, 15, 1009–1016. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; Zhang, L.; Zhao, X.; Xu, G.; Zhang, Y.; Roberts, A.I.; Zhao, R.C.; Shi, Y. Mesenchymal stem cell-mediated immunosuppression occurs via concerted action of chemokines and nitric oxide. Cell Stem Cell 2008, 2, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.X.; Zhang, Y.; Liu, B.; Zhang, S.X.; Wu, Y.; Yu, X.D.; Mao, N. Human mesenchymal stem cells inhibit differentiation and function of monocyte-derived dendritic cells. Blood 2005, 105, 4120–4126. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Ge, W.; Li, C.; You, S.; Liao, L.; Han, Q.; Deng, W.; Zhao, R.C. Effects of mesenchymal stem cells on differentiation, maturation, and function of human monocyte-derived dendritic cells. Stem Cells Dev. 2004, 13, 263–271. [Google Scholar] [CrossRef]
- Bernardo, M.E.; Fibbe, W.E. Mesenchymal stromal cells: Sensors and switchers of inflammation. Cell Stem Cell 2013, 13, 392–402. [Google Scholar] [CrossRef] [Green Version]
- Vasandan, A.B.; Jahnavi, S.; Shashank, C.; Prasad, P.; Kumar, A.; Prasanna, S.J. Human Mesenchymal stem cells program macrophage plasticity by altering their metabolic status via a PGE2-dependent mechanism. Sci. Rep. 2016, 6, 38308. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.; Lee, R.H.; Bazhanov, N.; Oh, J.Y.; Prockop, D.J. Anti-inflammatory protein TSG-6 secreted by activated MSCs attenuates zymosan-induced mouse peritonitis by decreasing TLR2/NF-kappaB signaling in resident macrophages. Blood 2011, 118, 330–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolaczkowska, E.; Kubes, P. Neutrophil recruitment and function in health and inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef] [PubMed]
- Di Nicola, M.; Carlo-Stella, C.; Magni, M.; Milanesi, M.; Longoni, P.D.; Matteucci, P.; Grisanti, S.; Gianni, A.M. Human bone marrow stromal cells suppress T-lymphocyte proliferation induced by cellular or nonspecific mitogenic stimuli. Blood 2002, 99, 3838–3843. [Google Scholar] [CrossRef] [PubMed]
- Le Blanc, K.; Tammik, L.; Sundberg, B.; Haynesworth, S.E.; Ringden, O. Mesenchymal stem cells inhibit and stimulate mixed lymphocyte cultures and mitogenic responses independently of the major histocompatibility complex. Scand. J. Immunol. 2003, 57, 11–20. [Google Scholar] [CrossRef]
- Potian, J.A.; Aviv, H.; Ponzio, N.M.; Harrison, J.S.; Rameshwar, P. Veto-like activity of mesenchymal stem cells: Functional discrimination between cellular responses to alloantigens and recall antigens. J. Immunol. 2003, 171, 3426–3434. [Google Scholar] [CrossRef] [Green Version]
- Tse, W.T.; Pendleton, J.D.; Beyer, W.M.; Egalka, M.C.; Guinan, E.C. Suppression of allogeneic T-cell proliferation by human marrow stromal cells: Implications in transplantation. Transplantation 2003, 75, 389–397. [Google Scholar] [CrossRef]
- Bartholomew, A.; Sturgeon, C.; Siatskas, M.; Ferrer, K.; McIntosh, K.; Patil, S.; Hardy, W.; Devine, S.; Ucker, D.; Deans, R.; et al. Mesenchymal stem cells suppress lymphocyte proliferation in vitro and prolong skin graft survival in vivo. Exp. Hematol. 2002, 30, 42–48. [Google Scholar] [CrossRef]
- Djouad, F.; Plence, P.; Bony, C.; Tropel, P.; Apparailly, F.; Sany, J.; Noel, D.; Jorgensen, C. Immunosuppressive effect of mesenchymal stem cells favors tumor growth in allogeneic animals. Blood 2003, 102, 3837–3844. [Google Scholar] [CrossRef] [Green Version]
- Ge, W.; Jiang, J.; Baroja, M.L.; Arp, J.; Zassoko, R.; Liu, W.; Bartholomew, A.; Garcia, B.; Wang, H. Infusion of mesenchymal stem cells and rapamycin synergize to attenuate alloimmune responses and promote cardiac allograft tolerance. Am. J. Transplant. 2009, 9, 1760–1772. [Google Scholar] [CrossRef]
- Waterman, R.S.; Tomchuck, S.L.; Henkle, S.L.; Betancourt, A.M. A new mesenchymal stem cell (MSC) paradigm: Polarization into a pro-inflammatory MSC1 or an Immunosuppressive MSC2 phenotype. PLoS ONE 2010, 5, e10088. [Google Scholar] [CrossRef]
- Heo, J.S.; Choi, Y.; Kim, H.O. Adipose-Derived Mesenchymal Stem Cells Promote M2 Macrophage Phenotype through Exosomes. Stem Cells Int. 2019, 2019, 7921760. [Google Scholar] [CrossRef] [PubMed]
- Morrison, T.J.; Jackson, M.V.; Cunningham, E.K.; Kissenpfennig, A.; McAuley, D.F.; O’Kane, C.M.; Krasnodembskaya, A.D. Mesenchymal Stromal Cells Modulate Macrophages in Clinically Relevant Lung Injury Models by Extracellular Vesicle Mitochondrial Transfer. Am. J. Respir. Crit. Care Med. 2017, 196, 1275–1286. [Google Scholar] [CrossRef] [PubMed]
- Park, D.; Spencer, J.A.; Koh, B.I.; Kobayashi, T.; Fujisaki, J.; Clemens, T.L.; Lin, C.P.; Kronenberg, H.M.; Scadden, D.T. Endogenous bone marrow MSCs are dynamic, fate-restricted participants in bone maintenance and regeneration. Cell Stem Cell 2012, 10, 259–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melief, S.M.; Schrama, E.; Brugman, M.H.; Tiemessen, M.M.; Hoogduijn, M.J.; Fibbe, W.E.; Roelofs, H. Multipotent stromal cells induce human regulatory T cells through a novel pathway involving skewing of monocytes toward anti-inflammatory macrophages. Stem Cells 2013, 31, 1980–1991. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, S.; Pittenger, M.F. Human mesenchymal stem cells modulate allogeneic immune cell responses. Blood 2005, 105, 1815–1822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beyth, S.; Borovsky, Z.; Mevorach, D.; Liebergall, M.; Gazit, Z.; Aslan, H.; Galun, E.; Rachmilewitz, J. Human mesenchymal stem cells alter antigen-presenting cell maturation and induce T-cell unresponsiveness. Blood 2005, 105, 2214–2219. [Google Scholar] [CrossRef] [Green Version]
- Glennie, S.; Soeiro, I.; Dyson, P.J.; Lam, E.W.; Dazzi, F. Bone marrow mesenchymal stem cells induce division arrest anergy of activated T cells. Blood 2005, 105, 2821–2827. [Google Scholar] [CrossRef]
- Deng, W.; Han, Q.; Liao, L.; You, S.; Deng, H.; Zhao, R.C. Effects of allogeneic bone marrow-derived mesenchymal stem cells on T and B lymphocytes from BXSB mice. DNA Cell Biol. 2005, 24, 458–463. [Google Scholar] [CrossRef]
- Corcione, A.; Benvenuto, F.; Ferretti, E.; Giunti, D.; Cappiello, V.; Cazzanti, F.; Risso, M.; Gualandi, F.; Mancardi, G.L.; Pistoia, V.; et al. Human mesenchymal stem cells modulate B-cell functions. Blood 2006, 107, 367–372. [Google Scholar] [CrossRef] [Green Version]
- Eggenhofer, E.; Hoogduijn, M.J. Mesenchymal stem cell-educated macrophages. Transplant. Res. 2012, 1, 12. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Tredget, E.E.; Wu, P.Y.; Wu, Y. Paracrine factors of mesenchymal stem cells recruit macrophages and endothelial lineage cells and enhance wound healing. PLoS ONE 2008, 3, e1886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drago, D.; Cossetti, C.; Iraci, N.; Gaude, E.; Musco, G.; Bachi, A.; Pluchino, S. The stem cell secretome and its role in brain repair. Biochimie 2013, 95, 2271–2285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, S.A.; Sherman, L.; Munoz, J.; Rameshwar, P. Immunological properties of mesenchymal stem cells and clinical implications. Arch. Immunol. Ther. Exp. 2008, 56, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Scholich, K.; Geisslinger, G. Is mPGES-1 a promising target for pain therapy? Trends Pharmacol. Sci. 2006, 27, 399–401. [Google Scholar] [CrossRef]
- Eibl, G.; Bruemmer, D.; Okada, Y.; Duffy, J.P.; Law, R.E.; Reber, H.A.; Hines, O.J. PGE(2) is generated by specific COX-2 activity and increases VEGF production in COX-2-expressing human pancreatic cancer cells. Biochem. Biophys. Res. Commun. 2003, 306, 887–897. [Google Scholar] [CrossRef]
- Kalinski, P. Regulation of immune responses by prostaglandin E2. J. Immunol. 2012, 188, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Park, J.Y.; Pillinger, M.H.; Abramson, S.B. Prostaglandin E2 synthesis and secretion: The role of PGE2 synthases. Clin. Immunol. 2006, 119, 229–240. [Google Scholar] [CrossRef]
- Seno, H.; Oshima, M.; Ishikawa, T.O.; Oshima, H.; Takaku, K.; Chiba, T.; Narumiya, S.; Taketo, M.M. Cyclooxygenase 2- and prostaglandin E(2) receptor EP(2)-dependent angiogenesis in Apc(Delta716) mouse intestinal polyps. Cancer Res. 2002, 62, 506–511. [Google Scholar]
- Castellone, M.D.; Teramoto, H.; Williams, B.O.; Druey, K.M.; Gutkind, J.S. Prostaglandin E2 promotes colon cancer cell growth through a Gs-axin-beta-catenin signaling axis. Science 2005, 310, 1504–1510. [Google Scholar] [CrossRef]
- Kim, H.S.; Shin, T.H.; Lee, B.C.; Yu, K.R.; Seo, Y.; Lee, S.; Seo, M.S.; Hong, I.S.; Choi, S.W.; Seo, K.W.; et al. Human umbilical cord blood mesenchymal stem cells reduce colitis in mice by activating NOD2 signaling to COX2. Gastroenterology 2013, 145, 1392–1403. [Google Scholar] [CrossRef]
- Kim, H.S.; Shin, T.H.; Yang, S.R.; Seo, M.S.; Kim, D.J.; Kang, S.K.; Park, J.H.; Kang, K.S. Implication of NOD1 and NOD2 for the differentiation of multipotent mesenchymal stem cells derived from human umbilical cord blood. PLoS ONE 2010, 5, e15369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.S.; Yun, J.W.; Shin, T.H.; Lee, S.H.; Lee, B.C.; Yu, K.R.; Seo, Y.; Lee, S.; Kang, T.W.; Choi, S.W.; et al. Human umbilical cord blood mesenchymal stem cell-derived PGE2 and TGF-beta1 alleviate atopic dermatitis by reducing mast cell degranulation. Stem Cells 2015, 33, 1254–1266. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kim, H.S.; Roh, K.H.; Lee, B.C.; Shin, T.H.; Yoo, J.M.; Kim, Y.L.; Yu, K.R.; Kang, K.S.; Seo, K.W. DNA methyltransferase inhibition accelerates the immunomodulation and migration of human mesenchymal stem cells. Sci. Rep. 2015, 5, 8020. [Google Scholar] [CrossRef] [Green Version]
- Yu, K.R.; Lee, J.Y.; Kim, H.S.; Hong, I.S.; Choi, S.W.; Seo, Y.; Kang, I.; Kim, J.J.; Lee, B.C.; Lee, S.; et al. A p38 MAPK-mediated alteration of COX-2/PGE2 regulates immunomodulatory properties in human mesenchymal stem cell aging. PLoS ONE 2014, 9, e102426. [Google Scholar] [CrossRef] [PubMed]
- Dohadwala, M.; Batra, R.K.; Luo, J.; Lin, Y.; Krysan, K.; Pold, M.; Sharma, S.; Dubinett, S.M. Autocrine/paracrine prostaglandin E2 production by non-small cell lung cancer cells regulates matrix metalloproteinase-2 and CD44 in cyclooxygenase-2-dependent invasion. J. Biol. Chem. 2002, 277, 50828–50833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gnecchi, M.; He, H.; Liang, O.D.; Melo, L.G.; Morello, F.; Mu, H.; Noiseux, N.; Zhang, L.; Pratt, R.E.; Ingwall, J.S.; et al. Paracrine action accounts for marked protection of ischemic heart by Akt-modified mesenchymal stem cells. Nat. Med. 2005, 11, 367–368. [Google Scholar] [CrossRef]
- Gnecchi, M.; He, H.; Noiseux, N.; Liang, O.D.; Zhang, L.; Morello, F.; Mu, H.; Melo, L.G.; Pratt, R.E.; Ingwall, J.S.; et al. Evidence supporting paracrine hypothesis for Akt-modified mesenchymal stem cell-mediated cardiac protection and functional improvement. FASEB J. 2006, 20, 661–669. [Google Scholar] [CrossRef]
- Hegyi, B.; Kudlik, G.; Monostori, E.; Uher, F. Activated T-cells and pro-inflammatory cytokines differentially regulate prostaglandin E2 secretion by mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2012, 419, 215–220. [Google Scholar] [CrossRef]
- Toulon, P.; Frere, E.; Bachmeyer, C.; Candia, N.; Blanche, P.; Sereni, D.; Sicard, D. Fibrin polymerization defect in HIV-infected patients—Evidence for a critical role of albumin in the prolongation of thrombin and reptilase clotting times. Thromb. Haemost. 1995, 73, 349–355. [Google Scholar] [CrossRef]
- Sattler, C.; Steinsdoerfer, M.; Offers, M.; Fischer, E.; Schierl, R.; Heseler, K.; Daubener, W.; Seissler, J. Inhibition of T-cell proliferation by murine multipotent mesenchymal stromal cells is mediated by CD39 expression and adenosine generation. Cell Transplant. 2011, 20, 1221–1230. [Google Scholar] [CrossRef] [Green Version]
- Tasso, R.; Ilengo, C.; Quarto, R.; Cancedda, R.; Caspi, R.R.; Pennesi, G. Mesenchymal stem cells induce functionally active T-regulatory lymphocytes in a paracrine fashion and ameliorate experimental autoimmune uveitis. Invest. Ophthalmol. Vis. Sci. 2012, 53, 786–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spaggiari, G.M.; Moretta, L. Interactions between mesenchymal stem cells and dendritic cells. Adv. Biochem. Eng. Biotechnol. 2013, 130, 199–208. [Google Scholar] [PubMed]
- Nauta, A.J.; Kruisselbrink, A.B.; Lurvink, E.; Willemze, R.; Fibbe, W.E. Mesenchymal stem cells inhibit generation and function of both CD34+-derived and monocyte-derived dendritic cells. J. Immunol. 2006, 177, 2080–2087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramasamy, R.; Fazekasova, H.; Lam, E.W.; Soeiro, I.; Lombardi, G.; Dazzi, F. Mesenchymal stem cells inhibit dendritic cell differentiation and function by preventing entry into the cell cycle. Transplantation 2007, 83, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Harizi, H.; Norbert, G. Inhibition of IL-6, TNF-alpha, and cyclooxygenase-2 protein expression by prostaglandin E2-induced IL-10 in bone marrow-derived dendritic cells. Cell. Immunol. 2004, 228, 99–109. [Google Scholar] [PubMed]
- Braun, D.; Longman, R.S.; Albert, M.L. A two-step induction of indoleamine 2,3 dioxygenase (IDO) activity during dendritic-cell maturation. Blood 2005, 106, 2375–2381. [Google Scholar] [CrossRef] [Green Version]
- Krause, P.; Singer, E.; Darley, P.I.; Klebensberger, J.; Groettrup, M.; Legler, D.F. Prostaglandin E2 is a key factor for monocyte-derived dendritic cell maturation: Enhanced T cell stimulatory capacity despite IDO. J. Leukoc. Biol. 2007, 82, 1106–1114. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, S.A.; Roobrouck, V.D.; Verfaillie, C.M.; Van Gool, S.W. Immunological characteristics of human mesenchymal stem cells and multipotent adult progenitor cells. Immunol. Cell Biol. 2013, 91, 32–39. [Google Scholar] [CrossRef]
- Krampera, M.; Cosmi, L.; Angeli, R.; Pasini, A.; Liotta, F.; Andreini, A.; Santarlasci, V.; Mazzinghi, B.; Pizzolo, G.; Vinante, F.; et al. Role for interferon-gamma in the immunomodulatory activity of human bone marrow mesenchymal stem cells. Stem Cells 2006, 24, 386–398. [Google Scholar] [CrossRef]
- DelaRosa, O.; Lombardo, E.; Beraza, A.; Mancheno-Corvo, P.; Ramirez, C.; Menta, R.; Rico, L.; Camarillo, E.; Garcia, L.; Abad, J.L.; et al. Requirement of IFN-gamma-mediated indoleamine 2,3-dioxygenase expression in the modulation of lymphocyte proliferation by human adipose-derived stem cells. Tissue Eng. Part A 2009, 15, 2795–2806. [Google Scholar] [CrossRef]
- Zhao, Q.; Ren, H.; Han, Z. Mesenchymal stem cells: Immunomodulatory capability and clinical potential in immune diseases. J. Cell. Immunother. 2016, 2, 3–20. [Google Scholar] [CrossRef] [Green Version]
- Meisel, R.; Zibert, A.; Laryea, M.; Gobel, U.; Daubener, W.; Dilloo, D. Human bone marrow stromal cells inhibit allogeneic T-cell responses by indoleamine 2,3-dioxygenase-mediated tryptophan degradation. Blood 2004, 103, 4619–4621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmusson, I.; Ringden, O.; Sundberg, B.; Le Blanc, K. Mesenchymal stem cells inhibit the formation of cytotoxic T lymphocytes, but not activated cytotoxic T lymphocytes or natural killer cells. Transplantation 2003, 76, 1208–1213. [Google Scholar] [CrossRef] [PubMed]
- Spaggiari, G.M.; Capobianco, A.; Becchetti, S.; Mingari, M.C.; Moretta, L. Mesenchymal stem cell-natural killer cell interactions: Evidence that activated NK cells are capable of killing MSCs, whereas MSCs can inhibit IL-2-induced NK-cell proliferation. Blood 2006, 107, 1484–1490. [Google Scholar] [CrossRef] [PubMed]
- Spaggiari, G.M.; Capobianco, A.; Abdelrazik, H.; Becchetti, F.; Mingari, M.C.; Moretta, L. Mesenchymal stem cells inhibit natural killer-cell proliferation, cytotoxicity, and cytokine production: Role of indoleamine 2,3-dioxygenase and prostaglandin E2. Blood 2008, 111, 1327–1333. [Google Scholar] [CrossRef]
- Sotiropoulou, P.A.; Perez, S.A.; Gritzapis, A.D.; Baxevanis, C.N.; Papamichail, M. Interactions between human mesenchymal stem cells and natural killer cells. Stem Cells 2006, 24, 74–85. [Google Scholar] [CrossRef]
- Ricklin, D.; Hajishengallis, G.; Yang, K.; Lambris, J.D. Complement: A key system for immune surveillance and homeostasis. Nat. Immunol. 2010, 11, 785–797. [Google Scholar] [CrossRef] [Green Version]
- Merle, N.S.; Noe, R.; Halbwachs-Mecarelli, L.; Fremeaux-Bacchi, V.; Roumenina, L.T. Complement System Part II: Role in Immunity. Front. Immuno 2015, 6, 257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merle, N.S.; Church, S.E.; Fremeaux-Bacchi, V.; Roumenina, L.T. Complement System Part I—Molecular Mechanisms of Activation and Regulation. Front. Immunol. 2015, 6, 262. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Lin, F. Mesenchymal stem cells are injured by complement after their contact with serum. Blood 2012, 120, 3436–3443. [Google Scholar] [CrossRef] [Green Version]
- Vorup-Jensen, T.; Jensen, R.K. Structural Immunology of Complement Receptors 3 and 4. Front. Immunol. 2018, 9, 2716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galleu, A.; Riffo-Vasquez, Y.; Trento, C.; Lomas, C.; Dolcetti, L.; Cheung, T.S.; von Bonin, M.; Barbieri, L.; Halai, K.; Ward, S.; et al. Apoptosis in mesenchymal stromal cells induces in vivo recipient-mediated immunomodulation. Sci. Transl. Med. 2017, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braza, F.; Dirou, S.; Forest, V.; Sauzeau, V.; Hassoun, D.; Chesne, J.; Cheminant-Muller, M.A.; Sagan, C.; Magnan, A.; Lemarchand, P. Mesenchymal Stem Cells Induce Suppressive Macrophages Through Phagocytosis in a Mouse Model of Asthma. Stem Cells 2016, 34, 1836–1845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Witte, S.F.H.; Luk, F.; Sierra Parraga, J.M.; Gargesha, M.; Merino, A.; Korevaar, S.S.; Shankar, A.S.; O’Flynn, L.; Elliman, S.J.; Roy, D.; et al. Immunomodulation By Therapeutic Mesenchymal Stromal Cells (MSC) Is Triggered Through Phagocytosis of MSC By Monocytic Cells. Stem Cells 2018, 36, 602–615. [Google Scholar] [CrossRef] [Green Version]
- Gavin, C.; Meinke, S.; Heldring, N.; Heck, K.A.; Achour, A.; Iacobaeus, E.; Hoglund, P.; Le Blanc, K.; Kadri, N. The Complement System Is Essential for the Phagocytosis of Mesenchymal Stromal Cells by Monocytes. Front. Immunol. 2019, 10, 2249. [Google Scholar] [CrossRef]
- Le Blanc, K.; Mougiakakos, D. Multipotent mesenchymal stromal cells and the innate immune syStem. Nat. Rev. Immunol. 2012, 12, 383–396. [Google Scholar] [CrossRef]
- Tu, Z.; Li, Q.; Bu, H.; Lin, F. Mesenchymal stem cells inhibit complement activation by secreting factor H. Stem Cells Dev. 2010, 19, 1803–1809. [Google Scholar] [CrossRef]
- Li, Y.; Fung, J.; Lin, F. Local Inhibition of Complement Improves Mesenchymal Stem Cell Viability and Function After Administration. Mol. Ther. 2016, 24, 1665–1674. [Google Scholar] [CrossRef] [Green Version]
- Keerthikumar, S.; Chisanga, D.; Ariyaratne, D.; Al Saffar, H.; Anand, S.; Zhao, K.; Samuel, M.; Pathan, M.; Jois, M.; Chilamkurti, N.; et al. ExoCarta: A Web-Based Compendium of Exosomal Cargo. J. Mol. Biol. 2016, 428, 688–692. [Google Scholar] [CrossRef] [Green Version]
- Blanc, L.; Vidal, M. New insights into the function of Rab GTPases in the context of exosomal secretion. Small GTPases 2018, 9, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Jin, X.; Hu, C.F.; Li, R.; Zhou, Z.; Shen, C.X. Exosomes Derived from Mesenchymal Stem Cells Rescue Myocardial Ischaemia/Reperfusion Injury by Inducing Cardiomyocyte Autophagy Via AMPK and Akt Pathways. Cell Physiol. Biochem. 2017, 43, 52–68. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Chen, Y.; Chen, Y.; Meng, Q.; Sun, J.; Shao, L.; Yu, Y.; Huang, H.; Hu, Y.; Yang, Z.; et al. MicroRNA-132, Delivered by Mesenchymal Stem Cell-Derived Exosomes, Promote Angiogenesis in Myocardial Infarction. Stem Cells Int. 2018, 2018, 3290372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Yang, J.; Yan, W.; Li, Y.; Shen, Z.; Asahara, T. Pretreatment of Cardiac Stem Cells With Exosomes Derived From Mesenchymal Stem Cells Enhances Myocardial Repair. J. Am. Heart Assoc. 2016, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Sun, X.; Cao, W.; Ma, J.; Sun, L.; Qian, H.; Zhu, W.; Xu, W. Exosomes Derived from Human Umbilical Cord Mesenchymal Stem Cells Relieve Acute Myocardial Ischemic Injury. Stem Cells Int. 2015, 2015, 761643. [Google Scholar] [CrossRef]
- Li, T.; Yan, Y.; Wang, B.; Qian, H.; Zhang, X.; Shen, L.; Wang, M.; Zhou, Y.; Zhu, W.; Li, W.; et al. Exosomes derived from human umbilical cord mesenchymal stem cells alleviate liver fibrosis. Stem Cells Dev. 2013, 22, 845–854. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Tan, Y.; Cai, M.; Zhao, T.; Mao, F.; Zhang, X.; Xu, W.; Yan, Z.; Qian, H.; Yan, Y. Human Umbilical Cord MSC-Derived Exosomes Suppress the Development of CCl4-Induced Liver Injury through Antioxidant Effect. Stem Cells Int. 2018, 2018, 6079642. [Google Scholar] [CrossRef] [Green Version]
- Qu, Y.; Zhang, Q.; Cai, X.; Li, F.; Ma, Z.; Xu, M.; Lu, L. Exosomes derived from miR-181-5p-modified adipose-derived mesenchymal stem cells prevent liver fibrosis via autophagy activation. J. Cell Mol. Med. 2017, 21, 2491–2502. [Google Scholar] [CrossRef]
- Phinney, D.G.; Pittenger, M.F. Concise Review: MSC-Derived Exosomes for Cell-Free Therapy. Stem Cells 2017, 35, 851–858. [Google Scholar] [CrossRef] [Green Version]
- Ono, M.; Kosaka, N.; Tominaga, N.; Yoshioka, Y.; Takeshita, F.; Takahashi, R.U.; Yoshida, M.; Tsuda, H.; Tamura, K.; Ochiya, T. Exosomes from bone marrow mesenchymal stem cells contain a microRNA that promotes dormancy in metastatic breast cancer cells. Sci. Signal 2014, 7, ra63. [Google Scholar] [CrossRef]
- Lee, J.K.; Park, S.R.; Jung, B.K.; Jeon, Y.K.; Lee, Y.S.; Kim, M.K.; Kim, Y.G.; Jang, J.Y.; Kim, C.W. Exosomes derived from mesenchymal stem cells suppress angiogenesis by down-regulating VEGF expression in breast cancer cells. PLoS ONE 2013, 8, e84256. [Google Scholar] [CrossRef] [Green Version]
- Qi, J.; Zhou, Y.; Jiao, Z.; Wang, X.; Zhao, Y.; Li, Y.; Chen, H.; Yang, L.; Zhu, H.; Li, Y. Exosomes Derived from Human Bone Marrow Mesenchymal Stem Cells Promote Tumor Growth Through Hedgehog Signaling Pathway. Cell Physiol. Biochem. 2017, 42, 2242–2254. [Google Scholar] [CrossRef] [PubMed]
- Reza, A.; Choi, Y.J.; Yasuda, H.; Kim, J.H. Human adipose mesenchymal stem cell-derived exosomal-miRNAs are critical factors for inducing anti-proliferation signalling to A2780 and SKOV-3 ovarian cancer cells. Sci. Rep. 2016, 6, 38498. [Google Scholar] [CrossRef] [PubMed]
- Le Blanc, K.; Davies, L.C. Mesenchymal stromal cells and the innate immune response. Immunol. Lett. 2015, 168, 140–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.; Su, J.; Roberts, A.I.; Shou, P.; Rabson, A.B.; Ren, G. How mesenchymal stem cells interact with tissue immune responses. Trends Immunol. 2012, 33, 136–143. [Google Scholar] [CrossRef] [Green Version]
- Selmani, Z.; Naji, A.; Zidi, I.; Favier, B.; Gaiffe, E.; Obert, L.; Borg, C.; Saas, P.; Tiberghien, P.; Rouas-Freiss, N.; et al. Human leukocyte antigen-G5 secretion by human mesenchymal stem cells is required to suppress T lymphocyte and natural killer function and to induce CD4+CD25highFOXP3+ regulatory T cells. Stem Cells 2008, 26, 212–222. [Google Scholar] [CrossRef]
- Chinnadurai, R.; Rajan, D.; Qayed, M.; Arafat, D.; Garcia, M.; Liu, Y.; Kugathasan, S.; Anderson, L.J.; Gibson, G.; Galipeau, J. Potency Analysis of Mesenchymal Stromal Cells Using a Combinatorial Assay Matrix Approach. Cell Rep. 2018, 22, 2504–2517. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.S.; Jang, I.K.; Lee, M.W.; Ko, Y.J.; Lee, D.H.; Lee, J.W.; Sung, K.W.; Koo, H.H.; Yoo, K.H. Enhanced Immunosuppressive Properties of Human Mesenchymal Stem Cells Primed by Interferon-gamma. EBioMedicine 2018, 28, 261–273. [Google Scholar] [CrossRef] [Green Version]
- Le Blanc, K. Immunomodulatory effects of fetal and adult mesenchymal stem cells. Cytotherapy 2003, 5, 485–489. [Google Scholar] [CrossRef]
- Li, H.; Jiang, Y.; Jiang, X.; Guo, X.; Ning, H.; Li, Y.; Liao, L.; Yao, H.; Wang, X.; Liu, Y.; et al. CCR7 guides migration of mesenchymal stem cell to secondary lymphoid organs: A novel approach to separate GvHD from GvL effect. Stem Cells 2014, 32, 1890–1903. [Google Scholar] [CrossRef]
- Le Blanc, K.; Frassoni, F.; Ball, L.; Locatelli, F.; Roelofs, H.; Lewis, I.; Lanino, E.; Sundberg, B.; Bernardo, M.E.; Remberger, M.; et al. Mesenchymal stem cells for treatment of steroid-resistant, severe, acute graft-versus-host disease: A phase II study. Lancet 2008, 371, 1579–1586. [Google Scholar]
- Herrmann, R.; Sturm, M.; Shaw, K.; Purtill, D.; Cooney, J.; Wright, M.; Phillips, M.; Cannell, P. Mesenchymal stromal cell therapy for steroid-refractory acute and chronic graft versus host disease: A phase 1 study. Int. J Hematol. 2012, 95, 182–188. [Google Scholar] [CrossRef]
- Prasad, V.K.; Lucas, K.G.; Kleiner, G.I.; Talano, J.A.; Jacobsohn, D.; Broadwater, G.; Monroy, R.; Kurtzberg, J. Efficacy and safety of ex vivo cultured adult human mesenchymal stem cells (Prochymal) in pediatric patients with severe refractory acute graft-versus-host disease in a compassionate use study. Biol. Blood Marrow Transplant. 2011, 17, 534–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Simon, J.A.; Lopez-Villar, O.; Andreu, E.J.; Rifon, J.; Muntion, S.; Diez Campelo, M.; Sanchez-Guijo, F.M.; Martinez, C.; Valcarcel, D.; Canizo, C.D. Mesenchymal stem cells expanded in vitro with human serum for the treatment of acute and chronic graft-versus-host disease: Results of a phase I/II clinical trial. Haematologica 2011, 96, 1072–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ankrum, J.A.; Dastidar, R.G.; Ong, J.F.; Levy, O.; Karp, J.M. Performance-enhanced mesenchymal stem cells via intracellular delivery of steroids. Sci. Rep. 2014, 4, 4645. [Google Scholar] [CrossRef] [Green Version]
- Dey, N.D.; Bombard, M.C.; Roland, B.P.; Davidson, S.; Lu, M.; Rossignol, J.; Sandstrom, M.I.; Skeel, R.L.; Lescaudron, L.; Dunbar, G.L. Genetically engineered mesenchymal stem cells reduce behavioral deficits in the YAC 128 mouse model of Huntington’s disease. Behav. Brain Res. 2010, 214, 193–200. [Google Scholar] [CrossRef]
- Kwon, Y.W.; Heo, S.C.; Jeong, G.O.; Yoon, J.W.; Mo, W.M.; Lee, M.J.; Jang, I.H.; Kwon, S.M.; Lee, J.S.; Kim, J.H. Tumor necrosis factor-alpha-activated mesenchymal stem cells promote endothelial progenitor cell homing and angiogenesis. Biochim. Biophys. Acta 2013, 1832, 2136–2144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsushita, T.; Kibayashi, T.; Katayama, T.; Yamashita, Y.; Suzuki, S.; Kawamata, J.; Honmou, O.; Minami, M.; Shimohama, S. Mesenchymal stem cells transmigrate across brain microvascular endothelial cell monolayers through transiently formed inter-endothelial gaps. Neurosci. Lett. 2011, 502, 41–45. [Google Scholar] [CrossRef]
- Zanier, E.R.; Pischiutta, F.; Riganti, L.; Marchesi, F.; Turola, E.; Fumagalli, S.; Perego, C.; Parotto, E.; Vinci, P.; Veglianese, P.; et al. Bone marrow mesenchymal stromal cells drive protective M2 microglia polarization after brain trauma. Neurother. J. Am. Soc. Exp. Neurother. 2014, 11, 679–695. [Google Scholar] [CrossRef] [Green Version]
- Hasan, A.; Deeb, G.; Rahal, R.; Atwi, K.; Mondello, S.; Marei, H.E.; Gali, A.; Sleiman, E. Mesenchymal Stem Cells in the Treatment of Traumatic Brain Injury. Front. Neurol. 2017, 8, 28. [Google Scholar] [CrossRef] [Green Version]
- Jo, C.H.; Chai, J.W.; Jeong, E.C.; Oh, S.; Shin, J.S.; Shim, H.; Yoon, K.S. Intra-articular Injection of Mesenchymal Stem Cells for the Treatment of Osteoarthritis of the Knee: A 2-Year Follow-up Study. Am. J. Sports Med. 2017, 45, 2774–2783. [Google Scholar] [CrossRef]
- Rizk, M.; Monaghan, M.; Shorr, R.; Kekre, N.; Bredeson, C.N.; Allan, D.S. Heterogeneity in Studies of Mesenchymal Stromal Cells to Treat or Prevent Graft-versus-Host Disease: A Scoping Review of the Evidence. Biol. Blood Marrow Transplant. 2016, 22, 1416–1423. [Google Scholar] [CrossRef] [Green Version]
- Connick, P.; Kolappan, M.; Crawley, C.; Webber, D.J.; Patani, R.; Michell, A.W.; Du, M.Q.; Luan, S.L.; Altmann, D.R.; Thompson, A.J.; et al. Autologous mesenchymal stem cells for the treatment of secondary progressive multiple sclerosis: An open-label phase 2a proof-of-concept study. Lancet. Neurol. 2012, 11, 150–156. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Niu, L.; Feng, X.; Yuan, X.; Zhao, S.; Zhang, H.; Liang, J.; Zhao, C.; Wang, H.; Hua, B.; et al. Long-term safety of umbilical cord mesenchymal stem cells transplantation for systemic lupus erythematosus: A 6-year follow-up study. Clin. Exp. Med. 2017, 17, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Sutton, M.T.; Fletcher, D.; Ghosh, S.K.; Weinberg, A.; van Heeckeren, R.; Kaur, S.; Sadeghi, Z.; Hijaz, A.; Reese, J.; Lazarus, H.M.; et al. Antimicrobial Properties of Mesenchymal Stem Cells: Therapeutic Potential for Cystic Fibrosis Infection, and Treatment. Stem Cells Int. 2016, 2016, 5303048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Ma, S.; Yang, B.; Huang, T.; Meng, N.; Xu, L.; Xing, Q.; Zhang, Y.; Zhang, K.; Li, Q.; et al. Resveratrol promotes hUC-MSCs engraftment and neural repair in a mouse model of Alzheimer’s disease. Behav. Brain Res. 2018, 339, 297–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simones, A.A.; Beisang, D.J.; Panoskaltsis-Mortari, A.; Roberts, K.D. Mesenchymal stem cells in the pathogenesis and treatment of bronchopulmonary dysplasia: A clinical review. Pediatric Res. 2018, 83, 308–317. [Google Scholar] [CrossRef] [Green Version]
- Ward, M.R.; Abadeh, A.; Connelly, K.A. Concise Review: Rational Use of Mesenchymal Stem Cells in the Treatment of Ischemic Heart Disease. Stem Cells Transl. Med. 2018, 7, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Yi, T.G.; Park, J.M.; Han, Y.M.; Kim, J.H.; Shin, D.H.; Tak, S.J.; Lee, K.; Lee, Y.S.; Jeon, M.S.; et al. Therapeutic effects of mouse bone marrow-derived clonal mesenchymal stem cells in a mouse model of inflammatory bowel disease. J. Clin. Biochem. Nutr. 2015, 57, 192–203. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Menocal, L.; Shareef, S.; Salgado, M.; Shabbir, A.; Van Badiavas, E. Role of whole bone marrow, whole bone marrow cultured cells, and mesenchymal stem cells in chronic wound healing. Stem Cell Res. Ther. 2015, 6, 24. [Google Scholar] [CrossRef] [Green Version]
- Milczarek, O.; Jarocha, D.; Starowicz-Filip, A.; Kwiatkowski, S.; Badyra, B.; Majka, M. Multiple Autologous Bone Marrow-Derived CD271(+) Mesenchymal Stem Cell Transplantation Overcomes Drug-Resistant Epilepsy in Children. Stem Cells Transl. Med. 2018, 7, 20–33. [Google Scholar] [CrossRef] [Green Version]
- Kubo, H.; Takamura, K.; Nagaya, N.; Ohgushi, H. The effect of serum on the proliferation of bone marrow-derived mesenchymal stem cells from aged donors and donors with or without chronic heart failure. J. Tissue Eng. Regen. Med. 2018, 12, e395–e397. [Google Scholar] [CrossRef]
- Wang, W.; Guo, H.; Li, H.; Yan, Y.; Wu, C.; Wang, X.; He, X.; Zhao, N. Interleukin-35 Gene-Modified Mesenchymal Stem Cells Protect Concanavalin A-Induced Fulminant Hepatitis by Decreasing the Interferon Gamma Level. Hum. Gene Ther. 2018, 29, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Perets, N.; Segal-Gavish, H.; Gothelf, Y.; Barzilay, R.; Barhum, Y.; Abramov, N.; Hertz, S.; Morozov, D.; London, M.; Offen, D. Long term beneficial effect of neurotrophic factors-secreting mesenchymal stem cells transplantation in the BTBR mouse model of autism. Behav. Brain Res. 2017, 331, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Kim, K.H.; Song, S.U.; Yi, T.G.; Yoon, S.H.; Park, S.R.; Choi, B.H. Transplantation of human bone marrow-derived clonal mesenchymal stem cells reduces fibrotic scar formation in a rat spinal cord injury model. J. Tissue Eng. Regen. Med. 2018, 12, e1034–e1045. [Google Scholar] [CrossRef]
- Galland, S.; Vuille, J.; Martin, P.; Letovanec, I.; Caignard, A.; Fregni, G.; Stamenkovic, I. Tumor-Derived Mesenchymal Stem Cells Use Distinct Mechanisms to Block the Activity of Natural Killer Cell Subsets. Cell Rep. 2017, 20, 2891–2905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fregni, G.; Quinodoz, M.; Moller, E.; Vuille, J.; Galland, S.; Fusco, C.; Martin, P.; Letovanec, I.; Provero, P.; Rivolta, C. Reciprocal modulation of mesenchymal stem cells and tumor cells promotes lung cancer metastasis. EBioMedicine 2018, 29, 128–145. [Google Scholar] [CrossRef] [Green Version]
- Timaner, M.; Letko-Khait, N.; Kotsofruk, R.; Benguigui, M.; Beyar-Katz, O.; Rachman-Tzemah, C.; Raviv, Z.; Bronshtein, T.; Machluf, M.; Shaked, Y. Therapy-Educated Mesenchymal Stem Cells Enrich for Tumor-Initiating Cells. Cancer Res. 2018, 78, 1253–1265. [Google Scholar] [CrossRef] [Green Version]
- Golay, A.; Bobbioni, E. The role of dietary fat in obesity. Int. J. Obes. Relat. Metab. Disord. 1997, 21 (Suppl. S3), S2–S11. [Google Scholar]
- Lin, H.L.; Lin, S.H.; Shen, K.P.; Chan, H.C.; Tseng, Y.H.; Yen, H.W.; Law, S.H.; Ke, L.Y. Efficiency comparison of PGBR extract and gamma-oryzanol in antioxidative stress and anti-inflammatory properties against metabolic syndrome. J. Food Biochem. 2020, 44, e13129. [Google Scholar] [CrossRef]
- Maharlooei, M.K.; Bagheri, M.; Solhjou, Z.; Jahromi, B.M.; Akrami, M.; Rohani, L.; Monabati, A.; Noorafshan, A.; Omrani, G.R. Adipose tissue derived mesenchymal stem cell (AD-MSC) promotes skin wound healing in diabetic rats. Diabetes. Res. Clin. Pract. 2011, 93, 228–234. [Google Scholar] [CrossRef]
- King, L.K.; March, L.; Anandacoomarasamy, A. Obesity & osteoarthritis. Indian J. Med. Res. 2013, 138, 185–193. [Google Scholar]
- Gersing, A.S.; Solka, M.; Joseph, G.B.; Schwaiger, B.J.; Heilmeier, U.; Feuerriegel, G.; Nevitt, M.C.; McCulloch, C.E.; Link, T.M. Progression of cartilage degeneration and clinical symptoms in obese and overweight individuals is dependent on the amount of weight loss: 48-month data from the Osteoarthritis Initiative. Osteoarthr. Cartil. 2016, 24, 1126–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francis, S.L.; Duchi, S.; Onofrillo, C.; Di Bella, C.; Choong, P.F.M. Adipose-Derived Mesenchymal Stem Cells in the Use of Cartilage Tissue Engineering: The Need for a Rapid Isolation Procedure. Stem Cells Int. 2018, 2018, 8947548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunnell, B.A.; Flaat, M.; Gagliardi, C.; Patel, B.; Ripoll, C. Adipose-derived stem cells: Isolation, expansion and differentiation. Methods 2008, 45, 115–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serhal, R.; Saliba, N.; Hilal, G.; Moussa, M.; Hassan, G.S.; El Atat, O.; Alaaeddine, N. Effect of adipose-derived mesenchymal stem cells on hepatocellular carcinoma: In vitro inhibition of carcinogenesis. World J. Gastroenterol. 2019, 25, 567–583. [Google Scholar] [CrossRef]
- Chui, P.C.; Guan, H.P.; Lehrke, M.; Lazar, M.A. PPARgamma regulates adipocyte cholesterol metabolism via oxidized LDL receptor 1. J. Clin. Investig. 2005, 115, 2244–2256. [Google Scholar] [CrossRef] [Green Version]
- Giralt, M.; Cereijo, R.; Villarroya, F. Adipokines and the Endocrine Role of Adipose Tissues. Handb. Exp. Pharmacol. 2016, 233, 265–282. [Google Scholar]
- Chen, G.; Deng, C.; Li, Y.P. TGF-beta and BMP signaling in osteoblast differentiation and bone formation. Int. J. Biol. Sci. 2012, 8, 272–288. [Google Scholar] [CrossRef] [Green Version]
- Kang, Q.; Song, W.X.; Luo, Q.; Tang, N.; Luo, J.; Luo, X.; Chen, J.; Bi, Y.; He, B.C.; Park, J.K.; et al. A comprehensive analysis of the dual roles of BMPs in regulating adipogenic and osteogenic differentiation of mesenchymal progenitor cells. Stem Cells Dev. 2009, 18, 545–559. [Google Scholar] [CrossRef] [Green Version]
- Van Zoelen, E.J.; Duarte, I.; Hendriks, J.M.; van der Woning, S.P. TGFbeta-induced switch from adipogenic to osteogenic differentiation of human mesenchymal stem cells: Identification of drug targets for prevention of fat cell differentiation. Stem Cell Res. Ther. 2016, 7, 123. [Google Scholar] [CrossRef] [Green Version]
- Wang, E.A.; Israel, D.I.; Kelly, S.; Luxenberg, D.P. Bone morphogenetic protein-2 causes commitment and differentiation in C3H10T1/2 and 3T3 cells. Growth Factors 1993, 9, 57–71. [Google Scholar] [CrossRef]
- Huang, H.; Song, T.J.; Li, X.; Hu, L.; He, Q.; Liu, M.; Lane, M.D.; Tang, Q.Q. BMP signaling pathway is required for commitment of C3H10T1/2 pluripotent stem cells to the adipocyte lineage. Proc. Natl. Acad. Sci. USA 2009, 106, 12670–12675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowers, R.R.; Kim, J.W.; Otto, T.C.; Lane, M.D. Stable stem cell commitment to the adipocyte lineage by inhibition of DNA methylation: Role of the BMP-4 gene. Proc. Natl. Acad. Sci. USA 2006, 103, 13022–13027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tontonoz, P.; Hu, E.; Spiegelman, B.M. Stimulation of adipogenesis in fibroblasts by PPAR gamma 2, a lipid-activated transcription factor. Cell 1994, 79, 1147–1156. [Google Scholar] [CrossRef]
- Barak, Y.; Nelson, M.C.; Ong, E.S.; Jones, Y.Z.; Ruiz-Lozano, P.; Chien, K.R.; Koder, A.; Evans, R.M. PPAR gamma is required for placental, cardiac, and adipose tissue development. Mol. Cell 1999, 4, 585–595. [Google Scholar] [CrossRef]
- Rosen, E.D.; Sarraf, P.; Troy, A.E.; Bradwin, G.; Moore, K.; Milstone, D.S.; Spiegelman, B.M.; Mortensen, R.M. PPAR gamma is required for the differentiation of adipose tissue in vivo and in vitro. Mol. Cell 1999, 4, 611–617. [Google Scholar] [CrossRef]
- Wang, F.; Mullican, S.E.; DiSpirito, J.R.; Peed, L.C.; Lazar, M.A. Lipoatrophy and severe metabolic disturbance in mice with fat-specific deletion of PPARgamma. Proc. Natl. Acad. Sci. USA 2013, 110, 18656–18661. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Rosen, E.D.; Brun, R.; Hauser, S.; Adelmant, G.; Troy, A.E.; McKeon, C.; Darlington, G.J.; Spiegelman, B.M. Cross-regulation of C/EBP alpha and PPAR gamma controls the transcriptional pathway of adipogenesis and insulin sensitivity. Mol. Cell 1999, 3, 151–158. [Google Scholar] [CrossRef]
- Freytag, S.O.; Paielli, D.L.; Gilbert, J.D. Ectopic expression of the CCAAT/enhancer-binding protein alpha promotes the adipogenic program in a variety of mouse fibroblastic cells. Genes Dev. 1994, 8, 1654–1663. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Bucher, N.L.; Farmer, S.R. Induction of peroxisome proliferator-activated receptor gamma during the conversion of 3T3 fibroblasts into adipocytes is mediated by C/EBPbeta, C/EBPdelta, and glucocorticoids. Mol. Cell Biol. 1996, 16, 4128–4136. [Google Scholar] [CrossRef] [Green Version]
- Choe, S.S.; Huh, J.Y.; Hwang, I.J.; Kim, J.I.; Kim, J.B. Adipose Tissue Remodeling: Its Role in Energy Metabolism and Metabolic Disorders. Front. Endocrinol. 2016, 7, 30. [Google Scholar] [CrossRef] [Green Version]
- Boutens, L.; Stienstra, R. Adipose tissue macrophages: Going off track during obesity. Diabetologia 2016, 59, 879–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guilherme, A.; Virbasius, J.V.; Puri, V.; Czech, M.P. Adipocyte dysfunctions linking obesity to insulin resistance and type 2 diabetes. Nat. Rev. Mol. Cell Biol. 2008, 9, 367–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bluher, M. Adipose tissue dysfunction in obesity. Exp. Clin. Endocrinol. Diabetes 2009, 117, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Ke, L.Y.; Chan, H.C.; Chan, H.C.; Kalu, F.C.U.; Lee, H.C.; Lin, I.L.; Jhuo, S.J.; Lai, W.T.; Tsao, C.R.; Sawamura, T.; et al. Electronegative Low-Density Lipoprotein L5 Induces Adipose Tissue Inflammation Associated With Metabolic Syndrome. J. Clin. Endocrinol. Metab. 2017, 102, 4615–4625. [Google Scholar] [CrossRef]
- Kattoor, A.J.; Goel, A.; Mehta, J.L. LOX-1: Regulation, Signaling and Its Role in Atherosclerosis. Antioxidants 2019, 8, 218. [Google Scholar] [CrossRef] [Green Version]
- Manteiga, S.; Choi, K.; Jayaraman, A.; Lee, K. Systems biology of adipose tissue metabolism: Regulation of growth, signaling and inflammation. Wiley Interdiscip. Rev. Syst. Biol. Med. 2013, 5, 425–447. [Google Scholar] [CrossRef]
- Waki, H.; Yamauchi, T.; Kadowaki, T. Regulation of differentiation and hypertrophy of adipocytes and adipokine network by PPARgamma. Nihon Rinsho Jpn. J. Clin. Med. 2010, 68, 210–216. [Google Scholar]
- Minteer, D.; Marra, K.G.; Rubin, J.P. Adipose-derived mesenchymal stem cells: Biology and potential applications. Adv. Biochem. Eng. Biotechnol. 2013, 129, 59–71. [Google Scholar]
- Barbagallo, I.; Li Volti, G.; Galvano, F.; Tettamanti, G.; Pluchinotta, F.R.; Bergante, S.; Vanella, L. Diabetic human adipose tissue-derived mesenchymal stem cells fail to differentiate in functional adipocytes. Exp. Biol. Med. 2017, 242, 1079–1085. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Tie, G.; Wang, S.; Messina, K.E.; DiDato, S.; Guo, S.; Messina, L.M. Type 2 diabetes restricts multipotency of mesenchymal stem cells and impairs their capacity to augment postischemic neovascularization in db/db mice. J. Am. Heart Assoc. 2012, 1, e002238. [Google Scholar] [CrossRef] [Green Version]
- Schroder, K.; Wandzioch, K.; Helmcke, I.; Brandes, R.P. Nox4 acts as a switch between differentiation and proliferation in preadipocytes. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 239–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahadev, K.; Motoshima, H.; Wu, X.; Ruddy, J.M.; Arnold, R.S.; Cheng, G.; Lambeth, J.D.; Goldstein, B.J. The NAD(P)H oxidase homolog Nox4 modulates insulin-stimulated generation of H2O2 and plays an integral role in insulin signal transduction. Mol. Cell Biol. 2004, 24, 1844–1854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellis, J.R.; McDonald, R.B.; Stern, J.S. A diet high in fat stimulates adipocyte proliferation in older (22 month) rats. Exp. Gerontol. 1990, 25, 141–148. [Google Scholar] [CrossRef]
- Johnson, P.R.; Stern, J.S.; Greenwood, M.R.; Hirsch, J. Adipose tissue hyperplasia and hyperinsulinemia on Zucker obese female rats: A developmental study. Metabolism 1978, 27 (Suppl. S2), 1941–1954. [Google Scholar] [CrossRef]
- Rigamonti, A.; Brennand, K.; Lau, F.; Cowan, C.A. Rapid cellular turnover in adipose tissue. PLoS ONE 2011, 6, e17637. [Google Scholar] [CrossRef] [Green Version]
- Hood, R.L.; Allen, C.E. Cellularity of porcine adipose tissue: Effects of growth and adiposity. J. Lipid Res. 1977, 18, 275–284. [Google Scholar]
- Salans, L.B.; Horton, E.S.; Sims, E.A. Experimental obesity in man: Cellular character of the adipose tissue. J. Clin. Investig. 1971, 50, 1005–1011. [Google Scholar] [CrossRef]
- Salans, L.B.; Cushman, S.W.; Weismann, R.E. Studies of human adipose tissue. Adipose cell size and number in nonobese and obese patients. Adipose cell size and number in nonobese and obese patients. J. Clin. Investig. 1973, 52, 929–941. [Google Scholar] [CrossRef] [Green Version]
- Spalding, K.L.; Arner, E.; Westermark, P.O.; Bernard, S.; Buchholz, B.A.; Bergmann, O.; Blomqvist, L.; Hoffstedt, J.; Naslund, E.; Britton, T.; et al. Dynamics of fat cell turnover in humans. Nature 2008, 453, 783–787. [Google Scholar] [CrossRef]
- Steinhauser, M.L.; Lechene, C.P. Quantitative imaging of subcellular metabolism with stable isotopes and multi-isotope imaging mass spectrometry. Semin. Cell Dev. Biol. 2013, 24, 661–667. [Google Scholar] [CrossRef] [Green Version]
- Steinhauser, M.L.; Bailey, A.P.; Senyo, S.E.; Guillermier, C.; Perlstein, T.S.; Gould, A.P.; Lee, R.T.; Lechene, C.P. Multi-isotope imaging mass spectrometry quantifies stem cell division and metabolism. Nature 2012, 481, 516–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, B.; Lu, Y.; Hajifathalian, K.; Bentham, J.; Di Cesare, M.; Danaei, G.; Lo, W.C. Worldwide trends in diabetes since 1980: A pooled analysis of 751 population-based studies with 4.4 million participants. Lancet 2016, 387, 1513–1530. [Google Scholar] [CrossRef] [Green Version]
- Keane, K.N.; Calton, E.K.; Carlessi, R.; Hart, P.H.; Newsholme, P. The bioenergetics of inflammation: Insights into obesity and type 2 diabetes. Eur. J. Clin. Nutr. 2017, 71, 904–912. [Google Scholar] [CrossRef] [PubMed]
- Karnieli, O.; Izhar-Prato, Y.; Bulvik, S.; Efrat, S. Generation of insulin-producing cells from human bone marrow mesenchymal stem cells by genetic manipulation. Stem Cells 2007, 25, 2837–2844. [Google Scholar] [CrossRef]
- Ding, Y.; Xu, D.; Feng, G.; Bushell, A.; Muschel, R.J.; Wood, K.J. Mesenchymal stem cells prevent the rejection of fully allogenic islet grafts by the immunosuppressive activity of matrix metalloproteinase-2 and -9. Diabetes 2009, 58, 1797–1806. [Google Scholar] [CrossRef] [Green Version]
- Berman, D.M.; Willman, M.A.; Han, D.; Kleiner, G.; Kenyon, N.M.; Cabrera, O.; Karl, J.A.; Wiseman, R.W.; O’Connor, D.H.; Bartholomew, A.M.; et al. Mesenchymal stem cells enhance allogeneic islet engraftment in nonhuman primates. Diabetes 2010, 59, 2558–2568. [Google Scholar] [CrossRef] [Green Version]
- Lu, D.; Chen, B.; Liang, Z.; Deng, W.; Jiang, Y.; Li, S.; Xu, J.; Wu, Q.; Zhang, Z.; Xie, B.; et al. Comparison of bone marrow mesenchymal stem cells with bone marrow-derived mononuclear cells for treatment of diabetic critical limb ischemia and foot ulcer: A double-blind, randomized, controlled trial. Diabetes Res. Clin. Pract. 2011, 92, 26–36. [Google Scholar] [CrossRef]
- Caplan, A.I.; Dennis, J.E. Mesenchymal stem cells as trophic mediators. J. Cell. Biochem. 2006, 98, 1076–1084. [Google Scholar] [CrossRef]
- Si, Y.; Zhao, Y.; Hao, H.; Liu, J.; Guo, Y.; Mu, Y.; Shen, J.; Cheng, Y.; Fu, X.; Han, W. Infusion of mesenchymal stem cells ameliorates hyperglycemia in type 2 diabetic rats: Identification of a novel role in improving insulin sensitivity. Diabetes 2012, 61, 1616–1625. [Google Scholar] [CrossRef] [Green Version]
- Hao, H.; Liu, J.; Shen, J.; Zhao, Y.; Liu, H.; Hou, Q.; Tong, C.; Ti, D.; Dong, L.; Cheng, Y.; et al. Multiple intravenous infusions of bone marrow mesenchymal stem cells reverse hyperglycemia in experimental type 2 diabetes rats. Biochem. Biophys. Res. Commun. 2013, 436, 418–423. [Google Scholar] [CrossRef]
- Gao, X.; Song, L.; Shen, K.; Wang, H.; Qian, M.; Niu, W.; Qin, X. Bone marrow mesenchymal stem cells promote the repair of islets from diabetic mice through paracrine actions. Mol. Cell. Endocrinol. 2014, 388, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.H.; Seo, M.J.; Reger, R.L.; Spees, J.L.; Pulin, A.A.; Olson, S.D.; Prockop, D.J. Multipotent stromal cells from human marrow home to and promote repair of pancreatic islets and renal glomeruli in diabetic NOD/scid mice. Proc. Nat. Acad. Sci. USA 2006, 103, 17438–17443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Liao, L.; Tan, J. Mesenchymal-stem-cell-based experimental and clinical trials: Current status and open questions. Expert Opin. Biol. Ther. 2011, 11, 893–909. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.R.; Pollock, K.; Hubel, A.; McKenna, D. Mesenchymal stem or stromal cells: A review of clinical applications and manufacturing practices. Transfusion 2014, 54, 1418–1437. [Google Scholar] [CrossRef] [PubMed]
- Ringden, O.; Uzunel, M.; Rasmusson, I.; Remberger, M.; Sundberg, B.; Lonnies, H.; Marschall, H.U.; Dlugosz, A.; Szakos, A.; Hassan, Z.; et al. Mesenchymal stem cells for treatment of therapy-resistant graft-versus-host disease. Transplantation 2006, 81, 1390–1397. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Hui, H.; Jia, X.; Zhang, J.; Liu, Y.; Xu, Q.; Zhu, D. Infusion with Human Bone Marrow-derived Mesenchymal Stem Cells Improves beta-cell Function in Patients and Non-obese Mice with Severe Diabetes. Sci. Rep. 2016, 6, 37894. [Google Scholar] [CrossRef] [Green Version]
- Bhansali, A.; Asokumar, P.; Walia, R.; Bhansali, S.; Gupta, V.; Jain, A.; Sachdeva, N.; Sharma, R.R.; Marwaha, N.; Khandelwal, N. Efficacy and safety of autologous bone marrow-derived stem cell transplantation in patients with type 2 diabetes mellitus: A randomized placebo-controlled study. Cell Transplant. 2014, 23, 1075–1085. [Google Scholar] [CrossRef]
- Jiang, R.; Han, Z.; Zhuo, G.; Qu, X.; Li, X.; Wang, X.; Shao, Y.; Yang, S.; Han, Z.C. Transplantation of placenta-derived mesenchymal stem cells in type 2 diabetes: A pilot study. Front. Med. 2011, 5, 94–100. [Google Scholar] [CrossRef]
- Kong, D.; Zhuang, X.; Wang, D.; Qu, H.; Jiang, Y.; Li, X.; Wu, W.; Xiao, J.; Liu, X.; Liu, J.; et al. Umbilical cord mesenchymal stem cell transfusion ameliorated hyperglycemia in patients with type 2 diabetes mellitus. Clin. Lab. 2014, 60, 1969–1976. [Google Scholar] [CrossRef]
- Hu, J.; Wang, Y.; Gong, H.; Yu, C.; Guo, C.; Wang, F.; Yan, S.; Xu, H. Long term effect and safety of Wharton’s jelly-derived mesenchymal stem cells on type 2 diabetes. Exp. Ther. Med. 2016, 12, 1857–1866. [Google Scholar] [CrossRef]
- Bhansali, A.; Upreti, V.; Khandelwal, N.; Marwaha, N.; Gupta, V.; Sachdeva, N.; Sharma, R.R.; Saluja, K.; Dutta, P.; Walia, R.; et al. Efficacy of autologous bone marrow-derived stem cell transplantation in patients with type 2 diabetes mellitus. Stem Cells Dev. 2009, 18, 1407–1416. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.B.; Jiang, X.B.; Yang, L. Differentiation of rat marrow mesenchymal stem cells into pancreatic islet beta-cells. World J. Gastroenterol. 2004, 10, 3016–3020. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.H.; Liu, C.P.; Xu, K.F.; Mao, X.D.; Zhu, J.; Jiang, J.J.; Cui, D.; Zhang, M.; Xu, Y.; Liu, C. Reversal of hyperglycemia in diabetic rats by portal vein transplantation of islet-like cells generated from bone marrow mesenchymal stem cells. World J. Gastroenterol. 2007, 13, 3342–3349. [Google Scholar] [CrossRef] [PubMed]
- Chandravanshi, B.; Bhonde, R.R. Shielding Engineered Islets with Mesenchymal Stem Cells Enhance Survival Under Hypoxia. J. Cell. Biochem. 2017, 118, 2672–2683. [Google Scholar] [CrossRef] [PubMed]
- Zang, L.; Hao, H.; Liu, J.; Li, Y.; Han, W.; Mu, Y. Mesenchymal stem cell therapy in type 2 diabetes mellitus. Diabetology Metab. Syndr. 2017, 9, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estrada, E.J.; Valacchi, F.; Nicora, E.; Brieva, S.; Esteve, C.; Echevarria, L.; Froud, T.; Bernetti, K.; Cayetano, S.M.; Velazquez, O.; et al. Combined treatment of intrapancreatic autologous bone marrow stem cells and hyperbaric oxygen in type 2 diabetes mellitus. Cell Transplant. 2008, 17, 1295–1304. [Google Scholar] [CrossRef]
- Wu, Z.; Cai, J.; Chen, J.; Huang, L.; Wu, W.; Luo, F.; Wu, C.; Liao, L.; Tan, J. Autologous bone marrow mononuclear cell infusion and hyperbaric oxygen therapy in type 2 diabetes mellitus: An open-label, randomized controlled clinical trial. Cytotherapy 2014, 16, 258–265. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Zheng, P.; Wang, X.; Dai, G.; Cheng, H.; Zhang, Z.; Hua, R.; Niu, X.; Shi, J.; An, Y. A preliminary evaluation of efficacy and safety of Wharton’s jelly mesenchymal stem cell transplantation in patients with type 2 diabetes mellitus. Stem Cell Res. Ther. 2014, 5, 57. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Li, C.; Wang, L.; Zhang, X.; Zhang, M.; Gao, H.; Yu, X.; Wang, F.; Zhao, W.; Yan, S.; et al. Long term effects of the implantation of autologous bone marrow mononuclear cells for type 2 diabetes mellitus. Endocr. J. 2012, 59, 1031–1039. [Google Scholar] [CrossRef] [Green Version]
- Sood, V.; Mittal, B.R.; Bhansali, A.; Singh, B.; Khandelwal, N.; Marwaha, N.; Jain, A. Biodistribution of 18F-FDG-Labeled Autologous Bone Marrow-Derived Stem Cells in Patients With Type 2 Diabetes Mellitus: Exploring Targeted and Intravenous Routes of Delivery. Clin. Nucl. Med. 2015, 40, 697–700. [Google Scholar] [CrossRef]
- Skyler, J.S.; Fonseca, V.A.; Segal, K.R.; Rosenstock, J.; Investigators, M.-D. Allogeneic Mesenchymal Precursor Cells in Type 2 Diabetes: A Randomized, Placebo-Controlled, Dose-Escalation Safety and Tolerability Pilot Study. Diabetes Care 2015, 38, 1742–1749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thom, S.R.; Bhopale, V.M.; Velazquez, O.C.; Goldstein, L.J.; Thom, L.H.; Buerk, D.G. Stem cell mobilization by hyperbaric oxygen. Am. J. Physiol. Heart Circ. Physiol. 2006, 290, H1378–H1386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Zhao, S.; Mao, H.; Zhou, L.; Wang, Z.J.; Wang, H.X. Autologous bone marrow stem cell transplantation for the treatment of type 2 diabetes mellitus. Chin. Med. J. 2011, 124, 3622–3628. [Google Scholar] [PubMed]
- Zhao, Y.; Jiang, Z.; Zhao, T.; Ye, M.; Hu, C.; Zhou, H.; Yin, Z.; Chen, Y.; Zhang, Y.; Wang, S.; et al. Targeting insulin resistance in type 2 diabetes via immune modulation of cord blood-derived multipotent stem cells (CB-SCs) in stem cell educator therapy: Phase I/II clinical trial. BMC Med. 2013, 11, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhansali, S.; Dutta, P.; Kumar, V.; Yadav, M.K.; Jain, A.; Mudaliar, S.; Bhansali, S.; Sharma, R.R.; Jha, V.; Marwaha, N.; et al. Efficacy of Autologous Bone Marrow-Derived Mesenchymal Stem Cell and Mononuclear Cell Transplantation in Type 2 Diabetes Mellitus: A Randomized, Placebo-Controlled Comparative Study. Stem Cells Dev. 2017, 26, 471–481. [Google Scholar] [CrossRef]
- Kinnaird, T.; Stabile, E.; Burnett, M.S.; Epstein, S.E. Bone-marrow-derived cells for enhancing collateral development: Mechanisms, animal data, and initial clinical experiences. Circ. Res. 2004, 95, 354–363. [Google Scholar] [CrossRef]
- Guan, L.X.; Guan, H.; Li, H.B.; Ren, C.A.; Liu, L.; Chu, J.J.; Dai, L.J. Therapeutic efficacy of umbilical cord-derived mesenchymal stem cells in patients with type 2 diabetes. Exp. Ther. Med. 2015, 9, 1623–1630. [Google Scholar] [CrossRef]
- El-Jawhari, J.J.; El-Sherbiny, Y.M.; Jones, E.A.; McGonagle, D. Mesenchymal stem cells, autoimmunity and rheumatoid arthritis. QJM 2014, 107, 505–514. [Google Scholar] [CrossRef] [Green Version]
- Maria, A.T.; Maumus, M.; Le Quellec, A.; Jorgensen, C.; Noel, D.; Guilpain, P. Adipose-Derived Mesenchymal Stem Cells in Autoimmune Disorders: State of the Art and Perspectives for Systemic Sclerosis. Clin. Rev. Allergy Immunol. 2017, 52, 234–259. [Google Scholar] [CrossRef]
- Giacoppo, S.; Bramanti, P.; Mazzon, E. The transplantation of mesenchymal stem cells derived from unconventional sources: An innovative approach to multiple sclerosis therapy. Arch. Immunol. Ther. Exp. 2017, 65, 363–379. [Google Scholar] [CrossRef]
- Guney, M.A.; Gannon, M. Pancreas cell fate. Birth Defects Res. Part C Embryo Today Rev. 2009, 87, 232–248. [Google Scholar] [CrossRef] [PubMed]
- Moriscot, C.; de Fraipont, F.; Richard, M.J.; Marchand, M.; Savatier, P.; Bosco, D.; Favrot, M.; Benhamou, P.Y. Human bone marrow mesenchymal stem cells can express insulin and key transcription factors of the endocrine pancreas developmental pathway upon genetic and/or microenvironmental manipulation in vitro. Stem Cells 2005, 23, 594–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Q.P.; Huang, H.; Xu, B.; Dong, X.; Gao, S.L.; Zhang, B.; Wu, Y.L. Human bone marrow mesenchymal stem cells differentiate into insulin-producing cells upon microenvironmental manipulation in vitro. Differ. Res. Biol. Divers. 2009, 77, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Nam, J.S.; Kang, H.M.; Kim, J.; Park, S.; Kim, H.; Ahn, C.W.; Park, J.O.; Kim, K.R. Transplantation of insulin-secreting cells differentiated from human adipose tissue-derived stem cells into type 2 diabetes mice. Biochem. Biophys. Res. Commun. 2014, 443, 775–781. [Google Scholar] [CrossRef]
- Asumda, F.Z. Age-associated changes in the ecological niche: Implications for mesenchymal stem cell aging. Stem Cell Res. Ther. 2013, 4, 47. [Google Scholar] [CrossRef] [Green Version]
- Lai, Y.; Sun, Y.; Skinner, C.M.; Son, E.L.; Lu, Z.; Tuan, R.S.; Jilka, R.L.; Ling, J.; Chen, X.D. Reconstitution of marrow-derived extracellular matrix ex vivo: A robust culture system for expanding large-scale highly functional human mesenchymal stem cells. Stem Cells Dev. 2010, 19, 1095–1107. [Google Scholar] [CrossRef] [Green Version]
- Block, T.J.; Marinkovic, M.; Tran, O.N.; Gonzalez, A.O.; Marshall, A.; Dean, D.D.; Chen, X.D. Restoring the quantity and quality of elderly human mesenchymal stem cells for autologous cell-based therapies. Stem Cell Res. Ther. 2017, 8, 239. [Google Scholar] [CrossRef] [Green Version]
- Wei, W.; Huang, Y.; Li, D.; Gou, H.F.; Wang, W. Improved therapeutic potential of MSCs by genetic modification. Gene Ther. 2018, 25, 538–547. [Google Scholar] [CrossRef]
- Francois, M.; Copland, I.B.; Yuan, S.; Romieu-Mourez, R.; Waller, E.K.; Galipeau, J. Cryopreserved mesenchymal stromal cells display impaired immunosuppressive properties as a result of heat-shock response and impaired interferon-gamma licensing. Cytotherapy 2012, 14, 147–152. [Google Scholar] [CrossRef] [Green Version]
- Cruz, F.F.; Borg, Z.D.; Goodwin, M.; Sokocevic, D.; Wagner, D.; McKenna, D.H.; Rocco, P.R.; Weiss, D.J. Freshly thawed and continuously cultured human bone marrow-derived mesenchymal stromal cells comparably ameliorate allergic airways inflammation in immunocompetent mice. Stem Cells Transl. Med. 2015, 4, 615–624. [Google Scholar] [CrossRef]
- Sart, S.; Ma, T.; Li, Y. Preconditioning stem cells for in vivo delivery. Biores. Open Access 2014, 3, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Pei, M. Environmental preconditioning rejuvenates adult stem cells’ proliferation and chondrogenic potential. Biomaterials 2017, 117, 10–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fehrer, C.; Brunauer, R.; Laschober, G.; Unterluggauer, H.; Reitinger, S.; Kloss, F.; Gully, C.; Gassner, R.; Lepperdinger, G. Reduced oxygen tension attenuates differentiation capacity of human mesenchymal stem cells and prolongs their lifespan. Aging Cell 2007, 6, 745–757. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Yu, S.P.; Fraser, J.L.; Lu, Z.; Ogle, M.E.; Wang, J.A.; Wei, L. Transplantation of hypoxia-preconditioned mesenchymal stem cells improves infarcted heart function via enhanced survival of implanted cells and angiogenesis. J. Thorac. Cardiovasc. Surg. 2008, 135, 799–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.S.; Lee, J.H.; Yoon, Y.M.; Yun, C.W.; Noh, H.; Lee, S.H. Hypoxia-induced expression of cellular prion protein improves the therapeutic potential of mesenchymal stem cells. Cell Death Dis. 2016, 7, e2395. [Google Scholar] [CrossRef] [PubMed]
- Bonnard, C.; Durand, A.; Peyrol, S.; Chanseaume, E.; Chauvin, M.A.; Morio, B.; Vidal, H.; Rieusset, J. Mitochondrial dysfunction results from oxidative stress in the skeletal muscle of diet-induced insulin-resistant mice. J. Clin. Investig. 2008, 118, 789–800. [Google Scholar] [CrossRef]
- Minamino, T.; Orimo, M.; Shimizu, I.; Kunieda, T.; Yokoyama, M.; Ito, T.; Nojima, A.; Nabetani, A.; Oike, Y.; Matsubara, H.; et al. A crucial role for adipose tissue p53 in the regulation of insulin resistance. Nat. Med. 2009, 15, 1082–1087. [Google Scholar] [CrossRef]
- Amiri, F.; Jahanian-Najafabadi, A.; Roudkenar, M.H. In vitro augmentation of mesenchymal stem cells viability in stressful microenvironments: In vitro augmentation of mesenchymal stem cells viability. Cell Stress Chaperones 2015, 20, 237–251. [Google Scholar] [CrossRef] [Green Version]
- Boregowda, S.V.; Krishnappa, V.; Chambers, J.W.; Lograsso, P.V.; Lai, W.T.; Ortiz, L.A.; Phinney, D.G. Atmospheric oxygen inhibits growth and differentiation of marrow-derived mouse mesenchymal stem cells via a p53-dependent mechanism: Implications for long-term culture expansion. Stem Cells 2012, 30, 975–987. [Google Scholar] [CrossRef] [Green Version]
- Cheng, N.C.; Hsieh, T.Y.; Lai, H.S.; Young, T.H. High glucose-induced reactive oxygen species generation promotes stemness in human adipose-derived stem cells. Cytotherapy 2016, 18, 371–383. [Google Scholar] [CrossRef]
- Lian, Q.; Zhang, Y.; Zhang, J.; Zhang, H.K.; Wu, X.; Zhang, Y.; Lam, F.F.; Kang, S.; Xia, J.C.; Lai, W.H.; et al. Functional mesenchymal stem cells derived from human induced pluripotent stem cells attenuate limb ischemia in mice. Circulation 2010, 121, 1113–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, W.M.; Liu, W.; Bi, Y.W.; He, X.P.; Sun, W.Y.; Pang, X.Y.; Gu, X.H.; Wang, X.P. Mesenchymal stem cells differentiate into an endothelial phenotype, reduce neointimal formation, and enhance endothelial function in a rat vein grafting model. Stem Cells Dev. 2008, 17, 785–793. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Chen, L.; Scott, P.G.; Tredget, E.E. Mesenchymal stem cells enhance wound healing through differentiation and angiogenesis. Stem Cells 2007, 25, 2648–2659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinnaird, T.; Stabile, E.; Burnett, M.S.; Lee, C.W.; Barr, S.; Fuchs, S.; Epstein, S.E. Marrow-derived stromal cells express genes encoding a broad spectrum of arteriogenic cytokines and promote in vitro and in vivo arteriogenesis through paracrine mechanisms. Circ. Res. 2004, 94, 678–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinnaird, T.; Stabile, E.; Burnett, M.S.; Shou, M.; Lee, C.W.; Barr, S.; Fuchs, S.; Epstein, S.E. Local delivery of marrow-derived stromal cells augments collateral perfusion through paracrine mechanisms. Circulation 2004, 109, 1543–1549. [Google Scholar] [CrossRef] [PubMed]
- Hudak, C.S.; Gulyaeva, O.; Wang, Y.; Park, S.M.; Lee, L.; Kang, C.; Sul, H.S. Pref-1 marks very early mesenchymal precursors required for adipose tissue development and expansion. Cell Rep. 2014, 8, 678–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
The central role of Mesenchymal Stem Cells (MSC) in immune responses. The above figure distinguishes the response and interaction of MSCs in pro-inflammatory and anti-inflammatory conditions on immune cells. These effects demonstrate cell-to-cell contact-mediated immunosuppression of B and T cell proliferation, induction and transforming growth factor-β (TGF-β)/hepatocyte growth factor (HGF) mediated regulation of regulatory T cells. Also, it shows the capacity of immunomodulation of MSCs by inhibiting the natural killer (NK) cells, dendritic cells (DCs) at various maturation stages as well as macrophage polarization dependency on the microenvironment.
Figure 1.
The central role of Mesenchymal Stem Cells (MSC) in immune responses. The above figure distinguishes the response and interaction of MSCs in pro-inflammatory and anti-inflammatory conditions on immune cells. These effects demonstrate cell-to-cell contact-mediated immunosuppression of B and T cell proliferation, induction and transforming growth factor-β (TGF-β)/hepatocyte growth factor (HGF) mediated regulation of regulatory T cells. Also, it shows the capacity of immunomodulation of MSCs by inhibiting the natural killer (NK) cells, dendritic cells (DCs) at various maturation stages as well as macrophage polarization dependency on the microenvironment.

Figure 2.
The autocrine and paracrine functions of MSC. The left side depicts the cyclooxygenase-2 (COX-2)/prostaglandin E2 (PGE2) axis for the maintenance of an autocrine/paracrine loop and COX-2 mediated PGE2 production in MSCs as a response to the surrounding microenvironment. The right side of the figure demonstrates the dominance of MSCs on immune cells (inhibiting cardiomyocyte apoptosis and DC differentiation, also promoting M2 macrophage polarization and T-reg cell proliferation) by producing several immunomodulatory factors and chemokines. Note: Figure represents both conditioned/modified and natural MSCs.
Figure 2.
The autocrine and paracrine functions of MSC. The left side depicts the cyclooxygenase-2 (COX-2)/prostaglandin E2 (PGE2) axis for the maintenance of an autocrine/paracrine loop and COX-2 mediated PGE2 production in MSCs as a response to the surrounding microenvironment. The right side of the figure demonstrates the dominance of MSCs on immune cells (inhibiting cardiomyocyte apoptosis and DC differentiation, also promoting M2 macrophage polarization and T-reg cell proliferation) by producing several immunomodulatory factors and chemokines. Note: Figure represents both conditioned/modified and natural MSCs.

Figure 3.
Progression of adipogenic MSCs differentiation and maturation into mature fat cells due to an excess of calories.
Figure 3.
Progression of adipogenic MSCs differentiation and maturation into mature fat cells due to an excess of calories.

Figure 4.
The above figure demonstrates the failure of overnutrition accommodation resulting in adipose tissue dysfunction. Insufficient pre-adipocytes pressure the matured adipose cells to undergo hypertrophy resulting in adipocyte fibrosis (macrophage accumulation and collagen deposition). Furthermore, these cells progress towards adipose dysfunction (ectopic lipid deposition leads to hypoxia and necrosis).
Figure 4.
The above figure demonstrates the failure of overnutrition accommodation resulting in adipose tissue dysfunction. Insufficient pre-adipocytes pressure the matured adipose cells to undergo hypertrophy resulting in adipocyte fibrosis (macrophage accumulation and collagen deposition). Furthermore, these cells progress towards adipose dysfunction (ectopic lipid deposition leads to hypoxia and necrosis).


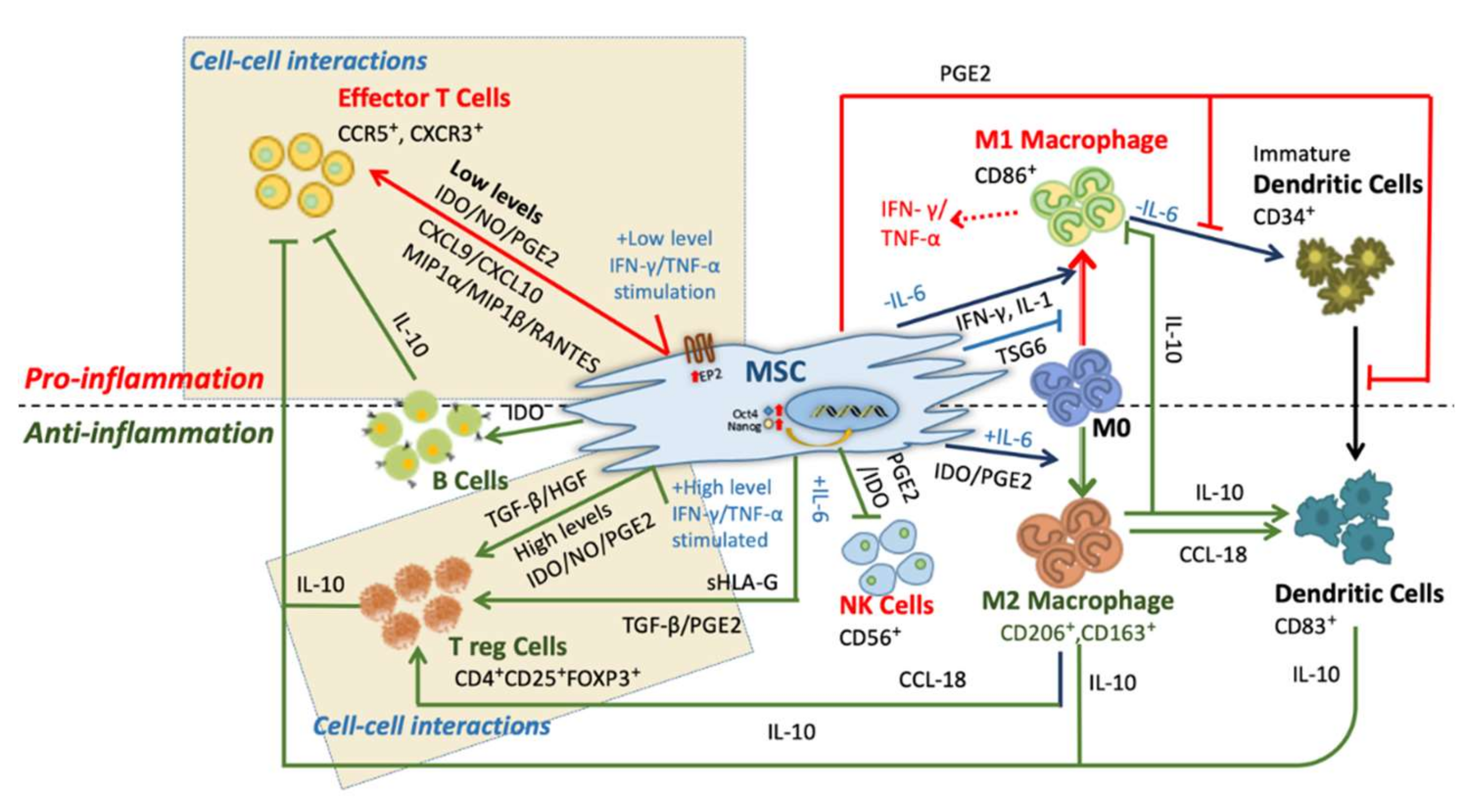{kind=link}
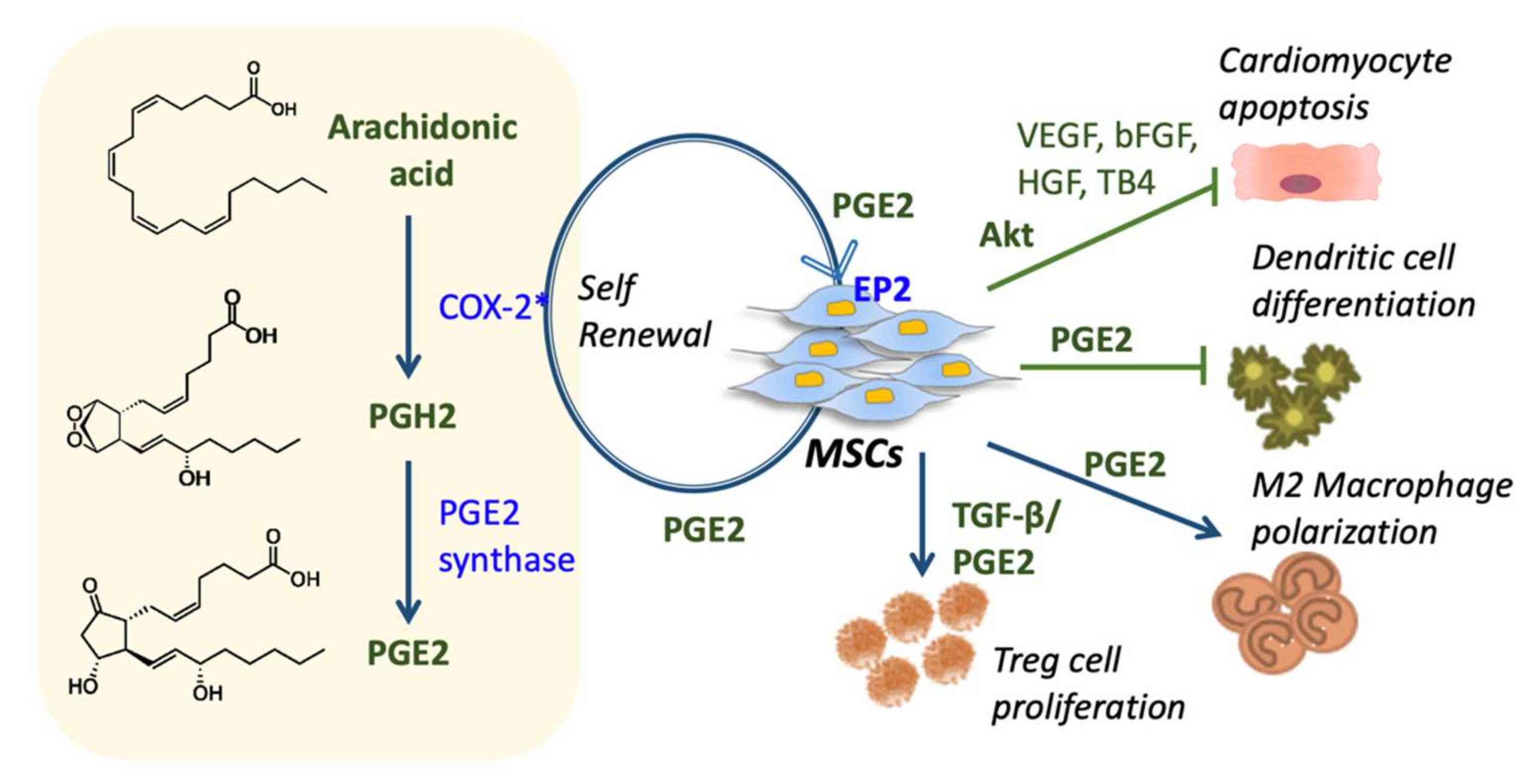{kind=link}
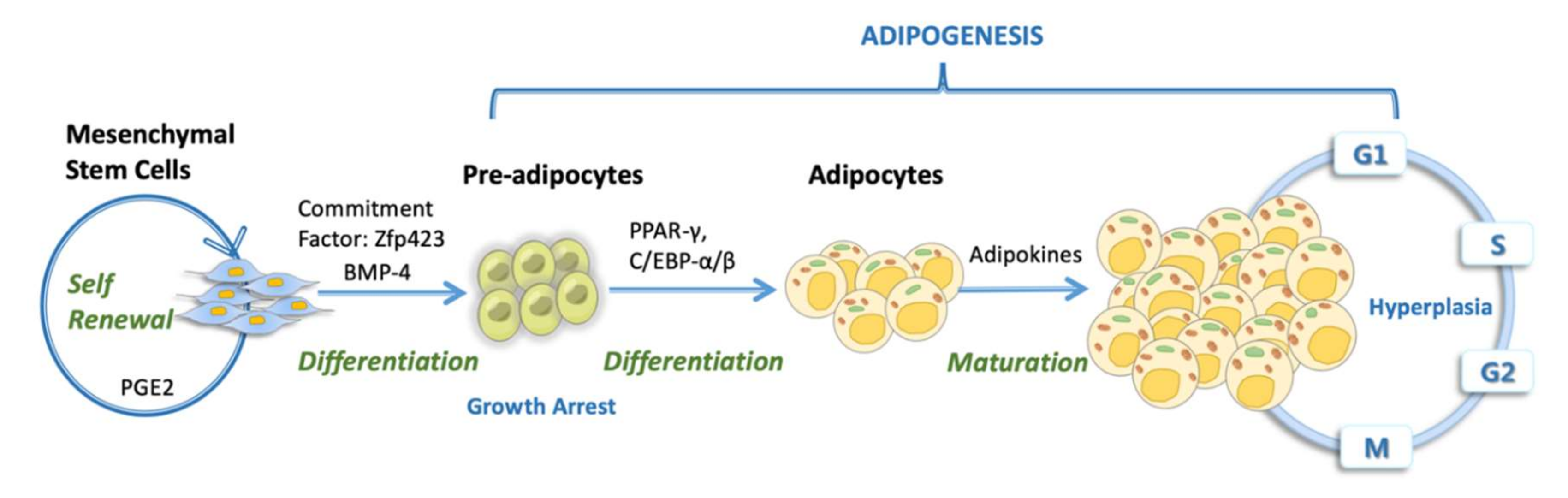{kind=link}
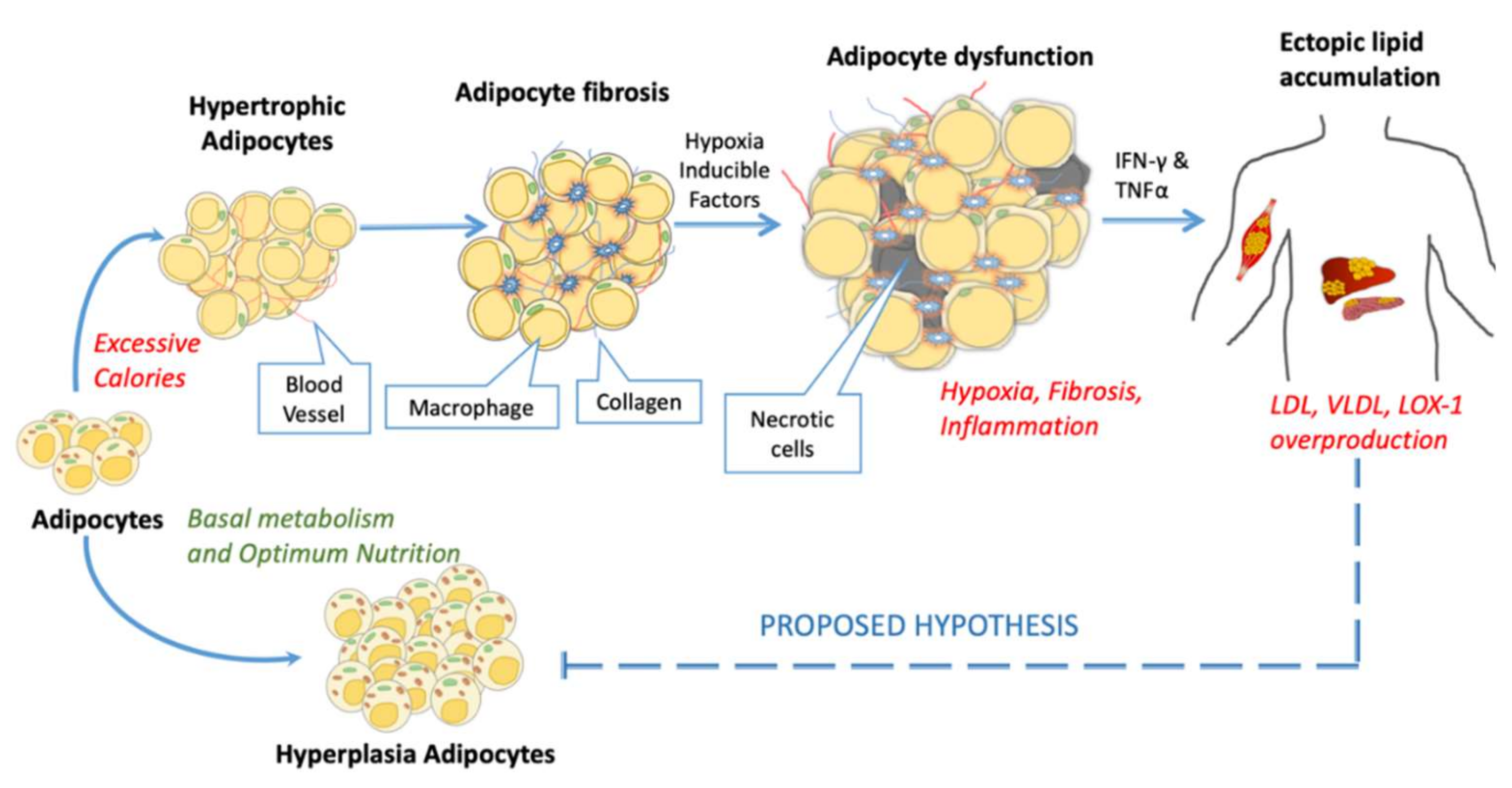{kind=link}
Table 1.
Therapeutic potential of MSCs tested.
| Site of Isolation | Subject tested | Role of MSCs | Modifications | Applications | Therapeutic Potential | Ref. |
|---|---|---|---|---|---|---|
| BM-MSC | Rat | Injury healing | _ | Autologous mode | Traumatic brain injury | [127] |
| BM-MSC | Mouse | Injury healing | _ | Allogenic mode | Traumatic brain injury | [128,129] |
| AD-MSC | Human | Knee injury healing | _ | Autologous | As a treatment for osteoarthritis | [130] |
| BM, AD, UC and WJ-MSC | Human | Immunosuppression | IFN-γ treated MSCs | Allogenic and autologous mode treatment and Prevention | GvHD | [131] |
| BM-MSC | Human | Immunosuppression | _ | Autologous | Multiple sclerosis | [132,133] |
| AD-MSC BM-MSC | Mouse | Antimicrobial and anti-infection | Collagenase 0.1% treatment | Allogenic and in vitro mode | Cystic fibrosis | [134] |
| UC-MSC | Mouse human | Immunosuppression | Vehicle-controlled without modification | Autologous and allogenic mode | Cirrhosis and autoimmune diseases | [135] |
| UC, AD, BM-MSC, Placenta | Human | Anti-inflammation, immunosuppression | _ | Exogenous MSCs | Bronchopulmonary dysplasia | [136] |
| BM-MSC | Human | Immunomodulation | _ | Autologous and Allogenic | Acute myocardial infarction, chronic ischemic heart disease, cardiomyopathy | [137] |
| BM-MSC | Mouse | Anti-inflammation, immunosuppression | Marrow-derived clonal MSC | Autologous | Inflammatory bowel disease | [138] |
| UC-MSC | Human | Immunosuppression | _ | Allogenic | Systemic lupus erythematosus | [133] |
| BM-MSC | Mouse | Anti-inflammation | _ | Allogenic | Chronic wound healing | [139] |
| BM-MSC | Human | Immunosuppression, immunomodulation | _ | Autologous | Drug-resistant epilepsy | [140] |
| BM-MSC | _ | MSCs cloning and proliferation | Feta bovine serum | Only Cultured | Chronic heart failure | [141] |
| AD-MSC | Mouse | Immunosuppression, immunomodulation | IL-35 gene modified | Exogenous | Con A-induced liver injury | [142] |
| BM-MSC (Purchased) | In vitro | Immunosuppression | Intracellular delivery of steroids | Allogenic Co-cultured | GvHD, Crohn’s disease | [124] |
| BM-MSC | YAC128 Mouse | Anti-inflammation, immunosuppression | Over-expressing BDNF and NGF genes | Autologous to mice models | Huntington’s disease | [125,143] |
| BM-MSC | Rat | Anti-inflammation, immunomodulation | Long term clonal MSCs | Reduces Fibrotic scars | Rat spinal cord injury | [144] |
BM-MSC: bone marrow-derived mesenchymal stem cell; AD-MSC: adipose tissue-derived MSC; UC-MSC: umbilical cord-derived mesenchymal stem cell; WJ-MSC: Wharton’s Jelly-derived MSC; IFN-γ: interferon-γ BDNF, brain-derived neurotrophic factors; NGF, Nerve growth factor; GvHD: graft versus host disease; Con A: concanavalin A.
Table 2.
Details of the clinical trials using MSCs on diabetes mellitus.
| Status | Outcomes/Complications | Criteria | NP | Treatment approach | Mode of intervention | Center/NCT No. | ||
|---|---|---|---|---|---|---|---|---|
| Primary | Secondary | Inclusions | Exclusions | |||||
| Completed | Reduction (≥50%) of insulin dose | HbA1c increases | T2DM for 5 years, 3 months’ medication before therapy, HbA1c range: 7.5% to 9% | Type 1 diabetes, chronic or severe diseases | 30 | 2 treatment at 6 months of interval | BM-MSCs, autologous inoculation | VRISCGT Hanoi Vietnam, NCT03343782 |
| Unknown; crossed the completion date | Reduction of insulin dose, change of C-peptide levels vs. baseline | Evaluation of adverse events e.g., fever, allergy | T2DM of age 18-80, ITT indicating insulin resistance, no infection | Chronic or severe diseases, HIV, Hepatitis B or C infection | 30 | 2 treatment at 3 months of interval | UC-MSCs, allograft and intravenous | Shandong University, China NCT01413035 |
| Completed | NF-κB inflammatory markers and osteoblast-specific gene expression | NF-κB inflammatory markers and apoptotic marker | T2DM of age 18 and above, HbA1c range between 6.5% | Receiving TZD, steroid or other medication, high serum creatinine 1.4 mg/dL for female and 1.5 for male | 75 | Cross-sectional 2-4 weeks of time frame | Not mentioned | Chiang Mai University, Thailand NCT02286128 |
| Unknown; crossed the completion date | Reduction of insulin dose, HbA1c increases | Adverse events | T2DM of age 18-75, ITT indicating insulin resistance | Severe diseases, HIV, Hepatitis B or C infection, pregnant | 24 | One-year time frame, 0 to 14 ±2 days 3 times | BM-MSCs intravenous | APGH, Beijing China, NCT01142050 |
| Completed | HbA1c monitored for 1 year, results not posted | Insulin dose, severity of adverse events | T2DM of age 35 or above, HbA1c range between 7.5% to 12% | Insulin requirement above 100 U/day, proteinuria, chronic or severe diseases receiving TZD, steroid or other medication | 22 | Cross-sectional for 1 year; Number of times not specified | Autologous BM-MSCs infused with BM-MNCs with insulin drug | FGH, Fuzhou China, NCT01719640 |
| Unknown; crossed the completion date | HbA1c monitored for 1 year | Fasting blood glucose monitored for 1 year | T2DM of age 35 to 65, HbA1c range between 7.5% to 11% | Insulin requirement above 100 U/day, proteinuria, chronic or severe diseases receiving TZD, steroid or other medication | 100 | Cross-sectional for 1 year; Number of times not specified | UC-MSCs infused with GLP-1 (Liraglutide) | Diabetes care center of Nanjing Military Command, Fuzhou China, NCT01954147 |
| Completed | Reduction (≥50%) of insulin dose | N.A. | T2DM of age 30 to 70, HbA1c below 7.5% | Type 1 diabetes, severe diseases, HIV, Hepatitis B or C infection | 30 | 6 months cross-sectional | Autologous BM-MSCs infused with vitamin B and MNCs | PIMER Chandigarh, India, NCT01759823 |
| Unknown; crossed the completion date | Osteoporosis in T2DM patients | N.A. | T2DM patients of age 40 to 99 | Organization people | 1200 | 3 years cross-sectional | Not provided | NTUH, Taipei, Taiwan, NCT01846533 |
| Active but not recruiting | CTCAE-assessment of 12 months and change in hypoglycemia | Fasting glucose monitored for 1 year, change of C-peptide and HbA1c | Type 1 diabetes detection less than 6 weeks, antibodies against pancreatic β-cells | Pregnant or breastfeeding, cancer or severe diseases, known HIV, Hepatitis B or C infection | 20 | 12 month cross-sectional, time frame of weeks 0 and 3 | Intravenous injection of autologous BM-MSCs | RIT, Tehran Iran, NCT04078308 |
| Completed | Change of C-peptide | N.A. | Type 1 diabetes detection within 10 days, fasting C-peptide below 0.12nmol/L | BMI, immuno-suppressive treatment, HIV, Hepatitis B or C infection, pregnant | 20 | 1-year follow-up study | Intravenous injection of autologous BM-MSCs | UUH, Uppsala, Sweden NCT01068951 |
| Terminated | Change of C-peptide | N.A. | Type 1 diabetes detection within 3 weeks, fasting C-peptide below 0.12nmol/L | BMI, immuno-suppressive treatment, HIV, Hepatitis B or C infection, pregnant | 50 | 2-year follow-up study | Intravenous injection of autologous BM-MSCs | UUH, Uppsala, Sweden NCT02057211 |
| Unknown; crossed the completion date | Change of C-peptide OGTT curve | Fasting blood glucose monitored for 1 year, a decrease in HbA1c | Type 1 diabetes of age 35 or below, HbA1c 7.5% or above | Insulin requirement above 100 U/day, proteinuria, chronic or severe diseases, receiving TZD, steroid or other medication | 44 | 1-year follow-up study | Intravenous injection of allograft UC –MSCs infused with pancreatic MNCs | FGH, Fuzhou China, NCT01374854 |
| Ongoing | Change of C-peptide | Change of C-peptide and change in β-cell function | Type 1 diabetes detection within 3 months, male and female of age 12 and 30 years | Body Mass Index < 14 or >35, HbA1c >12%, and/or fasting blood glucose >270 mg/d | 50 | 1-year follow-up study | Allograft BM–MSCs infused with plasmalyte 0.5% | MUSC, South Carolina USA NCT04061746 |
| Ongoing | Safe assessment of Allogenic use | N.A. | Type 1 diabetes of age 18 to 35, HbA1c change, baseline C-peptide | Type 1 diabetes below 18 and above 35 age, pregnant or comatose | 20 | 1-year follow-up study, 2 dosages at the interval of 6 months | Allogenic AD-MSCs infused with BM-MNCs intravenously | Cell Therapy Center, Amman, Jordan, NCT02940418 |
| Enrolling by invitation | Pancreatic β-cells monitoring by follow-up for 2 years and change of C-peptide analysis | Oral cholecalciferol 2000UI/day supplementation for 2 years | Type 1 diabetes detection within 4 months and pancreatic autoimmunity | HIV, Hepatitis B or C infection, pregnant, cancer | 30 | 1-year follow-up study | Allogenic AD-MSCs along with oral cholecalciferol supplementation and vitamin D | CFFUH Rio De Janeiro, Brazil, NCT03920397 |
NP: Number of participants; N.A.: Not Available; NF-κB: Nuclear factor-κB; VRISCGT: Vinmec Research Institute of Stem Cell and Gene Technology; ITT: Intravenous insulin tolerance test; APGH: Armed police general hospital; FGH: Fuzhou general hospital; PIMER: Postgraduate Institute of Medical Education and Research; NTUH: National Taiwan University Hospital; BM-MNCs: Bone marrow mononuclear cells; CTCAE: Common terminology criteria for adverse events; RIT: Royal Institute of Tehran; UUH: Uppsala University Hospital; MUSC: Medical University of South Carolina; CFFUH: Clementino Fraga Filho University Hospital; NCT number refer to its www.clinicaltrials.gov identifier.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mishra, V.K.; Shih, H.-H.; Parveen, F.; Lenzen, D.; Ito, E.; Chan, T.-F.; Ke, L.-Y. Identifying the Therapeutic Significance of Mesenchymal Stem Cells. Cells 2020, 9, 1145. https://doi.org/10.3390/cells9051145
AMA Style
Mishra VK, Shih H-H, Parveen F, Lenzen D, Ito E, Chan T-F, Ke L-Y. Identifying the Therapeutic Significance of Mesenchymal Stem Cells. Cells. 2020; 9(5):1145. https://doi.org/10.3390/cells9051145
Chicago/Turabian StyleMishra, Vineet Kumar, Hui-Hsuan Shih, Farzana Parveen, David Lenzen, Etsuro Ito, Te-Fu Chan, and Liang-Yin Ke. 2020. "Identifying the Therapeutic Significance of Mesenchymal Stem Cells" Cells 9, no. 5: 1145. https://doi.org/10.3390/cells9051145
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.