Screening of Bacterial Endophytes Able to Promote Plant Growth and Increase Salinity Tolerance

 ,
,  , ,
, ,  ,
, 
Abstract
:Featured Application
Abstract
1. Introduction
2. Materials and Methods
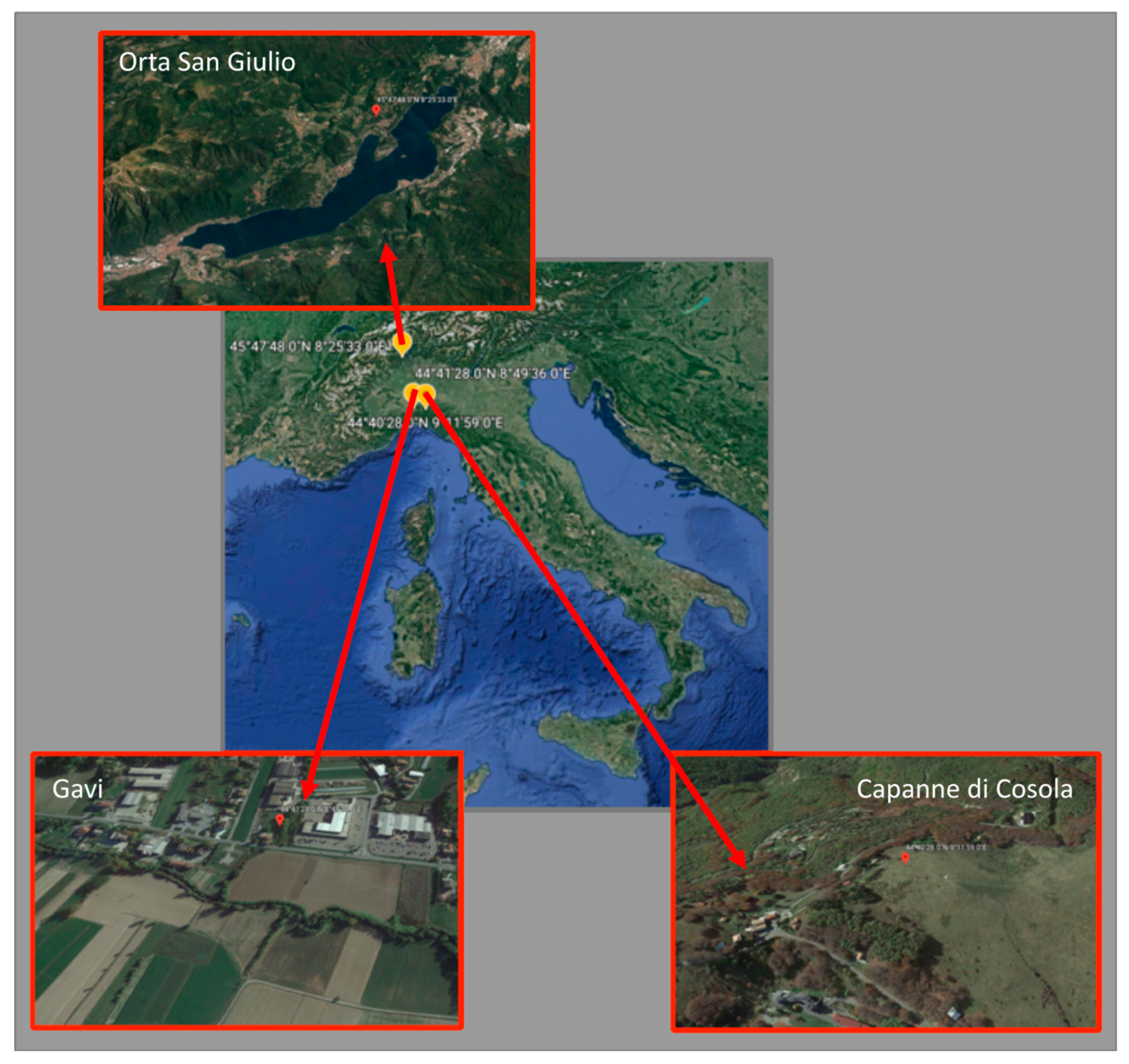
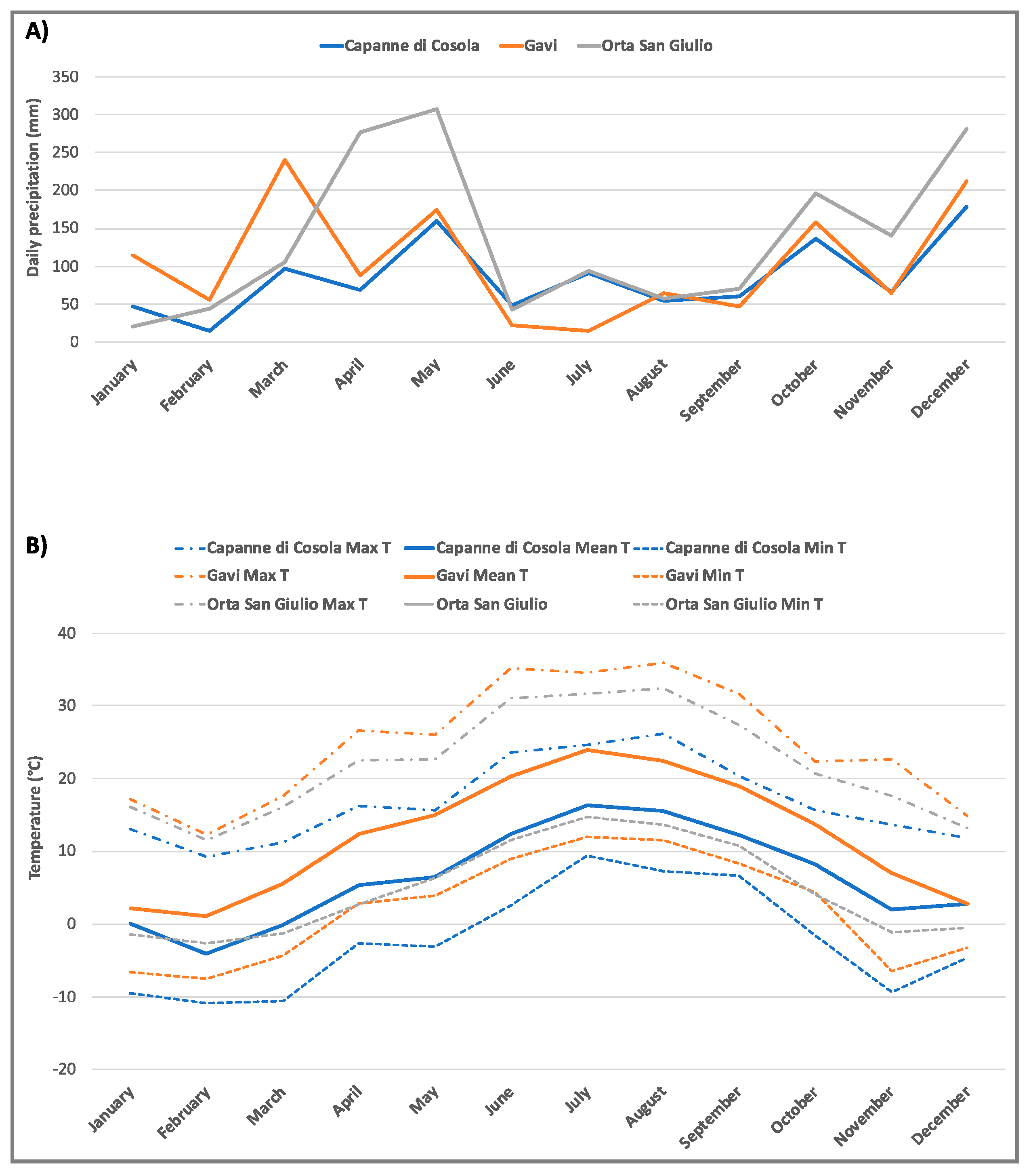
2.1. Sampling Site
2.2. Seed Sterilization
2.3. Isolation of Bacterial Endophytes
2.4. Bacterial Endophyte Identification
2.5. Characterization of Physiological Plant-Beneficial Activities
2.6. Antibiotic Resistance Profile
2.7. Determining Capability to Colonize Plant Tissue and Promote Plant Growth
2.8. Evaluation of ACC Deaminase Activity
2.9. Determination of Minimal Inhibitory Concentration for the Growth of Bacterial Strains in the Presence of Salt Stress
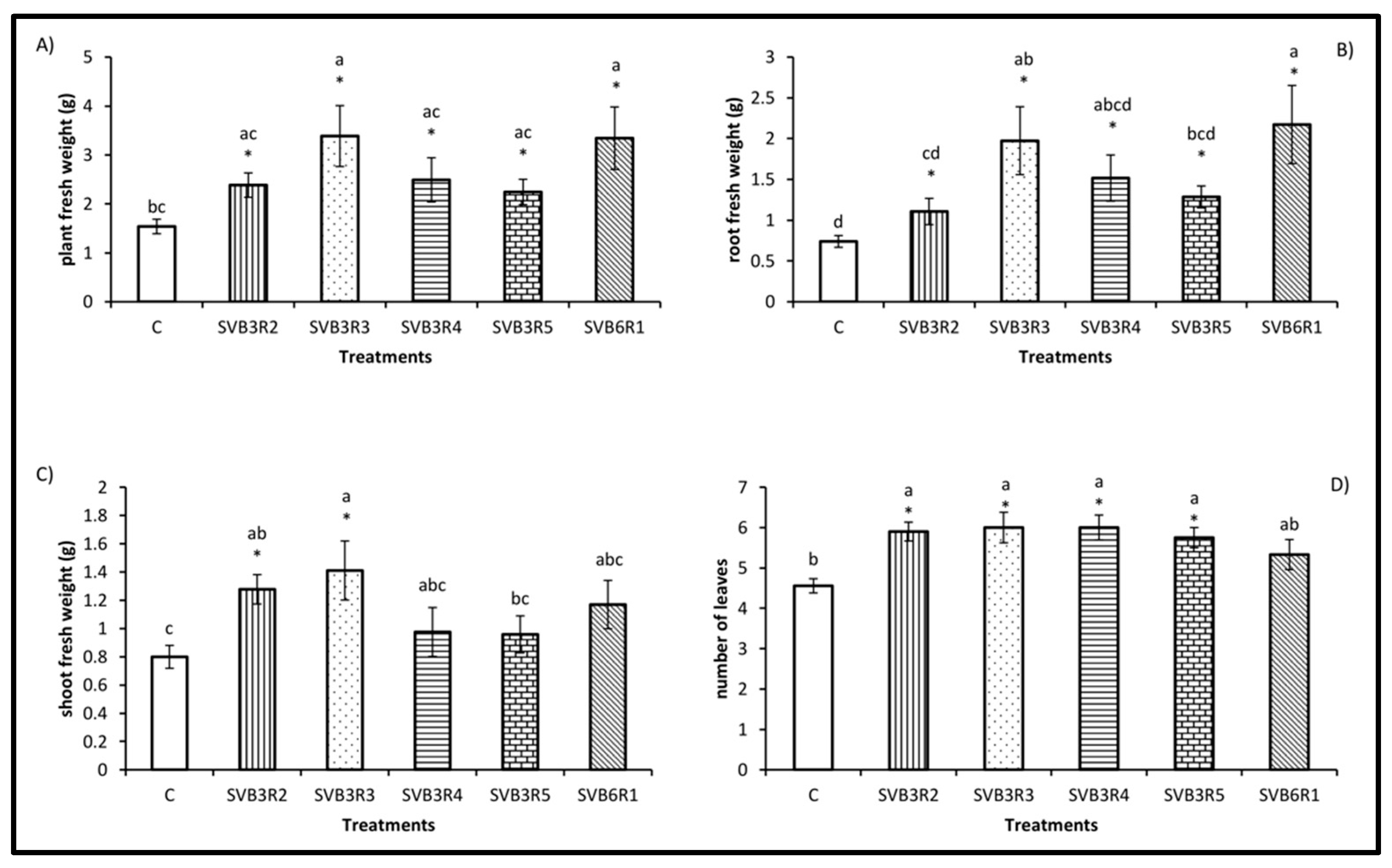
2.10. Effects of Bacterial Endophytes on Plants Exposed to Salt Stress
2.11. Statistical Analysis
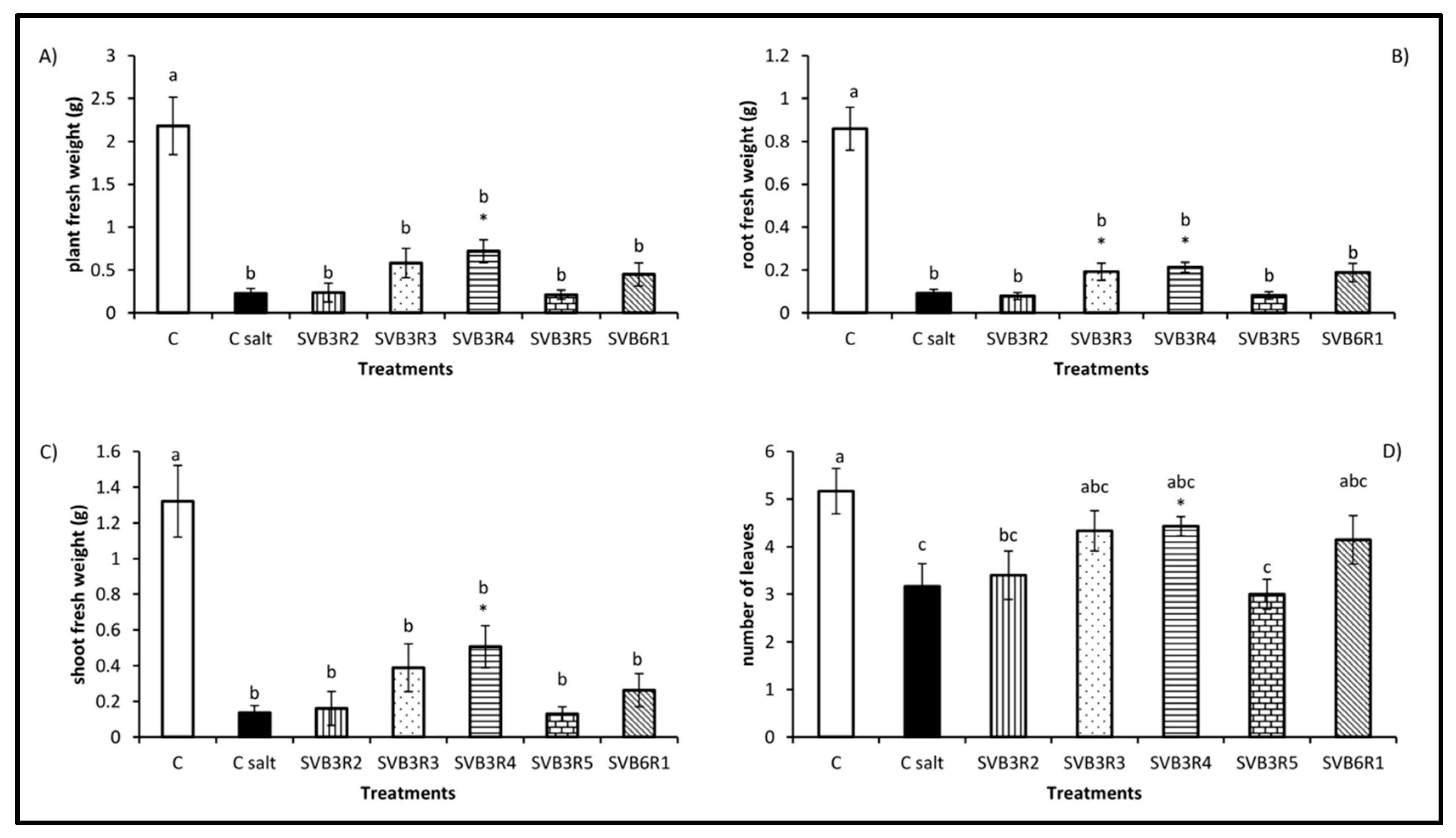
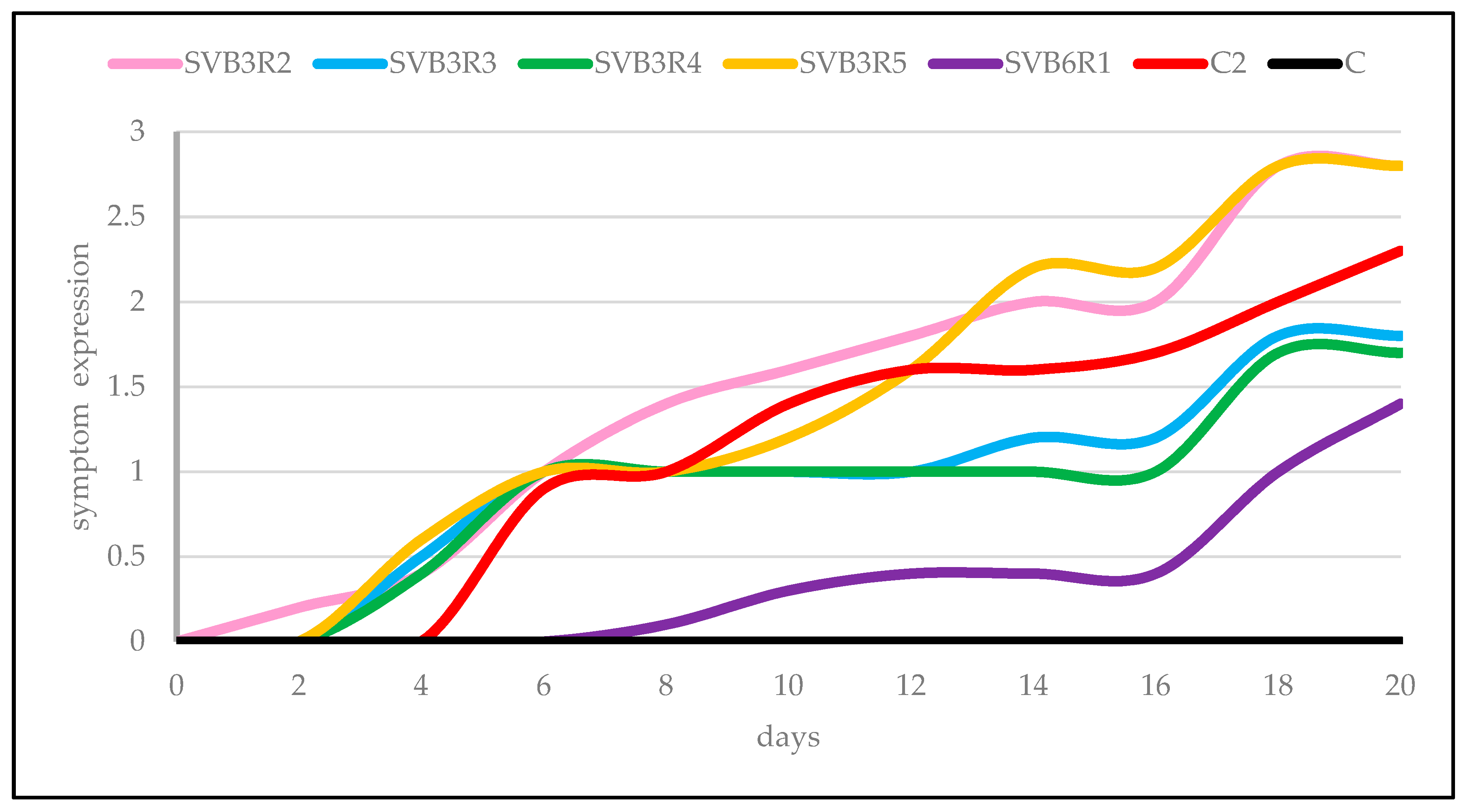
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wilson, D. Endophyte: The evolution of a term, and clarification of its use and definition. Oikos 1995, 73, 274. [Google Scholar] [CrossRef]
- Hallmann, J.; Quadt-Hallmann, A.; Mahaffee, A.; Kloepper, J. Endophytic bacteria in agricultural crops. Can. J. Microbiol. 1997, 43, 895–914. [Google Scholar] [CrossRef]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The hidden world within plants: Ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santoyo, G.; Moreno-Hagelsieb, G.; del Carmen Orozco-Mosqueda, M.; Glick, B.R. Plant growth-promoting bacterial endophytes. Microbiol. Res. 2016, 183, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Duan, J.; Charles, T.C.; Glick, B.R. A bioinformatics approach to the determination of genes involved in endophytic behavior in Burkholderia spp. J. Theor. Biol. 2014, 343, 193–198. [Google Scholar] [CrossRef]
- Ali, S.; Charles, T.C.; Glick, B.R. Delay of flower senescence by bacterial endophytes expressing 1-aminocyclopropane-1-carboxylate deaminase. J. Appl. Microbiol. 2012, 113, 1139–1144. [Google Scholar] [CrossRef] [PubMed]
- Coutinho, B.G.; Licastro, D.; Mendonça-Previato, L.; Cámara, M.; Venturi, V. Plant-Influenced Gene Expression in the Rice Endophyte Burkholderia kururiensis M130. Mol. Plant-Microbe Interact. 2015, 28, 10–21. [Google Scholar] [CrossRef] [Green Version]
- de Abreu, C.S.; Figueiredo, J.E.F.; Oliveira, C.A.; dos Santos, V.L.; Gomes, E.A.; Ribeiro, V.P.; Barros, B.A.; Lana, U.G.P.; Marriel, I.E. Maize endophytic bacteria as mineral phosphate solubilizers. Genet. Mol. Res. 2017, 16. [Google Scholar] [CrossRef]
- Shahzad, R.; Khan, A.; Bilal, S.; Waqas, M.; Kang, S.; Lee, I. Inoculation of abscissic acid-producing endophytic bacteria enances salinity stress tolerance in Oryza sativa. Environ. Exp. Bot. 2017, 136, 68–77. [Google Scholar] [CrossRef]
- Defez, R.; Andreozzi, A.; Bianco, C. The overproduction of Indole-3-Acetic Acid (IAA) in endophytes upregulates nitrogen fixation in both bacterial cultures and inoculated rice plants. Microb. Ecol. 2017, 74, 441–452. [Google Scholar] [CrossRef]
- Compant, S.; Reiter, B.; Sessitsch, A.; Nowak, J.; Clément, C.; Ait Barka, E. Endophytic colonization of Vitis vinifera L. by plant growth-promoting bacterium Burkholderia sp. Strain PsJN. Appl. Environ. Microbiol. 2005, 71, 1685–1693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowman, S.; Kim-Dura, S.; Mei, C.; Nowak, J. Strategies for enhancement of switchgrass (Panicum virgatum L.) performance under limited nitrogen supply based on utilization of N-fixing bacterial endophytes. Plant Soil 2016, 405, 47–63. [Google Scholar] [CrossRef]
- Shabanamol, S.; Dibya, K.; George, T.; Rishad, K.; Sreekumar, T.; Jisha, M. Characterization and in planta nitrogen fixation of plant-growth promoting endophytic diazotrophyc Lysinibacillus sphæricus isolated from rice (Oryza sativa). Physiol. Mol. Plant Pathol. 2018, 102, 46–54. [Google Scholar] [CrossRef]
- Verma, V.C.; Singh, S.K.; Prakash, S. Bio-control and plant growth promotion potential of siderophore producing endophytic Streptomyces from Azadirachta indica A. Juss. J. Basic Microbiol. 2011, 51, 550–556. [Google Scholar] [CrossRef] [PubMed]
- Rosconi, F.; Davyt, D.; Martínez, V.; Martínez, M.; Abin-Carriquiry, J.A.; Zane, H.; Butler, A.; de Souza, E.M.; Fabiano, E. Identification and structural characterization of serobactins, a suite of lipopeptide siderophores produced by the grass endophyte Herbaspirillum seropedicae: Siderophores of Herbaspirillum seropedicae. Environ. Microbiol. 2013, 15, 916–927. [Google Scholar] [CrossRef]
- Brader, G.; Compant, S.; Mitter, B.; Trognitz, F.; Sessitsch, A. Metabolic potential of endophytic bacteria. Curr. Opin. Biotechnol. 2014, 27, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Hong, C.E.; Park, J.M. Endophytic bacteria as biocontrol agents against plant pathogens: Current state-of-the-art. Plant Biotechnol. Rep. 2016, 10, 353–357. [Google Scholar] [CrossRef]
- Glick, B.R. Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol. Res. 2014, 169, 30–39. [Google Scholar] [CrossRef]
- Gamalero, E.; Glick, B.R. Bacterial modulation of plant ethylene levels. Plant Physiol. 2015, 169, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Gamalero, E.; Bona, E.; Todeschini, V.; Lingua, G. Saline and arid soils: Impact on bacteria, plants, and their interaction. Biology 2020, 9, 116. [Google Scholar] [CrossRef]
- Gamalero, E.; Trotta, A.; Massa, N.; Copetta, A.; Martinotti, M.G.; Berta, G. Impact of two fluorescent pseudomonads and an arbuscular mycorrhizal fungus on tomato plant growth, root architecture and P acquisition. Mycorrhiza 2004, 14, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Rashid, S.; Charles, T.; Glick, B. Isolation and characterization of new plant growth-promoting bacterial endophytes. Appl. Soil Ecol. 2012, 61, 217–224. [Google Scholar] [CrossRef]
- Surette, M.A.; Sturz, A.V.; Lada, R.R.; Nowak, J. Bacterial endophytes in processing carrots (Daucus carota L. var. sativus): Their localization, population density, biodiversity and their effects on plant growth. Plant Soil 2003, 253, 381–390. [Google Scholar] [CrossRef]
- Bona, E.; Lingua, G.; Manassero, P.; Cantamessa, S.; Marsano, F.; Todeschini, V.; Copetta, A.; D’Agostino, G.; Massa, N.; Avidano, L.; et al. AM fungi and PGP pseudomonads increase flowering, fruit production, and vitamin content in strawberry grown at low nitrogen and phosphorus levels. Mycorrhiza 2015, 25, 181–193. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Schwyn, B.; Neilands, J. Universal chemical assay for the detection and determination of siderophore. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- de Brito, A.M.; Gagne, S.; Antoun, H. Effect of compost on rhizosphere microflora of the tomato and on the incidence of plant growth-promoting rhizobacteria. Appl. Environ. Microbiol. 1995, 61, 194–199. [Google Scholar] [CrossRef] [Green Version]
- Penrose, D.M.; Glick, B.R. Methods for isolating and characterizing ACC deaminase-containing plant growth-promoting rhizobacteria. Physiol. Plant. 2003, 118, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Duan, J.; Jiang, W.; Cheng, Z.; Heikkila, J.J.; Glick, B.R. The complete genome sequence of the plant growth-promoting bacterium Pseudomonas sp. UW4. PLoS ONE 2013, 8, e58640. [Google Scholar] [CrossRef] [Green Version]
- Loper, J.E.; Haack, C.; Schroth, M.N. Population dynamics of soil pseudomonads in the rhizosphere of potato (Solanum tuberosum L.). Appl. Environ. Microbiol. 1985, 49, 416–422. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wu, X.; Chen, T.; Wang, W.; Liu, G.; Zhang, W.; Li, S.; Wang, M.; Zhao, C.; Zhou, H.; et al. Plant phenotypic traits eventually shape its microbiota: A common garden test. Front. Microbiol. 2018, 9, 2479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spaepen, S.; Vanderleyden, J.; Remans, R. Indole-3-acetic acid in microbial and microorganism-plant signaling. FEMS Microbiol. Rev. 2007, 31, 425–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duca, D.; Lorv, J.; Patten, C.L.; Rose, D.; Glick, B.R. Indole-3-acetic acid in plant–microbe interactions. Antonie Van Leeuwenhoek 2014, 106, 85–125. [Google Scholar] [CrossRef] [PubMed]
- Gamalero, E.; Glick, B.R. Plant growth-promoting bacteria in agriculture and stress environments. In Modern Soil Microbiology, 3rd ed.; Van Elsas, J.D., Trevors, J.T., Rosado, A.S., Nannipieri, P., Eds.; Springer: New Delhi, India, 2019; pp. 361–380. [Google Scholar]
- Khan, M.S.; Zaidi, A.; Wani, P.A. Role of phosphate-solubilizing microorganisms in sustainable agriculture —A review. Agron. Sustain. Dev. 2007, 27, 29–43. [Google Scholar] [CrossRef]
- Liang, Y.; Tóth, K.; Cao, Y.; Tanaka, K.; Espinoza, C.; Stacey, G. Lipochitooligosaccharide recognition: An ancient story. New Phytol. 2014, 204, 289–296. [Google Scholar] [CrossRef]
- Chabot, R.; Antoun, H.; Cescas, M.P. Growth promotion of maize and lettuce by phosphate-solubilizing Rhizobium leguminosarum biovar. phaseoli. Plant Soil 1996, 184, 311–321. [Google Scholar] [CrossRef]
- Loper, J.; Buyer, J. Siderophores in microbial interactions on plant surfaces. Mol. Plant. Microbe Interact. 1990, 4, 5–13. [Google Scholar] [CrossRef]
- Mimmo, T.; Del Buono, D.; Terzano, R.; Tomasi, N.; Vigani, G.; Crecchio, C.; Pinton, R.; Zocchi, G.; Cesco, S. Rhizospheric organic compounds in the soil-microorganism-plant system: Their role in iron availability. Eur. J. Soil Sci. 2014, 65, 629–642. [Google Scholar] [CrossRef]
- Marschner, H.; Römheld, V.; Kissel, M. Different strategies in higher plants in mobilization and uptake of iron. J. Plant Nutr. 1986, 9, 695–713. [Google Scholar] [CrossRef]
- Vansuyt, G.; Robin, A.; Briat, J.-F.; Curie, C.; Lemanceau, P. Iron Acquisition from Fe-Pyoverdine by Arabidopsis thaliana. Mol. Plant-Microbe Interact. 2007, 20, 441–447. [Google Scholar] [CrossRef] [Green Version]
- Aznar, A.; Dellagi, A. New insights into the role of siderophores as triggers of plant immunity: What can we learn from animals? J. Exp. Bot. 2015, 66, 3001–3010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vleesschauwer, D.; Djavaheri, M.; Bakker, P.A.H.M.; Höfte, M. Pseudomonas fluorescens WCS374r-Induced Systemic Resistance in Rice against Magnaporthe oryzae Is Based on Pseudobactin-Mediated Priming for a Salicylic Acid-Repressible Multifaceted Defense Response. Plant Physiol. 2008, 148, 1996–2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerqueira, F.; Matamoros, V.; Bayona, J.; Piña, B. Antibiotic resistance genes distribution in microbiomes from the soil-plant-fruit continuum in commercial Lycopersicon esculentum fields under different agricultural practices. Sci. Total Environ. 2019, 652, 660–670. [Google Scholar] [CrossRef] [PubMed]
- Burch, T.R.; Sadowsky, M.J.; LaPara, T.M. Effect of different treatment technologies on the fate of antibiotic resistance genes and Class 1 integrons when residual municipal wastewater solids are applied to soil. Environ. Sci. Technol. 2017, 51, 14225–14232. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Ye, M.; Schwab, A.; Li, X.; Wan, J.; Wei, Z.; Wu, J.; Frimann, V.; Liu, K.; Tian, D.; et al. Human migration activities drive the fluctuation of ARGs: Case study of landfills in Nanjing, Eastern China. J. Hazard. Mater. 2016, 315, 93–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, M.; Chu, L.M. Occurrence of antibiotics and antibiotic resistance genes in soils from wastewater irrigation areas in the Pearl River Delta region, southern China. Sci. Total Environ. 2018, 624, 145–152. [Google Scholar] [CrossRef]
- Tien, Y.-C.; Li, B.; Zhang, T.; Scott, A.; Murray, R.; Sabourin, L.; Marti, R.; Topp, E. Impact of dairy manure pre-application treatment on manure composition, soil dynamics of antibiotic resistance genes, and abundance of antibiotic-resistance genes on vegetables at harvest. Sci. Total Environ. 2017, 581, 32–39. [Google Scholar] [CrossRef]
- Berendonk, T.U.; Manaia, C.M.; Merlin, C.; Fatta-Kassinos, D.; Cytryn, E.; Walsh, F.; Bürgmann, H.; Sørum, H.; Norström, M.; Pons, M.-N.; et al. Tackling antibiotic resistance: The environmental framework. Nat. Rev. Microbiol. 2015, 13, 310–317. [Google Scholar] [CrossRef]
- Koch, B.J.; Hungate, B.A.; Price, L.B. Food-animal production and the spread of antibiotic resistance: The role of ecology. Front. Ecol. Environ. 2017, 15, 309–318. [Google Scholar] [CrossRef]
- Fang, H.; Wang, H.; Cai, L.; Yu, Y. Prevalence of antibiotic resistance genes and bacterial pathogens in long-term manured greenhouse soils as revealed by metagenomic survey. Environ. Sci. Technol. 2015, 49, 1095–1104. [Google Scholar] [CrossRef]
- Olaimat, A.N.; Holley, R.A. Factors influencing the microbial safety of fresh produce: A review. Food Microbiol. 2012, 32, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Deering, A.J.; Mauer, L.J.; Pruitt, R.E. Internalization of E. coli O157:H7 and Salmonella spp. in plants: A review. Food Res. Int. 2012, 45, 567–575. [Google Scholar] [CrossRef]
- Kljujev, I.; Raicevic, V.; Vujovic, B.; Rothballer, M.; Schmid, M. Salmonella as an endophytic colonizer of plants—A risk for health safety vegetable production. Microb. Pathog. 2018, 115, 199–207. [Google Scholar] [CrossRef] [PubMed]
- de Costa, L.E.O.; de Queiroz, M.V.; Borges, A.C.; de Moraes, C.A.; de Araújo, E.F. Isolation and characterization of endophytic bacteria isolated from the leaves of the common bean (Phaseolus vulgaris). Braz. J. Microbiol. 2012, 43, 1562–1575. [Google Scholar] [CrossRef]
- Gamalero, E.; Marzachì, C.; Galetto, L.; Veratti, F.; Massa, N.; Bona, E.; Novello, G.; Glick, B.R.; Ali, S.; Cantamessa, S.; et al. An 1-Aminocyclopropane-1-carboxylate (ACC) deaminase-expressing endophyte increases plant resistance to flavescence dorée phytoplasma infection. Plant Biosyst.-Int. J. Deal. Asp. Plant Biol. 2017, 151, 331–340. [Google Scholar] [CrossRef]
- Chung, B.-S.; Kang, H.-S.; Ahn, J.-W.; Chung, Y.R. A Bacterial endophyte, Pseudomonas brassicacearum YC5480, isolated from the root of Artemisia sp. producing antifungal and phytotoxic compounds. Plant Pathol. J. 2008, 24, 461–468. [Google Scholar] [CrossRef]
- Ludlow, M.M.; Muchow, R.C. A critical evaluation of traits for improving crop yields in water-limited environments. In Advances in Agronomy; Academic Press: Cambridge, UK, 1990; Volume 43, pp. 107–153. [Google Scholar]
- Niu, G.; Xu, W.; Rodriguez, D.; Sun, Y. Growth and physiological responses of maize and sorghum genotypes to salt stress. ISRN Agron. 2012, 2012, 145072. [Google Scholar] [CrossRef] [Green Version]
- Gamalero, E.; Glick, B. The use of microorganisms to facilitate the growth of plant in saline soil. In Microbial Strategies for Crop Improvement; Springer: Cham, Switzerland, 2009; pp. 1–22. [Google Scholar]
- Rho, H.; Hsieh, M.; Kandel, S.L.; Cantillo, J.; Doty, S.L.; Kim, S.-H. Do endophytes promote growth of host plants under stress? A meta-analysis on plant stress mitigation by endophytes. Microb. Ecol. 2018, 75, 407–418. [Google Scholar] [CrossRef]
- Eida, A.A.; Ziegler, M.; Lafi, F.F.; Michell, C.T.; Voolstra, C.R.; Hirt, H.; Saad, M.M. Desert plant bacteria reveal host influence and beneficial plant growth properties. PLoS ONE 2018, 13, e0208223. [Google Scholar] [CrossRef]
- Ali, S.; Charles, T.C.; Glick, B.R. Amelioration of high salinity stress damage by plant growth-promoting bacterial endophytes that contain ACC deaminase. Plant Physiol. Biochem. 2014, 80, 160–167. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Origin | Taxonomic Identification | GenBank Accession Number | IAA a (Intensity Colour Scale 0–5) | Siderophore (HD/CD b cm) | Phosphate Solubilisation (DCP c, HD/CD cm) |
|---|---|---|---|---|---|---|
| CVB2R4 | cucumber root, pasture soil | Herbaspirillum lusitanum | KX436986 | 3 | 0.00 | 0.00 |
| CVB2R5 | cucumber root, pasture soil | Herbaspirillum lusitanum | KX436987 | 3 | 0.00 | 0.00 |
| CVB3S1 | cucumber shoot, pasture soil | Acinetobacter johnsonii | KX436989 | 0 | 0.00 | 0.00 |
| CVB3S2 | cucumber shoot, pasture soil | Acinetobacter johnsonii | KX437624 | 1 | 0.00 | 0.00 |
| CVB3S4 | cucumber shoot, pasture soil | Acinetobacter johnsonii | KX437625 | 0 | 0.00 | 0.00 |
| CVB3S5 | cucumber shoot, pasture soil | Stenotrophomonas rhizophila | KX437653 | 3 | 0.00 | 0.00 |
| CO3R3 | cucumber root, vegetable garden | Agrobacterium tumefaciens | KX429744 | 4 | 2.96 | 0.00 |
| CO3L1 | cucumber leaf, vegetable garden | Agrobacterium tumefaciens | KX429748 | 0 | 0.00 | 0.00 |
| CO4R1 | cucumber root, vegetable garden | Agrobacterium tumefaciens | KX429749 | 0 | 0.00 | 0.00 |
| CO4S1 | cucumber shoot, vegetable garden | Rhizobium radiobacter | MF993501 | 4 | 3.53 | 0.00 |
| CO4S3 | cucumber shoot, vegetable garden | Agrobacterium tumefaciens | KX436985 | 2 | 2.73 | 0.00 |
| CO5R1 | cucumber root, vegetable garden | Micrococcus yunnanensis | KX436988 | 0 | 0.00 | 0.00 |
| CL1S1 | cucumber shoot, chestnut woody soil | Paenibacillus graminis | KX404972 | 0 | 0.00 | 0.00 |
| CL1S2 | cucumber shoot, chestnut woody soil | Bacillus pumilus | KX421200 | 2 | 0.00 | 0.00 |
| CL6S1 | cucumber shoot, chestnut woody soil | Bacillus cereus | KX421202 | 0 | 0.00 | 0.00 |
| CL6S2 | cucumber shoot, chestnut woody soil | Bacillus pumilus | KX429743 | 2 | 0.00 | 1.72 |
| CL6S5 | cucumber shoot, chestnut woody soil | Bacillus muralis | KX429745 | 0 | 0.00 | 0.00 |
| CL6S9 | cucumber shoot, chestnut woody soil | Bacillus muralis | KX429746 | 0 | 3.44 | 0.00 |
| CL6S10 | cucumber shoot, chestnut woody soil | Terribacillus goriensis | KX429747 | 0 | 0.00 | 0.00 |
| SVB3R1 | sorghum root, pasture soil | Bacillus cereus | KX444208 | 0 | 0.00 | 0.00 |
| SVB3R2 | sorghum root, pasture soil | Pseudomonas migulae | MF993051 | 0 | 4.51 | 1.45 |
| SVB3R3 | sorghum root, pasture soil | Pseudomonas migulae | MF993115 | 0 | 5.63 | 0.62 |
| SVB3R4 | sorghum root, pasture soil | Pseudomonas migulae | MF993122 | 0 | 4.10 | 1.45 |
| SVB3R5 | sorghum root, pasture soil | Pseudomonas sp. | KX447593 | 0 | 3.93 | 1.72 |
| SVB6R1 | sorghum root, pasture soil | Pseudomonas brassicacearum | KX444672 | 3 | 5.55 | 2.92 |
| SVB6R2 | sorghum root, pasture soil | Paenibacillus lautus | KX444671 | 1 | 0.00 | 0.00 |
| SL3R1 | sorghum root, chestnut wood | Brevibacterium frigoritolerans | KX447586 | 0 | 2.78 | 0.00 |
| SL3R3 | sorghum root, chestnut wood | Bacillus anthracis | KX442610 | 0 | 0.00 | 0.00 |
| SL3R4 | sorghum root, chestnut wood | Paenibacillus illinoisensis | KX442643 | 0 | 0.00 | 1.52 |
| SL3R5 | sorghum root, chestnut wood | Bacillus muralis | KX444201 | 0 | 2.97 | 0.00 |
| SL3R6 | sorghum root, chestnut wood | Pseudomonas sp. | MF993023 | 2 | 2.76 | 1.50 |
| SL3R8 | sorghum root, chestnut wood | Bacillaceae bacterium | KX447594 | 0 | 0.00 | 0.00 |
| SL3L1 | sorghum root, chestnut wood | Micrococcus luteus | KX447587 | 1 | 0.00 | 0.00 |
| PVB1S1 | tomato shoot, pasture soil | Bacillus safensis | KX440184 | 0 | 0.00 | 1.54 |
| PVB1S2 | tomato shoot, pasture soil | Bacillaceae bacterium | KX442613 | 0 | 0.00 | 1.50 |
| PVB1S3 | tomato shoot, pasture soil | Acinetobacter lwoffii | KX440393 | 0 | 0.00 | 0.00 |
| PVB1S5 | tomato shoot, pasture soil | Bacillus cereus | KX440517 | 0 | 0.00 | 0.00 |
| PVB1L1 | tomato leaf, pasture soil | Bacillus thuringiensis | KX442563 | 0 | 0.00 | 0.00 |
| PVB1L2 | tomato leaf, pasture soil | Bacillus muralis | KX437755 | 0 | 5.04 | 0.00 |
| PVB1L3 | tomato leaf, pasture soil | Bacillus cereus | KX437756 | 0 | 0.00 | 1.37 |
| PVB1L5 | tomato leaf, pasture soil | Bacillus cereus | KX438058 | 0 | 0.00 | 0.00 |
| PVB1L6 | tomato leaf, pasture soil | Bacillus megaterium | KX438317 | 0 | 2.92 | 0.00 |
| PVB1L7 | tomato leaf, pasture soil | Bacillus tequilensis | KX438316 | 0 | 2.92 | 1.96 |
| PVB1L8 | tomato leaf, pasture soil | Bacillus cereus | KX438379 | 0 | 0.00 | 0.00 |
| PVB1R1 | tomato root, pasture soil | Bacillus aerophilus | KX440185 | 0 | 0.00 | 0.00 |
| PVB6R2 | tomato root, pasture soil | Bacillus muralis | KX440975 | 0 | 4.96 | 0.00 |
| PVB6R3 | tomato root, pasture soil | Bacillus thuringiensis | KX443415 | 0 | 0.00 | 1.42 |
| PVB6R4 | tomato root, pasture soil | Bacillaceae bacterium | KX443572 | 0 | 0.00 | 0.00 |
| PVB6R5 | tomato root, pasture soil | Bacillus thuringiensis | KX443561 | 0 | 0.00 | 0.00 |
| PVB6L1 | tomato leaf, pasture soil | Acinetobacter johnsonii | KX440619 | 2 | 0.00 | 0.00 |
| PVB6L2 | tomato leaf, pasture soil | Microbacterium schleiferi | KX440956 | 0 | 0.00 | 1.79 |
| PVB6L3 | tomato leaf, pasture soil | Acinetobacter johnsonii | KX440957 | 1 | 0.00 | 0.00 |
| PVB6L4 | tomato leaf, pasture soil | Bacillus cereus | KX440973 | 0 | 0.00 | 1.55 |
| PVB7R1 | tomato root, pasture soil | Bacillus subtilis | KX442589 | 0 | 1.58 | 2.00 |
| PVB7R2 | tomato root, pasture soil | Bacillus cereus | KX442609 | 0 | 0.00 | 0.00 |
| PO2R2 | tomato root, vegetable garden | Paenibacillus sp. | KX442611 | 0 | 0.00 | 0.00 |
| PO2L1 | tomato leaf, vegetable garden | Bacillus niacini | KX437751 | 0 | 0.00 | 0.00 |
| PO3S1 | tomato shoot, vegetable garden | Bacillus thuringiensis | KX442562 | 0 | 0.00 | 0.00 |
| PO7S1 | tomato shoot, vegetable garden | Kochuria palustris | KX442561 | 0 | 1.83 | 1.61 |
| H. lusitanum CVB2R5 | S. rhizophila CVB3S5 | P. migulae SVB3R2 | P. migulae SVB3R3 | P. migulae SVB3R4 | Pseudomonas sp. SVB3R5 | P. brassicacearum SVB6R1 | Pseudomonas sp. SL3R6 | |
|---|---|---|---|---|---|---|---|---|
| Ceftazidime 30 µg | S | S | S | S | S | S | S | S |
| Cotrimoxazole 25 µg | S | S | S | S | S | S | S | S |
| Gentamicin 10 µg | S | S | S | S | S | S | S | S |
| Ciprofloxacin 5 µg | S | S | S | S | S | S | S | S |
| Nalidixic acid 30 µg | S | S | S | S | S | S | S | S |
| Nitrofurans 100 µg | R | R | R | R | R | R | R | R |
| Cefoperazone30 µg | S | R | S | S | S | S | S | S |
| Phosphomycin 50 µg | R | S | S | S | S | S | S | S |
| Cefixime 10 µg | S | R | S | S | S | S | R | S |
| Norfloxacin 10 µg | S | S | S | S | S | S | S | S |
| Bacterial Strain | Log CFU/g Plant Tissue | |
|---|---|---|
| Root | Leaves | |
| Pseudomonas migulae SVB3R2 | 3.48 ± 1.01 | 3.76 ± 1.19 |
| Pseudomonas migulae SVB3R4 | 3.47 ± 1.15 | n.d. |
| Pseudomonas brassicacearum SVB6R1 | 3.18 ± 1.11 | 3.43 ± 1.09 |
| Pseudomonas migulae SVB3R3 | 3.37 ± 1.00 | 3.53 ± 1.02 |
| Pseudomonas sp. SVB3R5 | 3.63 ± 1.25 | 2.83 ± 1.07 |
| Pseudomonas sp. SL3R6 | n.d. | n.d. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gamalero, E.; Favale, N.; Bona, E.; Novello, G.; Cesaro, P.; Massa, N.; Glick, B.R.; Orozco-Mosqueda, M.d.C.; Berta, G.; Lingua, G. Screening of Bacterial Endophytes Able to Promote Plant Growth and Increase Salinity Tolerance. Appl. Sci. 2020, 10, 5767. https://doi.org/10.3390/app10175767
Gamalero E, Favale N, Bona E, Novello G, Cesaro P, Massa N, Glick BR, Orozco-Mosqueda MdC, Berta G, Lingua G. Screening of Bacterial Endophytes Able to Promote Plant Growth and Increase Salinity Tolerance. Applied Sciences. 2020; 10(17):5767. https://doi.org/10.3390/app10175767
Chicago/Turabian StyleGamalero, Elisa, Nicoletta Favale, Elisa Bona, Giorgia Novello, Patrizia Cesaro, Nadia Massa, Bernard R. Glick, Ma del Carmen Orozco-Mosqueda, Graziella Berta, and Guido Lingua. 2020. "Screening of Bacterial Endophytes Able to Promote Plant Growth and Increase Salinity Tolerance" Applied Sciences 10, no. 17: 5767. https://doi.org/10.3390/app10175767