
Advances in Biomarkers and Endogenous Regulation of Breast Cancer Stem Cells

1
Key Laboratory of Animal Models and Human Disease Mechanisms, Kunming Institute of Zoology, Chinese Academy of Sciences and Yunnan Province, Kunming 650201, China
2
Kunming College of Life Sciences, The University of the Chinese Academy of Sciences, Kunming 650201, China
3
Fudan University Shanghai Cancer Center & Institutes of Biomedical Sciences, State Key Laboratory of Genetic Engineering, Cancer Institutes, Key Laboratory of Breast Cancer in Shanghai, The Shanghai Key Laboratory of Medical Epigenetics, Shanghai Key Laboratory of Radiation Oncology, The International Co-laboratory of Medical Epigenetics and Metabolism, Ministry of Science and Technology; Shanghai Medical College, Fudan University, Shanghai 200032, China
4
Jiangsu Key Lab of Cancer Biomarkers, Prevention and Treatment, Collaborative Innovation Center for Cancer Medicine, Nanjing Medical University, Nanjing 211166, China
5
Academy of Biomedical Engineering, Kunming Medical University, Kunming 650500, China
6
The Third Affiliated Hospital, Kunming Medical University, Kunming 650118, China
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Cells 2022, 11(19), 2941; https://doi.org/10.3390/cells11192941
Submission received: 24 August 2022
/
Revised: 13 September 2022
/
Accepted: 14 September 2022
/
Published: 20 September 2022
(This article belongs to the Special Issue Mammary Gland Stem Cells and the Cells of Origin of Breast Cancers)
Abstract
:Breast cancer is one of the most common cancers. Even if breast cancer patients initially respond to treatment, developed resistance can lead to a poor prognosis. Cancer stem cells (CSCs) are a group of undifferentiated cells with self-renewal and multipotent differentiation characteristics. Existing evidence has shown that CSCs are one of the determinants that contribute to the heterogeneity of primary tumors. The emergence of CSCs causes tumor recurrence, metastasis, and therapeutic resistance. Previous studies indicated that different stemness-associated surface markers can identify other breast cancer stem cell (BCSC) subpopulations. Deciphering the critical signaling networks that are involved in the induction and maintenance of stemness is essential to develop novel BCSC-targeting strategies. In this review, we reviewed the biomarkers of BCSCs, critical regulators of BCSCs, and the signaling networks that regulate the stemness of BCSCs.
1. Introduction
Cancer has become one of the world’s public health problems. According to the latest statistics, there are 19.3 million new cancer cases worldwide, and 2.26 million new cases of breast cancer among them, therefore breast cancer is ranked first [1]. In China, according to cancer statistics in 2018, it was found that the incidence of breast cancer is increasing, and the onset age is becoming younger [2]. Research suggests that breast cancer is closely related to genetic and environmental factors [3,4]. Breast cancer is a highly heterogeneous disease, divided into four subtypes, including Luminal A, Luminal B, HER2 positive, and triple-negative breast cancer (TNBC) [5].
Despite the rapid development of medical technology, breast cancer is still a recurrence risk. BCSCs are often associated with recurrence, metastasis, therapeutic resistance, and other biological behaviors [6]. Emerging evidence indicates that, although BCSCs account for only a small subset of tumor cells, they have tumorigenic potential, multipotent differentiation, and self-renewal capabilities [7]. BCSCs are derived from various sources, including normal stem-cell-gained mutations and de-differentiation of cancer cells induced by microenvironment components [8].
Targeting BCSCs seems to be an effective way to improve breast cancer treatment. The more we know about BCSCs, the more we understand how to eliminate them. In this review, we summarized the latest research progress on BCSCs, focusing on the identification biomarker of BCSCs and signal pathways regulating their stemness.
2. Identification of BCSCs
Since the concept of CSCs was proposed, scientists have tried identifying and isolating CSCs in various cancers. In breast cancer, BCSCs were initially identified as the CD24−/CD44+ phenotype. As few as 100 CD24−/CD44+ BCSCs isolated from breast cancer tissues of patients were able to form tumors in immunocompromised mice [9]. In vitro studies further verified that the CD24−/CD44+ cancer cell population exhibited a self-renewal capacity and could differentiate into bulk cancer cells [10]. Although cell surface markers confer specificity to identify and isolate BCSCs, scientists have also tried to utilize stem-associated characteristics to define stem cells. Aldehyde dehydrogenases (ALDHs) are enzymes responsible for aldehydes’ oxidation (dehydrogenation). Ginestier et al. discovered that the phenotype and function of cells with ALDH hyperactivity were like those of breast cancer stem cells. They used the ALDEFLUOR assay to identify and isolate cells with high ALDH enzyme activity (ALDH+ cells). The efficiency of the mammosphere formation of ALDH+ cells was approximately 13%, while no mammospheres formed from ALDH− cells [11]. ALDH+ and CD24−/CD44+ cell populations partially overlap and are interchangeable. Subsequently, Mollie et al. found that some of the cell populations isolated from a BRCA1-deficient mouse mammary tumor were composed of a low percentage of CD24−/CD44+ cells and harbored 2–5.9% CD133+ cells. These CD133+ breast cancer cells exhibited strong sphere formation, tumor initiation, and drug resistance capabilities, suggesting that CD133 was another BCSC marker. Notably, the observation that CD133+ BCSCs did not overlap with CD24−/CD44+ BCSCs raised the hypothesis that there might be heterogeneity in BCSCs [12].
Several new membrane proteins and prognostic markers have been reported to regulate stemness in specific subtypes of breast cancer, such as protein C receptor (PROCR) [13], tetraspanin 8 (TSPAN8) [14], tumor endothelial marker 8 (TEM8) [15], epsilon sarcoglycan (SGCE) [16], and Ki-67 [17]. PROCR, as a surface marker on mammary stem cells, was first identified as being expressed on multipotent mammary stem cells located in the basal layer of the mammary gland [18]. PROCR-positive cells exhibited a high self-renewal capacity in transplantation assays and was differentiated into multiple lineages, as indicated by lineage tracing. In breast cancer, interfering with PROCR expression significantly decreased the number of CSCs, and dampened tumor growth and recurrence in TNBC [13,19]. More research, such as lineage tracing, is needed to elucidate the characteristics of these newly identified BCSC subpopulations. Ki-67, an established prognostic indicator for assessing malignancy, was reported as being required to maintain the cancer stem cell niche in breast cancer [17]. Equivalent or higher levels of Ki-67 were observed in the metastatic lesion, indicating Ki-67 might be strongly associated with metastatic potential. However, the knockout of Ki-67 in the breast epithelia cell line MCF10A did not decrease cell proliferation [17]. The development and fertility of the Ki-67-knockout mice were normal [20].
Another way to identify CSCs is based on their features of biological behaviors, such as employing Hoechst 33,342 staining, or using a PKH26-based label retention assay to discriminate stem cells. The Hoechst 33,342 dye is a cell-permeant fluorescent stain used to label double-stranded DNA. Although all types of cells take up Hoechst, stem cells efflux faster than other cells. Based on this, cells in the side area of the Hoechst-positive cells were gated as stem cells using fluorescence-activated cell sorting (FACS) analysis and named side population (SP). SP cells not only express high levels of stemness genes but are also resistant to chemotherapy and radiotherapy. SP cells have been isolated in several cancers, especially those without surface markers [21]. PKH26, as a cell membrane dye, is mainly used to study the approach of cell division. Because PKH26 is equally inherited by daughter cells, the intensity of PKH26 reflects the number of cell divisions. Usually, stem cells or CSCs divide much more slowly and retain more of the PKH26 dye in daughter cells than normal cells or cancer cells. Thus, it is convenient to isolate or track these stem cells in a quiescent state in vivo using the PKH26 staining assay.
3. Endogenous Factors That Regulate BCSCs
3.1. Epigenetic Factors
BCSCs have a unique gene expression signature, predominantly determined by an array of epigenetic and transcription factors. B lymphoma Mo-MLV insertion region 1 homolog (BMI1) is a polycomb-group (PcG) transcriptional repressor that is well documented in promoting BCSC self-renewal and tumorigenicity [22]. The molecular chaperone heat shock protein 90 alpha (HSP90α) can maintain the expression of BMI1, subsequently increasing the self-renewal ability of BCSCs [23]. In contrast, miR-494-3p inhibits BCSC self-renewal by targeting BMI1 [24]. Recently, the activated interleukin-1 receptor type 2 (IL1R2) was shown to recruit USP15 in order to stabilize BMI1 and promote the self-renewal and metastasis of BCSCs [25].
Zeste homolog 2 (EZH2) is the catalytic subunit of polycomb repressor complex 2 (PRC2), which methylates histone H3 lysine 27 (H3K27) to inhibit transcription, and enhances the mammosphere-forming ability of BCSCs [26]. EZH2 promotes the self-renewal of BCSCs and increases the percentage of BCSCs by activating the Wnt pathway [27,28]. Li et al. discovered that the protein arginine methyltransferase 1 (PRMT1) catalyzes the asymmetric methylation of EZH2 to foster breast cancer proliferation, metastasis, and tumorigenesis [29,30].
Lysine-specific demethylase 1 (LSD1) selectively acts on histone H3K4 and H3K9 through a yellow flavin adenine dinucleotide (FAD)-dependent oxidative reaction. LSD1 has the effect of dual-transcriptional activators and inhibitors [31,32]. LSD1 may indirectly modulate CSCs via cancer-associated fibroblasts (CAFs) and the tumor microenvironment [32].
PRMT1 is an asymmetric arginine N-methyltransferase in mammalian cells that catalyzes the asymmetric demethylation of histone H4 arginine 3 (H4R3) and modifies the active chromatin [33]. In addition, PRMT1 modulates cell function through the methylation of forkhead box O1 (FOXO1) and ERα proteins [34]. PRMT1 can induce MCF10A cells to develop stem cell characteristics and self-renewal abilities [33]. PRMT1 may increase the attributes of TNBC stem cells by activating the signal transducer and activator of transcription 3 (STAT3). Moreover, the epidermal growth factor receptor (EGFR) signal mediated by PRMT1 may contribute to the upregulation of ZEB1 and the promotion of BCSC generation [34]. The same family member PRMT5 is a type II methyltransferase that controls the symmetrical demethylation of arginine residues on target proteins in the cytoplasm and nucleus. It modulates transcription through the methylation of transcription factors, such as nuclear factor kappa B (NF-κB), p53, and E2F Transcription Factor 1 (E2F1) [35]. PRMT5 promotes the expansion of stem cells through histone methylation and the expression of forkhead box P1 (FOXP1) and Kruppel-like Factor 4 (KLF4), thereby enabling the development of breast tumors and chemotherapy resistance [36]. Similarly, PRMT5 methylates KLF5 to prevent its phosphorylation, ubiquitination, degradation, and facilitates the transcription of downstream target genes, thereby promoting the maintenance and proliferation of BCSCs [37].
Epigenetic-factor histone deacetylases (histone deacetylases, HDACs) target lysine residues to facilitate chromatin condensation and to regulate biological processes such as mitosis, differentiation, autophagy, and apoptosis [38,39]. HDAC1, HDAC5, and HDAC7 are necessary to maintain BCSCs. HDAC7 overexpression is sufficient to increase the CSC phenotype, manifested in the increases in sphere formation and tumor-initiating cell frequency [39]. HDAC5 silencing inhibits the growth, migration, and invasion of BCSCs and increases apoptosis [38]. HDAC1 stabilizes KLF5 protein by preventing its ubiquitination and degradation [40]. The expression of HDAC1 and KLF5 is positively associated with breast cancer [40]. HDAC inhibitors inhibit the expression of KLF5 and the tumorigenesis of breast cancer in vivo [38,40]. At the same time, another study showed that HDAC inhibitors partially expanded the number of BCSCs through the β-catenin signaling pathway [38,41].
3.2. Non-coding RNAs
Many microRNAs (miRNAs) were reported to regulate BCSCs (Table 1). The miR-200c family was reported to inhibit BCSCs by targeting peptidylprolyl cis/trans isomerase, NIMA-interacting 1 (Pin1) [42]. It also suppresses the expression of BMI1 and the recombinant suppressor of zeste 12 homolog (Suz12) [42]. Additionally, the Let-7 family reduces the self-renewal capacity of BCSCs by inhibiting the Wnt signaling pathway [43]. In contrast, miRNA-221/222 promotes BCSC self-renewal by downregulating PTEN expression [44,45], and miR-20b-5p promotes the proliferation and inhibits the apoptosis of BCSCs [46].
Similarly, long noncoding RNAs (lncRNAs) also regulate BCSCs (Table 2). HOTAIR promotes the characteristics of BCSCs through regulating miR-34a/SRY-box transcription factor 2 (SOX2) [61]. SOX21-AS1 maintains BCSC stemness by enhancing the nuclear localization ability of the Yes-associated protein (YAP) [62]. Conversely, FGF13-AS1 inhibits the stemness of BCSCs, which destroys the interaction between insulin-like growth factor 2 mRNA-binding proteins (IGF2BPs) and MYC [63].
3.3. Transcription Factors and Signal Transduction Pathways
3.3.1. Transcription Factors and Co-Activators
In addition to epigenetic factors, BCSCs are also regulated by transcription factors (Table 3). Octamer-binding transcription factor-4 (OCT4) plays a vital role in stem cell self-renewal. The high expression of OCT4 in 4T1 breast cancer cells enhances the mammosphere formation of CSCs in vitro [77]. The knockdown of OCT4 in an MCF-7 tumor-model induced apoptosis and inhibited tumor growth [78]. Estrogen induced OCT4 expression in MCF-7 cells and promoted mammosphere formation [79]. Programmed death ligand (PD-L1) maintains CSC stemness by promoting OCT4 phosphorylation, but the histone demethylase jumonji domain-containing protein-3 (JMJD3) decreases breast cancer stem cell-like properties by downregulating OCT4 [80,81].
KLF4 and KLF5 play oncogenic roles in breast tumors. KLF4 overexpression increases the proportion of CSCs [82] whereas miR-7 hinders the self-renewal and invasion abilities of CSCs by targeting KLF4 [83]. Lysine demethylase 7A (KDM7A) maintains BCSCs by upregulating KLF4 and c-MYC [84]. The downregulation of dual-specificity tyrosine phosphorylation-regulated kinase 2 (DYRK2) increases the expression of KLF4 and the proportion of BCSCs [85]. Our previous study found that KLF5 was essential for maintaining the stemness of normal breast stem cells and BCSCs in basal-like breast cancer (BLBC) [58,86]. PRMT5 increases the stemness of CSCs in BLBC by stabilizing KLF5 protein [37]. Mifepristone and metformin suppress BCSCs by inhibiting KLF5 expression [58,87]. Interestingly, several KLF5 downstream target genes, including SLUG [86,88] and NANOG [89,90], also increase the stemness of BCSCs.
C-MYC and SOX2 are two stem cell transcription factors. The overexpression of c-MYC in MDA-MB-468 cells facilitates the expression of ALDH [91]. Caveolin-1 inhibits the c-MYC-mediated metabolic reprogramming function of BCSCs [92]. In addition, p62 enhances the stem-like properties of BCSCs by stabilizing c-MYC [93]. SOX2 knockdown reduces BCSC stemness [94]. The tumor suppressor transcriptional repressor GATA binding 1 (TRPS1) inhibits SOX2 expression and the tumorigenic ability of CSCs [95]. Similarly, FOXO3a inhibits SOX2 expression and BCSC tumorigenicity [96]. Additionally, knockdown of SOX9 significantly inhibits the tumorigenicity of MDA-MB-231 cells, with a 70-fold decrease in tumor initiation capacity and a 40-fold increase in the ability to inhibit lung metastasis compared with the control [97]. More importantly, SOX9 and SLUG have significant advantages in the synergistic reversal of differentiated luminal cells into mammary stem cells [97].
3.3.2. Signal Transduction Pathways
The signal transduction pathways control gene expression in response to environmental stimuli (Table 4 and Figure 1). In TNBC, NOTCH-1 maintains CSC stemness, and inhibition of NOTCH-1 significantly reduces the number of CSCs [113,114]. Additionally, NOTCH-1 maintains the survival of BCSCs by inhibiting PTEN and activating extracellular signal-regulated kinase 1/2 (ERK1/2) [115]. NOTCH-2 knockdown can reduce the percentage of BCSCs [116]. NOTCH-3 inhibits the epithelial-to-mesenchymal transition (EMT) of BCSCs via the Kibra-mediated Hippo pathway [116], and inhibits the self-renewal of BCSCs by the interleukin-6 (IL6)/STAT3 signaling pathway [117]. NOTCH-4 plays a specific role in differentiating BCSCs into progenitor cells and reduces the expression level of NOTCH-4, impacting on its ability to form mammospheres [114].
Leucine-rich repeat-containing G protein-coupled receptor 4 (LGR4) acts as the primary positive regulator of the Wnt/β-catenin signaling pathway to maintain BCSCs [118]. CDH11 targets β-catenin, thereby inhibiting the stemness of TNBC cells [119]. Silencing LDL receptor-related protein 8 (LRP8) can reduce the percentage of BCSCs in TNBC [120]. β1,4-Galactosyltransferase V (B4GalT5) stabilizes Frizzled-1 by glycosylation and maintains BCSC stemness [121].
The activation of the Hedgehog signaling pathway also increases the number of BCSCs and the formation of mammospheres. TSPAN8 is upregulated in BCSCs, interacts with the SHH-PTCH1 complex, and promotes CSC stemness [14]. TSPAN8 was recently found to enter the nucleus by binding to 14-3-3θ, importin-β, and cholesterol [14]. Ubiquitin-specific peptidase 37 (USP37) activates the Hedgehog pathway by increasing Smo and GLI1 expression levels to enhance the characteristics of BCSCs [122]. The GLI1-derived tumors amplify a portion of CSCs featured by Keratin 6 and BMI1, supporting the role of Hedgehog signaling in breast cancer development [123].
TAZ, an effector of the Hippo pathway, is necessary to maintain the self-renewal ability of BCSCs [124]. Vascular endothelial growth factor (VEGF)/neuropilin-2 (NRP2) signaling participates in TAZ activation through a Rac1-dependent mechanism, enhancing the sphere-forming ability of BCSCs [125]. Silencing of the discs large homolog 5 (DLG5) might improve the activity of TAZ, thereby maintaining the self-renewal ability and stemness of BSCSs [126]. We previously reported that TAZ can stabilize KLF5 [127] and that tumor necrosis factor-alpha (TNFɑ) induces TAZ expression to increase the stemness of BCSCs [128]. Similarly, YAP was reported to promote the stemness of BCSCs [129]. Activating the competing endogenous RNA (ceRNA) network associated with star-related lipid transfer domain-containing 13 (STARD13) reduces YAP/YAZ activity, thereby inhibiting the formation of BCSCs [130]. YAP interacts with β-catenin, and TEA domain transcription factor 4 (TEAD4) cooperates in the nucleus to modulate CSCs in BLBC [131].
Receptor tyrosine kinases (RTKs) can activate the downstream PI3K/AKT/mTOR and MAPK signaling pathways. Type 1 insulin-like growth factor receptor (IGF-1R) maintains BCSCs by activating the downstream PI3K/Akt/mTOR pathway [132]. The downregulation of hypoxia-inducible factor 2 alpha (HIF-2α) expression can inhibit BCSCs by inhibiting the PI3K/AKT/mTOR signaling pathway [133]. B7-H3 activates MEK by binding to major vault protein (MVP), which regulates the MAPK kinase pathway and increases the proportion of BCSCs [134]. EGFR upregulates cyclooxygenase 2 (COX-2) to activate Nodal signaling and promote BCSC self-renewal [135]. SGCE stabilizes the level of EGFR by breaking the interaction between c-Cbl and EGFR, activating the PI3K/Akt pathway to foster breast cell stemness [16].
Wang et al. reported that the JAK/STAT3 pathway promotes BCSC differentiation by regulating lipid metabolism and that inhibition of this pathway blocks BCSC self-renewal [136]. WW domain-containing oxidoreductase (WWOX) inhibits the proliferation and metastasis of breast cancer cells by inhibiting the phosphorylation of JAK2 to hinder STAT3 activation [137]. WWOX also inhibits the expression of KLF5 in breast cancer [138]. In contrast, in TNBC, EGFR activates the JAK/STAT3 pathway by promoting the phosphorylation of STAT3, thereby facilitating the proliferation and invasion of tumor cells [139].
Alec et al. found that transforming growth factor-beta (TGF-β) activated the cytokine receptor leukemia inhibitory factor-receptor (LIFR) to initiate the JAK-STAT signaling pathway, subsequently driving the formation of BCSCs [140]. Jun et al. showed that TGF-β enhances the self-renewal capacity of BCSCs by upregulating fibronectin and Smad3-dependent COX-2 expression [141].
The transcription factor NF-κB is overexpressed in BCSCs, and high NF-κB activity regulates the self-renewal and differentiation of BCSCs [142]. Interleukin-1 alpha (IL-1α) mediates the HER2-induced NF-κB pathway to maintain BCSCs [143]. TNF-α promotes the self-renewal of BCSCs in human breast cancer cell lines by upregulating TAZ expression through the atypical NF-κB pathway [128]. HGFL-RON signaling increases the self-renewal capacity of BCSCs by activating β-catenin and its effector NF-κB [144]. Heregulin (HRG), the ligand of Erb-B2 receptor tyrosine kinase 3 (ErbB3), has been proved to stimulate mammosphere formation, which is achieved by activating the PI3K/NF-κB pathway [145]. Heat shock protein 27 (Hsp27) degrades IκBα to activate NF-κB and maintain BCSCs [146].
The scaffold protein SH3 domain containing ring finger 3 (SH3RF3), which is upregulated in BCSCs, interacts with c-Jun N-terminal kinases (JNK) in a JNK-interacting protein (JIP)-dependent manner and phosphorylates the latter, thereby activating the JNK-JUN pathway. This promotes the characteristics of BCSCs by enhancing the expression of pentraxin 3 (PTX3) [147].
4. Conclusions
Although the 5-year survival rate of breast cancer has been dramatically improved, recurrence is still a big challenge for breast cancer treatment. BCSCs are closely associated with recurrence. Notwithstanding that biomarkers have demonstrated their value in identifying BCSCs, it is still difficult to distinguish between normal stem cells and BCSCs. Due to the heterogeneity of breast cancer, the expressions of BCSC markers are varied and show distinct clinical values in different breast cancer subtypes. Moreover, distinct BCSC subclones may co-exist with a heterogeneous tumor, and new BCSC subclones may be induced during tumor treatment. Therefore, it is necessary to combine the analysis of BCSC markers in certain phases during tumor progression to improve BCSC-based prognosis. The continuous updating of breast cancer-specific biomarkers has an important guiding significance for CSC identification and separation. Novel therapeutic strategies that target BCSC and BCSC niches and the rest of the cancer are required to target the entire cancer and prevent metastasis or recurrence. Currently, there are several treatments for BCSCs, including cytotherapy (NCT02915445), antibody-based biopharmaceuticals (NCT01954355), synthetic small molecule compounds (NCT00645333), natural compounds and their products (NCT01608867), and nucleic acid medicines. In the I-SPY2 (Investigation of serial studies to predict your therapeutic response with imaging and molecular analysis) clinical trial platform, 17 new agents were evaluated in combination with neoadjuvant chemotherapy for women with locally advanced breast cancer [148]. By conducting such clinical trials, promising drugs were approved more quickly, and the drug development process was shortened [149].
Cancer occurrence is a multi-factor, multi-stage, multi-gene mutation accumulation process. Various signaling pathways may be involved in cancer occurrence and development simultaneously. Most of the signaling pathways regulating BCSCs are evolutionarily conserved and shared with normal stem cells, which makes them inappropriate as therapeutic targets. With a deep understanding of BCSCs, we expect more specific regulation of BCSCs pathway factors to be found. The relationship between multiple pathways is closely supported, providing a more theoretical basis for developing new targeted therapies in order to overcome current breast cancer treatment limitations.
Author Contributions
W.C. and L.Z. wrote and edited the paper and generated the tables and figure. S.L. and C.C. revised the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Key R&D Program of China (2020YFA0112300 and 2020YFA0803200); the National Natural Science Foundation of China (U2102203 and 81830087); and the Yunnan Fundamental Research Projects (202101AS070050); The “Ten Thousand Plan”-National High-Level Talents Special Support Plan (WR-YK5202101); the Program for Outstanding Leading Talents in Shanghai; the Program of Shanghai Academic/Technology Research Leader (20XD1400700); the Program for Outstanding Medical Academic Leader in Shanghai (2019LJ04); and the innovative research team of high-level local university in Shanghai.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Feng, R.M.; Zong, Y.N.; Cao, S.M.; Xu, R.H. Current cancer situation in China: Good or bad news from the 2018 Global Cancer Statistics? Cancer Commun. 2019, 39, 22. [Google Scholar] [CrossRef]
- Loibl, S.; Poortmans, P.; Morrow, M.; Denkert, C.; Curigliano, G. Breast cancer. Lancet 2021, 397, 1750–1769. [Google Scholar] [CrossRef]
- Łukasiewicz, S.; Czeczelewski, M.; Forma, A.; Baj, J.; Sitarz, R.; Stanisławek, A. Breast Cancer-Epidemiology, Risk Factors, Classification, Prognostic Markers, and Current Treatment Strategies-An Updated Review. Cancers 2021, 13, 4287. [Google Scholar] [CrossRef]
- Yeo, S.K.; Guan, J.L. Breast Cancer: Multiple Subtypes within a Tumor? Trends Cancer 2017, 3, 753–760. [Google Scholar] [CrossRef]
- Gu, H.F.; Mao, X.Y.; Du, M. Prevention of breast cancer by dietary polyphenols—role of cancer stem cells. Crit. Rev. Food Sci. Nutr. 2019, 60, 810–825. [Google Scholar] [CrossRef]
- Wang, C.; Xu, K.; Wang, R.; Han, X.; Tang, J.; Guan, X. Heterogeneity of BCSCs contributes to the metastatic organotropism of breast cancer. J. Exp. Clin. Cancer Res. 2021, 40, 1–14. [Google Scholar] [CrossRef]
- Bai, X.; Ni, J.; Beretov, J.; Graham, P.; Li, Y. Cancer stem cell in breast cancer therapeutic resistance. Cancer Treat. Rev. 2018, 69, 152–163. [Google Scholar] [CrossRef]
- Al-Hajj, M.; Wicha, M.S.; Benito-Hernandez, A.; Morrison, S.J.; Clarke, M.F. Prospective identification of tumorigenic breast cancer cells. Proc. Natl. Acad. Sci. 2003, 100, 3983–3988. [Google Scholar] [CrossRef]
- Ponti, D.; Costa, A.; Zaffaroni, N.; Pratesi, G.; Petrangolini, G.; Coradini, D.; Pilotti, S.; Pierotti, M.A.; Daidone, M.G. Isolation and In vitro Propagation of Tumorigenic Breast Cancer Cells with Stem/Progenitor Cell Properties. Cancer Res. 2005, 65, 5506–5511. [Google Scholar] [CrossRef] [Green Version]
- Ginestier, C.; Hur, M.H.; Charafe-Jauffret, E.; Monville, F.; Dutcher, J.; Brown, M.; Jacquemier, J.; Viens, P.; Kleer, C.G.; Liu, S.; et al. ALDH1 Is a Marker of Normal and Malignant Human Mammary Stem Cells and a Predictor of Poor Clinical Outcome. Cell Stem Cell 2007, 1, 555–567. [Google Scholar] [CrossRef]
- Wright, M.H.; Calcagno, A.M.; Salcido, C.D.; Carlson, M.D.; Ambudkar, S.V.; Varticovski, L. Brca1 breast tumors contain distinct CD44+/CD24- and CD133+ cells with cancer stem cell characteristics. Breast Cancer Res. 2008, 10, R10. [Google Scholar] [CrossRef]
- Wang, D.; Cai, C.; Dong, X.; Yu, Q.C.; Zhang, X.-O.; Yang, L.; Zeng, Y.A. Identification of multipotent mammary stem cells by protein C receptor expression. Nature 2014, 517, 81–84. [Google Scholar] [CrossRef]
- Zhu, R.; Gires, O.; Zhu, L.; Liu, J.; Li, J.; Yang, H.; Ju, G.; Huang, J.; Ge, W.; Chen, Y.; et al. TSPAN8 promotes cancer cell stemness via activation of sonic Hedgehog signaling. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef]
- Xu, J.; Yang, X.; Deng, Q.; Yang, C.; Wang, D.; Jiang, G.; Yao, X.; He, X.; Ding, J.; Qiang, J.; et al. TEM8 marks neovasculogenic tumor-initiating cells in triple-negative breast cancer. Nat. Commun. 2021, 12, 1–15. [Google Scholar] [CrossRef]
- Zhao, L.; Qiu, T.; Jiang, D.; Xu, H.; Zou, L.; Yang, Q.; Chen, C.; Jiao, B. SGCE Promotes Breast Cancer Stem Cells by Stabilizing EGFR. Adv. Sci. 2020, 7, 1903700. [Google Scholar] [CrossRef]
- Cidado, J.; Wong, H.Y.; Rosen, D.M.; Cimino-Mathews, A.; Garay, J.P.; Fessler, A.G.; Rasheed, Z.A.; Hicks, J.; Cochran, R.L.; Croessmann, S.; et al. Ki-67 is required for maintenance of cancer stem cells but not cell proliferation. Oncotarget 2016, 7, 6281–6293. [Google Scholar] [CrossRef]
- Wang, D.; Liu, C.; Wang, J.; Jia, Y.; Hu, X.; Jiang, H.; Shao, Z.-M.; Zeng, Y.A. Protein C receptor stimulates multiple signaling pathways in breast cancer cells. J. Biol. Chem. 2018, 293, 1413–1424. [Google Scholar] [CrossRef]
- Xue, G.; Wang, K.; Zhou, D.; Zhong, H.; Pan, Z. Light-Induced Protein Degradation with Photocaged PROTACs. J. Am. Chem. Soc. 2019, 141, 18370–18374. [Google Scholar] [CrossRef]
- Sobecki, M.; Mrouj, K.; Camasses, A.; Parisis, N.; Nicolas, E.; Llères, D.; Gerbe, F.; Prieto, S.; Krasinska, L.; David, A.; et al. The cell proliferation antigen Ki-67 organises heterochromatin. eLife 2016, 5, e13722. [Google Scholar] [CrossRef]
- Shimoda, M.; Ota, M.; Okada, Y. Isolation of Cancer Stem Cells by Side Population Method. Methods Mol. Biol. 2017, 1692, 49–59. [Google Scholar] [CrossRef]
- Srinivasan, M.; Bharali, D.J.; Sudha, T.; Khedr, M.; Guest, I.; Sell, S.; Glinsky, G.V.; Mousa, S.A. Downregulation of Bmi1 in breast cancer stem cells suppresses tumor growth and proliferation. Oncotarget 2017, 8, 38731–38742. [Google Scholar] [CrossRef]
- Lee, Y.-C.; Chang, W.-W.; Chen, Y.-Y.; Tsai, Y.-H.; Chou, Y.-H.; Tseng, H.-C.; Chen, H.-L.; Wu, C.-C.; Chang-Chien, J.; Lee, H.-T.; et al. Hsp90α Mediates BMI1 Expression in Breast Cancer Stem/Progenitor Cells through Facilitating Nuclear Translocation of c-Myc and EZH2. Int. J. Mol. Sci. 2017, 18, 1986. [Google Scholar] [CrossRef]
- Chen, S.-M.; Wang, B.-Y.; Lee, C.-H.; Lee, H.-T.; Li, J.-J.; Hong, G.-C.; Hung, Y.-C.; Chien, P.-J.; Chang, C.-Y.; Hsu, L.-S.; et al. Hinokitiol up-regulates miR-494-3p to suppress BMI1 expression and inhibits self-renewal of breast cancer stem/progenitor cells. Oncotarget 2017, 8, 76057–76068. [Google Scholar] [CrossRef]
- Zhang, L.; Qiang, J.; Yang, X.; Wang, D.; Rehman, A.U.; He, X.; Chen, W.; Sheng, D.; Zhou, L.; Jiang, Y.; et al. IL1R2 Blockade Suppresses Breast Tumorigenesis and Progression by Impairing USP15-Dependent BMI1 Stability. Adv. Sci. 2019, 7, 1901728. [Google Scholar] [CrossRef]
- van Vlerken, L.E.; Kiefer, C.M.; Morehouse, C.; Li, Y.; Groves, C.; Wilson, S.D.; Yao, Y.; Hollingsworth, R.E.; Hurt, E.M. EZH2 is required for breast and pancreatic cancer stem cell maintenance and can be used as a functional cancer stem cell reporter. Stem Cells Transl. Med. 2013, 2, 43–52. [Google Scholar] [CrossRef]
- Li, J.; Xi, Y.; Li, W.; McCarthy, R.L.; A Stratton, S.; Zou, W.; Dent, S.Y.; Jain, A.K.; Barton, M.C. TRIM28 interacts with EZH2 and SWI/SNF to activate genes that promote mammosphere formation. Oncogene 2017, 36, 2991–3001. [Google Scholar] [CrossRef]
- Zhao, Z.; Li, S.; Song, E.; Liu, S. The roles of ncRNAs and histone-modifiers in regulating breast cancer stem cells. Protein Cell 2015, 7, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, D.; Chen, X.; Wang, W.; Wang, P.; Hou, P.; Li, M.; Chu, S.; Qiao, S.; Zheng, J.; et al. PRMT1-mediated EZH2 methylation promotes breast cancer cell proliferation and tumorigenesis. Cell Death Dis. 2021, 12, 1–11. [Google Scholar] [CrossRef]
- Li, Z.; Wang, D.; Lu, J.; Huang, B.; Wang, Y.; Dong, M.; Fan, D.; Li, H.; Gao, Y.; Hou, P.; et al. Methylation of EZH2 by PRMT1 regulates its stability and promotes breast cancer metastasis. Cell Death Differ. 2020, 27, 3226–3242. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, Y.; Yang, X.H.; Kang, T.; Zhao, Y.; Wang, C.; Evers, B.M.; Zhou, B.P. The Deubiquitinase USP28 Stabilizes LSD1 and Confers Stem-Cell-like Traits to Breast Cancer Cells. Cell Rep. 2013, 5, 224–236. [Google Scholar] [CrossRef]
- Boulding, T.; McCuaig, R.D.; Tan, A.; Hardy, K.; Wu, F.; Dunn, J.; Kalimutho, M.; Sutton, C.R.; Forwood, J.; Bert, A.G.; et al. Author Correction: LSD1 activation promotes inducible EMT programs and modulates the tumour microenvironment in breast cancer. Sci. Rep. 2019, 9, 18771. [Google Scholar] [CrossRef]
- Gao, Y.; Zhao, Y.; Zhang, J.; Lu, Y.; Liu, X.; Geng, P.; Huang, B.; Zhang, Y.; Lu, J. The dual function of PRMT1 in modulating epithelial-mesenchymal transition and cellular senescence in breast cancer cells through regulation of ZEB1. Sci. Rep. 2016, 6, 19874. [Google Scholar] [CrossRef]
- Nakai, K.; Xia, W.; Liao, H.-W.; Saito, M.; Hung, M.-C.; Yamaguchi, H. The role of PRMT1 in EGFR methylation and signaling in MDA-MB-468 triple-negative breast cancer cells. Breast Cancer 2017, 25, 74–80. [Google Scholar] [CrossRef]
- Lattouf, H.; Kassem, L.; Jacquemetton, J.; Choucair, A.; Poulard, C.; Tredan, O.; Corbo, L.; Diab-Assaf, M.; Hussein, N.; Treilleux, I.; et al. LKB1 regulates PRMT5 activity in breast cancer. Int. J. Cancer 2019, 144, 595–606. [Google Scholar] [CrossRef]
- Zhou, Z.; Feng, Z.; Hu, D.; Yang, P.; Gur, M.; Bahar, I.; Cristofanilli, M.; Gradishar, W.J.; Xie, X.-Q.; Wan, Y. A novel small-molecule antagonizes PRMT5-mediated KLF4 methylation for targeted therapy. eBioMedicine 2019, 44, 98–111. [Google Scholar] [CrossRef]
- Wang, X.; Qiu, T.; Wu, Y.; Yang, C.; Li, Y.; Du, G.; He, Y.; Liu, W.; Liu, R.; Chen, C.-H.; et al. Arginine methyltransferase PRMT5 methylates and stabilizes KLF5 via decreasing its phosphorylation and ubiquitination to promote basal-like breast cancer. Cell Death Differ. 2021, 28, 2931–2945. [Google Scholar] [CrossRef]
- Hsieh, T.H.; Hsu, C.Y.; Tsai, C.F.; Long, C.Y.; Wu, C.H.; Wu, D.C.; Lee, J.N.; Chang, W.C.; Tsai, E.M. HDAC inhibitors target HDAC5, upregulate microRNA-125a-5p, and induce apoptosis in breast cancer cells. Mol. Ther. 2015, 23, 656–666. [Google Scholar] [CrossRef]
- Witt, A.E.; Lee, C.-W.; Lee, T.I.; Azzam, D.J.; Wang, B.; Caslini, C.; Petrocca, F.; Grosso, J.; Jones, M.; Cohick, E.B.; et al. Identification of a cancer stem cell-specific function for the histone deacetylases, HDAC1 and HDAC7, in breast and ovarian cancer. Oncogene 2017, 36, 1707–1720. [Google Scholar] [CrossRef]
- Kong, Y.; Ren, W.; Fang, H.; Shah, N.A.; Shi, Y.; You, D.; Zhou, C.; Jiang, D.; Yang, C.; Liang, H.; et al. Histone Deacetylase Inhibitors (HDACi) Promote KLF5 Ubiquitination and Degradation in Basal-like Breast Cancer. Int. J. Biol. Sci. 2022, 18, 2104–2115. [Google Scholar] [CrossRef]
- Debeb, B.G.; Lacerda, L.; Xu, W.; Larson, R.; Solley, T.; Atkinson, R.; Sulman, E.P.; Ueno, N.T.; Krishnamurthy, S.; Reuben, J.M.; et al. Histone deacetylase inhibitors stimulate dedifferentiation of human breast cancer cells through WNT/beta-catenin signaling. Stem Cells 2012, 30, 2366–2377. [Google Scholar] [CrossRef] [PubMed]
- Prabhu, K.; Raza, A.; Karedath, T.; Raza, S.; Fathima, H.; Ahmed, E.; Kuttikrishnan, S.; Therachiyil, L.; Kulinski, M.; Dermime, S.; et al. Non-Coding RNAs as Regulators and Markers for Targeting of Breast Cancer and Cancer Stem Cells. Cancers 2020, 12, 351. [Google Scholar] [CrossRef]
- Sun, X.; Xu, C.; Tang, S.-C.; Wang, J.; Wang, H.; Wang, P.; Du, N.; Qin, S.; Li, G.; Xu, S.; et al. Let-7c blocks estrogen-activated Wnt signaling in induction of self-renewal of breast cancer stem cells. Cancer Gene Ther. 2016, 23, 83–89. [Google Scholar] [CrossRef]
- Li, B.; Lu, Y.; Yu, L.; Han, X.; Wang, H.; Mao, J.; Shen, J.; Wang, B.; Tang, J.; Li, C.; et al. miR-221/222 promote cancer stem-like cell properties and tumor growth of breast cancer via targeting PTEN and sustained Akt/NF-kappaB/COX-2 activation. Chem. Biol. Interact 2017, 277, 33–42. [Google Scholar] [CrossRef]
- Li, B.; Lu, Y.; Wang, H.; Han, X.; Mao, J.; Li, J.; Yu, L.; Wang, B.; Fan, S.; Yu, X.; et al. miR-221/222 enhance the tumorigenicity of human breast cancer stem cells via modulation of PTEN/Akt pathway. Biomed. Pharmacother. 2016, 79, 93–101. [Google Scholar] [CrossRef]
- Xia, L.; Li, F.; Qiu, J.; Feng, Z.; Xu, Z.; Chen, Z.; Sun, J. Oncogenic miR-20b-5p contributes to malignant behaviors of breast cancer stem cells by bidirectionally regulating CCND1 and E2F1. BMC Cancer 2020, 20, 1–13. [Google Scholar] [CrossRef]
- Zhang, H.; Li, N.; Zhang, J.; Jin, F.; Shan, M.; Qin, J.; Wang, Y. The influence of miR-34a expression on stemness and cytotoxic susceptibility of breast cancer stem cells. Cancer Biol. Ther. 2016, 17, 614–624. [Google Scholar] [CrossRef]
- Liang, R.; Li, Y.; Wang, M.; Tang, S.-C.; Xiao, G.; Sun, X.; Li, G.; Du, N.; Liu, D.; Ren, H. MiR-146a promotes the asymmetric division and inhibits the self-renewal ability of breast cancer stem-like cells via indirect upregulation of Let-7. Cell Cycle 2018, 17, 1445–1456. [Google Scholar] [CrossRef]
- Han, M.-L.; Wang, F.; Gu, Y.-T.; Pei, X.-H.; Ge, X.; Guo, G.-C.; Li, L.; Duan, X.; Zhu, M.-Z.; Wang, Y.-M. MicroR-760 suppresses cancer stem cell subpopulation and breast cancer cell proliferation and metastasis: By down-regulating NANOG. Biomed. Pharmacother. 2016, 80, 304–310. [Google Scholar] [CrossRef]
- Zou, Y.; Chen, Y.; Yao, S.; Deng, G.; Liu, D.; Yuan, X.; Liu, S.; Rao, J.; Xiong, H.; Yuan, X.; et al. MiR-422a weakened breast cancer stem cells properties by targeting PLP2. Cancer Biol. Ther. 2018, 19, 436–444. [Google Scholar] [CrossRef] [Green Version]
- Troschel, F.M.; Böhly, N.; Borrmann, K.; Braun, T.; Schwickert, A.; Kiesel, L.; Eich, H.T.; Götte, M.; Greve, B. miR-142-3p attenuates breast cancer stem cell characteristics and decreases radioresistance in vitro. Tumor Biol. 2018, 40, 1010428318791887. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Wang, T.; He, D.; Li, X.; Jiang, Y. miR-1 inhibits the proliferation of breast cancer stem cells by targeting EVI-1. OncoTargets Ther. 2018, 11, 8773–8781. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wu, N.; Liu, L.; Dong, H.; Liu, X. microRNA-128-3p overexpression inhibits breast cancer stem cell characteristics through suppression of Wnt signalling pathway by down-regulating NEK2. J. Cell. Mol. Med. 2020, 24, 7353–7369. [Google Scholar] [CrossRef]
- Lin, Q.-Y.; Wang, J.-Q.; Wu, L.-L.; Zheng, W.-E.; Chen, P.-R. miR-638 represses the stem cell characteristics of breast cancer cells by targeting E2F2. Breast Cancer 2019, 27, 147–158. [Google Scholar] [CrossRef]
- Zhao, F.; Zhong, M.; Pei, W.; Tian, B.; Cai, Y. miR-376c-3p modulates the properties of breast cancer stem cells by targeting RAB2A. Exp. Ther. Med. 2020, 20, 68. [Google Scholar] [CrossRef]
- Chen, J.H.; Huang, W.C.; Bamodu, O.A.; Chang, P.M.; Chao, T.Y.; Huang, T.H. Monospecific antibody targeting of CDH11 inhibits epithelial-to-mesenchymal transition and represses cancer stem cell-like phenotype by up-regulating miR-335 in metastatic breast cancer, in vitro and in vivo. BMC Cancer 2019, 19, 1–13. [Google Scholar] [CrossRef]
- Liang, H.; Xiao, J.; Zhou, Z.; Wu, J.; Ge, F.; Li, Z.; Zhang, H.; Sun, J.; Li, F.; Liu, R.; et al. Hypoxia induces miR-153 through the IRE1alpha-XBP1 pathway to fine tune the HIF1alpha/VEGFA axis in breast cancer angiogenesis. Oncogene 2018, 37, 1961–1975. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Shi, P.; Nie, Z.; Liang, H.; Zhou, Z.; Chen, W.; Chen, H.; Dong, C.; Yang, R.; Liu, S.; et al. Mifepristone Suppresses Basal Triple-Negative Breast Cancer Stem Cells by Down-regulating KLF5 Expression. Theranostics 2016, 6, 533–544. [Google Scholar] [CrossRef]
- Polytarchou, C.; Iliopoulos, D.; Struhl, K. An integrated transcriptional regulatory circuit that reinforces the breast cancer stem cell state. Proc. Natl. Acad. Sci. 2012, 109, 14470–14475. [Google Scholar] [CrossRef]
- Zhao, Q.; Liu, Y.; Wang, T.; Yang, Y.; Ni, H.; Liu, H.; Guo, Q.; Xi, T.; Zheng, L. MiR-375 inhibits the stemness of breast cancer cells by blocking the JAK2/STAT3 signaling. Eur. J. Pharmacol. 2020, 884, 173359. [Google Scholar] [CrossRef]
- Deng, J.; Yang, M.; Jiang, R.; An, N.; Wang, X.; Liu, B. Long Non-Coding RNA HOTAIR Regulates the Proliferation, Self-Renewal Capacity, Tumor Formation and Migration of the Cancer Stem-Like Cell (CSC) Subpopulation Enriched from Breast Cancer Cells. PLoS ONE 2017, 12, e0170860. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Meng, D.; Wang, R. Long non-coding RNA SOX21-AS1 enhances the stemness of breast cancer cells via the Hippo pathway. FEBS Open Bio 2020, 11, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Liu, X.; Zhou, S.; Li, W.; Liu, C.; Chadwick, M.; Qian, C. Long non-coding RNA FGF13-AS1 inhibits glycolysis and stemness properties of breast cancer cells through FGF13-AS1/IGF2BPs/Myc feedback loop. Cancer Lett. 2019, 450, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.; Guo, C.; Xia, T.; Zhang, R.; Zen, K.; Pan, Y.; Jin, L. LncCCAT1 Promotes Breast Cancer Stem Cell Function through Activating WNT/beta-catenin Signaling. Theranostics 2019, 9, 7384–7402. [Google Scholar] [CrossRef]
- Peng, F.; Li, T.T.; Wang, K.L.; Xiao, G.Q.; Wang, J.H.; Zhao, H.D.; Kang, Z.J.; Fan, W.J.; Zhu, L.L.; Li, M.; et al. H19/let-7/LIN28 reciprocal negative regulatory circuit promotes breast cancer stem cell maintenance. Cell Death Dis. 2017, 8, e2569. [Google Scholar] [CrossRef]
- Song, X.; Zhang, X.; Wang, X.; Chen, L.; Jiang, L.; Zheng, A.; Zhang, M.; Zhao, L.; Wei, M. LncRNA SPRY4-IT1 regulates breast cancer cell stemness through competitively binding miR-6882-3p with TCF7L2. J. Cell Mol. Med. 2020, 24, 772–784. [Google Scholar] [CrossRef]
- Lu, G.; Li, Y.; Ma, Y.; Lu, J.; Chen, Y.; Jiang, Q.; Qin, Q.; Zhao, L.; Huang, Q.; Luo, Z.; et al. Long noncoding RNA LINC00511 contributes to breast cancer tumourigenesis and stemness by inducing the miR-185-3p/E2F1/Nanog axis. J. Exp. Clin. Cancer Res. 2018, 37, 289. [Google Scholar] [CrossRef]
- Han, L.; Yan, Y.; Zhao, L.; Liu, Y.; Lv, X.; Zhang, L.; Zhao, Y.; Zhao, H.; He, M.; Wei, M. LncRNA HOTTIP facilitates the stemness of breast cancer via regulation of miR-148a-3p/WNT1 pathway. J. Cell. Mol. Med. 2020, 24, 6242–6252. [Google Scholar] [CrossRef]
- Zheng, A.; Song, X.; Zhang, L.; Lu, G.; Zhao, L.; Mao, X.; Wei, M.; Jin, F. Long non-coding RNA LUCAT1/miR-5582-3p/TCF7L2 axis regulates breast cancer stemness via Wnt/beta-catenin pathway. J. Exp. Clin. Cancer Res. 2019, 38, 305. [Google Scholar] [CrossRef]
- Zhang, Z.; Sun, L.; Zhang, Y.; Lu, G.; Li, Y.; Wei, Z. Long non-coding RNA FEZF1-AS1 promotes breast cancer stemness and tumorigenesis via targeting miR-30a/Nanog axis. J. Cell. Physiol. 2018, 233, 8630–8638. [Google Scholar] [CrossRef]
- Wen, S.; Qin, Y.; Wang, R.; Yang, L.; Zeng, H.; Zhu, P.; Li, Q.; Qiu, Y.; Chen, S.; Liu, Y.; et al. A novel Lnc408 maintains breast cancer stem cell stemness by recruiting SP3 to suppress CBY1 transcription and increasing nuclear beta-catenin levels. Cell Death Dis. 2021, 12, 437. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Liu, C.; Zhao, Q.; Lü, J.; Ding, X.; Luo, A.; He, J.; Wang, G.; Li, Y.; Cai, Z.; et al. Long non-coding RNA CCAT2 promotes oncogenesis in triple-negative breast cancer by regulating stemness of cancer cells. Pharmacol. Res. 2020, 152, 104628. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Hou, Y.; Yang, G.; Zhang, H.; Tu, G.; Du, Y.E.; Wen, S.; Xu, L.; Tang, X.; Tang, S.; et al. LncRNA-Hh Strengthen Cancer Stem Cells Generation in Twist-Positive Breast Cancer via Activation of Hedgehog Signaling Pathway. Stem Cells 2016, 34, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Hou, Y.; Liu, S.; Zhu, P.; Wan, X.; Zhao, M.; Peng, M.; Zeng, H.; Li, Q.; Jin, T.; et al. A Novel Long Non-Coding RNA lnc030 Maintains Breast Cancer Stem Cell Stemness by Stabilizing SQLE mRNA and Increasing Cholesterol Synthesis. Adv. Sci. 2020, 8, 2002232. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Cen, Y.; Chen, J. Long non-coding RNA MALAT-1 contributes to maintenance of stem cell-like phenotypes in breast cancer cells. Oncol. Lett. 2018, 15, 2117–2122. [Google Scholar] [CrossRef]
- Liu, S.; Sun, Y.; Hou, Y.; Yang, L.; Wan, X.; Qin, Y.; Liu, Y.; Wang, R.; Zhu, P.; Teng, Y.; et al. A novel lncRNA ROPM-mediated lipid metabolism governs breast cancer stem cell properties. J. Hematol. Oncol. 2021, 14, 1–23. [Google Scholar] [CrossRef]
- Kim, R.-J.; Nam, J.-S. OCT4 Expression Enhances Features of Cancer Stem Cells in a Mouse Model of Breast Cancer. Lab. Anim. Res. 2011, 27, 147–152. [Google Scholar] [CrossRef]
- Hu, T.; Liu, S.; Breiter, D.R.; Wang, F.; Tang, Y.; Sun, S. Octamer 4 Small Interfering RNA Results in Cancer Stem Cell–Like Cell Apoptosis. Cancer Res. 2008, 68, 6533–6540. [Google Scholar] [CrossRef]
- Bak, M.J.; Furmanski, P.; Shan, N.L.; Lee, H.J.; Bao, C.; Lin, Y.; Shih, W.J.; Yang, C.S.; Suh, N. Tocopherols inhibit estrogen-induced cancer stemness and OCT4 signaling in breast cancer. Carcinogenesis 2018, 39, 1045–1055. [Google Scholar] [CrossRef]
- Almozyan, S.; Colak, D.; Mansour, F.; Alaiya, A.; Al-Harazi, O.; Qattan, A.; Al-Mohanna, F.; Al-Alwan, M.; Ghebeh, H. PD-L1 promotes OCT4 and Nanog expression in breast cancer stem cells by sustaining PI3K/AKT pathway activation. Int. J. Cancer 2017, 141, 1402–1412. [Google Scholar] [CrossRef] [Green Version]
- Xun, J.; Wang, D.; Shen, L.; Gong, J.; Gao, R.; Du, L.; Chang, A.; Song, X.; Xiang, R.; Tan, X. JMJD3 suppresses stem cell-like characteristics in breast cancer cells by downregulation of Oct4 independently of its demethylase activity. Oncotarget 2017, 8, 21918–21929. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Li, J.; Chen, H.; Fu, J.; Ray, S.; Huang, S.; Zheng, H.; Ai, W. Kruppel-like factor 4 (KLF4) is required for maintenance of breast cancer stem cells and for cell migration and invasion. Oncogene 2011, 30, 2161–2172. [Google Scholar] [CrossRef] [PubMed]
- Okuda, H.; Xing, F.; Pandey, P.R.; Sharma, S.; Watabe, M.; Pai, S.K.; Mo, Y.-Y.; Iiizumi-Gairani, M.; Hirota, S.; Liu, Y.; et al. miR-7 Suppresses Brain Metastasis of Breast Cancer Stem-Like Cells By Modulating KLF4. Cancer Res. 2013, 73, 1434–1444. [Google Scholar] [CrossRef] [PubMed]
- Meng, Z.; Liu, Y.; Wang, J.; Fan, H.; Fang, H.; Li, S.; Yuan, L.; Liu, C.; Peng, Y.; Zhao, W.; et al. Histone demethylase KDM7A is required for stem cell maintenance and apoptosis inhibition in breast cancer. J. Cell. Physiol. 2019, 235, 932–943. [Google Scholar] [CrossRef]
- Mimoto, R.; Imawari, Y.; Hirooka, S.; Takeyama, H.; Yoshida, K. Impairment of DYRK2 augments stem-like traits by promoting KLF4 expression in breast cancer. Oncogene 2016, 36, 1862–1872. [Google Scholar] [CrossRef]
- Liu, R.; Shi, P.; Zhou, Z.; Zhang, H.; Li, W.; Zhang, H.; Chen, C. Krüpple-like factor 5 is essential for mammary gland development and tumorigenesis. J. Pathol. 2018, 246, 497–507. [Google Scholar] [CrossRef]
- Shi, P.; Liu, W.; Tala; Wang, H.; Li, F.; Zhang, H.; Wu, Y.; Kong, Y.; Zhou, Z.; Wang, C.; et al. Metformin suppresses triple-negative breast cancer stem cells by targeting KLF5 for degradation. Cell Discov. 2017, 3, 17010. [Google Scholar] [CrossRef] [PubMed]
- Samanta, S.R.; Sun, H.; Goel, H.L.; Pursell, B.M.; Chang, C.; Khan, A.Q.; Greiner, D.L.; Cao, S.; Lim, E.; Shultz, L.D.; et al. IMP3 promotes stem-like properties in triple-negative breast cancer by regulating SLUG. Oncogene 2015, 35, 1111–1121. [Google Scholar] [CrossRef]
- Hu, C.; Xu, L.; Liang, S.; Zhang, Z.; Zhang, Y.; Zhang, F. Lentivirus-mediated shRNA targeting Nanog inhibits cell proliferation and attenuates cancer stem cell activities in breast cancer. J. Drug Target. 2015, 24, 422–432. [Google Scholar] [CrossRef]
- Zhang, C.; Samanta, D.; Lu, H.; Bullen, J.W.; Zhang, H.; Chen, I.; He, X.; Semenza, G.L. Hypoxia induces the breast cancer stem cell phenotype by HIF-dependent and ALKBH5-mediated m 6 A-demethylation of NANOG mRNA. Proc. Natl. Acad. Sci. 2016, 113, E2047–E2056. [Google Scholar] [CrossRef] [Green Version]
- Yin, S.; Cheryan, V.T.; Xu, L.; Rishi, A.K.; Reddy, K.B. Myc mediates cancer stem-like cells and EMT changes in triple negative breast cancers cells. PLoS ONE 2017, 12, e0183578. [Google Scholar] [CrossRef]
- Wang, S.; Wang, N.; Zheng, Y.; Yang, B.; Liu, P.; Zhang, F.; Li, M.; Song, J.; Chang, X.; Wang, Z. Caveolin-1 inhibits breast cancer stem cells via c-Myc-mediated metabolic reprogramming. Cell Death Dis. 2020, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.Z.; Li, S.S.; Zhou, W.; Kang, Z.J.; Zhang, Q.X.; Kamran, M.; Xu, J.; Liang, D.P.; Wang, C.L.; Hou, Z.J.; et al. p62/SQSTM1 enhances breast cancer stem-like properties by stabilizing MYC mRNA. Oncogene 2017, 36, 304–317. [Google Scholar] [CrossRef]
- Zhou, L.; Zhao, L.-C.; Jiang, N.; Wang, X.-L.; Zhou, X.-N.; Luo, X.-L.; Ren, J. MicroRNA miR-590-5p inhibits breast cancer cell stemness and metastasis by targeting SOX2. Eur. Rev. Med Pharmacol. Sci. 2017, 21, 87–94. [Google Scholar]
- Gong, X.; Liu, W.; Wu, L.; Ma, Z.; Wang, Y.; Yu, S.; Zhang, J.; Xie, H.; Wei, G.; Ma, F.; et al. Transcriptional repressor GATA binding 1–mediated repression of SRY-box 2 expression suppresses cancer stem cell functions and tumor initiation. J. Biol. Chem. 2018, 293, 18646–18654. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Song, Y.; Qiu, H.; Liu, Y.; Luo, K.; Yi, Y.; Jiang, G.; Lu, M.; Zhang, Z.; Yin, J.; et al. Downregulation of FOXO3a by DNMT1 promotes breast cancer stem cell properties and tumorigenesis. Cell Death Differ. 2019, 27, 966–983. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Keckesova, Z.; Donaher, J.L.; Shibue, T.; Tischler, V.; Reinhardt, F.; Itzkovitz, S.; Noske, A.; Zürrer-Härdi, U.; Bell, G.; et al. Slug and Sox9 cooperatively determine the mammary stem cell state. Cell 2012, 148, 1015–1028. [Google Scholar] [CrossRef]
- Li, X.; Li, Y.; Du, X.; Wang, X.; Guan, S.; Cao, Y.; Jin, F.; Li, F. HES1 promotes breast cancer stem cells by elevating Slug in triple-negative breast cancer. Int. J. Biol. Sci. 2021, 17, 247–258. [Google Scholar] [CrossRef]
- Zhou, L.; Wang, D.; Sheng, D.; Xu, J.; Chen, W.; Qin, Y.; Du, R.; Yang, X.; He, X.; Xie, N.; et al. NOTCH4 maintains quiescent mesenchymal-like breast cancer stem cells via transcriptionally activating SLUG and GAS1 in triple-negative breast cancer. Theranostics 2020, 10, 2405–2421. [Google Scholar] [CrossRef]
- Zhang, F.; Liu, B.; Deng, Q.; Sheng, D.; Xu, J.; He, X.; Zhang, L.; Liu, S. UCP1 regulates ALDH-positive breast cancer stem cells through releasing the suppression of Snail on FBP1. Cell Biol. Toxicol. 2020, 37, 277–291. [Google Scholar] [CrossRef]
- Doherty, M.R.; Parvani, J.G.; Tamagno, I.; Junk, D.J.; Bryson, B.L.; Cheon, H.J.; Stark, G.R.; Jackson, M.W. The opposing effects of interferon-beta and oncostatin-M as regulators of cancer stem cell plasticity in triple-negative breast cancer. Breast Cancer Res. 2019, 21, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Shen, W.; Zhang, X.; Tang, J.; Zhang, Z.; Du, R.; Luo, D.; Liu, X.; Xia, Y.; Li, Y.; Wang, S.; et al. CCL16 maintains stem cell-like properties in breast cancer by activating CCR2/GSK3beta/beta-catenin/OCT4 axis. Theranostics 2021, 11, 2297–2317. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Cao, J.; Tian, J.H.; Yu, C.Y.; Huang, Q.; Yu, J.J.; Ma, R.; Wang, J.; Xu, F.; Wang, L.B. Mortalin maintains breast cancer stem cells stemness via activation of Wnt/GSK3beta/beta-catenin signaling pathway. Am. J. Cancer Res. 2021, 11, 2696–2716. [Google Scholar] [PubMed]
- Zhang, F.; Li, P.; Liu, S.; Yang, M.; Zeng, S.; Deng, J.; Chen, D.; Yi, Y.; Liu, H. Beta-Catenin-CCL2 feedback loop mediates crosstalk between cancer cells and macrophages that regulates breast cancer stem cells. Oncogene 2021, 40, 5854–5865. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Xu, T.; Tian, T.; Fu, X.; Wang, W.; Li, S.; Shi, T.; Suo, A.; Ruan, Z.; Guo, H.; et al. Tripartite motif 16 suppresses breast cancer stem cell properties through regulation of Gli-1 degradation via the ubiquitin-proteasome pathway. Oncol. Rep. 2015, 35, 1204–1212. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, Y.; Fan, C.; Gao, P.; Wang, X.; Wei, G.; Wei, J. Estrogen promotes stemness and invasiveness of ER-positive breast cancer cells through Gli1 activation. Mol. Cancer 2014, 13, 137. [Google Scholar] [CrossRef]
- Choi, H.S.; Kim, J.H.; Kim, S.L.; Lee, D.S. Disruption of the NF-kappaB/IL-8 Signaling Axis by Sulconazole Inhibits Human Breast Cancer Stem Cell Formation. Cells 2019, 8, 1007. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.; Zhang, X.-T.; Wang, M.-L.; Zheng, H.-Y.; Liu, L.-J.; Wang, Z.-Y. ER-α36-Mediated Rapid Estrogen Signaling Positively Regulates ER-Positive Breast Cancer Stem/Progenitor Cells. PLoS ONE 2014, 9, e88034. [Google Scholar] [CrossRef]
- Bartucci, M.; Dattilo, R.; Moriconi, C.; Pagliuca, A.; Mottolese, M.; Federici, G.; Di Benedetto, A.; Todaro, M.; Stassi, G.; Sperati, F.; et al. TAZ is required for metastatic activity and chemoresistance of breast cancer stem cells. Oncogene 2014, 34, 681–690. [Google Scholar] [CrossRef]
- Xiang, L.; Gilkes, D.M.; Hu, H.; Takano, N.; Luo, W.; Lu, H.; Bullen, J.W.; Samanta, D.; Liang, H.; Semenza, G.L. Hypoxia-inducible factor 1 mediates TAZ expression and nuclear localization to induce the breast cancer stem cell phenotype. Oncotarget 2014, 5, 12509–12527. [Google Scholar] [CrossRef]
- Li, P.; Wang, Y.; Mao, X.; Jiang, Y.; Liu, J.; Li, J.; Wang, J.; Wang, R.; She, J.; Zhang, J.; et al. CRB3 downregulation confers breast cancer stem cell traits through TAZ/beta-catenin. Oncogenesis 2017, 6, e322. [Google Scholar] [CrossRef] [Green Version]
- Ko, Y.-C.; Choi, H.S.; Liu, R.; Lee, D.-S. Physalin A, 13,14-Seco-16, 24-Cyclo-Steroid, Inhibits Stemness of Breast Cancer Cells by Regulation of Hedgehog Signaling Pathway and Yes-Associated Protein 1 (YAP1). Int. J. Mol. Sci. 2021, 22, 8718. [Google Scholar] [CrossRef] [PubMed]
- Gharaibeh, L.F.; Elmadany, N.; Alwosaibai, K.; Alshaer, W. Notch1 in Cancer Therapy: Possible Clinical Implications and Challenges. Mol. Pharmacol. 2020, 98, 559–576. [Google Scholar] [CrossRef] [PubMed]
- Giuli, M.V.; Giuliani, E.; Screpanti, I.; Bellavia, D.; Checquolo, S. Notch Signaling Activation as a Hallmark for Triple-Negative Breast Cancer Subtype. J. Oncol. 2019, 2019, 1–15. [Google Scholar] [CrossRef]
- Baker, A.; Wyatt, D.; Bocchetta, M.; Li, J.; Filipovic, A.; Green, A.; Peiffer, D.S.; Fuqua, S.; Miele, L.; Albain, K.S.; et al. Notch-1-PTEN-ERK1/2 signaling axis promotes HER2+ breast cancer cell proliferation and stem cell survival. Oncogene 2018, 37, 4489–4504. [Google Scholar] [CrossRef] [PubMed]
- Dittmer, J. Breast cancer stem cells: Features, key drivers and treatment options. Semin. Cancer Biol. 2018, 53, 59–74. [Google Scholar] [CrossRef]
- Wang, D.; Xu, J.; Liu, B.; He, X.; Zhou, L.; Hu, X.; Qiao, F.; Zhang, A.; Xu, X.; Zhang, H.; et al. IL6 blockade potentiates the anti-tumor effects of gamma-secretase inhibitors in Notch3-expressing breast cancer. Cell Death Differ. 2018, 25, 330–339. [Google Scholar] [CrossRef]
- Yue, Z.; Yuan, Z.; Zeng, L.; Wang, Y.; Lai, L.; Li, J.; Sun, P.; Xue, X.; Qi, J.; Yang, Z.; et al. LGR4 modulates breast cancer initiation, metastasis, and cancer stem cells. FASEB J. 2017, 32, 2422–2437. [Google Scholar] [CrossRef]
- Satriyo, P.B.; Bamodu, O.A.; Chen, J.-H.; Aryandono, T.; Haryana, S.M.; Yeh, C.-T.; Chao, T.-Y. Cadherin 11 Inhibition Downregulates β-catenin, Deactivates the Canonical WNT Signalling Pathway and Suppresses the Cancer Stem Cell-Like Phenotype of Triple Negative Breast Cancer. J. Clin. Med. 2019, 8, 148. [Google Scholar] [CrossRef]
- Lin, C.-C.; Lo, M.-C.; Moody, R.; Jiang, H.; Harouaka, R.; Stevers, N.; Tinsley, S.; Gasparyan, M.; Wicha, M.; Sun, D. Targeting LRP8 inhibits breast cancer stem cells in triple-negative breast cancer. Cancer Lett. 2018, 438, 165–173. [Google Scholar] [CrossRef]
- Tang, W.; Li, M.; Qi, X.; Li, J. Beta1,4-Galactosyltransferase V Modulates Breast Cancer Stem Cells through Wnt/beta-catenin Signaling Pathway. Cancer Res. Treat. 2020, 52, 1084–1102. [Google Scholar]
- Qin, T.; Li, B.; Feng, X.; Fan, S.; Liu, L.; Liu, D.; Mao, J.; Lu, Y.; Yang, J.; Yu, X.; et al. Abnormally elevated USP37 expression in breast cancer stem cells regulates stemness, epithelial-mesenchymal transition and cisplatin sensitivity. J. Exp. Clin. Cancer Res. 2018, 37, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Wellner, U.; Schubert, J.; Burk, U.C.; Schmalhofer, O.; Zhu, F.; Sonntag, A.; Waldvogel, B.; Vannier, C.; Darling, D.; zur Hausen, A.; et al. The EMT-activator ZEB1 promotes tumorigenicity by repressing stemness-inhibiting microRNAs. Nat. Cell Biol. 2009, 11, 1487–1495. [Google Scholar] [CrossRef] [PubMed]
- Crabtree, J.S.; Miele, L. Breast Cancer Stem Cells. Biomedicines 2018, 6, 77. [Google Scholar] [CrossRef] [PubMed]
- Elaimy, A.L.; Guru, S.; Chang, C.; Ou, J.; Amante, J.J.; Zhu, L.J.; Goel, H.L.; Mercurio, A.M. VEGF-neuropilin-2 signaling promotes stem-like traits in breast cancer cells by TAZ-mediated repression of the Rac GAP beta2-chimaerin. Sci. Signal 2018, 11, eaao6897. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, J.; Li, P.; Jiang, Y.; Chen, H.; Wang, R.; Cao, F.; Liu, P. DLG5 suppresses breast cancer stem cell-like characteristics to restore tamoxifen sensitivity by inhibiting TAZ expression. J. Cell. Mol. Med. 2018, 23, 512–521. [Google Scholar] [CrossRef]
- Zhao, D.; Zhi, X.; Zhou, Z.; Chen, C. TAZ antagonizes the WWP1-mediated KLF5 degradation and promotes breast cell proliferation and tumorigenesis. Carcinogenesis 2011, 33, 59–67. [Google Scholar] [CrossRef]
- Liu, W.; Lu, X.; Shi, P.; Yang, G.; Zhou, Z.; Li, W.; Mao, X.; Jiang, D.; Chen, C. TNF-alpha increases breast cancer stem-like cells through up-regulating TAZ expression via the non-canonical NF-kappaB pathway. Sci. Rep. 2020, 10, 1804. [Google Scholar] [CrossRef]
- Zhi, X.; Zhao, D.; Zhou, Z.; Liu, R.; Chen, C. YAP Promotes Breast Cell Proliferation and Survival Partially through Stabilizing the KLF5 Transcription Factor. Am. J. Pathol. 2012, 180, 2452–2461. [Google Scholar] [CrossRef]
- Zheng, L.; Xiang, C.; Li, X.; Guo, Q.; Gao, L.; Ni, H.; Xia, Y.; Xi, T. STARD13-correlated ceRNA network-directed inhibition on YAP/TAZ activity suppresses stemness of breast cancer via co-regulating Hippo and Rho-GTPase/F-actin signaling. J. Hematol. Oncol. 2018, 11, 1–18. [Google Scholar] [CrossRef]
- Quinn, H.M.; Vogel, R.; Popp, O.; Mertins, P.; Lan, L.; Messerschmidt, C.; Landshammer, A.; Lisek, K.; Château-Joubert, S.; Marangoni, E.; et al. YAP and beta-Catenin Cooperate to Drive Oncogenesis in Basal Breast Cancer. Cancer Res. 2021, 81, 2116–2127. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.-W.; Lin, R.-J.; Yu, J.; Fu, C.-H.; Lai, A.C.-Y.; Yu, J.-C.; Yu, A.L. The expression and significance of insulin-like growth factor-1 receptor and its pathway on breast cancer stem/progenitors. Breast Cancer Res. 2013, 15, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, J.; Chen, W.-B.; Zhang, X.-Y.; Kang, X.-N.; Jin, L.-J.; Zhang, H.; Wang, Z.-Y. HIF-2α regulates CD44 to promote cancer stem cell activation in triple-negative breast cancer via PI3K/AKT/mTOR signaling. World J. Stem Cells 2020, 12, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhang, W.; Phillips, J.B.; Arora, R.; McClellan, S.; Li, J.; Kim, J.-H.; Sobol, R.W.; Tan, M. Immunoregulatory protein B7-H3 regulates cancer stem cell enrichment and drug resistance through MVP-mediated MEK activation. Oncogene 2018, 38, 88–102. [Google Scholar] [CrossRef]
- Wang, X.; Reyes, M.E.; Zhang, D.; Funakoshi, Y.; Trape, A.P.; Gong, Y.; Kogawa, T.; Eckhardt, B.L.; Masuda, H.; Jr, D.A.P.; et al. EGFR signaling promotes inflammation and cancer stem-like activity in inflammatory breast cancer. Oncotarget 2017, 8, 67904–67917. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Fahrmann, J.F.; Lee, H.; Li, Y.J.; Tripathi, S.C.; Yue, C.; Zhang, C.; Lifshitz, V.; Song, J.; Yuan, Y.; et al. JAK/STAT3-Regulated Fatty Acid beta-Oxidation Is Critical for Breast Cancer Stem Cell Self-Renewal and Chemoresistance. Cell Metab. 2018, 27, 1357. [Google Scholar] [CrossRef]
- Chang, R.; Song, L.; Xu, Y.; Wu, Y.; Dai, C.; Wang, X.; Sun, X.; Hou, Y.; Li, W.; Zhan, X.; et al. Loss of Wwox drives metastasis in triple-negative breast cancer by JAK2/STAT3 axis. Nat Commun. 2018, 9, 3486. [Google Scholar] [CrossRef] [PubMed]
- Ge, F.; Chen, W.; Yang, R.; Zhou, Z.; Chang, N.; Chen, C.; Zou, T.; Liu, R.; Tan, J.; Ren, G. WWOX suppresses KLF5 expression and breast cancer cell growth. Chin. J. Cancer Res. 2014, 26, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Liu, Z.; Yu, Z. EGFR Promotes the Development of Triple Negative Breast Cancer Through JAK/STAT3 Signaling. Cancer Manag. Res. 2020, 12, 703–717. [Google Scholar] [CrossRef]
- Woosley, A.N.; Dalton, A.C.; Hussey, G.S.; Howley, B.V.; Mohanty, B.K.; Grelet, S.; Dincman, T.; Bloos, S.; Olsen, S.K.; Howe, P.H. TGFbeta promotes breast cancer stem cell self-renewal through an ILEI/LIFR signaling axis. Oncogene 2019, 38, 3794–3811. [Google Scholar] [CrossRef]
- Tian, J.; Hachim, M.Y.; Hachim, I.Y.; Dai, M.; Lo, C.; Raffa, F.A.; Ali, S.; Lebrun, J.J. Cyclooxygenase-2 regulates TGFbeta-induced cancer stemness in triple-negative breast cancer. Sci. Rep. 2017, 7, 40258. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Santillan, K.; Melendez-Zajgla, J.; Jimenez-Hernandez, L.E.; Gaytan-Cervantes, J.; Muñoz-Galindo, L.; Piña-Sanchez, P.; Martinez-Ruiz, G.; Torres, J.; Garcia-Lopez, P.; Gonzalez-Torres, C.; et al. NF-kappaBeta-inducing kinase regulates stem cell phenotype in breast cancer. Sci. Rep. 2016, 6, 37340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Lee, J.S.; Jie, C.; Park, M.H.; Iwakura, Y.; Patel, Y.; Soni, M.; Reisman, D.; Chen, H. HER2 Overexpression Triggers an IL1α Proinflammatory Circuit to Drive Tumorigenesis and Promote Chemotherapy Resistance. Cancer Res. 2018, 78, 2040–2051. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Torres, S.J.; Benight, N.M.; Karns, R.A.; Lower, E.E.; Guan, J.L.; Waltz, S.E. HGFL-mediated RON signaling supports breast cancer stem cell phenotypes via activation of non-canonical beta-catenin signaling. Oncotarget 2017, 8, 58918–58933. [Google Scholar] [CrossRef]
- Hinohara, K.; Kobayashi, S.; Kanauchi, H.; Shimizu, S.; Nishioka, K.; Tsuji, E.; Tada, K.; Umezawa, K.; Mori, M.; Ogawa, T.; et al. ErbB receptor tyrosine kinase/NF-kappaB signaling controls mammosphere formation in human breast cancer. Proc. Natl. Acad. Sci. USA 2012, 109, 6584–6589. [Google Scholar] [CrossRef]
- Wei, L.; Liu, T.T.; Wang, H.H.; Hong, H.M.; Yu, A.L.; Feng, H.P.; Chang, W.W. Hsp27 participates in the maintenance of breast cancer stem cells through regulation of epithelial-mesenchymal transition and nuclear factor-kappaB. Breast Cancer Res. 2011, 13, R101. [Google Scholar] [CrossRef]
- Zhang, P.; Liu, Y.; Lian, C.; Cao, X.; Wang, Y.; Li, X.; Cong, M.; Tian, P.; Zhang, X.; Wei, G.; et al. SH3RF3 promotes breast cancer stem-like properties via JNK activation and PTX3 upregulation. Nat. Commun. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Barker, A.D.; Sigman, C.C.; Kelloff, G.J.; Hylton, N.M.; A Berry, D.; Esserman, L.J. I-SPY 2: An Adaptive Breast Cancer Trial Design in the Setting of Neoadjuvant Chemotherapy. Clin. Pharmacol. Ther. 2009, 86, 97–100. [Google Scholar] [CrossRef]
- Wang, H.; Yee, D. I-SPY 2: A Neoadjuvant Adaptive Clinical Trial Designed to Improve Outcomes in High-Risk Breast Cancer. Curr. Breast Cancer Rep. 2019, 11, 303–310. [Google Scholar] [CrossRef]
Figure 1.
Critical signal transduction pathway networks associated with BCSCs.


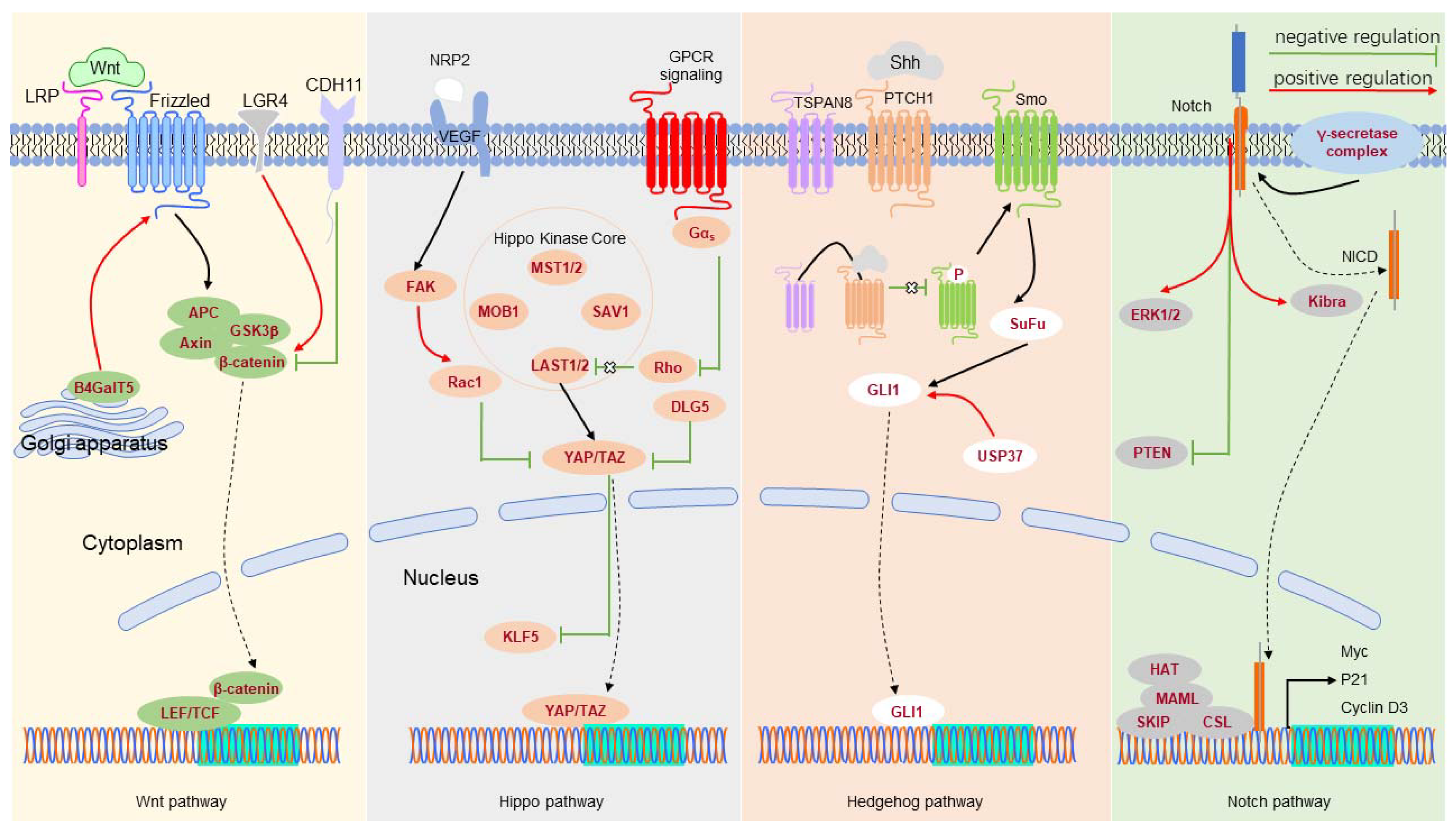{kind=link}
Table 1.
miRNAs regulating BCSCs.
| Names | Mechanism | References |
|---|---|---|
| miRNA-200c | inhibits the expression of Pin1, BMI1 and Suz12. | [42] |
| Let-7 family | inhibits the Wnt signaling pathway. | [43] |
| miR-34 family | targets Notch1. | [47] |
| miRNA-146a | promotes the asymmetric division of BCSCs. | [48] |
| miRNA-760 | inhibits the expression of NANOG. | [49] |
| miR-422a | reduces the expression of Proteolipid Protein 2 (PLP2). | [50] |
| miRNA-142-3p | targets β-catenin. | [51] |
| miRNA-1 | targets ecotropic virus integration site 1 (EVI1). | [52] |
| miRNA-128-3p | downregulates NIMA related kinase 2 (NEK2) to inhibit the Wnt signaling pathway. | [53] |
| miRNA-638 | reduces the expression of E2F2. | [54] |
| miR-376c-3p | reduces the expression of RAB2A. | [55] |
| miRNA-221/222 | inhibits PTEN expression. | [44] |
| miR-20b-5p | bidirectionally regulates cyclin D1 and E2F1. | [46] |
| miR-335 | inhibits cadherin 11 (CDH11), β-catenin, and vimentin. | [56] |
| miR-153 | downregulates hypoxia-inducible factor 1 subunit alpha (HIF1ɑ) and KLF5. | [57,58] |
| miR-145 | suppresses BCSCs growth by inhibiting KLF4. | [59] |
| miR-375 | decreases BCSCs by interrupting the JAK2-STAT3 pathway. | [60] |
Table 2.
LncRNAs regulating BCSCs.
| Names | Mechanism | References |
|---|---|---|
| HOTAIR | regulates miR-34a to upregulate the expression of SOX2 in BCSCs. | [61] |
| SOX21-AS1 | inhibits the Hippo signaling pathway. | [62] |
| CCAT1 | enhances the expression of T-cell factor 4 (TCF4) to activate the Wnt signaling pathway. | [64] |
| H19 | forms a two-way negative feedback loop with miRNA let-7 and LIN28. | [65] |
| SPRY4-IT1 | sponges miR-6882-3p. | [66] |
| LINC00511 | regulates the miR-185-3p/E2F1/NANOG axis. | [67] |
| HOTTIP | acts as an miR-148a-3p sponge and regulates Wnt 1. | [68] |
| LUCAT1 | competitively binds to miR-5582-3p and transcription factor 7 like 2 (TCF7L2) to enhance the Wnt/β-catenin pathway. | [69] |
| FEZF1-AS1 | regulates the miR-30a/NANOG signal pathway. | [70] |
| Lnc408 | recruit specificity protein 3 (Sp3) to inhibit chibby family member 1 (CBY1) and β-catenin expression. | [71] |
| CCAT2 | upregulates OCT4-PG1 and the miR-205-Notch1 pathway. | [72] |
| Hh | stimulates hedgehog signaling. | [73] |
| Lnc030 | interacts with poly (RC) binding protein 2 (PCBP2) to stabilize squalene epoxidase (SQLE) and activate the PI3K/Akt signaling pathway. | [74] |
| MALAT1 | positively regulates SOX2. | [75] |
| ROPM | maintains group XVI phospholipase A2 (PLA2G16) to facilitate lipid metabolism, thereby activating the Wnt/β-catenin pathway. | [76] |
| FGF13-AS1 | regulates the IGF2BPs/Myc feedback loop. | [63] |
Table 3.
Transcription factors and co-activators regulating BCSCs.
| Names | Hallmarks | References |
|---|---|---|
| OCT4 | OCT4 promotes sphere formation of BCSCs in vitro, while inhibition of OCT4 induces apoptosis, reduces BCSC characteristics, and inhibits tumor growth. | [77,78,79] |
| KLF4 | KDM7A and DYRK2 increase BCSCs by upregulating KLF4 expression, and miR-7 inhibits KLF4 to inhibit BCSCs self-renewal and invasion. | [83,84,85] |
| KLF5 | Mifepristone and metformin inhibit KLF5 and BCSC. PRMT5 increases stemness of BCSC by stabilizing KLF5. | [37,58,87] |
| C-MYC | Caveoli-1 inhibits C-MYC-mediated BSCS metabolic reprogramming and p62 stabilizes C-MYC to enhance BCSC properties. | [92,93] |
| SOX2 | Knockdown of SOX2 attenuates stemness of BCSC. TRPS1 and FOXO3a inhibit SOX2 the expression and tumorigenesis of BCSCs. | [94,95,96] |
| SOX9 | Knockdown of SOX9 significantly inhibits the tumorigenicity of MDA-MB-231 cells. | [97] |
| SLUG | Hes family BHLH transcription factor 1 (HES1) increases SLUG transcription and BCSC stemness. Interestingly, the Notch4/SLUG/Gas1 axis maintains mesenchymal-like BCSCs. | [98,99] |
| SNAIL | Uncoupling Protein 1 (UCP1)-mediated fructose-bisphosphatase 1 (FBP1) expression promotes BCSC properties, which can be reversed by SNAIL. Interferon beta (IFN-β restrains SNAIL-induced tumor initiation. | [100,101] |
| β-catenin | β-catenin facilitates BCSC properties through CCL2-mediated macrophage polarization and infiltration. CCL16 and mortalin maintain the stemness of BCSCs by promoting the translocation of β-catenin. | [102,103,104] |
| GLI1 | Tripartite motif 16 (TRIM16) inhibits BCSCs partially via Glioma-related homologue 1 (GLI1). In contrast, estrogen promotes BCSCs by activating GLI1. | [105,106] |
| p65 | p65 is important for BCSC survival. | [107] |
| ERα | Reduction in expression of ER inhibits CSC tumor-seeding efficiency. | [108] |
| FOXO3a | FOXO3a inhibits the characteristics and tumorigenesis of BCSCs by negatively regulating SOX2. | [96] |
| TAZ | Overexpression of transcription activator with PDZ-binding motif (TAZ) in BCSCs enhances tumorigenicity and cell migration. The ability of HIF1 and Crumbs homolog 3 (CRB3) to maintain or induce BCSC properties is partially achieved by activating TAZ. | [109,110,111] |
| YAP | Downregulation of YAP1 has a negative effect on BCSC tumorigenicity and stemness markers. | [112] |
Table 4.
Signal transduction pathways regulating BCSCs.
| Related Signaling Pathway Factors | Mechanism | References | |
|---|---|---|---|
| NOTCH | NOTCH-1 | inhibits PTEN and activates ERK1/2 to maintain BCSCs. | [115] |
| NOTCH-2 | promotes BCSC survival. | [116] | |
| NOTCH-3 | inhibits BCSC self-renewal by IL6/STAT3. | [117] | |
| NOTCH-4 | promotes mammosphere formation. | [114] | |
| WNT | LGR4 | promotes BCSCs. | [118] |
| CDH11 | inhibits TNBC cell stemness. | [119] | |
| LRP8 | decreases the percentage of BCSCs. | [120] | |
| B4GalT5 | maintains BCSCs by stabilizing Frizzled. | [121] | |
| HH | TSPAN8 | interacts with the SHH-PTCH1 complex to promote stemness of breast cancer. | [14] |
| USP37 | increases Smo and GLI1 expression to enhance BCSC characteristics. | [122] | |
| HIPPO | VEGF/NRP2 | activates TAZ to enhance BCSC sphere-forming ability. | [125] |
| DLG5 | enhances TAZ activity to maintain BCSC self-renewal. | [126] | |
| TNF-ɑ | induces TAZ expression to increase BCSC stemness. | [127,128] | |
| STARD13 | reduces YAP/YAZ activity, thereby inhibiting the formation of BCSCs. | [130] | |
| RTK | IGF-1R | maintains BCSCs by activating the PI3K/Akt/mTOR pathway. | [132] |
| HIF-2ɑ | inhibits BCSCs by inhibiting the PI3K/Akt/mTOR pathway. | [133] | |
| B7-H3 | activates MEK and increases BCSC proportions. | [134] | |
| SGCE | stabilizes EGFR levels, thereby fostering breast cell stemness. | [16] | |
| JAK/STAT3 | WWOX | hinders STAT3 activation to block breast cancer cell proliferation and metastasis. | [137] |
| EGFR | promotes STAT3 phosphorylation to facilitate tumor cell proliferation and invasion. | [139] | |
| TGF-β | LIFR | drives the formation of BCSCs. | [140] |
| Fibronectin, COX2 | enhances the self-renewal capacity of BCSCs. | [141] | |
| NF-κB | IL-1α | maintains BCSCs. | [143] |
| HGFL-RON | supports the self-renewal capacity of BCSCs. | [144] | |
| HRG | stimulates mammosphere formation. | [145] | |
| Hsp27 | degrades IκBα to maintain BCSCs. | [146] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chen, W.; Zhang, L.; Liu, S.; Chen, C. Advances in Biomarkers and Endogenous Regulation of Breast Cancer Stem Cells. Cells 2022, 11, 2941. https://doi.org/10.3390/cells11192941
AMA Style
Chen W, Zhang L, Liu S, Chen C. Advances in Biomarkers and Endogenous Regulation of Breast Cancer Stem Cells. Cells. 2022; 11(19):2941. https://doi.org/10.3390/cells11192941
Chicago/Turabian StyleChen, Wenmin, Lu Zhang, Suling Liu, and Ceshi Chen. 2022. "Advances in Biomarkers and Endogenous Regulation of Breast Cancer Stem Cells" Cells 11, no. 19: 2941. https://doi.org/10.3390/cells11192941
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.