
Integrating the Roles of Midbrain Dopamine Circuits in Behavior and Neuropsychiatric Disease


1
Department of Neurobiology and Behavior, Renaissance School of Medicine at Stony Brook University, Stony Brook, NY 11794, USA
2
Medical Scientist Training Program, Renaissance School of Medicine at Stony Brook University, Stony Brook, NY 11794, USA
*
Author to whom correspondence should be addressed.
Biomedicines 2021, 9(6), 647; https://doi.org/10.3390/biomedicines9060647
Submission received: 29 April 2021
/
Revised: 1 June 2021
/
Accepted: 3 June 2021
/
Published: 7 June 2021
(This article belongs to the Special Issue Dopamine in Health and Disease)
Abstract
:Dopamine (DA) is a behaviorally and clinically diverse neuromodulator that controls CNS function. DA plays major roles in many behaviors including locomotion, learning, habit formation, perception, and memory processing. Reflecting this, DA dysregulation produces a wide variety of cognitive symptoms seen in neuropsychiatric diseases such as Parkinson’s, Schizophrenia, addiction, and Alzheimer’s disease. Here, we review recent advances in the DA systems neuroscience field and explore the advancing hypothesis that DA’s behavioral function is linked to disease deficits in a neural circuit-dependent manner. We survey different brain areas including the basal ganglia’s dorsomedial/dorsolateral striatum, the ventral striatum, the auditory striatum, and the hippocampus in rodent models. Each of these regions have different reported functions and, correspondingly, DA’s reflecting role in each of these regions also has support for being different. We then focus on DA dysregulation states in Parkinson’s disease, addiction, and Alzheimer’s Disease, emphasizing how these afflictions are linked to different DA pathways. We draw upon ideas such as selective vulnerability and region-dependent physiology. These bodies of work suggest that different channels of DA may be dysregulated in different sets of disease. While these are great advances, the fine and definitive segregation of such pathways in behavior and disease remains to be seen. Future studies will be required to define DA’s necessity and contribution to the functional plasticity of different striatal regions.
Keywords:
dopamine; dorsal striatum; ventral striatum; substantia nigra pars compacta; ventral tegmental area; neural circuits; auditory striatum; dorsomedial striatum; dorsolateral striatum; nucleus accumbens; hippocampus; Parkinson’s disease; addiction; Alzheimer’s disease; schizophrenia; D1 Receptor; D2 Receptor; dopamine transporter1. Introduction
A central goal in neuroscience is to understand the basic functions of the brain and how these functions may go awry in neuropsychiatric and behavioral disorders. In successful cases, there is a bidirectional relationship in which basic studies inform and learn from pathological phenomena. An emerging exemplar of this is dopamine (DA), a major neurotransmitter released throughout the brain that has drawn widespread interest from basic and clinical neurosciences. DA neurons degenerate and a lack of striatal DA is linked to the movement deficits seen in Parkinson’s disease, a major neurodegenerative disorder. Indeed, clinical treatments and rodent models of recovery draw upon DA repletion in the CNS [1,2]. These observations form the basis of major neuroscience curricula and how the basal ganglia contribute to movement. Another major success story in cognitive neuroscience is the finding that DA acts as a reward prediction error (RPE) signal and contributes to reinforcement learning. Linked to major brain functions in movement and reward processing, it is unsurprising that many neuropsychiatric diseases arise from DA dysregulation. The goal of this review is to dissect the neural circuit-dependent roles of DA in behavior and attempt to link these functions to DA’s dysregulation in different brain diseases.
We attempt to draw insight from what we can learn from both basic and pathological perspectives on DA. While there have been major advances, many lines of DA investigation in health and disease have remained largely separate. It is critical to cross fields and start asking questions such as “Do models of Parkinson’s disease rely on DA dysregulation across the entire striatum or specific subregions? What roles do striatal subdivisions have in addiction pathogenesis? Or schizophrenia?”. When considering the cognitive and perceptual deficits of Parkinson’s disease and other DA dysregulation disorders, it is tantalizing to speculate on the nuances of DA’s function. We adopt a current view in the literature that there may be separate channels through which DA functions, as determined by specific anatomical projections [3,4,5]. While we provide some evidence that supports this hypothesis, it should be noted that it is still an open question whether a single or single set of principles underlie DA’s function in multiple behaviors. Nevertheless, we systematically discuss the basic function of major neural circuits regulated by DA. Specifically, we dissect circuits in the dorsal striatum, the ventral striatum, and the hippocampus. We attempt to be comprehensive in this regard by delineating general function, Dopamine Receptor-1 (D1R) vs. Dopamine Receptor-2 (D2R) medium spiny neuron pathway differences, and DA modulation therein. We note along the way the importance of the D1R/D2R dichotomy, as this relates to theoretical and clinical aspects of using dopaminergic pharmacological agents. Within this paradigm, it is widely appreciated that DA tends to increase the activity of D1R-expressing neurons and decrease the activity of D2R-expressing neurons [6,7,8,9]. We discuss how this physiological mechanism extends to behavioral phenomena. Then, we discuss how major brain diseases are related to this circuit-function framework. As the literature regarding DA in behavior and disease is expansive, we primarily focused on recent advances in DA regulation and do not cover the remarkable literature regarding the role of non-DA cell types in the regulation of DA output. For these relevant studies, please see [10,11].
2. Behavior to Neuroanatomical Circuit-Function
2.1. Anatomical Overview
It has long been appreciated that DA transmission occurs broadly throughout the brain and exerts effects on an arsenal of behaviors. Canonical organization of dopamine-containing nuclei was established in the 1960s and 1970s with the development of the then introduced formaldehyde histochemical techniques [12]. Carlsson and colleagues took advantage of the chemical interaction of catecholamines and formaldehyde, which produces fluorescent by-products [13,14]. Later, the development of immunohistochemical methods for the detection of catecholamine-synthesizing enzymes allowed for the elaboration of all catecholaminergic nuclei [15,16]. Critically, this work determined that the substantia nigra pars compacta (SNpc) and the ventral tegmental area (VTA) are major midbrain complexes that are dopaminergic in nature. A general picture arises in which dopaminergic neurons are densely populated in the VTA and more so in the SNpc [11,17,18]. The SNpc primarily projects to the dorsal striatum (DS; caudate/putamen in primates) while the VTA primarily projects to the ventral striatum (VS)/nucleus accumbens (NAc). Previous studies have created a dichotomous understanding of the DA nuclei that implicates the SNpc’s nigrostriatal pathway in movement and the VTA’s mesolimbic pathway in reward processing and motivation [19,20,21,22]. This dichotomy, while oversimplified considering recent studies (see [3,23,24,25]), has been useful to understand neural and behavioral dysfunction in disease. We will discuss the basic function of the medial and lateral DS subdivision (DMS and DLS) and relevant studies on the role of DA modulation shown here. Similarly, we will discuss parallel roles in behavior for the VS.
Along the way, we also discuss the respective functions of the D1R and D2R genetically defined anatomical pathways. We note here that the D1R and D2R receptors are part of the D1-like family (including D1R and D5R) and D2-like family (including D2R, D3R, and D4R), respectively [7,8,26]. For the sake of focusing on behavioral functions of DA, we have limited our discussion to the D1R and D2R pathways, which are the primary constituents of both the dorsal and ventral striatum [27,28]. However, it is important to note that the other receptors (D3-5) are pharmacologically, behaviorally, and clinically relevant. For instance, the D3R is expressed in and regulates the prefrontal cortex, with implications in modulating memory function in schizophrenia [26,29]. While not delineated in this review, we encourage the consideration of all DA receptor subtypes in order to adopt a complete framework of DA in behavior and disease. After discussing the major striatal subregions, we lastly add in research performed in the hippocampal field regarding DA. In Figure 1, we summarize the different behavioral functions related to the different parts of the DS, as well as the different roles of DA associated with the striatum in these regions. In Table 1, we summarize the key points from our review of recent research. We note here that these ongoing observations and definitive conclusions about different striatal subregions may require further systematic study.
2.2. SNpc and the Dorsal Striatum
The bulk of the DS projects to the SNpc and receives projections back from SNpc DA neurons [19,21]. Overall, the DS has been divided into two major behavioral pathways on the basis of molecular and anatomical evidence, the D1 “direct” and D2 “indirect” pathways. In addition, the DS has traditionally been divided into two behaviorally distinct regions, the DMS and the DLS. The function of these two structures have been attributed to goal-directed behaviors and habit-based behaviors, respectively [30,31,32]. A related categorization, the DMS is denoted as the associative striatum, while the DLS is the sensorimotor striatum. There is a parallel anatomy that suggests that the DMS and DLS receive differential cortical inputs, with the DMS receiving more limbic inputs and the DLS receiving more motor inputs [33,34]. We could consider that, if the DMS and DLS serve different functions, it is perhaps the case that their respective DA inputs also serve different functions.
2.2.1. DMS Circuits in Goal-Directed Behavior and Beyond
DMS General Function: The DMS is a striatal subregion important for goal-directed behaviors. Observed in a variety of reward-based behaviors, as animals first learn a task to obtain reward, they tend to choose a goal-oriented rather than a stimulus-oriented strategy. For early lever pressing or navigational tasks, this means that the animal interacts with the environment more flexibly in order to obtain a reward. In early studies, groups found that, after lesioning the DMS, rats would perform water maze tasks in a more cue-responsive manner than a goal directed one [35,36]. The opposite was the case for animals with DLS lesions. This divide between the DMS/DLS was further supported by in vivo recordings that demonstrate that DMS activity is greater earlier on during instrumental training, whereas DLS activity is more prominent after extended training. Thus, the current view is that the DMS is important for goal-directed behaviors whereas the DLS is more important for habit-based or skill-honing behaviors.
In line with a role in goal-oriented behavior, DMS neuronal activity correlates with flexible switches in task contingencies in rats that perform a dynamic go/no-go task [37]. In another task dimension, it has been demonstrated that the DLS performs action generalization whereas the DMS is better suited for action discrimination and differential utilization. This study suggests that the DMS and DLS may behaviorally compete with one another for adopting particularly motor strategies when engaging in the environment. Adding to its role in flexible action selection, the DMS is also implicated in perceptual actions and sensory encoding. This poises the DMS as a structure to encode actions within the perceptual contexts of the environment, which is related to previous studies that implicated the DMS in navigational and water maze tasks.
The DMS indeed receives visual, somatosensory, and some sources of auditory input [33,34]. Interestingly, the DMS functionally responds to both somatosensory and visual input, whereas the DLS primarily responds to somatosensory inputs [38]. In an interesting brain-machine interface study, mice were trained to control visual cortical activity in order to generate favorable online task outcomes [39]. They found that inactivation of the DMS impairs the ability of mice to learn this visual cortical control task. This study allows us to speculate whether visual corticostriatal projections or other parts of this basal ganglia segment may allow for special types of visual learning. Furthermore, activation of DMS striatal neurons during a perceptual decision-making task causes contralateral bias. Importantly, in this task, the mode of activation occurred only during stimulus onset, suggesting a role for the visual stimulus encoding in the DMS. In support of this, a recent systematic study utilized neuropixel probes and widefield calcium imaging to simultaneous probe the activity of cortical and striatal activity across the medial–lateral axis. During a visual action association task, they found that DMS neurons responded to visual stimuli in a manner consistent with cortical input activity, whereas DLS neurons responded more to rewarded sensation or licking action. This study dynamically demonstrated distributed perceptual responses and is consistent with previous studies.
D1 vs. D2 in DMS: Many studies that provide evidence for the canonical divide between the direct and indirect pathway have been carried out in the dorsomedial striatum [40]. Mechanisms of DA strongly support the behavioral divide in that DA tends to excite or enhance synaptic transmission in D1R MSNs and depress activity in D2R MSNs [6,8]. This divide has been investigated using the early adoption of optogenetic tools. These early experiments indicate that D1R MSNs reinforce lever pressing behavior whereas D2R MSN stimulation serves to signal transient punishment for the animal. Consistent with this, Balliene’s group recently demonstrated that learning-associated plasticity has opposite signs for the two pathways in the DMS, with synaptic enhancement of D1R MSNs and depression in D2R MSNs [41]. Other studies in value-based decision making have demonstrated that both D1R and D2R’s MSNs play a role in decisions. In line with this, it was previously demonstrated that the DMS direct pathway is involved in action selection [42]. Here, optogenetic stimulation of the D1R MSN population induces contraversive bias, whereas the D2R population induces ipsiversive bias. It is important to note that the above two studies dissect different behaviors, with the former focusing on outcome evaluation and the latter on ongoing decision output structure. Thus, regarding the direct vs. indirect pathway dichotomy, both appear to play opposing roles in different forms of goal-directed decision-making and learning. While more work will be required to dissect anatomical differences in this opponency, it explains different aspects of dopaminergic antagonists such as loss of motivation and deficits in learning.
DA in Dorsomedial Striatum: DA release in the DMS has been found to correspond to reward, movement, and aversive behavioral cues. Having discussed the behavioral divide between the DMS and DLS, several recent studies have attempted to address differences in DA signaling in these two regions. For instance, Lerner et al. studied the SNpc-DMS and SNpc-DLS circuits as being separate both in anatomy and function [18]. Using whole brain mapping with rabies viral targeting and brain clearing strategies, they found that DLS-projecting SNpc neurons receive both DLS and DMS inputs but have a bias for DLS input. The converse is true for DMS-projecting SNpc DA neurons. This suggests that the SNpc DA neurons projecting to the DMS or DLS are differentially regulated by reciprocal GABAergic MSN inputs. In agreement with the prior literature, they also demonstrated that the DLS-projection SNpc neurons are located more laterally in the SNpc, while the DMS-projected neurons are located more medially. Interestingly, they found that other subcortical and cortical areas appear to project to these compartments of the SNpc somewhat equivalently. Consistent with work demonstrating the role of DMS in action selection, DA inputs to this region appear to encode contralateral actions in contrast with reward prediction error encoding demonstrated by the ventral striatal DA. Interestingly, in a recent preprint study by Lerner’s group, DA responses in the DMS were found to correspond to compulsive behaviors. They showed that task-relevant increases in DMS DA corresponded to which individual animal would express compulsive behavior. Optogenetic stimulation of DA terminals in the DMS increased compulsive but not habitual types of behavior. This has implications for our understanding of DA pharmacological agents in OCD and attention disorders [32]. While there is a gradient of encoding a variety of variables amongst DA neurons [3], it appears that DMS DA may correlate well with what the striatal target neurons seem to perform in terms of goal directed behavior.
2.2.2. DLS Circuits—Habit Formation and Beyond
DLS General Function: A large body of literature suggests that the DLS is involved in sensorimotor encoding and habit formation. Appropriately, in comparison to the DMS, the DLS receives more input from the primary somatosensory and primary motor cortices [33,34]. In contrast with the DMS, DLS striatal neurons appear to have higher activity in rodents that have undergone extended bouts of training, and thus appear to be more correlated with skill honing and habit execution. In line with this, a recent study showed elegantly that the DLS is increasingly modulated by DA as an animal becomes an expert at an instrumental task. However, just as there is regional topographic segregation in the motor cortex, one can ask whether this applies to the DMS/DLS subdivisions for action control. A recent study topographically mapped out the different pathways in basal ganglia, comparing the DMS, the DLS, and the ventrolateral dorsal striatum [43]. This group showed that the DMS pathway is important for contraversive turning whereas the ventrolateral dorsal striatum is important for contraversive licking. They argue that, while previous studies have implicated the DLS in both contralateral licking and turning, other regions of the DS more potently control these specific actions. This is overall consistent with the literature that suggests that basal ganglia circuits are important for contralateral actions. It will be important to consider how these data fit in the context of goal-directed vs. habitual dichotomies. Further, it will be important to understand how sensory input into these different regions may map onto specific behaviors. For instance, given the preponderance of S1 inputs in the lateral DS, one would predict that whisker stimulation best triggers licking behaviors in a habitual manner.
Previous studies show the importance of the dorsolateral striatum in simple discrimination tasks [44]. In addition, the DLS has also been implicated in perceptual-based behaviors that require somatosensation. Whisker-based choices recruit the DLS circuit and appear to be important for the timing of actions initiated by the DLS [45]. In addition, the DLS striatal neurons seem particularly to respond to somatosensory stimulation, whereas DMS striatal neurons also respond to visual stimuli [38]. Lastly, the DLS striatonigral pathway is involved in encoding whisker stimulation responses in a Go/No-go paradigm. In light of these studies, the DLS is poised to perform basic sensorimotor integration with strong communication with motor and somatosensory cortices (Figure 1).
D1 vs. D2: As with the DMS, the DLS also consists primarily of MSNs of the direct and indirect pathways. Compared with the DMS, the evidence for segregated D1 and D2 pathways in the DLS is less abundant. Recordings in the central part of the DS are consistent with this, demonstrating that both D1 and D2 pathways are concurrently active during movement [46]. A newer emerging model postulates that the pathways are intertwined, and both are required for purposeful action execution. Consistent with this, evidence suggests that the D1 and D2 pathways in the DLS both act to reinforce optogenetically linked lever press actions. However, the action strategies supported by the D1 and D2 pathways are different. In tangent with this study, other groups have also shown nuanced learning roles for the D1 and D2 pathways in the DLS [47,48]. These studies indicate that the D1R pathway in the DLS is critical for initial action contingency adoption, while the D2R pathway may play more of a role in sculpting and refining behavioral strategies. In the context of habit formation and expression, an imaging study conducted indicates that both pathways play time-specific roles in habit expression [49]. Interestingly, in this study, activation of the D2R pathway did not predict habit suppression, and both pathways underwent strengthened plasticity during habit formation. In relation to sensorimotor behaviors, Sippy et al. demonstrated that D1R pathway activity correlates with whisker sensory responses and that optogenetic inhibition of this pathway reduces conditioned licking responses. In conclusion, both D1R/D2R pathways in the DLS appear to be important for learning and habits but to different extents.
DA in Dorsolateral Striatum: DA in the DLS has been documented to correlate with sensory, motor, and learning variables. This is similar but distinct from the variables seen in the DMS. As lesions of the DLS lead to impairment of habit formation, removing DA input to this region via neurotoxin delivery also results in impaired habit formation [50,51]. In comparison to the DMS, Lerner et al. found the DLS DA dynamics appear to have similar reward processing but a greater response to aversive events. A recent study showed elegantly that the DLS becomes increasingly modulated by DA as an animal becomes an expert at an instrumental task. In an interesting set of studies, Palmiter’s group showed that, in dopamine deficient mice, rescuing DA only in the DLS is sufficient to preserve the ability to learn cognitive tasks, albeit with a slower rate of learning [52]. This is important because it indicates that DA processing in the DLS may contribute to aspects of learning and motivation, which are attributes primarily associated with the DMS or VS. It should be noted that the mice generated in these studies have been found to have severe deficits in learning and motivation that are comparable to those in mice with bilateral OHDA depletion models. Corresponding to the previously discussed sensorimotor role of the DLS, Ketzef et al. found that DA depletion in the DLS impairs bilateral somatosensory processing in MSNs [53]. This work may reflect some of the sensory deficits seen in Parkinson’s disease.
2.2.3. Auditory Striatum
General Function: Early on in the striatal literature, LeDoux’s group characterized tonal responses in the striatum of anesthetized cats. Since then, a variety of groups have probed the sensory representation in the striatum across visual, auditory, and somatosensory modalities. The auditory cortex projects strongly, particularly to the tail or caudal extreme of the DS. These projections are not defined or experimentally investigated by studies on the more rostral DMS and DLS. Based on its strong auditory cortical input, here, we synonymously refer to the “tail” or “caudal” striatum (as denoted in previous literature) as the auditory striatum. We note here that we do not discuss the auditory striatum in non-rodent species; there is anatomical and behavioral evidence for an auditory striatum correlates in nonhuman primate (extensively reviewed here). There are several anatomical characteristics of the auditory striatum that distinguish it from the DMS and DLS (Figure 1). Anatomically, the auditory striatum receives projections primarily from sensory cortices and thalami [33,34]. Additionally, it is innervated by the lateral region of the SNpc or the substantia nigra pars lateralis which receives efferent projections that differs from the rest of the SNpc. Furthermore, this region has a subregion that is particularly rich in D1R MSNs (adjacent to the globus pallidus externus), unlike the rest of the dorsal striatum, which has an approximately even distribution of D1R and D2R MSNs [54]. Taken together, the auditory striatum is anatomically poised to perform functions that differ from the DMS/DLS by specifically integrating sensory inputs. However, as reviewed in the above sections, there is electrophysiological and in vivo evidence for strong visual and somatosensory processing in the DMS and DLS, respectively.
Zador’s group found that auditory corticostriatal projections functionally control arbitrary sound–action decisions in rats. This study is important because it depicts a potential pathway through which arbitrary sound representation in the auditory cortex can be translated into actions through the basal ganglia. This serves as a general mechanism through which the basal ganglia may contribute to perceptual decision making. Furthermore, we found that the potentiation of auditory corticostriatal synapses underlies the acquisition of sound–action learning [55]. This depicts a neural mechanism through which sound–action learning can occur through specific corticostriatal interactions. This is consistent with DA theory in how DA modulates learning in the striatum by potentiating corticostriatal synapses. Beyond cortical connections, we have also shown that the auditory thalamus functionally regulates the region of the auditory striatum [56]. These thalamic connections appear to be important for controlling the gain of sound representation within the auditory striatum. Similar to corticostriatal or thalamostriatal intervention, optogenetic activation of D1R MSNs in the auditory striatum demonstrates that it can bias auditory decisions in a manner that is consistent with tonal stimuli but not simple contraversive choice selection [57]. Thus, the striatal MSNs in the region are important for stimulus encoding and decision-making enforcement. However, how this region fits into the VS to DLS continuum is unclear and it may potentially be the case that this region has specialized sensory functions. More studies examining the auditory striatum in different behavioral paradigms are needed to draw critical comparisons.
D1R vs. D2R in the Auditory Striatum: Like its general role in the basal ganglia, the auditory striatum’s direct and indirect pathways remain to be clarified. As mentioned previously, Jaramillo’s group optogenetically manipulated the D1R pathway and demonstrated a role for auditory-guided decisions that was distinct from a general movement bias demonstrated in other regions of the dorsal striatum [57]. A recent paper from Zhang’s group characterized the role of the auditory striatum in defensive behaviors [58]. They demonstrated that auditory corticostriatal projections targeting D2R MSNs are particularly important for freezing behavior in response to fear-inducing auditory cues. With these two studies combined, it appears that the function of D1R/D2R MSNs may support traditional Go/No-go basal ganglia models specifically in auditory or sensory-dependent behavioral contexts. Further studies will be required to fully characterize the function and input/output dynamics of these pathways.
DA in the Auditory Striatum: It has been shown that the posterior extreme of the striatum receives differential cortical, thalamic, and dopaminergic input [33,34,59]. In particular, the SNpc DA neurons that project to the auditory striatum appear to reside in the lateral most part of the SNpc (denoted as the substania nigra pars lateralis, SNL), and receive a different set of inputs in comparison to other SNpc/VTA DA neurons. In addition, these neurons appear to respond to novel, intense, and aversive stimuli [60,61]. Menegas et al. demonstrated that these neurons do not appear to signal a traditional RPE that has been found in the SNpc and VTA neurons that presumably project to the more canonical DS and VS. DA antagonism and OHDA depletion of DA in the auditory striatum reduces an animal’s ability to learn about threatening stimuli. In this study, they used air puffs and bitter taste to signal aversive stimuli. In comparison with DA signals in other striatal regions, auditory striatal DA release seems to be the most distinct. Future studies will be important for definitively demarcating this region and determining whether it is sufficient to denote this overall region as the “tail striatum” or whether it is based on specific sensory inputs. Furthermore, it will be important to investigate whether the auditory striatum’s DA input will be important for other types of sensory learning and decision making.
2.3. VTA and the Ventral Striatum
Ventral Striatum—Motivation and learning: Having discussed the differences in the lateral and medial subdivision of the dorsal striatum, it is important to consider the foremost subdivision of the striatum across the dorsal-ventral axis. In comparison with the DMS, the VS or nucleus accumbens is thought to be more related to emotional, social, and reward processing. Anatomically the ventral striatum has been demarcated as consisting of the ventral and medial parts of the dorsal striatum, the nucleus accumbens, as well as the olfactory tubercle system. Here, we primarily consider the nucleus accumbens, which is subdivided into core and shell components, on the basis of immunocytochemical architecture, as well as input–output connectivity [11]. Excitotoxic lesions of the nucleus accumbens core impact the learning of task acquisitions [62]. Akin to lesions in the DMS previously discussed, the core lesions’ effect on stimulus–response learning mainly impacts early learning and had little effect if rats had already learned the task. In comparison to the core’s role in reinforcement learning, the shell appears to play more of a role in motivated behavior and the locomotion of conditioned response expression [63]. Overall, the nucleus accumbens core and shell appear to play dual roles in motivation and learning. This divide, while not complete, appears to be analogous to the DMS and DLS divide previously discussed.
D1 vs. D2: Both the core and shell of the nucleus accumbens are comprised of D1R and D2R MSNs. However, in comparison to the dorsal striatum, these pathways project to different but analogous regions. In canonical contexts, the D1R MSNs project to the VTA while the D2R MSNs project to the ventral pallidum. Paralleling the neuroanatomical dichotomy, studies suggest that the D1R pathway encodes positive valence while the D2R pathway encodes negative valence [28]. However, a major study by Kupchik and colleagues demonstrated that the D1R and D2R pathways are not label-lined, and thus genetic or pharmacological interventions of the respective receptors should be interpreted with caution [64]. Another complicating factor is that there are populations of MSNs that co-express D1R and D2R [65]. Thus, as with the DLS and DMS, the evidence for a D1R/D2R functional and behavioral divide is apparent but complex. In comparison to the DLS/DMS, where influential models of Parkinson’s disease support a D1R/D2R opponency, the divide is less clarified in the VS.
DA in Ventral Striatum: In comparison with the DMS and DLS, DA release in the VS appears to correspond more directly with reward processing and hedonic value. However, this divide is not clear cut and it is important to consider recent studies. The VTA has major projections to the frontal cortex and the ventral striatum (VS) in the respective mesocorticolimbic and mesolimbic systems. Traditionally, these associations have led to the canon conclusion that, in comparison to the SNpc’s role in movement and ongoing behavioral modulation, the VTA is more involved in emotion, learning, and motivational processing. It is important to note here that this divide is neither distinct nor absolute [5,23,24,25]. Advancing the SNpc’s role in movement, recent and previous studies have shown that nigrostriatal activity can correspond to transiently defined movements [23,66,67,68]. However, supporting a role for the VTA system in movement, early studies have shown that DA depletion or antagonism of the VS results in hypoactivity in rodents. Conversely, DA agonism in the VS causes an increase in spontaneous movements. Consistent with this, recent studies have reinstated the importance of movement and kinematic modulation in VTA moment-to-moment responses [3,69,70]. Hughes et al. recently discovered quantitative and causal relationships between VTA activity and movement impulse vectors. Importantly, they showed that VTA DA neuronal activity correspond with different types of movement, regardless of reward or aversive contexts. While these more recent experiments consider the dissociation or confounding of movement with general motivation, it is still theoretically difficult to separate these variables, especially in the context of learning. Thus, further studies will need to identify how the VTA modulates movement variables and how this mode of DA release contributes to motivated behaviors. Additionally, it will be important to consider how DA instantaneous modulates downstream cellular targets in order to enforce movement changes.
From both anterograde and retrograde tracing studies, it has been identified that the lateral portion of the VTA generally projects to the core of the nucleus accumbens, whereas the medial portion projects to the shell [47,69]. This has been found to be significant considering the large body of literature that distinguishes these two accumbal regions in terms of their capacity to process reward, aversive stimuli, and motivation. It appears that VTA DA neurons projecting to the medial nucleus accumbens are involved in aversive fear conditioning, as controlled by the lateral hypothalamus.
Beier et al. systematically characterized the input–output relationships of the VTA. They found that the VTA population projecting to the medial nucleus accumbens is a distinct population of DA neurons that is largely non-overlapping with the VTA population projecting to the lateral nucleus accumbens. Like other studies, they also found that these populations had different roles in motivating reward behavior. A recent study found that the lateral VTA projections to the NAcc core is specifically involved in generating ongoing motivational behavior [71]. They found that, in general, the VTA DA population encodes reward prediction error, the signal attributed to reinforcement learning. Conceptually, however, such RPE learning signals would not be attributed to a role in enforcing ongoing behavior, as such behaviors depend on already-learned contingencies. Thus, the researchers hypothesized that separate neuronal mechanisms must be involved in order to dissociate signals for learning and motivation. Moreover, they hypothesized that both could be attributed to the mesolimbic pathway. They found that DA levels in the NAcc specifically fluctuated with ongoing states of motivation, whereas the levels in other regions of the NAcc only appeared to change with RPE signaling. Thus, this study points towards the differences in function of different VTA DA projections.
2.4. VTA and the Hippocampus
Thus far, we have discussed the striatum and its function as a major DA afferent recipient. While these projections represent major circuit pathways through which DA modulates behavioral output, DA nuclei also provide input to major cortical and hippocampal regions. Here, we limit our discussion to the role of DA in hippocampal circuits, as subsequent sections will highlight a role for this modulation in health and disease. Early evidence utilizing electrolytic lesions of dopaminergic nuclei indicates that the hippocampal formation receives input from both the VTA and SNpc [72]. In addition, circuit plasticity of the hippocampal formation was seen to be modulated by DA pharmacological agents [73]. In a variety of studies, D1R receptors have been found to be expressed in the molecular layer of the dentate gyrus, while D2R receptors have been found to be expressed in the hilus. Thus, at the anatomical and histochemical level, evidence points towards a role for DA in hippocampal function.
Influential theories about how the VTA and the hippocampus are involved in a novelty–reward loop plays into how memories are formed. Briefly, hippocampal connections to the VTA trigger firing in response to novel information. This increased VTA firing is proposed to increase DA in the hippocampus and therefore support the synaptic plasticity underlying different types of memory formation. The optogenetic activation of VTA DA cells have been shown to trigger CA1 replay activity and improve spatial memory. Consistent with this, VTA DA neuron activation can bidirectionally modulate Schaffer collateral synapses. Furthermore, one group demonstrated that concurrent activation of CA1 and the medial forebrain bundle was sufficient to create place preference and a memory trace [74]. Other work demonstrates that the source of DA for modulation of hippocampal circuits may originate from the locus coeruleus in addition to the VTA [75,76]. Nevertheless, the role of DA as an important modulator for hippocampal memory function points toward a potential role in cognitive function and dysfunction.
2.5. Conclusion to Behavior to Neuroanatomical Circuit-Function
Emerging studies of DA and striatal subregion-dependent function provoke multiple themes. Support arises for fundamental differences between striatal regions including the DMS, the DLS, the VS, and the auditory striatum. The VS and DMS have a greater influence on flexible, goal-directed behaviors, while the DLS is more related to stimulus–action encoding and habitual expression. With caveats, original lesion and DA pharmacological studies have been supported by newer sophisticated neuroanatomical tracing, optogenetics, and behavioral paradigms. While there is support for differences in DA function in each of the subregions, aside from auditory striatal DA, there is a degree of convergence at the physiological level. Thus, the specific function of DA in each region may rely mostly on the integrated processing of corticostriatal, thalamostriatal, and nigrostriatal/mesolimbic inputs. Future studies are needed to clarify these neuromodulatory circuits. A thought-provoking circuit question that arises is how to think about D1R and D2R MSNs in each respective region. We have cited studies that primarily refine or refute the original models in movement and reward learning. One trend that needs to be verified is whether the D1/D2 dichotomies hold more strongly when studying the VS and DMS, whereas breakdown occurs when studying the DLS. It is perhaps the case that the DLS as a sensorimotor region is more strongly influenced by complex ongoing locomotor behaviors that may obscure sensory/learning neural dichotomies during experiments. In conclusion, working models have been and continue to be refined but, to deeply understand DA’s role in behavior, there needs to be further clarification of DA receptor pathways in the different regions of the basal ganglia.
3. DA in Neuropsychiatric Disease
Overview: As we have reviewed, DA plays major behavioral roles in modulating major brain regions of the CNS. Thus, it is tantalizing to hypothesize that such functions go awry in neuropsychiatric diseases. Indeed, there is strong support for this in theoretical accounts of dopaminergic pharmacological therapies. In these next sections, we highlight neuropsychiatric diseases and afflicted dopaminergic pathways. We emphasize that we adopt a biased DA-centric view in this regard, and we refer readers to other sources in order to adopt a holistic view of these disorders. We discuss literature that highlights the molecular and genetic basis for selective vulnerability of DLS-projecting SNpc neurons. We then discuss the circuits involved in addiction, highlighting the VTA-VS pathway in this disease. We also discuss recent evidence that indicates an amyloidogenic vulnerability of the VTA–hippocampal pathway in Alzheimer’s Disease. Lastly, recent evidence has emerged regarding the dopaminergic hypothesis of schizophrenia and how it relates to rodent neural circuits. We summarize these recent findings in Figure 2, where we illustrate the specific dopaminergic circuits implicated in disease. Overall, this serves to describe how particular DA circuits are co-opted in different neuropsychiatric diseases.
3.1. Parkinson’s Disease and Dopamine Depletion
One of the most well-known neurological disorders caused by dopamine dysregulation is Parkinson’s Disease (PD). PD is the second largest cause of neurodegeneration [77], which is marked by dopaminergic neuron loss in SNpc [78]. The degeneration of SNpc dopaminergic neurons is thought to be the likely cause the dysfunction of their projection target—the dorsal striatum [79]. Since the dorsal striatum (DS) is responsible for action selection and movement control [46,80,81,82], dysfunction of the DS will lead to movement deficits, such as rigidity, tremor, and shuffling gaits [27,83]. These movement difficulties are the first-found symptoms [84] and are hallmarks in PD cases, serving as evidence for a clinical PD diagnosis [85].
The mechanisms of SNpc neuronal death in PD are a topic of current investigation and debate. Alpha-synuclein protienopathy and mitochondrial dysfunction are two major mechanisms that have been highly investigated. Given its molecular and clinical precedence, alpha-synuclein pathology has been believed to be one of the most probable causes of PD [86,87]. Alpha-synuclein proteins are expressed by neurons across different brain regions and interact with pre-synaptic dopaminergic machinery [88,89,90], such as the dopamine transporter [91]. In Parkinson’s Disease, α-synuclein are aggregated in the cytoplasm and neurites of SNpc dopaminergic neurons, forming Lewy Bodies and Lewy neurites [92]. These aggregated α-synuclein are toxic to neurons and lead to neuronal cell death molecular cascades. However, early neuronal loss in SNpc has also been shown to precede α-synuclein aggregation. Other than α-synuclein pathology, mitochondrial dysfunction is also believed to be responsible for dopaminergic neuron degeneration. Consistent with this, mutations of mitochondria function-related genes have been found to contribute to familiar PD. However, in sporadic PD, which has a later onset in comparison to genetic causes, it has been difficult to prove whether mitochondrial dysfunction is the original cause of dopaminergic neuron loss or just a result of other pathological events. Whether these two major molecular causes underlie selective vulnerability of SNpc DA neurons has not been investigated.
3.2. SNpc DA Neurons’ Selective Vulnerability in PD
As we discussed, motor dysfunction is the hallmark of this neurodegenerative disorder. There were cognitive disorders (e.g., dementia) reported in many of PD cases but there are also PD patients without cognitive abnormalities [93,94]. This phenotype is consistent with the fact that SNpcs degenerate earlier and more severely than the degeneration of VTA in PD brains. In both human PD cases and animal PD models, neurodegeneration follows this conserved pattern in midbrain dopaminergic nuclei [95,96,97]. Dopaminergic neurons in SNpc degenerate more severely than the VTA and within SNpc, and dopaminergic neurons in ventral SNpc degenerate more severely than the dorsal tier of SNr [98,99,100,101]. Different hypotheses have been brought up to explain this selectivity. Differential expression of transcription factors and differences in bioenergetics or metabolism could mark the different vulnerability of SNpc and VTA DA neurons. We discuss several of the major markers of vulnerability below.
Calbindin: One of the markers for selective invulnerability or neurodegeneration-protective is calbindin. Early in 1990, there were papers reporting that, in PD cases, calbindin-(+) SNpc DA neurons were more spared than calbindin-(−) counterparts [102]. Therefore, calbindin has been considered to be a marker of selective invulnerability. The distribution of calbindin-(+) dopamine neurons is mostly concentrated in the VTA and denser in the dorsal tier of SNpc than in the ventral tier [102,103]. This distribution is thus consistent with the vulnerability pattern of PD. Calbindin-(+) dopamine neurons in the dorsal SNpc mainly projects to the medial part of the caudate-putamen, through the rostro-caudal axis [103]. As described in a previous paper, calbindin-(+) dopaminergic projections to the DMS are more resistant to PD pathology, while DLS dopamine projections will degenerate faster [104]. Moreover, the VS receives more calbindin-(+) dopaminergic input from VTA than DS and, therefore, the dorsal striatum suffers from a more severe dopamine deficiency than the ventral striatum in PD. Different vulnerabilities in subregions of the striatum are consistent with the fact that movement deficits are more common and pervasive than other symptoms in PD.
The mechanism of the protective role of calbindin in dopaminergic system has been thought to be related to calbindin’s Ca2+ buffering capacity. High supraphysiological concentrations of Ca2+ in neurons are generally toxic and such Ca2+ could also trigger the aggregation of α-synuclein [105]. This hypothesis is also supported by research indicating that calbindin-(−) dopaminergic neurons (ventral tier of SNpc) have a prominent T-type calcium current and a higher Ca2+ amplitude. This is in contrast with the DA neurons of the dorsal tier of SNpc, which only exhibit a small T-type current. Additionally, VTA DA neuron spiking does not depend on calcium channels [98,106,107]. In vitro assays also showed that overexpression of calbindin in cultured dopaminergic neurons could attenuate the death induced by MPP+, a toxin mimicking PD. However, there are arguments about the relationship between calbindin and protection in selective vulnerability, as a paper from 2012 notes by reporting only 2% calbindin-(+) dopaminergic neurons in the dorsal SNpc. The authors argued that the small number might not be enough for neuroprotective function [98,108]. Later, in a macaque study, a much higher number of calbindin+/TH+ co-expression in the dSNpc appeared (34.7%) [109]. This might be due to the different species in the experiments, but this suggests that calbindin might still be able to serve as a selective invulnerability or protective marker.
Electrophysiological property and ion channels: Previous research reported that calbindin shows a subpopulation-specific expression in SNpc DA neurons [106,110]. Therefore, researchers considered calbindin as a marker for different SNpc DA populations. Studies have been focused on the differences of electrophysiological properties of calbindin-(+) and calbindin-(-) DA neurons. Calbindin-(-) DA neurons, which project to DLS, have low-threshold Ca2+ depolarization in hyperpolarized conditions, which is absent in calbindin-(+) DA neurons in dorsal SNpc [106]. This low-threshold Ca2+ depolarization is accompanied by a large and prolonged Ca2+ transient in dendrites of calbindin- DA neurons. As we discussed in the previous section, a high level of Ca2+ concentration directly contributes to the vulnerability of ventral SNpc DA neurons. Ca2+ could bind to α-synuclein and mediate the interaction of α-synuclein with pre-synaptic sites [105]. A recent paper suggested that calcium promotes the aggregation of α-synuclein [105]. On the other hand, aggregated α-synuclein could bind to calcium pump and further enhance the cytosolic concentration of Ca2+ [111]. This positive feedback loop explains why ventral SNpc DAs are at higher risk of α-synuclein aggregation and neurodegeneration. Except for α-synuclein aggregation, higher levels of cytosolic Ca2+ also add to the oxidative stress of mitochondria [112,113]. Therefore, ventral SNpc DA neurons have a more vulnerable ER-mitochondrial system, considering the higher levels of cytosol Ca2+ concentration. These special electrophysiological properties and higher “daily” Ca2+ burdens explain why vSNpc DA neurons degenerate faster and more severely in PD cases, and therefore lead to significant deficits in DLS function.
Studies have also tried to understand the different Ca2+ concentrations and different electrophysiological properties of dorsal and ventral SNpc DA neurons through the different expressions and functions of ion channels. Several important ion channels, mostly potassium and calcium channels, have the possibility of being markers of selective vulnerability. Ion channels such as GIRK2 (G-protein-regulated inward-rectifier potassium channel 2), SKs (calcium-activated potassium channels), T-type calcium channels, and L-type calcium channels have been analyzed through immunostaining assays [114,115]. The general idea is that the expression of these ion channels in SNpc subpopulations remains similar, although there are debates regarding some other channels in different research [115,116]. These conflicts are likely due to the affinity or selectivity shortage of immunostaining protocols. Although the expression of channels shows no significant difference, as we mentioned above, the function of T-type calcium channels could serve as a marker of selective vulnerability. Ventral SNpc DA neurons depend more on T-type calcium channels than dorsal SNpc DA neurons [106,107]. Dependence of calcium channels might contribute to the higher calcium concentration in vSNpc DA neurons and therefore contribute to selective vulnerability.
ALDH1A1: Another marker for selective vulnerability is aldehyde dehydrogenase 1 (ALDH1A1). ALDH1A1+ dopamine neurons express mostly in the ventral tier of the substantia nigra, and therefore serve as a marker separating dorsal and ventral SNpcs [110,117]. This group of dopaminergic neurons preferentially project to dorsal and lateral parts of the rostral and intermediate striatum, and less so to the caudal parts of the striatum [110,118,119,120]. Especially in the DLS, ALDH1A1+ dopaminergic neuron projections are enriched in striosomal DLS regions [110,119]. According to a previous paper, this part of the striatum receives massive cortical projections from the primary and secondary motor cortexes [33]. Thus, this is consistent with the fact that, in Parkinson’s disease, ventral SNpcs with more ALDH1A1 positive neurons degenerate more severely and movement is more affected. Further molecular evidence was shown in experiments conducted on ALDH1A1-deletion animals which suffer from motor skill learning impairments [120]. This is consistent with our discussed motor or habit roles for the DLS in comparison to the more cognitively skewed DMS [9,36,51,67,82,121]. However, it is important to consider the non-motor cognitive and reward learning aspects of the DLS that may also underlie some PD symptomatology [21,52,122,123]. Nevertheless, the selective vulnerability of DLS-projecting DA neurons in comparison to the DMS-projecting DA neurons is broadly consistent with the DMS/DLS dichotomy.
DCC: Having discussed physiological and metabolic molecular factors, it is important to note that recent work has highlighted the role of developmental cascades in PD. For example, deleted in colorectal cancer (DCC) is a cancer/developmental gene that has received attention for its role in PD. DCC was first characterized in colorectal cancers in 1990 [124]. DCC was later discovered to be a ligand of Netrin-1 in the nervous system, mediating axon guidance and neurodevelopment [125,126]. Interestingly, in the rodent brain, DCC has been found to have preferential expression in the ventral tier of SNpc DA neurons compared to dorsal SNpc and VTA [98,114]. This finding is supported by colocalization studies of calbindin and DCC in substantia nigra, which revealed that DCC was rarely colocalized with calbindin in SNpc [127]. Related to our above motor circuit hypothesis, DCC is enriched DA neurons that primarily project to the DLS. Thus, DCC’s expression pattern is consistent with the vulnerability selectivity in PD DA neuron degeneration.
Although we know that Netrin-1/DCC is still expressed in adult SNpcs, the mechanism of DCC-mediated selective vulnerability has been elusive until recently. Using loss-of-function rodent experiments, it was found that Netrin-1 deletion leads to α-synuclein aggregation and that unbound DCC could trigger the cell death pathways [128]. In this work, the authors also showed that overexpression of Netrin-1 could be neuroprotective in PD models. These facts indicated that loss of Netrin-1 and artificially unbound DCC lead to vulnerability in ventral SNpc in an α-synuclein-dependent manner. Thus, with the knowledge of DCC expression in the ventral tier of the SNpc, this work serves to provide molecular mechanistic evidence for the circuit-based motor deficits seen in PD.
PD-related genetic expression: As we discussed above, different expression levels of calbindin, ALDH1A1, and DCC in the dorsal and ventral tiers of SNpc DA neurons and VTA either directly or indirectly contribute to selective vulnerability in DA neurons. Besides these major genes, transcriptomic analyses have been performed to investigate the different gene expression profiles among VTA, dSNpc, and vSNpc. It has been long known that neuroprotective genes are more expressed in VTA and metabolic genes are more expressed in SNpc, which indicates a higher metabolic burden in SNpc [98]. Recent studies have started to dive into subpopulations in the SNpc to further establish the gene expression differences in human tissue. Consistent with what has been discussed, relatively more neuroprotective genes are expressed in the VTA and dorsal tier of SNpc, while more PD-related genes are expressed in the ventral tier of SNpc [129]. Taken together, these studies explain the molecular culprits underlying selectively vulnerability of ventral SNpc, although more studies need to be performed in rodent models for evidence. Here, we invoke the hypothesis that the motor biased symptoms, together with cognitive symptoms, are related to this selective degeneration of the SNpc ventral tier, and thus a preferential loss of DLS-projecting SNpc neurons.
3.3. VTA and Disease
As we have seen so far, the VTA is one of the major areas of the brain that embodies dopaminergic neurons, with projections to the ventral striatum and hippocampus. More importantly, the VTA is the origin of the reward pathway where dopamine is released to allow hedonic sensations while also playing a crucial role in memory and learning. Having such important functions in the brain, it is reasonable to assume that there will be diseases related to the dysfunction of VTA DA neurons. Sure enough, addiction, Alzheimer’s disease, eating disorders, depression, learning deficits, schizophrenia, ADHD, and many more have been linked to abnormalities in the VTA and its dopaminergic system [130,131,132]. In this section, however, we will focus on reviewing the recent literature on drug addiction and Alzheimer’s disease in relation to the VTA DA neurons.
3.3.1. Drug Addiction and VTA
Drug addiction has been defined as a chronic and relapsing neuropsychiatric disorder that manifests as compulsive drug-seeking behavior. In addition to the consistent problem with alcohol and cigarette smoking, the opioid epidemic in the United States caused more than 63,000 deaths in 2016 alone [133]. Thus, much time and effort has been attributed to researching the mechanism and circuitry of drug addiction. There has been much advancement in our understanding of drug addiction, providing us with numerous therapeutics to combat it [134]. In this section, we will focus on the VTA-NAc pathway and its function in drug addiction.
The dopamine theory of addiction made major breakthroughs in the late 1900s, with studies showing evidence that brain areas with DA neurons displayed positive reinforcement [135]. Recurrent electrical stimulation in these midbrain regions lead to positive reinforcement and, furthermore, blocking dopamine receptors in these areas prevents the reinforcing effects of psychostimulants. Furthermore, one group suggested that not only stimulants but a broad range of drugs (i.e., opiates, central depressants, cholinergic agonists) can increase extracellular dopamine through different mechanisms [136]. Therefore, dopamine neurons have been the focus of addiction research for the past 60 years. The main source of dopamine is from the DA neurons of the midbrain, including the VTA, which was found to mediate the rewarding effects of several drugs. Animals were able to learn to self-administer drugs into the VTA (nicotine: [137,138], opiates: [139,140,141,142], cocaine: [143,144,145], ethanol: [146,147]), and this effect was diminished with the addition of dopamine antagonists or with lesions to the VTA DA neurons [137,138,148,149]. Moreover, this seems to be specific to the VTA and the regions it projects, as the SNpc has almost no release of dopamine in response to intravenous drug administration or abstinence [150,151]. This is thus broadly consistent with the hypothesis that circuit-specific DA neuromodulation underlies addiction symptomatology.
How do addictive substances impact neural circuits of the VTA? In the early 2000s, an interesting hypothesis was proposed where it was argued that neuronal alterations could occur in the mesolimbic pathway following drug exposure, leading to long-term behavioral changes [152]. This stemmed from observations that DA neurons of the VTA were able to express NMDAR-dependent long-term potentiation (LTP) and long-term depression (LTD) at their excitatory synapses [153]. Subsequent studies were able to show that drugs including cocaine, amphetamine, morphine, ethanol, and nicotine induced LTP on the excitatory synapses of VTA DA neurons [153,154,155,156,157]. This suggested that the synaptic adaptation occurring on the VTA DA neurons played a role in drug experience. However, it was noted that repeated exposure to cocaine did not show further alterations in the synaptic plasticity of VTA DA neurons [158]. Having seen these changes, the next questions to ask were “What mediates this drug-induced LTP in the VTA?” and “Is there a behavioral change due to the drug-induced synaptic plasticity?”. The trafficking of AMPA receptors with specific subunits has been observed to mediate LTP in different regions of the brain [159,160,161,162], but it was not known whether drug-induced LTP followed the same mechanism. With further investigation, it was found that GluR1-containing AMPARs contributed to drug-induced synaptic activity [163] and resulted in behavioral deficits [164,165]. When a GluR1 knockout mouse model was used, the administration of cocaine did not induce the LTP that was observed previously, and the experimentally-induced conditioned place preference was absent [165]. On the other hand, overexpressing this subunit in the VTA resulted in sensitized behavior, a process where repeated administration of stimulants leads to a gradual increase in motor-stimulant response, to morphine [165]. Putting these together, it is clear that exposure to drugs causes alterations at the synapses of VTA DA neurons. However, the lack of additional change with subsequent drug exposure suggests that the VTA plasticity and activity is more involved in the early stages of addiction, contributing to the initial reward stimulus of the drug experience [154,166].
One of the areas that the VTA DA neurons transmits information is the nucleus accumbens (NAc), which is also part of the reward circuit. With the drug-induced synaptic plasticity in DA neurons of the VTA, one could assume that drug exposure would affect the NAc. Indeed, the NAc has also been implicated for addiction, as the depletion of dopamine in the NAc with 6-hydroxydopamine (6-OHDA) led to diminished rewarding effects of cocaine and amphetamine [149,167,168] while increasing the intake of morphine. Moreover, application of amphetamine or DA receptor agonists to the NAc induced conditioned place preference [169,170], which was attenuated by DA receptor antagonists. The NAc is comprised largely of two anatomical parts, the NAc core and the NAc shell. Although there is evidence that both play a role in addiction, the NAc shell has been identified to be more crucial in terms of the rewarding effects of drugs, while the NAc core has been shown to be more involved with drug-seeking behavior. Animals learned to self-administer stimulants or dopamine receptor antagonists into the NAc shell but not the core. Moreover, injections of dopaminergic antagonists into the shell, but not the core, led to decreased rewarding effects induced by drugs [171]. From the pharmacological manipulations, we can see that the dopaminergic system is again mediating these behavioral outcomes.
The differences between the NAc shell and core in addiction may potentially be explained by the different genetic expression of DA neurons in the VTA. One study tried to map the different dopamine subtypes and its connectivity, and it was found that Sox6+ DA neurons of the dorsolateral VTA projected to the NAc core and the lateral shell, while vesicular glutamate transporter 2 (Vglut2+) and aldehyde dehydrogenase 1 family member A1 (ALDH1a1+) DA neurons of the VTA projected to the NAc medial shell and olfactory tubercle (OT) [110]. In addition, cholecystokinin (CCK+) DA neurons were found to project to all regions of the NAc and OT [110]. Interestingly, VGlut2+ DA neurons have been identified to be essential for psychostimulant-mediated behavior activation [172]. ALDH1a1 expression in DA neurons was shown to mediate GABA co-release, and decreased ALDH1a1 expression led to increased alcohol consumption and preference [173]. Although it seems that there are differential projections of VTA DA neuron subtypes, more investigation is needed into what role each DA neuron subtype may have in mediating addiction-related behaviors. Furthermore, it will be informative to link these genetic studies to the input–output relationships seen in the VTA [174], and to determine whether the substance-induced plasticity in DA neurons may have a molecular identity or cause.
The VTA DA neurons are known to project onto GABAergic medium spiny neurons (MSNs), which are the NAc’s major cell type. Interestingly, it has been suggested that there are also alterations in the excitatory synaptic inputs, specifically excitatory cortical afferents, on the MSNs of the NAc shell. Similar to what we saw with the VTA, drug administration leads to altered synaptic strength. Application of cocaine induces long-term depression (LTD) at excitatory synapses found on the GABAergic medium spiny cells, and the absence of this LTD prevented the expression of behavioral sensitization, the process of progressively increasing behavioral response due to repeated administration of stimulants [175,176]. However, in contrast to the VTA, cocaine induced this depression following a period of chronic administration rather than a single dose. This suggests that the NAc is more related to the long-lasting behavioral responses due to previous or repeated drug exposure. To examine the behavioral aspects of this synaptic plasticity in the NAc, GluR1, which facilitates LTP, as mentioned before, was examined. Overexpression of GluR1 in the NAc was found to diminish multiple aspects of drug-associated behavior (drug-seeking behavior and rewarding effect) [177,178], while overexpression of a pore-dead GluR1 led to diminished synaptic strength, amplifying the drug-associated behaviors [179]. Moreover, the role of the NAc shell was emphasized in these studies, as the NAc core was not shown to have alterations in GluR1 subunit AMPA receptors in response to chronic cocaine self-administration [177], again suggesting that the NAc shell has a more prominent role in addiction. In addition, it was recently discovered that the NAc medial shell provides inhibitory control to mesolimbic DA neurons through GABA inhibition [180]. Depression of the excitatory synapses on the GABAergic medial spiny neurons may decrease the activity of this pathway, leading to the overactivity of DA neurons and potentiating the rewarding effects of dopamine. Together, these data suggest that the dopamine system of the VTA and NAc play crucial roles in addiction, with the VTA focused on the early stages of disease development and the NAc responsible for behaviors further along.
Putting all this information together, it is convincing that the VTA-NAc pathway is important and plays a role in understanding drug addiction. Moreover, as the NAc shell receives more input from the medial VTA, one may infer that the medial VTA to NAc shell circuit is more involved in the process of initial drug acquisition. On the other hand, the lateral VTA projecting to the NAc core has been demonstrated to have a role in ongoing motivation [71]. In addition, not only is the VTA compartmentalized by its medial-lateral axis, but it also has differences along its anterior-posterior axis, especially in terms of its projections. Some studies have suggested that the posterior VTA is more involved with opioid drug processing [181]. However, the synaptic changes or behavioral changes depending on the different regions of VTA have not been explored extensively. With more studies elucidating these differences, we will be able to form therapeutics that target certain cell types or regions to combat each individual drug specifically.
3.3.2. Alzheimer’s Disease and VTA
As the average age of our population grows, there is an increasing burden on our healthcare system. Dementia is one of the most prevalent diseases in the aged population, with Alzheimer’s disease (AD) being the primary culprit. AD is a devastating neurological disease characterized by progressive memory loss and brain atrophy. The pathology of this disease has continuously been attributed to amyloid plaques and neurofibrillary tangles, causing massive destruction including neuronal loss and synaptic failure. Therefore, we have constantly been focused on the hippocampus and cortex, which are the areas directly related to the symptoms. However, the extrinsic connections to the hippocampus are commonly ignored when discussing the hippocampus, even though they have been identified to be disrupted in AD [182]. The hippocampus receives sub-cortical inputs from various regions, and specifically receives dopamine from the VTA [183,184,185] and locus coeruleus (LC) [75,76] through tyrosine hydroxylase positive (TH+) fibers. In this section, we will be focusing on the recent literature regarding the VTA-hippocampus-NAc pathway in relation to AD.
Dopamine is known to be crucial for modulating synaptic plasticity and memory encoding in the hippocampus [76,183,184,185,186]. As VTA is one of the major sources of dopamine, one can hypothesize that alterations of the VTA DA neurons may lead to the memory deficits we observe in AD. Sure enough, there have been reports that the DAergic system is altered in AD patients, and it is characterized by decreased levels of dopamine and dopamine receptors [187,188]. Moreover, a recent study provided evidence that there was selective neuronal loss in the VTA causing degeneration of dopamine neurons, even before the formation of amyloid plaques in an AD mouse model [189]. Interestingly, other areas including the LC, the hippocampus, and cortical regions were not affected. To further support these findings, a recent clinical study in humans showed that there was a strong association with VTA size to both hippocampal size and memory index in healthy subjects [190]. In addition, hippocampal volume and memory performance, markers for AD, were found to be associated with the functional connectivity between the VTA and hippocampus. Similar to what we saw in mice, other regions did not share this relationship. The authors who discovered the VTA DA neuronal loss in the AD mouse model also reported that the dopamine outflow to the NAc is decreased [189]. As we have seen, the NAc is also a region that the VTA DA neurons project to, modulating reward and motivation processing. This suggests that some non-cognitive function deficits found in AD, such as behavioral deficits or neuropsychiatric symptoms, may be partially due to alterations of the VTA-NAc pathway. One recent study showed that chemogenetic and optogenetic stimulation in the VTA of a dopamine transporter (DAT)-Cre transgenic mice caused hyperactive locomotion and induced reinforcement, while inhibition or pharmacological interventions (DA receptor antagonist) showed opposite effects [191], supporting this hypothesis.
The hippocampus projects glutamatergic transmissions to the NAc core, thus completing a VTA-hippocampal-NAc-VTA loop [192,193]. The NAc core is known to have functions related to memory, learning [194], and reward behavior [195]. Therefore, it was questioned whether alterations in the DA outflow to the hippocampus can cause changes in the hippocampus-NAc core connection. Indeed, the same AD mouse model above was found to show impaired glutamatergic transmission from the hippocampus to the NAc core (reduced excitability and impaired LTP, which was attributed to the loss of VTA DA neurons [196]). Moreover, the administration of L-DOPA rescued the mouse from all deficits. Putting these together, the loss of VTA DA neurons seems to occur in the preclinical stages of AD, causing deficits in the VTA-hippocampus-NAc core functional pathway, thus causing symptoms of AD.
The studies regarding VTA DA neurons and AD show promising results, especially as a marker or early indicator of AD, as these deficits are found mainly in the preclinical stages of AD. To achieve this goal, outlining the specific anatomical relationships is crucial to understanding this pathway. Only a small portion (6–18%) of the projections from the VTA to the hippocampus are from DA neurons [17,197]. Moreover, these DA neurons predominantly project almost exclusively to the CA1 region of the hippocampus, while the NAc core receives its glutamatergic input from the ventral hippocampus [198,199]. These specific regional relationships suggest a specialized role for these projections. Nevertheless, to understand the full extent of how dysfunction of the DAergic system affects AD, much more investigation of the VTA-hippocampus-NAc pathway is needed.
3.4. Schizophrenia, Perception, and Striatal Dopamine
As discussed, major bodies of evidence point toward both reward and motor accounts of DA’s contributions to behavior. However, in each section, we also discussed how striatal circuitry and the dopaminergic modulation thereof may regulate multiple modalities of sensory perception [38,39,53,55,56,57,58,60,200,201,202,203]. Like the reward and motor deficits seen in Parkinson’s disease and addiction, previous and emerging evidence points towards a role for DA dysregulation in clinical perceptual deficits [204,205]. Chief in this regard, schizophrenia and other psychotic disorders are majorly characterized by perceptual disturbances and hallucinations in multiple sensory modalities. Related to DA, DA antagonism has long served as treatment that curtails hallucinations and other positive symptoms seen in schizophrenia. This pharmacological and other neuroimaging evidence supports the dopaminergic hypothesis of schizophrenia, that excessive dopamine underlies schizophrenia and perceptual symptomatology [206,207,208,209]. In addition, in terms of negative symptomatology, decreased phasic DA responses have been thought to underlie decreased levels of learning, attention, motivation, and memory in schizophrenic patients [210]. Consistent with this, DA agonists, amphetamine, and L-DOPA have been reported to induce hallucinations clinically [211]. In mechanistic terms, the system’s relationship between human perception and hallucinations is a topic of heavy investigation [212,213,214]. Current influential theories of how the brain encodes hallucinations is based upon “priors” or a set of perceptual expectations and predictions that the brain makes about the environment [215]. This draws upon the idea that the brain does not encode percepts passively but actively with the engagement of subcortical and memory circuits. In this framework, subjects that develop hallucinatory tendencies may have unusually strong priors or engage in circuits in a manner that emphasizes prior sensory predictions in the absence of real stimuli [204,215].
How do neural circuits (striatal or not) regulate sensory perception and hallucinatory events? Landmark studies of perception and basic choice discrimination support the hypothesis that sensory pathways indeed play a causal role in active perception [216,217,218]. In many of these early studies, cortical microstimulations and microlesions have demonstrated a causal role for sensory pathways in generating perception. Advances in optogenetics and deep brain recordings have paved the way to mechanistically study perceptual decision making and perceptual reporting in deep brain structures, such as the striatum, of animal models. As mentioned previously, optogenetic manipulation of auditory corticostriatal, thamlamostriatal, and striatal neurons has an impact on mouse auditory discrimination [56,57,201]. Similar approaches have provided evidence for striatal perception in other modalities such as somatosensation and vision [38,39,202,203]. While these experiments do not point towards mechanisms of hallucination per se, they invoke the argument that striatal circuits can causally mediate forms of perception.
In a technically and conceptually innovative study, Schmack et al. modeled hallucination-like perception in humans and mice [219]. In their study, they showed that normal humans and mice can report hallucinatory events in an alternative-forced choice task. Briefly, wild-type mice and humans were trained to report the detection of acoustic stimuli and were presented with a range of volumes, from a non-detectable 0 dB tone to 30 dB. Both subject pools reported a small but significant number of choices that corresponded to hearing a sound when none could actually be detected. This paradigm is consistent with previous research supporting strong prior models and detailing such halluncinaton-like events in the absence of neuropsychiatric disease [215,220]. Their systematic behavioral paradigm was validated by the use of hallucinogenic agents, which induced an increased report of false alarms (i.e., hallucination-like events). Interestingly, they reported that an increase in both ventral and tail (auditory) striatal DA preceded trials that resulted in a false report. Furthermore, they demonstrated that optogenetically increasing DA levels in the tail striatum causally increases false alarm report rates. This remarkable and thought-provoking evidence for the DA hypothesis of schizophrenia triggers further questions about the auditory striatum. Further work detailing the nature of auditory corticostriatal activity, MSN stimulus representation, and a characterization of different sensory modalities for hallucinatory-like events will be informative.
3.5. Dopamine System Genes and Neuropsychiatric Disease
We have, thus far, discussed the role of DA in various diseases from a systems and neural circuit perspective. However, as further evidence, DA-related genes are implicated as molecular modulators or risk factors for neuropsychiatric diseases such as schizophrenia and addiction [221,222,223,224]. Many of these implicated genes are involved in DA recycling and thus DA homeostasis. Genetic polymorphisms of a dopamine transporter (DAT), a primary enzyme responsible for recycling DA, impact treatment resistance susceptibility and working memory function in patients with schizophrenia [222,225,226]. Furthermore, DAT polymorphisms have been associated with behavioral deficits in anxiety and attention [227,228]. Consistent with this, animal models of DAT genetic manipulation or knockout have shown deficits in executive and cortical function [229]. While DAT levels are high and responsible for recycling dynamics within the striatum, in cortical regions, DAT levels are low and, therefore, DA levels are thought to depend more on degradation dynamics. The catechol-O-methyltransferase (COMT) enzyme is responsible for DA dynamics, and polymorphisms of the COMT gene are also associated with treatment response, cortical activity, and cognitive functioning in schizophrenia [230,231]. Related to this, the relatively common COMT Val158Met polymorphism has been associated with differences in behavioral traits such as reward processing and anxiety [232,233]. Together, these genetic studies implicate DA metabolism machinery and thus region-specific DA levels in the expression of behavior in both clinically normal and neuropsychiatric contexts.
4. Overall Conclusions/Summary
Since the discovery of DA as a functional neurotransmitter in the 1960s, its role in behavior and disease in the CNS has been heavily investigated. With the advent of genetic, physiological, and systems neuroscience techniques, the fields have begun to draw major conclusions regarding DA. Firstly, DA strongly modulates a diverse range of neural systems, such as the repertoire of the basal ganglia, the cortex and subcortex, and the hippocampus. Through the vast portfolio of the basal ganglia, many behavioral functions arise, such as locomotion, reward processing, and learning. We show that there is a gradient of function across the striatum, with the VS more related to reward/value learning and the DLS more related to sensorimotor habit execution. DA’s specific role in these regions seems to be one of functional support, but there are still open-ended questions to answer. For instance, the clearly distinct roles for DA transmission in each subregion are not quite clear, with auditory striatal DA appearing to be the most distinct. Nevertheless, different diseases have implicated different DA pathways, further pointing toward subregional roles for DA. In rodent and human models of Parkinson’s, the SNpc-DLS pathway seems to be particularly vulnerable, famously sparing the VTA DA neurons. In addition, the role of the VTA in reward processing becomes maladaptive. Additionally, recent evidence indicates that the VTA-hippocampal pathways may be selectively vulnerable to insults from experimentally introduced amyloid. With these exciting advances, there are still many questions that remain open-ended. In the reviewed studies and in future studies, new circuit level technologies such as the use of DA sensors and in vivo imaging will allow us to dissect the contributions of circuits to disease [234,235]. What behaviors do specific dopamine receptor pathways (D1-D5) support and how are they related to DA loss in disease? How does the D1/D2 canonical striatal pathways differ in function in each striatal subdivision? Can subtype specific dopamine agents be fruitful for different neuropsychiatric diseases? Are there major metabolic or genetic principles that underlie the SNpc’s/VTA’s ability to cope with aging or inflammation? These many questions are bound to be asked and addressed as the field develops further understanding through newly introduced technologies.
Author Contributions
A.P.C., L.C., T.A.K., and Q.X. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Institutes of Health (DC016746 and DC017470 to Q.X., and 1F30DCDC018214 to A.P.C.).
Institutional Review Board Statement
Not Applicable.
Informed Consent Statement
Not Applicable.
Data Availability Statement
Not Applicable.
Conflicts of Interest
The authors declare no competing interests.
References
- Schultz, W. Dopamine reward prediction-error signalling: A two-component response. Nat. Rev. Neurosci. 2016, 17, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Zhai, S.; Tanimura, A.; Graves, S.M.; Shen, W.; Surmeier, D.J. Striatal synapses, circuits, and Parkinson’s disease. Curr. Opin. Neurobiol. 2018, 48, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Engelhard, B.; Finkelstein, J.; Cox, J.; Fleming, W.; Jang, H.J.; Ornelas, S.; Koay, S.A.; Thiberge, S.Y.; Daw, N.D.; Tank, D.W.; et al. Specialized coding of sensory, motor and cognitive variables in VTA dopamine neurons. Nature 2019, 570, 509–513. [Google Scholar] [CrossRef]
- Collins, A.L.; Saunders, B.T. Heterogeneity in striatal dopamine circuits: Form and function in dynamic reward seeking. J. Neurosci. Res. 2020, 98, 1046–1069. [Google Scholar] [CrossRef] [PubMed]
- Lerner, T.N.; Holloway, A.L.; Seiler, J.L. Dopamine, Updated: Reward Prediction Error and Beyond. Curr. Opin. Neurobiol. 2020, 67, 123–130. [Google Scholar] [CrossRef]
- Shen, W.; Flajolet, M.; Greengard, P.; Surmeier, D.J. Dichotomous dopaminergic control of striatal synaptic plasticity. Science 2008, 321, 848–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaulieu, J.M.; Gainetdinov, R.R. The physiology, signaling, and pharmacology of dopamine receptors. Pharmacol. Rev. 2011, 63, 182–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerfen, C.R.; Surmeier, D.J. Modulation of striatal projection systems by dopamine. Annu. Rev. Neurosci. 2011, 34, 441–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, J.G.; Marshall, J.D.; Ahanonu, B.; Wu, Y.W.; Kim, T.H.; Grewe, B.F.; Zhang, Y.; Li, J.Z.; Ding, J.B.; Ehlers, M.D.; et al. Diametric neural ensemble dynamics in parkinsonian and dyskinetic states. Nature 2018, 557, 177–182. [Google Scholar] [CrossRef]
- Tritsch, N.X.; Granger, A.J.; Sabatini, B.L. Mechanisms and functions of GABA co-release. Nat. Rev. Neurosci. 2016, 17, 139–145. [Google Scholar] [CrossRef]
- Morales, M.; Margolis, E.B. Ventral tegmental area: Cellular heterogeneity, connectivity and behaviour. Nat. Rev. Neurosci. 2017, 18, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Bjorklund, A.; Dunnett, S.B. Dopamine neuron systems in the brain: An update. Trends Neurosci. 2007, 30, 194–202. [Google Scholar] [CrossRef]
- Carlsson, A.; Falck, B.; Hillarp, N.A. Cellular localization of brain monoamines. Acta Physiol. Scand. Suppl. 1962, 56, 1–28. [Google Scholar]
- Falck, B.; Torp, A. New evidence for the localization of noradrenalin in the adrenergic nerve terminals. Med. Exp. Int. J. Exp. Med. 1962, 6, 169–172. [Google Scholar] [CrossRef]
- Everitt, B.J.; Hokfelt, T.; Wu, J.Y.; Goldstein, M. Coexistence of tyrosine hydroxylase-like and gamma-aminobutyric acid-like immunoreactivities in neurons of the arcuate nucleus. Neuroendocrinology 1984, 39, 189–191. [Google Scholar] [CrossRef] [PubMed]
- Hokfelt, T.; Smith, C.B.; Norell, G.; Peters, A.; Crane, A.; Goldstein, M.; Brownstein, M.; Sokoloff, L. Attempts to combine 2-deoxyglucose autoradiography and tyrosine hydroxylase immunohistochemistry. Neuroscience 1984, 13, 495–512. [Google Scholar] [CrossRef]
- Swanson, L.W. The projections of the ventral tegmental area and adjacent regions: A combined fluorescent retrograde tracer and immunofluorescence study in the rat. Brain Res. Bull. 1982, 9, 321–353. [Google Scholar] [CrossRef]
- Yetnikoff, L.; Lavezzi, H.N.; Reichard, R.A.; Zahm, D.S. An update on the connections of the ventral mesencephalic dopaminergic complex. Neuroscience 2014, 282, 23–48. [Google Scholar] [CrossRef] [Green Version]
- Haber, S.N. The place of dopamine in the cortico-basal ganglia circuit. Neuroscience 2014, 282, 248–257. [Google Scholar] [CrossRef] [Green Version]
- Haber, S.N.; Fudge, J.L.; McFarland, N.R. Striatonigrostriatal pathways in primates form an ascending spiral from the shell to the dorsolateral striatum. J. Neurosci. 2000, 20, 2369–2382. [Google Scholar] [CrossRef] [PubMed]
- Pennartz, C.M.; Berke, J.D.; Graybiel, A.M.; Ito, R.; Lansink, C.S.; van der Meer, M.; Redish, A.D.; Smith, K.S.; Voorn, P. Corticostriatal Interactions during Learning, Memory Processing, and Decision Making. J. Neurosci. 2009, 29, 12831–12838. [Google Scholar] [CrossRef]
- Schultz, W. Multiple dopamine functions at different time courses. Annu Rev. Neurosci. 2007, 30, 259–288. [Google Scholar] [CrossRef] [Green Version]
- Coddington, L.T.; Dudman, J.T. The timing of action determines reward prediction signals in identified midbrain dopamine neurons. Nat. Neurosci. 2018, 21, 1563–1573. [Google Scholar] [CrossRef]
- Saunders, B.T.; Richard, J.M.; Margolis, E.B.; Janak, P.H. Dopamine neurons create Pavlovian conditioned stimuli with circuit-defined motivational properties. Nat. Neurosci. 2018, 21, 1072–1083. [Google Scholar] [CrossRef]
- Lee, K.; Claar, L.D.; Hachisuka, A.; Bakhurin, K.I.; Nguyen, J.; Trott, J.M.; Gill, J.L.; Masmanidis, S.C. Temporally restricted dopaminergic control of reward-conditioned movements. Nat. Neurosci. 2020, 23, 209–216. [Google Scholar] [CrossRef]
- Martel, J.C.; Gatti McArthur, S. Dopamine Receptor Subtypes, Physiology and Pharmacology: New Ligands and Concepts in Schizophrenia. Front. Pharmacol. 2020, 11, 1003. [Google Scholar] [CrossRef] [PubMed]
- Kravitz, A.V.; Freeze, B.S.; Parker, P.R.; Kay, K.; Thwin, M.T.; Deisseroth, K.; Kreitzer, A.C. Regulation of parkinsonian motor behaviours by optogenetic control of basal ganglia circuitry. Nature 2010, 466, 622–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobo, M.K.; Covington, H.E., 3rd; Chaudhury, D.; Friedman, A.K.; Sun, H.; Damez-Werno, D.; Dietz, D.M.; Zaman, S.; Koo, J.W.; Kennedy, P.J.; et al. Cell type-specific loss of BDNF signaling mimics optogenetic control of cocaine reward. Science 2010, 330, 385–390. [Google Scholar] [CrossRef] [Green Version]
- Clarkson, R.L.; Liptak, A.T.; Gee, S.M.; Sohal, V.S.; Bender, K.J. D3 Receptors Regulate Excitability in a Unique Class of Prefrontal Pyramidal Cells. J. Neurosci. 2017, 37, 5846–5860. [Google Scholar] [CrossRef] [Green Version]
- Crittenden, J.R.; Graybiel, A.M. Basal Ganglia disorders associated with imbalances in the striatal striosome and matrix compartments. Front. Neuroanat. 2011, 5, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redgrave, P.; Rodriguez, M.; Smith, Y.; Rodriguez-Oroz, M.C.; Lehericy, S.; Bergman, H.; Agid, Y.; DeLong, M.R.; Obeso, J.A. Goal-directed and habitual control in the basal ganglia: Implications for Parkinson’s disease. Nat. Rev. Neurosci. 2010, 11, 760–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shepherd, G.M. Corticostriatal connectivity and its role in disease. Nat. Rev. Neurosci. 2013, 14, 278–291. [Google Scholar] [CrossRef] [PubMed]
- Hintiryan, H.; Foster, N.N.; Bowman, I.; Bay, M.; Song, M.Y.; Gou, L.; Yamashita, S.; Bienkowski, M.S.; Zingg, B.; Zhu, M.; et al. The mouse cortico-striatal projectome. Nat. Neurosci. 2016, 19, 1100–1114. [Google Scholar] [CrossRef] [PubMed]
- Hunnicutt, B.J.; Jongbloets, B.C.; Birdsong, W.T.; Gertz, K.J.; Zhong, H.; Mao, T. A comprehensive excitatory input map of the striatum reveals novel functional organization. Elife 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Devan, B.D.; White, N.M. Parallel information processing in the dorsal striatum: Relation to hippocampal function. J. Neurosci. 1999, 19, 2789–2798. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.H.; Knowlton, B.J. The role of the basal ganglia in habit formation. Nat. Rev. Neurosci. 2006, 7, 464–476. [Google Scholar] [CrossRef]
- Kimchi, E.Y.; Laubach, M. Dynamic encoding of action selection by the medial striatum. J. Neurosci. 2009, 29, 3148–3159. [Google Scholar] [CrossRef] [PubMed]
- Reig, R.; Silberberg, G. Multisensory integration in the mouse striatum. Neuron 2014, 83, 1200–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neely, R.M.; Koralek, A.C.; Athalye, V.R.; Costa, R.M.; Carmena, J.M. Volitional Modulation of Primary Visual Cortex Activity Requires the Basal Ganglia. Neuron 2018, 97, 1356–1368.e1354. [Google Scholar] [CrossRef] [Green Version]
- Soares-Cunha, C.; Coimbra, B.; Sousa, N.; Rodrigues, A.J. Reappraising striatal D1- and D2-neurons in reward and aversion. Neurosci. Biobehav. Rev. 2016, 68, 370–386. [Google Scholar] [CrossRef]
- Shan, Q.; Ge, M.; Christie, M.J.; Balleine, B.W. The acquisition of goal-directed actions generates opposing plasticity in direct and indirect pathways in dorsomedial striatum. J. Neurosci. 2014, 34, 9196–9201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tai, L.H.; Lee, A.M.; Benavidez, N.; Bonci, A.; Wilbrecht, L. Transient stimulation of distinct subpopulations of striatal neurons mimics changes in action value. Nat. Neurosci. 2012, 15, 1281–1289. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Wang, W.; Sabatini, B.L. Anatomically segregated basal ganglia pathways allow parallel behavioral modulation. Nat. Neurosci. 2020, 23, 1388–1398. [Google Scholar] [CrossRef] [PubMed]
- Featherstone, R.E.; McDonald, R.J. Dorsal striatum and stimulus-response learning: Lesions of the dorsolateral, but not dorsomedial, striatum impair acquisition of a simple discrimination task. Behav. Brain Res. 2004, 150, 15–23. [Google Scholar] [CrossRef]
- Hidalgo-Balbuena, A.E.; Luma, A.Y.; Pimentel-Farfan, A.K.; Pena-Rangel, T.; Rueda-Orozco, P.E. Sensory representations in the striatum provide a temporal reference for learning and executing motor habits. Nat. Commun. 2019, 10, 4074. [Google Scholar] [CrossRef] [Green Version]
- Cui, G.; Jun, S.B.; Jin, X.; Pham, M.D.; Vogel, S.S.; Lovinger, D.M.; Costa, R.M. Concurrent activation of striatal direct and indirect pathways during action initiation. Nature 2013, 494, 238–242. [Google Scholar] [CrossRef]
- Bergstrom, H.C.; Lipkin, A.M.; Lieberman, A.G.; Pinard, C.R.; Gunduz-Cinar, O.; Brockway, E.T.; Taylor, W.W.; Nonaka, M.; Bukalo, O.; Wills, T.A.; et al. Dorsolateral Striatum Engagement Interferes with Early Discrimination Learning. Cell Rep. 2018, 23, 2264–2272. [Google Scholar] [CrossRef] [Green Version]
- Matamales, M.; McGovern, A.E.; Mi, J.D.; Mazzone, S.B.; Balleine, B.W.; Bertran-Gonzalez, J. Local D2- to D1-neuron transmodulation updates goal-directed learning in the striatum. Science 2020, 367, 549–555. [Google Scholar] [CrossRef]
- O’Hare, J.K.; Ade, K.K.; Sukharnikova, T.; Van Hooser, S.D.; Palmeri, M.L.; Yin, H.H.; Calakos, N. Pathway-Specific Striatal Substrates for Habitual Behavior. Neuron 2016, 89, 472–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faure, A.; Haberland, U.; Conde, F.; El Massioui, N. Lesion to the nigrostriatal dopamine system disrupts stimulus-response habit formation. J. Neurosci. 2005, 25, 2771–2780. [Google Scholar] [CrossRef]
- Yin, H.H.; Mulcare, S.P.; Hilario, M.R.; Clouse, E.; Holloway, T.; Davis, M.I.; Hansson, A.C.; Lovinger, D.M.; Costa, R.M. Dynamic reorganization of striatal circuits during the acquisition and consolidation of a skill. Nat. Neurosci. 2009, 12, 333–341. [Google Scholar] [CrossRef] [Green Version]
- Darvas, M.; Palmiter, R.D. Restriction of dopamine signaling to the dorsolateral striatum is sufficient for many cognitive behaviors. Proc. Natl. Acad. Sci. USA 2009, 106, 14664–14669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ketzef, M.; Spigolon, G.; Johansson, Y.; Bonito-Oliva, A.; Fisone, G.; Silberberg, G. Dopamine Depletion Impairs Bilateral Sensory Processing in the Striatum in a Pathway-Dependent Manner. Neuron 2017, 94, 855–865.e855. [Google Scholar] [CrossRef] [PubMed]
- Gangarossa, G.; Espallergues, J.; Mailly, P.; De Bundel, D.; de Kerchove d’Exaerde, A.; Herve, D.; Girault, J.A.; Valjent, E.; Krieger, P. Spatial distribution of D1R- and D2R-expressing medium-sized spiny neurons differs along the rostro-caudal axis of the mouse dorsal striatum. Front. Neural Circuits 2013, 7, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Q.; Znamenskiy, P.; Zador, A.M. Selective corticostriatal plasticity during acquisition of an auditory discrimination task. Nature 2015, 521, 348–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Wang, X.; Ge, S.; Xiong, Q. Medial geniculate body and primary auditory cortex differentially contribute to striatal sound representations. Nat. Commun. 2019, 10, 418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, L.; Walker, W.I.; Ponvert, N.D.; Penix, P.L.; Jaramillo, S. Stable representation of sounds in the posterior striatum during flexible auditory decisions. Nat. Commun. 2018, 9, 1534. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Wei, J.X.; Zhang, G.W.; Huang, J.J.; Zingg, B.; Wang, X.; Tao, H.W.; Zhang, L.I. Corticostriatal control of defense behavior in mice induced by auditory looming cues. Nat. Commun. 2021, 12, 1040. [Google Scholar] [CrossRef]
- Menegas, W.; Bergan, J.F.; Ogawa, S.K.; Isogai, Y.; Venkataraju, K.U.; Osten, P.; Uchida, N.; Watabe-Uchida, M. Dopamine neurons projecting to the posterior striatum form an anatomically distinct subclass. Elife 2015, 4, e10032. [Google Scholar] [CrossRef] [PubMed]
- Menegas, W.; Babayan, B.M.; Uchida, N.; Watabe-Uchida, M. Opposite initialization to novel cues in dopamine signaling in ventral and posterior striatum in mice. Elife 2017, 6. [Google Scholar] [CrossRef]
- Menegas, W.; Akiti, K.; Amo, R.; Uchida, N.; Watabe-Uchida, M. Dopamine neurons projecting to the posterior striatum reinforce avoidance of threatening stimuli. Nat. Neurosci. 2018, 21, 1421–1430. [Google Scholar] [CrossRef] [PubMed]
- Kelley, A.E.; Smith-Roe, S.L.; Holahan, M.R. Response-reinforcement learning is dependent on N-methyl-D-aspartate receptor activation in the nucleus accumbens core. Proc. Natl. Acad. Sci. USA 1997, 94, 12174–12179. [Google Scholar] [CrossRef] [Green Version]
- Parkinson, J.A.; Olmstead, M.C.; Burns, L.H.; Robbins, T.W.; Everitt, B.J. Dissociation in effects of lesions of the nucleus accumbens core and shell on appetitive pavlovian approach behavior and the potentiation of conditioned reinforcement and locomotor activity by D-amphetamine. J. Neurosci. 1999, 19, 2401–2411. [Google Scholar] [CrossRef] [PubMed]
- Kupchik, Y.M.; Brown, R.M.; Heinsbroek, J.A.; Lobo, M.K.; Schwartz, D.J.; Kalivas, P.W. Coding the direct/indirect pathways by D1 and D2 receptors is not valid for accumbens projections. Nat. Neurosci. 2015, 18, 1230–1232. [Google Scholar] [CrossRef] [Green Version]
- Gagnon, D.; Petryszyn, S.; Sanchez, M.G.; Bories, C.; Beaulieu, J.M.; De Koninck, Y.; Parent, A.; Parent, M. Striatal Neurons Expressing D1 and D2 Receptors are Morphologically Distinct and Differently Affected by Dopamine Denervation in Mice. Sci. Rep. 2017, 7, 41432. [Google Scholar] [CrossRef] [Green Version]
- Howe, M.W.; Dombeck, D.A. Rapid signalling in distinct dopaminergic axons during locomotion and reward. Nature 2016, 535, 505–510. [Google Scholar] [CrossRef] [Green Version]
- Parker, N.F.; Cameron, C.M.; Taliaferro, J.P.; Lee, J.; Choi, J.Y.; Davidson, T.J.; Daw, N.D.; Witten, I.B. Reward and choice encoding in terminals of midbrain dopamine neurons depends on striatal target. Nat. Neurosci. 2016, 19, 845–854. [Google Scholar] [CrossRef]
- da Silva, J.A.; Tecuapetla, F.; Paixao, V.; Costa, R.M. Dopamine neuron activity before action initiation gates and invigorates future movements. Nature 2018, 554, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Hughes, R.N.; Watson, G.D.R.; Petter, E.A.; Kim, N.; Bakhurin, K.I.; Yin, H.H. Precise Coordination of Three-Dimensional Rotational Kinematics by Ventral Tegmental Area GABAergic Neurons. Curr. Biol. 2019, 29, 3244–3255.e3244. [Google Scholar] [CrossRef]
- Hughes, R.N.; Bakhurin, K.I.; Petter, E.A.; Watson, G.D.R.; Kim, N.; Friedman, A.D.; Yin, H.H. Ventral Tegmental Dopamine Neurons Control the Impulse Vector during Motivated Behavior. Curr. Biol. 2020, 30, 2681–2694.e2685. [Google Scholar] [CrossRef]
- Mohebi, A.; Pettibone, J.R.; Hamid, A.A.; Wong, J.T.; Vinson, L.T.; Patriarchi, T.; Tian, L.; Kennedy, R.T.; Berke, J.D. Publisher Correction: Dissociable dopamine dynamics for learning and motivation. Nature 2019, 571, E3. [Google Scholar] [CrossRef]
- Scatton, B.; Simon, H.; Le Moal, M.; Bischoff, S. Origin of dopaminergic innervation of the rat hippocampal formation. Neurosci. Lett. 1980, 18, 125–131. [Google Scholar] [CrossRef]
- Bach, M.E.; Barad, M.; Son, H.; Zhuo, M.; Lu, Y.F.; Shih, R.; Mansuy, I.; Hawkins, R.D.; Kandel, E.R. Age-related defects in spatial memory are correlated with defects in the late phase of hippocampal long-term potentiation in vitro and are attenuated by drugs that enhance the cAMP signaling pathway. Proc. Natl. Acad. Sci. USA 1999, 96, 5280–5285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Lavilleon, G.; Lacroix, M.M.; Rondi-Reig, L.; Benchenane, K. Explicit memory creation during sleep demonstrates a causal role of place cells in navigation. Nat. Neurosci. 2015, 18, 493–495. [Google Scholar] [CrossRef]
- Kempadoo, K.A.; Mosharov, E.V.; Choi, S.J.; Sulzer, D.; Kandel, E.R. Dopamine release from the locus coeruleus to the dorsal hippocampus promotes spatial learning and memory. Proc. Natl. Acad. Sci. USA 2016, 113, 14835–14840. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, T.; Duszkiewicz, A.J.; Sonneborn, A.; Spooner, P.A.; Yamasaki, M.; Watanabe, M.; Smith, C.C.; Fernández, G.; Deisseroth, K.; Greene, R.W.; et al. Locus coeruleus and dopaminergic consolidation of everyday memory. Nature 2016, 537, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Surmeier, D.J. Determinants of dopaminergic neuron loss in Parkinson’s disease. FEBS J. 2018, 285, 3657–3668. [Google Scholar] [CrossRef] [Green Version]
- Poewe, W.; Seppi, K.; Tanner, C.M.; Halliday, G.M.; Brundin, P.; Volkmann, J.; Schrag, A.E.; Lang, A.E. Parkinson disease. Nat. Rev. Dis. Primers 2017, 3, 17013. [Google Scholar] [CrossRef]
- Samii, A.; Nutt, J.G.; Ransom, B.R. Parkinson’s disease. Lancet 2004, 363, 1783–1793. [Google Scholar] [CrossRef] [Green Version]
- Balleine, B.W.; O’Doherty, J.P. Human and rodent homologies in action control: Corticostriatal determinants of goal-directed and habitual action. Neuropsychopharmacology 2010, 35, 48–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grahn, J.A.; Parkinson, J.A.; Owen, A.M. The cognitive functions of the caudate nucleus. Prog. Neurobiol. 2008, 86, 141–155. [Google Scholar] [CrossRef]
- Yttri, E.A.; Dudman, J.T. Opponent and bidirectional control of movement velocity in the basal ganglia. Nature 2016, 533, 402–406. [Google Scholar] [CrossRef]
- Alcacer, C.; Andreoli, L.; Sebastianutto, I.; Jakobsson, J.; Fieblinger, T.; Cenci, M.A. Chemogenetic stimulation of striatal projection neurons modulates responses to Parkinson’s disease therapy. J. Clin. Investig. 2017, 127, 720–734. [Google Scholar] [CrossRef]
- Parkinson, J. An essay on the shaking palsy. 1817. J. Neuropsychiatry Clin. Neurosci. 2002, 14, 223–236. [Google Scholar] [CrossRef] [PubMed]
- Tysnes, O.B.; Storstein, A. Epidemiology of Parkinson’s disease. J. Neural Transm 2017, 124, 901–905. [Google Scholar] [CrossRef]
- Conway, K.A.; Lee, S.J.; Rochet, J.C.; Ding, T.T.; Williamson, R.E.; Lansbury, P.T., Jr. Acceleration of oligomerization, not fibrillization, is a shared property of both alpha-synuclein mutations linked to early-onset Parkinson’s disease: Implications for pathogenesis and therapy. Proc. Natl. Acad. Sci. USA 2000, 97, 571–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iljina, M.; Garcia, G.A.; Horrocks, M.H.; Tosatto, L.; Choi, M.L.; Ganzinger, K.A.; Abramov, A.Y.; Gandhi, S.; Wood, N.W.; Cremades, N.; et al. Kinetic model of the aggregation of alpha-synuclein provides insights into prion-like spreading. Proc. Natl. Acad. Sci. USA 2016, 113, E1206–E1215. [Google Scholar] [CrossRef] [Green Version]
- Burre, J.; Sharma, M.; Sudhof, T.C. Cell Biology and Pathophysiology of alpha-Synuclein. Cold Spring Harb. Perspect. Med. 2018, 8. [Google Scholar] [CrossRef]
- Burre, J.; Sharma, M.; Tsetsenis, T.; Buchman, V.; Etherton, M.R.; Sudhof, T.C. Alpha-synuclein promotes SNARE-complex assembly in vivo and in vitro. Science 2010, 329, 1663–1667. [Google Scholar] [CrossRef] [Green Version]
- Jakes, R.; Spillantini, M.G.; Goedert, M. Identification of two distinct synucleins from human brain. FEBS Lett. 1994, 345, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Longhena, F.; Faustini, G.; Missale, C.; Pizzi, M.; Bellucci, A. Dopamine Transporter/alpha-Synuclein Complexes Are Altered in the Post Mortem Caudate Putamen of Parkinson’s Disease: An In Situ Proximity Ligation Assay Study. Int. J. Mol. Sci. 2018, 19, 1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehra, S.; Sahay, S.; Maji, S.K. alpha-Synuclein misfolding and aggregation: Implications in Parkinson’s disease pathogenesis. Biochim. Biophys. Acta Proteins Proteom. 2019, 1867, 890–908. [Google Scholar] [CrossRef]
- Dujardin, K.; Moonen, A.J.; Behal, H.; Defebvre, L.; Duhamel, A.; Duits, A.A.; Plomhause, L.; Tard, C.; Leentjens, A.F. Cognitive disorders in Parkinson’s disease: Confirmation of a spectrum of severity. Parkinsonism Relat. Disord. 2015, 21, 1299–1305. [Google Scholar] [CrossRef] [Green Version]
- Roheger, M.; Kalbe, E.; Liepelt-Scarfone, I. Progression of Cognitive Decline in Parkinson’s Disease. J. Parkinsons Dis. 2018, 8, 183–193. [Google Scholar] [CrossRef] [Green Version]
- Kanaan, N.M.; Kordower, J.H.; Collier, T.J. Age-related changes in dopamine transporters and accumulation of 3-nitrotyrosine in rhesus monkey midbrain dopamine neurons: Relevance in selective neuronal vulnerability to degeneration. Eur. J. Neurosci. 2008, 27, 3205–3215. [Google Scholar] [CrossRef] [Green Version]
- Liang, C.L.; Nelson, O.; Yazdani, U.; Pasbakhsh, P.; German, D.C. Inverse relationship between the contents of neuromelanin pigment and the vesicular monoamine transporter-2: Human midbrain dopamine neurons. J. Comp. Neurol. 2004, 473, 97–106. [Google Scholar] [CrossRef]
- Reyes, S.; Fu, Y.; Double, K.L.; Cottam, V.; Thompson, L.H.; Kirik, D.; Paxinos, G.; Watson, C.; Cooper, H.M.; Halliday, G.M. Trophic factors differentiate dopamine neurons vulnerable to Parkinson’s disease. Neurobiol. Aging 2013, 34, 873–886. [Google Scholar] [CrossRef] [Green Version]
- Brichta, L.; Greengard, P. Molecular determinants of selective dopaminergic vulnerability in Parkinson’s disease: An update. Front. Neuroanat. 2014, 8, 152. [Google Scholar] [CrossRef] [Green Version]
- Cai, H.; Liu, G.; Sun, L.; Ding, J. Aldehyde Dehydrogenase 1 making molecular inroads into the differential vulnerability of nigrostriatal dopaminergic neuron subtypes in Parkinson’s disease. Transl. Neurodegener. 2014, 3, 27. [Google Scholar] [CrossRef] [Green Version]
- Fearnley, J.M.; Lees, A.J. Ageing and Parkinson’s disease: Substantia nigra regional selectivity. Brain 1991, 114 Pt 5, 2283–2301. [Google Scholar] [CrossRef]
- Liu, G.; Yu, J.; Ding, J.; Xie, C.; Sun, L.; Rudenko, I.; Zheng, W.; Sastry, N.; Luo, J.; Rudow, G.; et al. Aldehyde dehydrogenase 1 defines and protects a nigrostriatal dopaminergic neuron subpopulation. J. Clin. Investig. 2014, 124, 3032–3046. [Google Scholar] [CrossRef]
- Yamada, T.; McGeer, P.L.; Baimbridge, K.G.; McGeer, E.G. Relative sparing in Parkinson’s disease of substantia nigra dopamine neurons containing calbindin-D28K. Brain Res. 1990, 526, 303–307. [Google Scholar] [CrossRef]
- Poulin, J.F.; Zou, J.; Drouin-Ouellet, J.; Kim, K.Y.; Cicchetti, F.; Awatramani, R.B. Defining midbrain dopaminergic neuron diversity by single-cell gene expression profiling. Cell Rep. 2014, 9, 930–943. [Google Scholar] [CrossRef] [Green Version]
- Ricke, K.M.; Pass, T.; Kimoloi, S.; Fahrmann, K.; Jungst, C.; Schauss, A.; Baris, O.R.; Aradjanski, M.; Trifunovic, A.; Faelker, T.M.E.; et al. Mitochondrial Dysfunction Combined with High Calcium Load Leads to Impaired Antioxidant Defense Underlying the Selective Loss of Nigral Dopaminergic Neurons. J. Neurosci. 2020, 40, 1975–1986. [Google Scholar] [CrossRef]
- Lautenschlager, J.; Stephens, A.D.; Fusco, G.; Strohl, F.; Curry, N.; Zacharopoulou, M.; Michel, C.H.; Laine, R.; Nespovitaya, N.; Fantham, M.; et al. C-terminal calcium binding of alpha-synuclein modulates synaptic vesicle interaction. Nat. Commun. 2018, 9, 712. [Google Scholar] [CrossRef] [Green Version]
- Evans, R.C.; Zhu, M.; Khaliq, Z.M. Dopamine Inhibition Differentially Controls Excitability of Substantia Nigra Dopamine Neuron Subpopulations through T-Type Calcium Channels. J. Neurosci. 2017, 37, 3704–3720. [Google Scholar] [CrossRef]
- Gantz, S.C.; Ford, C.P.; Morikawa, H.; Williams, J.T. The Evolving Understanding of Dopamine Neurons in the Substantia Nigra and Ventral Tegmental Area. Annu. Rev. Physiol. 2018, 80, 219–241. [Google Scholar] [CrossRef]
- Fu, Y.; Yuan, Y.; Halliday, G.; Rusznak, Z.; Watson, C.; Paxinos, G. A cytoarchitectonic and chemoarchitectonic analysis of the dopamine cell groups in the substantia nigra, ventral tegmental area, and retrorubral field in the mouse. Brain Struct. Funct. 2012, 217, 591–612. [Google Scholar] [CrossRef] [Green Version]
- Dopeso-Reyes, I.G.; Rico, A.J.; Roda, E.; Sierra, S.; Pignataro, D.; Lanz, M.; Sucunza, D.; Chang-Azancot, L.; Lanciego, J.L. Calbindin content and differential vulnerability of midbrain efferent dopaminergic neurons in macaques. Front. Neuroanat. 2014, 8, 146. [Google Scholar] [CrossRef] [Green Version]
- Poulin, J.F.; Caronia, G.; Hofer, C.; Cui, Q.; Helm, B.; Ramakrishnan, C.; Chan, C.S.; Dombeck, D.A.; Deisseroth, K.; Awatramani, R. Mapping projections of molecularly defined dopamine neuron subtypes using intersectional genetic approaches. Nat. Neurosci. 2018, 21, 1260–1271. [Google Scholar] [CrossRef] [PubMed]
- Betzer, C.; Lassen, L.B.; Olsen, A.; Kofoed, R.H.; Reimer, L.; Gregersen, E.; Zheng, J.; Cali, T.; Gai, W.P.; Chen, T.; et al. Alpha-synuclein aggregates activate calcium pump SERCA leading to calcium dysregulation. EMBO Rep. 2018, 19. [Google Scholar] [CrossRef]
- Rivero-Rios, P.; Gomez-Suaga, P.; Fdez, E.; Hilfiker, S. Upstream deregulation of calcium signaling in Parkinson’s disease. Front. Mol. Neurosci. 2014, 7, 53. [Google Scholar] [CrossRef] [Green Version]
- Ludtmann, M.H.R.; Abramov, A.Y. Mitochondrial calcium imbalance in Parkinson’s disease. Neurosci. Lett. 2018, 663, 86–90. [Google Scholar] [CrossRef]
- Reyes, S.; Fu, Y.; Double, K.; Thompson, L.; Kirik, D.; Paxinos, G.; Halliday, G.M. GIRK2 expression in dopamine neurons of the substantia nigra and ventral tegmental area. J. Comp. Neurol. 2012, 520, 2591–2607. [Google Scholar] [CrossRef] [Green Version]
- Dufour, M.A.; Woodhouse, A.; Goaillard, J.M. Somatodendritic ion channel expression in substantia nigra pars compacta dopaminergic neurons across postnatal development. J. Neurosci. Res. 2014, 92, 981–999. [Google Scholar] [CrossRef] [PubMed]
- Wolfart, J.; Neuhoff, H.; Franz, O.; Roeper, J. Differential expression of the small-conductance, calcium-activated potassium channel SK3 is critical for pacemaker control in dopaminergic midbrain neurons. J. Neurosci. 2001, 21, 3443–3456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCaffery, P.; Drager, U.C. High levels of a retinoic acid-generating dehydrogenase in the meso-telencephalic dopamine system. Proc. Natl. Acad. Sci. USA 1994, 91, 7772–7776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, J.; Yu, J.; Sun, L.; Xie, C.; Chang, L.; Wu, J.; Hawes, S.; Saez-Atienzar, S.; Zheng, W.; Kung, J.; et al. ALDH1A1 regulates postsynaptic mu-opioid receptor expression in dorsal striatal projection neurons and mitigates dyskinesia through transsynaptic retinoic acid signaling. Sci. Rep. 2019, 9, 3602. [Google Scholar] [CrossRef] [PubMed]
- Sgobio, C.; Wu, J.; Zheng, W.; Chen, X.; Pan, J.; Salinas, A.G.; Davis, M.I.; Lovinger, D.M.; Cai, H. Aldehyde dehydrogenase 1-positive nigrostriatal dopaminergic fibers exhibit distinct projection pattern and dopamine release dynamics at mouse dorsal striatum. Sci. Rep. 2017, 7, 5283. [Google Scholar] [CrossRef]
- Wu, J.; Kung, J.; Dong, J.; Chang, L.; Xie, C.; Habib, A.; Hawes, S.; Yang, N.; Chen, V.; Liu, Z.; et al. Distinct Connectivity and Functionality of Aldehyde Dehydrogenase 1a1-Positive Nigrostriatal Dopaminergic Neurons in Motor Learning. Cell Rep. 2019, 28, 1167–1181.e1167. [Google Scholar] [CrossRef] [Green Version]
- Peters, A.J.; Fabre, J.M.J.; Steinmetz, N.A.; Harris, K.D.; Carandini, M. Striatal activity topographically reflects cortical activity. Nature 2021. [Google Scholar] [CrossRef]
- Hilario, M.; Holloway, T.; Jin, X.; Costa, R.M. Different dorsal striatum circuits mediate action discrimination and action generalization. Eur. J. Neurosci. 2012, 35, 1105–1114. [Google Scholar] [CrossRef] [PubMed]
- Lerner, T.N.; Shilyansky, C.; Davidson, T.J.; Evans, K.E.; Beier, K.T.; Zalocusky, K.A.; Crow, A.K.; Malenka, R.C.; Luo, L.; Tomer, R.; et al. Intact-Brain Analyses Reveal Distinct Information Carried by SNc Dopamine Subcircuits. Cell 2015, 162, 635–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fearon, E.R.; Cho, K.R.; Nigro, J.M.; Kern, S.E.; Simons, J.W.; Ruppert, J.M.; Hamilton, S.R.; Preisinger, A.C.; Thomas, G.; Kinzler, K.W.; et al. Identification of a chromosome 18q gene that is altered in colorectal cancers. Science 1990, 247, 49–56. [Google Scholar] [CrossRef]
- Kang, D.S.; Yang, Y.R.; Lee, C.; Park, B.; Park, K.I.; Seo, J.K.; Seo, Y.K.; Cho, H.; Lucio, C.; Suh, P.G. Netrin-1/DCC-mediated PLCgamma1 activation is required for axon guidance and brain structure development. EMBO Rep. 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Marsh, A.P.; Heron, D.; Edwards, T.J.; Quartier, A.; Galea, C.; Nava, C.; Rastetter, A.; Moutard, M.L.; Anderson, V.; Bitoun, P.; et al. Mutations in DCC cause isolated agenesis of the corpus callosum with incomplete penetrance. Nat. Genet. 2017, 49, 511–514. [Google Scholar] [CrossRef] [PubMed]
- Osborne, P.B.; Halliday, G.M.; Cooper, H.M.; Keast, J.R. Localization of immunoreactivity for deleted in colorectal cancer (DCC), the receptor for the guidance factor netrin-1, in ventral tier dopamine projection pathways in adult rodents. Neuroscience 2005, 131, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Jasmin, M.; Ahn, E.H.; Voutilainen, M.H.; Fombonne, J.; Guix, C.; Viljakainen, T.; Kang, S.S.; Yu, L.Y.; Saarma, M.; Mehlen, P.; et al. Netrin-1 and its receptor DCC modulate survival and death of dopamine neurons and Parkinson’s disease features. EMBO J. 2021, 40, e105537. [Google Scholar] [CrossRef]
- Monzon-Sandoval, J.; Poggiolini, I.; Ilmer, T.; Wade-Martins, R.; Webber, C.; Parkkinen, L. Human-Specific Transcriptome of Ventral and Dorsal Midbrain Dopamine Neurons. Ann. Neurol. 2020, 87, 853–868. [Google Scholar] [CrossRef] [Green Version]
- Conio, B.; Martino, M.; Magioncalda, P.; Escelsior, A.; Inglese, M.; Amore, M.; Northoff, G. Opposite effects of dopamine and serotonin on resting-state networks: Review and implications for psychiatric disorders. Mol. Psychiatry 2020, 25, 82–93. [Google Scholar] [CrossRef]
- Hu, H. Reward and Aversion. Annu. Rev. Neurosci. 2016, 39, 297–324. [Google Scholar] [CrossRef] [Green Version]
- Solinas, M.; Belujon, P.; Fernagut, P.O.; Jaber, M.; Thiriet, N. Dopamine and addiction: What have we learned from 40 years of research. J. Neural Transm 2019, 126, 481–516. [Google Scholar] [CrossRef]
- Scholl, L.; Seth, P.; Kariisa, M.; Wilson, N.; Baldwin, G. Drug and Opioid-Involved Overdose Deaths-United States, 2013–2017. Morb. Mortal. Wkly. Rep. 2018, 67, 1419–1427. [Google Scholar] [CrossRef]
- Harrison, T.K.; Kornfeld, H.; Aggarwal, A.K.; Lembke, A. Perioperative Considerations for the Patient with Opioid Use Disorder on Buprenorphine, Methadone, or Naltrexone Maintenance Therapy. Anesthesiol. Clin. 2018, 36, 345–359. [Google Scholar] [CrossRef]
- Olds, J.; Milner, P. Positive reinforcement produced by electrical stimulation of septal area and other regions of rat brain. J. Comp. Physiol. Psychol. 1954, 47, 419–427. [Google Scholar] [CrossRef]
- Di Chiara, G.; Imperato, A. Drugs abused by humans preferentially increase synaptic dopamine concentrations in the mesolimbic system of freely moving rats. Proc. Natl. Acad. Sci. USA 1988, 85, 5274–5278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- David, V.; Besson, M.; Changeux, J.P.; Granon, S.; Cazala, P. Reinforcing effects of nicotine microinjections into the ventral tegmental area of mice: Dependence on cholinergic nicotinic and dopaminergic D1 receptors. Neuropharmacology 2006, 50, 1030–1040. [Google Scholar] [CrossRef] [PubMed]
- Maskos, U.; Molles, B.E.; Pons, S.; Besson, M.; Guiard, B.P.; Guilloux, J.P.; Evrard, A.; Cazala, P.; Cormier, A.; Mameli-Engvall, M.; et al. Nicotine reinforcement and cognition restored by targeted expression of nicotinic receptors. Nature 2005, 436, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Bozarth, M.A.; Wise, R.A. Intracranial self-administration of morphine into the ventral tegmental area in rats. Life Sci. 1981, 28, 551–555. [Google Scholar] [CrossRef]
- Corre, J.; van Zessen, R.; Loureiro, M.; Patriarchi, T.; Tian, L.; Pascoli, V.; Lüscher, C. Dopamine neurons projecting to medial shell of the nucleus accumbens drive heroin reinforcement. Elife 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- David, V.; Cazala, P. A comparative study of self-administration of morphine into the amygdala and the ventral tegmental area in mice. Behav. Brain Res. 1994, 65, 205–211. [Google Scholar] [CrossRef]
- Devine, D.P.; Wise, R.A. Self-administration of morphine, DAMGO, and DPDPE into the ventral tegmental area of rats. J. Neurosci. 1994, 14, 1978–1984. [Google Scholar] [CrossRef] [Green Version]
- Boyson, C.O.; Holly, E.N.; Shimamoto, A.; Albrechet-Souza, L.; Weiner, L.A.; DeBold, J.F.; Miczek, K.A. Social stress and CRF-dopamine interactions in the VTA: Role in long-term escalation of cocaine self-administration. J. Neurosci. 2014, 34, 6659–6667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- David, V.; Segu, L.; Buhot, M.C.; Ichaye, M.; Cazala, P. Rewarding effects elicited by cocaine microinjections into the ventral tegmental area of C57BL/6 mice: Involvement of dopamine D1 and serotonin1B receptors. Psychopharmacology 2004, 174, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Rodd, Z.A.; Bell, R.L.; Kuc, K.A.; Zhang, Y.; Murphy, J.M.; McBride, W.J. Intracranial self-administration of cocaine within the posterior ventral tegmental area of Wistar rats: Evidence for involvement of serotonin-3 receptors and dopamine neurons. J. Pharmacol. Exp. Ther. 2005, 313, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Rodd, Z.A.; Melendez, R.I.; Bell, R.L.; Kuc, K.A.; Zhang, Y.; Murphy, J.M.; McBride, W.J. Intracranial self-administration of ethanol within the ventral tegmental area of male Wistar rats: Evidence for involvement of dopamine neurons. J. Neurosci. 2004, 24, 1050–1057. [Google Scholar] [CrossRef] [PubMed]
- Rodd-Henricks, Z.A.; McKinzie, D.L.; Crile, R.S.; Murphy, J.M.; McBride, W.J. Regional heterogeneity for the intracranial self-administration of ethanol within the ventral tegmental area of female Wistar rats. Psychopharmacology 2000, 149, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Corrigall, W.A.; Coen, K.M.; Adamson, K.L. Self-administered nicotine activates the mesolimbic dopamine system through the ventral tegmental area. Brain Res. 1994, 653, 278–284. [Google Scholar] [CrossRef]
- Roberts, D.C.; Koob, G.F. Disruption of cocaine self-administration following 6-hydroxydopamine lesions of the ventral tegmental area in rats. Pharmacol. Biochem. Behav. 1982, 17, 901–904. [Google Scholar] [CrossRef]
- Keath, J.R.; Iacoviello, M.P.; Barrett, L.E.; Mansvelder, H.D.; McGehee, D.S. Differential modulation by nicotine of substantia nigra versus ventral tegmental area dopamine neurons. J. Neurophysiol. 2007, 98, 3388–3396. [Google Scholar] [CrossRef]
- Salin, A.; Lardeux, V.; Solinas, M.; Belujon, P. Protracted abstinence from extended cocaine self-administration is associated with hypodopaminergic activity in the VTA but not in the SNc. Int. J. Neuropsychopharmacol. 2020. [Google Scholar] [CrossRef]
- Bowers, M.S.; Chen, B.T.; Bonci, A. AMPA receptor synaptic plasticity induced by psychostimulants: The past, present, and therapeutic future. Neuron 2010, 67, 11–24. [Google Scholar] [CrossRef] [Green Version]
- Mansvelder, H.D.; McGehee, D.S. Long-term potentiation of excitatory inputs to brain reward areas by nicotine. Neuron 2000, 27, 349–357. [Google Scholar] [CrossRef] [Green Version]
- Gao, M.; Jin, Y.; Yang, K.; Zhang, D.; Lukas, R.J.; Wu, J. Mechanisms involved in systemic nicotine-induced glutamatergic synaptic plasticity on dopamine neurons in the ventral tegmental area. J. Neurosci. 2010, 30, 13814–13825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lüscher, C.; Malenka, R.C. Drug-evoked synaptic plasticity in addiction: From molecular changes to circuit remodeling. Neuron 2011, 69, 650–663. [Google Scholar] [CrossRef] [Green Version]
- Saal, D.; Dong, Y.; Bonci, A.; Malenka, R.C. Drugs of abuse and stress trigger a common synaptic adaptation in dopamine neurons. Neuron 2003, 37, 577–582. [Google Scholar] [CrossRef] [Green Version]
- Ungless, M.A.; Whistler, J.L.; Malenka, R.C.; Bonci, A. Single cocaine exposure in vivo induces long-term potentiation in dopamine neurons. Nature 2001, 411, 583–587. [Google Scholar] [CrossRef]
- Borgland, S.L.; Malenka, R.C.; Bonci, A. Acute and chronic cocaine-induced potentiation of synaptic strength in the ventral tegmental area: Electrophysiological and behavioral correlates in individual rats. J. Neurosci. 2004, 24, 7482–7490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bredt, D.S.; Nicoll, R.A. AMPA receptor trafficking at excitatory synapses. Neuron 2003, 40, 361–379. [Google Scholar] [CrossRef] [Green Version]
- Malenka, R.C.; Bear, M.F. LTP and LTD: An embarrassment of riches. Neuron 2004, 44, 5–21. [Google Scholar] [CrossRef] [Green Version]
- Malinow, R.; Malenka, R.C. AMPA receptor trafficking and synaptic plasticity. Annu. Rev. Neurosci. 2002, 25, 103–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, I.; Huganir, R.L. Regulation of AMPA receptors during synaptic plasticity. Trends Neurosci. 2002, 25, 578–588. [Google Scholar] [CrossRef]
- Bellone, C.; Lüscher, C. Cocaine triggered AMPA receptor redistribution is reversed in vivo by mGluR-dependent long-term depression. Nat. Neurosci. 2006, 9, 636–641. [Google Scholar] [CrossRef]
- Carlezon, W.A., Jr.; Boundy, V.A.; Haile, C.N.; Lane, S.B.; Kalb, R.G.; Neve, R.L.; Nestler, E.J. Sensitization to morphine induced by viral-mediated gene transfer. Science 1997, 277, 812–814. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Saal, D.; Thomas, M.; Faust, R.; Bonci, A.; Robinson, T.; Malenka, R.C. Cocaine-induced potentiation of synaptic strength in dopamine neurons: Behavioral correlates in GluRA(−/−) mice. Proc. Natl. Acad. Sci. USA 2004, 101, 14282–14287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heikkinen, A.E.; Möykkynen, T.P.; Korpi, E.R. Long-lasting modulation of glutamatergic transmission in VTA dopamine neurons after a single dose of benzodiazepine agonists. Neuropsychopharmacology 2009, 34, 290–298. [Google Scholar] [CrossRef]
- Lyness, W.H.; Friedle, N.M.; Moore, K.E. Destruction of dopaminergic nerve terminals in nucleus accumbens: Effect on d-amphetamine self-administration. Pharmacol. Biochem. Behav. 1979, 11, 553–556. [Google Scholar] [CrossRef]
- Pettit, H.O.; Ettenberg, A.; Bloom, F.E.; Koob, G.F. Destruction of dopamine in the nucleus accumbens selectively attenuates cocaine but not heroin self-administration in rats. Psychopharmacology 1984, 84, 167–173. [Google Scholar] [CrossRef]
- Carr, G.D.; White, N.M. Anatomical disassociation of amphetamine’s rewarding and aversive effects: An intracranial microinjection study. Psychopharmacology 1986, 89, 340–346. [Google Scholar] [CrossRef] [PubMed]
- White, N.M.; Packard, M.G.; Hiroi, N. Place conditioning with dopamine D1 and D2 agonists injected peripherally or into nucleus accumbens. Psychopharmacology 1991, 103, 271–276. [Google Scholar] [CrossRef]
- Fenu, S.; Spina, L.; Rivas, E.; Longoni, R.; Di Chiara, G. Morphine-conditioned single-trial place preference: Role of nucleus accumbens shell dopamine receptors in acquisition, but not expression. Psychopharmacology 2006, 187, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Birgner, C.; Nordenankar, K.; Lundblad, M.; Mendez, J.A.; Smith, C.; le Grevès, M.; Galter, D.; Olson, L.; Fredriksson, A.; Trudeau, L.E.; et al. VGLUT2 in dopamine neurons is required for psychostimulant-induced behavioral activation. Proc. Natl. Acad. Sci. USA 2010, 107, 389–394. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.I.; Ganesan, S.; Luo, S.X.; Wu, Y.W.; Park, E.; Huang, E.J.; Chen, L.; Ding, J.B. Aldehyde dehydrogenase 1a1 mediates a GABA synthesis pathway in midbrain dopaminergic neurons. Science 2015, 350, 102–106. [Google Scholar] [CrossRef] [Green Version]
- Beier, K.T.; Steinberg, E.E.; DeLoach, K.E.; Xie, S.; Miyamichi, K.; Schwarz, L.; Gao, X.J.; Kremer, E.J.; Malenka, R.C.; Luo, L. Circuit Architecture of VTA Dopamine Neurons Revealed by Systematic Input-Output Mapping. Cell 2015, 162, 622–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brebner, K.; Wong, T.P.; Liu, L.; Liu, Y.; Campsall, P.; Gray, S.; Phelps, L.; Phillips, A.G.; Wang, Y.T. Nucleus accumbens long-term depression and the expression of behavioral sensitization. Science 2005, 310, 1340–1343. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.J.; Beurrier, C.; Bonci, A.; Malenka, R.C. Long-term depression in the nucleus accumbens: A neural correlate of behavioral sensitization to cocaine. Nat. Neurosci. 2001, 4, 1217–1223. [Google Scholar] [CrossRef]
- Sutton, M.A.; Schmidt, E.F.; Choi, K.H.; Schad, C.A.; Whisler, K.; Simmons, D.; Karanian, D.A.; Monteggia, L.M.; Neve, R.L.; Self, D.W. Extinction-induced upregulation in AMPA receptors reduces cocaine-seeking behaviour. Nature 2003, 421, 70–75. [Google Scholar] [CrossRef]
- Todtenkopf, M.S.; Parsegian, A.; Naydenov, A.; Neve, R.L.; Konradi, C.; Carlezon, W.A., Jr. Brain reward regulated by AMPA receptor subunits in nucleus accumbens shell. J. Neurosci. 2006, 26, 11665–11669. [Google Scholar] [CrossRef]
- Bachtell, R.K.; Choi, K.H.; Simmons, D.L.; Falcon, E.; Monteggia, L.M.; Neve, R.L.; Self, D.W. Role of GluR1 expression in nucleus accumbens neurons in cocaine sensitization and cocaine-seeking behavior. Eur. J. Neurosci. 2008, 27, 2229–2240. [Google Scholar] [CrossRef]
- Yang, H.; de Jong, J.W.; Tak, Y.; Peck, J.; Bateup, H.S.; Lammel, S. Nucleus Accumbens Subnuclei Regulate Motivated Behavior via Direct Inhibition and Disinhibition of VTA Dopamine Subpopulations. Neuron 2018, 97, 434–449.e434. [Google Scholar] [CrossRef]
- Sánchez-Catalán, M.J.; Hipólito, L.; Zornoza, T.; Polache, A.; Granero, L. Motor stimulant effects of ethanol and acetaldehyde injected into the posterior ventral tegmental area of rats: Role of opioid receptors. Psychopharmacology 2009, 204, 641–653. [Google Scholar] [CrossRef]
- Van Hoesen, G.W.; Hyman, B.T. Hippocampal formation: Anatomy and the patterns of pathology in Alzheimer’s disease. Prog. Brain Res. 1990, 83, 445–457. [Google Scholar] [CrossRef]
- Broussard, J.I.; Yang, K.; Levine, A.T.; Tsetsenis, T.; Jenson, D.; Cao, F.; Garcia, I.; Arenkiel, B.R.; Zhou, F.M.; De Biasi, M.; et al. Dopamine Regulates Aversive Contextual Learning and Associated In Vivo Synaptic Plasticity in the Hippocampus. Cell Rep. 2016, 14, 1930–1939. [Google Scholar] [CrossRef] [Green Version]
- McNamara, C.G.; Tejero-Cantero, A.; Trouche, S.; Campo-Urriza, N.; Dupret, D. Dopaminergic neurons promote hippocampal reactivation and spatial memory persistence. Nat. Neurosci. 2014, 17, 1658–1660. [Google Scholar] [CrossRef]
- Rossato, J.I.; Bevilaqua, L.R.; Izquierdo, I.; Medina, J.H.; Cammarota, M. Dopamine controls persistence of long-term memory storage. Science 2009, 325, 1017–1020. [Google Scholar] [CrossRef]
- Yang, K.; Broussard, J.I.; Levine, A.T.; Jenson, D.; Arenkiel, B.R.; Dani, J.A. Dopamine receptor activity participates in hippocampal synaptic plasticity associated with novel object recognition. Eur. J. Neurosci. 2017, 45, 138–146. [Google Scholar] [CrossRef] [Green Version]
- Gibb, W.R.; Mountjoy, C.Q.; Mann, D.M.; Lees, A.J. The substantia nigra and ventral tegmental area in Alzheimer’s disease and Down’s syndrome. J. Neurol Neurosurg. Psychiatry. 1989, 52, 193–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storga, D.; Vrecko, K.; Birkmayer, J.G.; Reibnegger, G. Monoaminergic neurotransmitters, their precursors and metabolites in brains of Alzheimer patients. Neurosci. Lett. 1996, 203, 29–32. [Google Scholar] [CrossRef]
- Nobili, A.; Latagliata, E.C.; Viscomi, M.T.; Cavallucci, V.; Cutuli, D.; Giacovazzo, G.; Krashia, P.; Rizzo, F.R.; Marino, R.; Federici, M.; et al. Dopamine neuronal loss contributes to memory and reward dysfunction in a model of Alzheimer’s disease. Nat. Commun. 2017, 8, 14727. [Google Scholar] [CrossRef] [PubMed]
- De Marco, M.; Venneri, A. Volume and Connectivity of the Ventral Tegmental Area are Linked to Neurocognitive Signatures of Alzheimer’s Disease in Humans. J. Alzheimers Dis. 2018, 63, 167–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, M.Y.; Han, X.; Zhao, T.Y.; Wang, Z.Y.; Lu, G.Y.; Wu, N.; Song, R.; Li, J. Re-examining the role of ventral tegmental area dopaminergic neurons in motor activity and reinforcement by chemogenetic and optogenetic manipulation in mice. Metab. Brain Dis. 2019, 34, 1421–1430. [Google Scholar] [CrossRef]
- Grace, A.A.; Onn, S.P. Morphology and electrophysiological properties of immunocytochemically identified rat dopamine neurons recorded in vitro. J. Neurosci. 1989, 9, 3463–3481. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, P.; Grace, A.A. Synaptic interactions among excitatory afferents to nucleus accumbens neurons: Hippocampal gating of prefrontal cortical input. J. Neurosci. 1995, 15, 3622–3639. [Google Scholar] [CrossRef] [Green Version]
- Smith-Roe, S.L.; Kelley, A.E. Coincident activation of NMDA and dopamine D1 receptors within the nucleus accumbens core is required for appetitive instrumental learning. J. Neurosci. 2000, 20, 7737–7742. [Google Scholar] [CrossRef] [Green Version]
- LeGates, T.A.; Kvarta, M.D.; Tooley, J.R.; Francis, T.C.; Lobo, M.K.; Creed, M.C.; Thompson, S.M. Reward behaviour is regulated by the strength of hippocampus-nucleus accumbens synapses. Nature 2018, 564, 258–262. [Google Scholar] [CrossRef] [PubMed]
- Cordella, A.; Krashia, P.; Nobili, A.; Pignataro, A.; La Barbera, L.; Viscomi, M.T.; Valzania, A.; Keller, F.; Ammassari-Teule, M.; Mercuri, N.B.; et al. Dopamine loss alters the hippocampus-nucleus accumbens synaptic transmission in the Tg2576 mouse model of Alzheimer’s disease. Neurobiol. Dis. 2018, 116, 142–154. [Google Scholar] [CrossRef] [PubMed]
- Gasbarri, A.; Verney, C.; Innocenzi, R.; Campana, E.; Pacitti, C. Mesolimbic dopaminergic neurons innervating the hippocampal formation in the rat: A combined retrograde tracing and immunohistochemical study. Brain Res. 1994, 668, 71–79. [Google Scholar] [CrossRef]
- Bagot, R.C.; Parise, E.M.; Pena, C.J.; Zhang, H.X.; Maze, I.; Chaudhury, D.; Persaud, B.; Cachope, R.; Bolanos-Guzman, C.A.; Cheer, J.F.; et al. Ventral hippocampal afferents to the nucleus accumbens regulate susceptibility to depression. Nat. Commun. 2015, 6, 7062. [Google Scholar] [CrossRef] [PubMed]
- Rosen, Z.B.; Cheung, S.; Siegelbaum, S.A. Midbrain dopamine neurons bidirectionally regulate CA3-CA1 synaptic drive. Nat. Neurosci. 2015, 18, 1763–1771. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Gold, J.I. The basal ganglia’s contributions to perceptual decision making. Neuron 2013, 79, 640–649. [Google Scholar] [CrossRef] [Green Version]
- Znamenskiy, P.; Zador, A.M. Corticostriatal neurons in auditory cortex drive decisions during auditory discrimination. Nature 2013, 497, 482–485. [Google Scholar] [CrossRef]
- Sippy, T.; Lapray, D.; Crochet, S.; Petersen, C.C. Cell-Type-Specific Sensorimotor Processing in Striatal Projection Neurons during Goal-Directed Behavior. Neuron 2015, 88, 298–305. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Rangarajan, K.V.; Gerfen, C.R.; Krauzlis, R.J. Activation of Striatal Neurons Causes a Perceptual Decision Bias during Visual Change Detection in Mice. Neuron 2018, 98, 669. [Google Scholar] [CrossRef]
- Horga, G.; Abi-Dargham, A. An integrative framework for perceptual disturbances in psychosis. Nat. Rev. Neurosci. 2019, 20, 763–778. [Google Scholar] [CrossRef] [PubMed]
- McCarthy-Jones, S.; Longden, E. Auditory verbal hallucinations in schizophrenia and post-traumatic stress disorder: Common phenomenology, common cause, common interventions? Front. Psychol. 2015, 6, 1071. [Google Scholar] [CrossRef] [Green Version]
- Abi-Dargham, A.; Rodenhiser, J.; Printz, D.; Zea-Ponce, Y.; Gil, R.; Kegeles, L.S.; Weiss, R.; Cooper, T.B.; Mann, J.J.; Van Heertum, R.L.; et al. Increased baseline occupancy of D2 receptors by dopamine in schizophrenia. Proc. Natl. Acad. Sci. USA 2000, 97, 8104–8109. [Google Scholar] [CrossRef] [Green Version]
- Brugger, S.P.; Angelescu, I.; Abi-Dargham, A.; Mizrahi, R.; Shahrezaei, V.; Howes, O.D. Heterogeneity of Striatal Dopamine Function in Schizophrenia: Meta-analysis of Variance. Biol. Psychiatry 2020, 87, 215–224. [Google Scholar] [CrossRef]
- McCutcheon, R.A.; Abi-Dargham, A.; Howes, O.D. Schizophrenia, Dopamine and the Striatum: From Biology to Symptoms. Trends Neurosci. 2019, 42, 205–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinstein, J.J.; Chohan, M.O.; Slifstein, M.; Kegeles, L.S.; Moore, H.; Abi-Dargham, A. Pathway-Specific Dopamine Abnormalities in Schizophrenia. Biol. Psychiatry 2017, 81, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Maia, T.V.; Frank, M.J. An Integrative Perspective on the Role of Dopamine in Schizophrenia. Biol. Psychiatry 2017, 81, 52–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liechti, M. Novel psychoactive substances (designer drugs): Overview and pharmacology of modulators of monoamine signaling. Swiss Med. Wkly. 2015, 145, w14043. [Google Scholar] [CrossRef]
- Vercammen, A.; de Haan, E.H.; Aleman, A. Hearing a voice in the noise: Auditory hallucinations and speech perception. Psychol. Med. 2008, 38, 1177–1184. [Google Scholar] [CrossRef]
- Bentall, R.P.; Slade, P.D. Reality testing and auditory hallucinations: A signal detection analysis. Br. J. Clin. Psychol. 1985, 24 Pt 3, 159–169. [Google Scholar] [CrossRef]
- Cassidy, C.M.; Balsam, P.D.; Weinstein, J.J.; Rosengard, R.J.; Slifstein, M.; Daw, N.D.; Abi-Dargham, A.; Horga, G. A Perceptual Inference Mechanism for Hallucinations Linked to Striatal Dopamine. Curr. Biol. 2018, 28, 503–514.e504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corlett, P.R.; Horga, G.; Fletcher, P.C.; Alderson-Day, B.; Schmack, K.; Powers, A.R., 3rd. Hallucinations and Strong Priors. Trends Cogn. Sci. 2019, 23, 114–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salzman, C.D.; Britten, K.H.; Newsome, W.T. Cortical microstimulation influences perceptual judgements of motion direction. Nature 1990, 346, 174–177. [Google Scholar] [CrossRef]
- Murphey, D.K.; Maunsell, J.H.; Beauchamp, M.S.; Yoshor, D. Perceiving electrical stimulation of identified human visual areas. Proc. Natl. Acad. Sci. USA 2009, 106, 5389–5393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dehaene, S.; Changeux, J.P. Experimental and theoretical approaches to conscious processing. Neuron 2011, 70, 200–227. [Google Scholar] [CrossRef] [Green Version]
- Schmack, K.; Bosc, M.; Ott, T.; Sturgill, J.F.; Kepecs, A. Striatal dopamine mediates hallucination-like perception in mice. Science 2021, 372. [Google Scholar] [CrossRef]
- Powers, A.R.; Mathys, C.; Corlett, P.R. Pavlovian conditioning-induced hallucinations result from overweighting of perceptual priors. Science 2017, 357, 596–600. [Google Scholar] [CrossRef] [Green Version]
- Birnbaum, R.; Weinberger, D.R. Genetic insights into the neurodevelopmental origins of schizophrenia. Nat. Rev. Neurosci. 2017, 18, 727–740. [Google Scholar] [CrossRef]
- Ye, J.; Ji, F.; Jiang, D.; Lin, X.; Chen, G.; Zhang, W.; Shan, P.; Zhang, L.; Zhuo, C. Polymorphisms in Dopaminergic Genes in Schizophrenia and Their Implications in Motor Deficits and Antipsychotic Treatment. Front. Neurosci. 2019, 13, 355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Leon, S.; Gonzalez-Giraldo, Y.; Wegman-Ostrosky, T.; Forero, D.A. Molecular genetics of substance use disorders: An umbrella review. Neurosci. Biobehav. Rev. 2021, 124, 358–369. [Google Scholar] [CrossRef]
- Popescu, A.; Marian, M.; Dragoi, A.M.; Costea, R.V. Understanding the genetics and neurobiological pathways behind addiction (Review). Exp. Ther. Med. 2021, 21, 544. [Google Scholar] [CrossRef] [PubMed]
- Zivkovic, M.; Mihaljevic-Peles, A.; Bozina, N.; Sagud, M.; Nikolac-Perkovic, M.; Vuksan-Cusa, B.; Muck-Seler, D. The association study of polymorphisms in DAT, DRD2, and COMT genes and acute extrapyramidal adverse effects in male schizophrenic patients treated with haloperidol. J. Clin. Psychopharmacol. 2013, 33, 593–599. [Google Scholar] [CrossRef]
- Bilic, P.; Jukic, V.; Vilibic, M.; Savic, A.; Bozina, N. Treatment-resistant schizophrenia and DAT and SERT polymorphisms. Gene 2014, 543, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Chmielowiec, J.; Chmielowiec, K.; Suchanecka, A.; Trybek, G.; Mroczek, B.; Malecka, I.; Grzywacz, A. Associations Between the Dopamine D4 Receptor and DAT1 Dopamine Transporter Genes Polymorphisms and Personality Traits in Addicted Patients. Int. J. Environ. Res. Public Health 2018, 15, 2076. [Google Scholar] [CrossRef] [Green Version]
- Kuc, K.; Bielecki, M.; Racicka-Pawlukiewicz, E.; Czerwinski, M.B.; Cybulska-Klosowicz, A. The SLC6A3 gene polymorphism is related to the development of attentional functions but not to ADHD. Sci. Rep. 2020, 10, 6176. [Google Scholar] [CrossRef] [Green Version]
- Illiano, P.; Leo, D.; Gainetdinov, R.R.; Pardo, M. Early Adolescence Prefrontal Cortex Alterations in Female Rats Lacking Dopamine Transporter. Biomedicines 2021, 9, 157. [Google Scholar] [CrossRef]
- Kaneko, H.; Miura, I.; Kanno-Nozaki, K.; Horikoshi, S.; Hino, M.; Yabe, H. COMT Val 108/158 Met polymorphism and treatment response to aripiprazole in patients with acute schizophrenia. Neuropsychiatr. Dis. Treat. 2018, 14, 1657–1663. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Garcia, P.; Cristobal-Huerta, A.; Espinoza, L.Y.; Molero, P.; Sanchez-Pedreno, F.O.; Hernandez-Tamames, J.A. The influence of the COMT genotype in the underlying functional brain activity of context processing in schizophrenia and in relatives. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2016, 71, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Julio-Costa, A.; Martins, A.A.S.; Wood, G.; de Almeida, M.P.; de Miranda, M.; Haase, V.G.; Carvalho, M.R.S. Heterosis in COMT Val158Met Polymorphism Contributes to Sex-Differences in Children’s Math Anxiety. Front. Psychol. 2019, 10, 1013. [Google Scholar] [CrossRef] [PubMed]
- Sindermann, C.; Luo, R.; Zhang, Y.; Kendrick, K.M.; Becker, B.; Montag, C. The COMT Val158Met Polymorphism and Reaction to a Transgression: Findings of Genetic Associations in Both Chinese and German Samples. Front. Behav. Neurosci. 2018, 12, 148. [Google Scholar] [CrossRef] [PubMed]
- Patriarchi, T.; Mohebi, A.; Sun, J.; Marley, A.; Liang, R.; Dong, C.; Puhger, K.; Mizuno, G.O.; Davis, C.M.; Wiltgen, B.; et al. An expanded palette of dopamine sensors for multiplex imaging in vivo. Nat. Methods 2020, 17, 1147–1155. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Zhou, J.; Dai, B.; Qian, T.; Zeng, J.; Li, X.; Zhuo, Y.; Zhang, Y.; Wang, Y.; Qian, C.; et al. Next-generation GRAB sensors for monitoring dopaminergic activity in vivo. Nat. Methods 2020, 17, 1156–1166. [Google Scholar] [CrossRef]
Figure 1.
The medial–lateral midbrain projection system to the anterior–posterior striatal axis: Different regions of the dorsal striatum receive dopaminergic inputs from distinct but overlapping populations of the SNpc and the VTA. The posterior auditory striatum primarily receives nigrostriatal input from the lateral portions of the SNpc or what has been termed the substantia nigra pars lateralis (SNL). Moving anteriorly, there is relatively more contribution from the medial portions of the SNpc, and then subsequently contributions from the VTA. On the opposite end of the spectrum, the NAcc ventral striatum receives input primarily from the VTA, with lateral portions of the VTA projecting to the NAcc core and medial portions projecting to the shell. Major input across the posterior to anterior dorsal striatum/NAcc gradually shifts from sensory to limbic cortices. Here, we listed the differential inputs and associated behavioral functions of each subregion. Abbreviations: Dopamine, DA; dorsolateral striatum, DLS; dorsomedial striatum, DMS; nucleus accumbens, NAcc; primary motor cortex, M1; anterior cingulate cortex, ACC; orbitofrontal cortex, OFC; secondary motor cortex, M2; dorsal prefrontal cortex, dPFC; ventromedial prefrontal cortex, vmPFC.
Figure 1.
The medial–lateral midbrain projection system to the anterior–posterior striatal axis: Different regions of the dorsal striatum receive dopaminergic inputs from distinct but overlapping populations of the SNpc and the VTA. The posterior auditory striatum primarily receives nigrostriatal input from the lateral portions of the SNpc or what has been termed the substantia nigra pars lateralis (SNL). Moving anteriorly, there is relatively more contribution from the medial portions of the SNpc, and then subsequently contributions from the VTA. On the opposite end of the spectrum, the NAcc ventral striatum receives input primarily from the VTA, with lateral portions of the VTA projecting to the NAcc core and medial portions projecting to the shell. Major input across the posterior to anterior dorsal striatum/NAcc gradually shifts from sensory to limbic cortices. Here, we listed the differential inputs and associated behavioral functions of each subregion. Abbreviations: Dopamine, DA; dorsolateral striatum, DLS; dorsomedial striatum, DMS; nucleus accumbens, NAcc; primary motor cortex, M1; anterior cingulate cortex, ACC; orbitofrontal cortex, OFC; secondary motor cortex, M2; dorsal prefrontal cortex, dPFC; ventromedial prefrontal cortex, vmPFC.

Figure 2.
Summary of reviewed DA-related diseases categorized according to different brain regions. Diseases associated with the SNpc include, but are not limited to, Parkison’s, schizophrenia, and OCD. The DLS is subject to metabolic and molecular vulnerability in Parkinson’s disease. The tail striatum has recently been implicated in hallucinatory-like percepts in schizophrenia models. Diseases associated with the VTA include, but are not limited to, addiction, Alzheimer’s disease, and ADHD. The shell of the nucleus accumbens have specifically been linked to hedonic processing and plasticity in initial drug acquisition. The VTA also projects to the hippocampus and it has been found that these neurons are vulnerable in amyloidogenic mouse models.
Figure 2.
Summary of reviewed DA-related diseases categorized according to different brain regions. Diseases associated with the SNpc include, but are not limited to, Parkison’s, schizophrenia, and OCD. The DLS is subject to metabolic and molecular vulnerability in Parkinson’s disease. The tail striatum has recently been implicated in hallucinatory-like percepts in schizophrenia models. Diseases associated with the VTA include, but are not limited to, addiction, Alzheimer’s disease, and ADHD. The shell of the nucleus accumbens have specifically been linked to hedonic processing and plasticity in initial drug acquisition. The VTA also projects to the hippocampus and it has been found that these neurons are vulnerable in amyloidogenic mouse models.


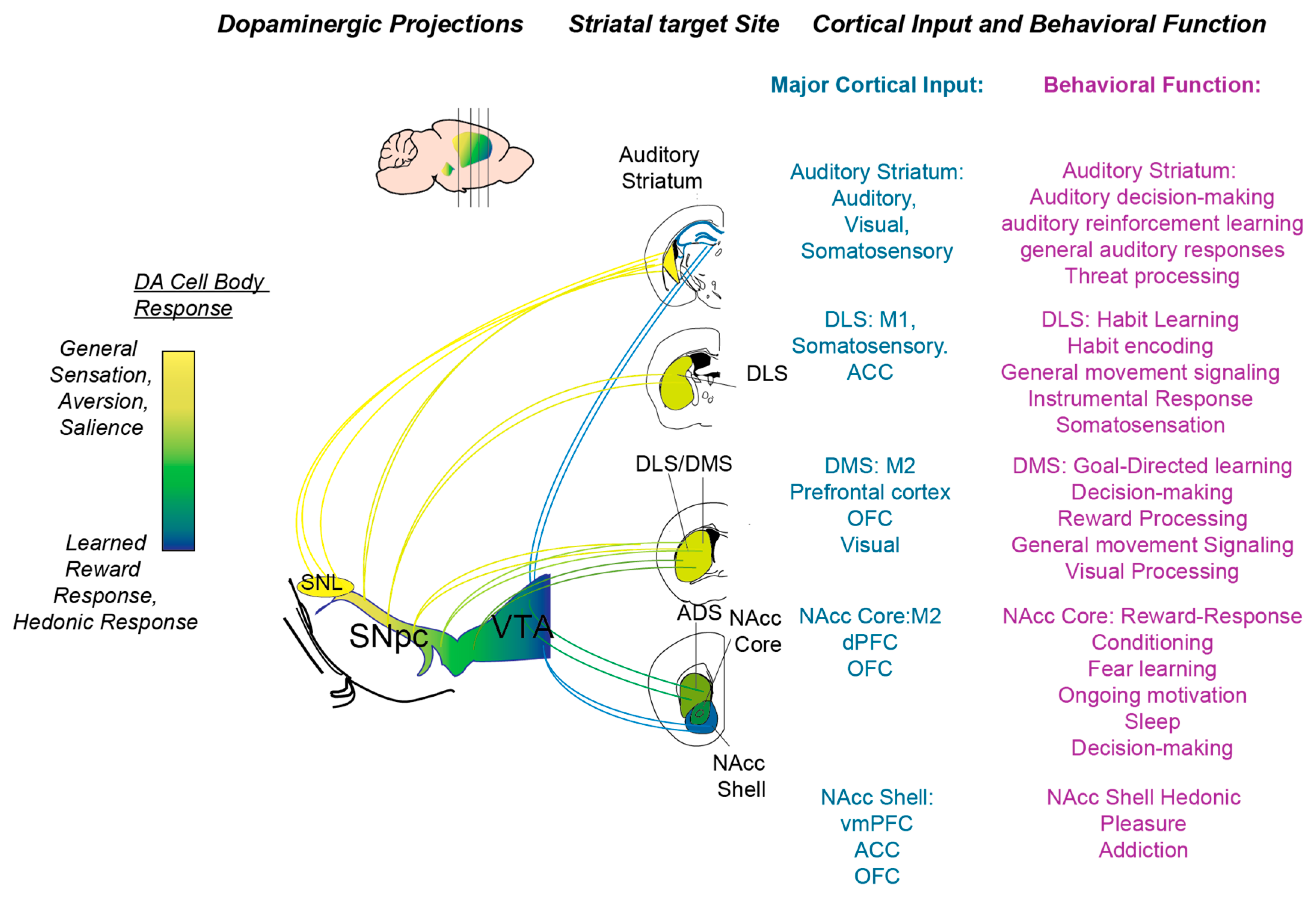{kind=link}
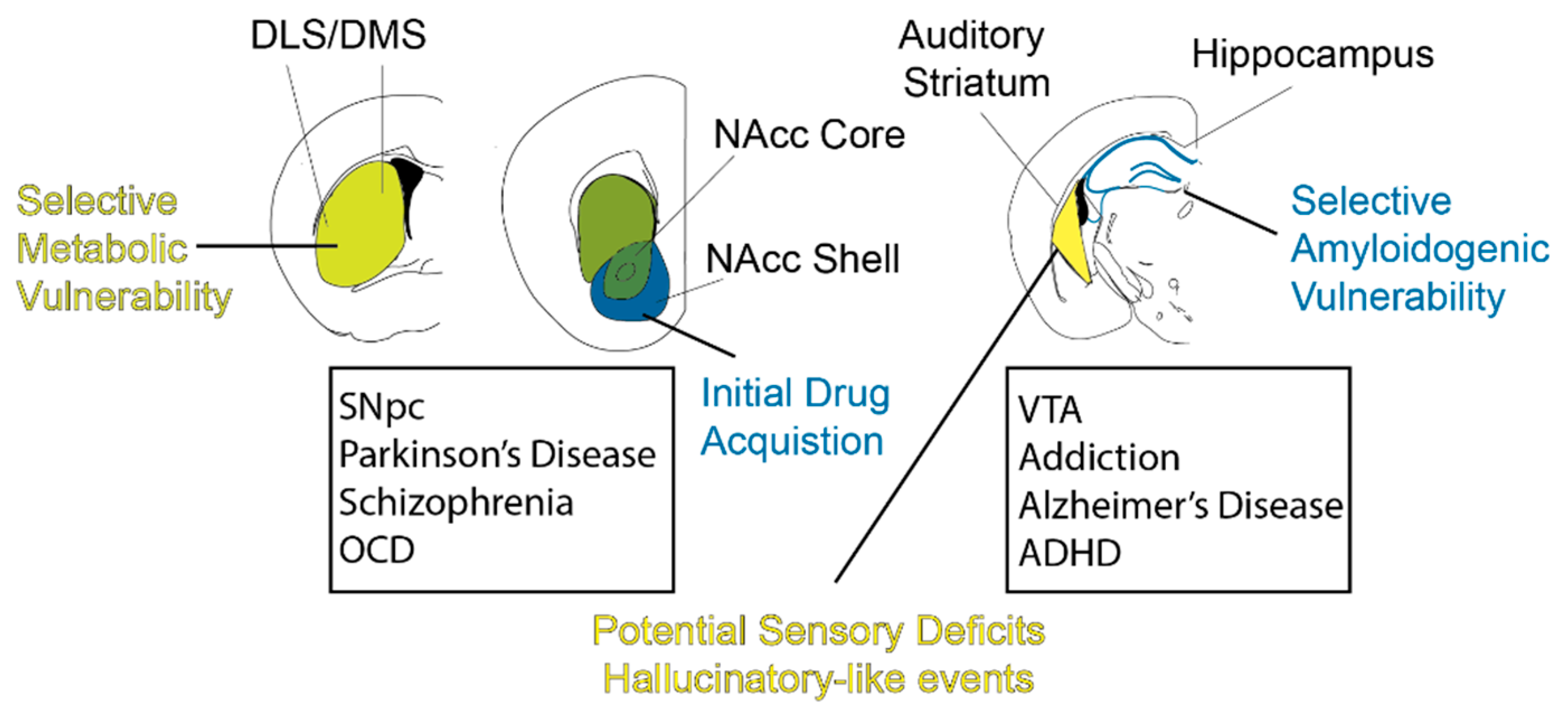{kind=link}
Table 1.
Highlighted summary of details characterizing each striatal subregion.
| Striatal Subregion | Overall Function | D1R/D2R Function | DA Function |
|---|---|---|---|
| DMS | Goal-directed behavior, visual perception, movement | D1R: Positive reinforcement | Reward, aversive processing, movement |
| D2R: Punishment | |||
| DLS | Habit formation, skill refinement, sensorimotor integration, somatosensation, movement | D1R: Habit formation and skill execution | Reward, aversive processing, movement |
| D2R: Habit/skill refinement | |||
| VS | Motivation, learning | Reward and aversion opponency | Reward, aversive processing, motivation, movement |
| Auditory/tail Striatum | Auditory decision making, salience, threat processing, sensory processing | D1R MSNs in driving auditory decisions, D2R MSNs in auditory-induced freezing behavior | Threat learning, sensory processing |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chen, A.P.; Chen, L.; Kim, T.A.; Xiong, Q. Integrating the Roles of Midbrain Dopamine Circuits in Behavior and Neuropsychiatric Disease. Biomedicines 2021, 9, 647. https://doi.org/10.3390/biomedicines9060647
AMA Style
Chen AP, Chen L, Kim TA, Xiong Q. Integrating the Roles of Midbrain Dopamine Circuits in Behavior and Neuropsychiatric Disease. Biomedicines. 2021; 9(6):647. https://doi.org/10.3390/biomedicines9060647
Chicago/Turabian StyleChen, Allen PF, Lu Chen, Thomas A. Kim, and Qiaojie Xiong. 2021. "Integrating the Roles of Midbrain Dopamine Circuits in Behavior and Neuropsychiatric Disease" Biomedicines 9, no. 6: 647. https://doi.org/10.3390/biomedicines9060647
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.