 Sara Sigurlásdóttir
Sara Sigurlásdóttir Gabriela M. Wassing
Gabriela M. Wassing Fanglei Zuo
Fanglei Zuo Melanie Arts
Melanie Arts Ann-Beth Jonsson
Ann-Beth Jonsson- Department of Molecular Biosciences, The Wenner-Gren Institute, Stockholm University, Stockholm, Sweden
Neisseria meningitidis is a Gram-negative bacterium that asymptomatically colonizes the human nasopharyngeal mucosa. Pilus-mediated initial adherence of N. meningitidis to the epithelial mucosa is followed by the formation of three-dimensional aggregates, called microcolonies. Dispersal from microcolonies contributes to the transmission of N. meningitidis across the epithelial mucosa. We have recently discovered that environmental concentrations of host cell-derived lactate influences N. meningitidis microcolony dispersal. Here, we examined the ability of N. meningitidis mutants deficient in lactate metabolism to form biofilms. A lactate dehydrogenease A (ldhA) mutant had an increased level of biofilm formation. Deletion of ldhA increased the N. meningitidis cell surface hydrophobicity and aggregation. In this study, we used FAM20, which belongs to clonal complex ST-11 that forms biofilms independently of extracellular DNA (eDNA). However, treatment with DNase I abolished the increased biofilm formation and aggregation of the ldhA-deficient mutant, suggesting a critical role for eDNA. Compared to wild-type, the ldhA-deficient mutant exhibited an increased autolytic rate, with significant increases in the eDNA concentrations in the culture supernatants and in biofilms. Within the ldhA mutant biofilm, the transcription levels of the capsule, pilus, and bacterial lysis genes were downregulated, while norB, which is associated with anaerobic respiration, was upregulated. These findings suggest that the absence of ldhA in N. meningitidis promotes biofilm formation and aggregation through autolysis-mediated DNA release.
Introduction
The Gram-negative human-restricted pathogen Neisseria meningitidis is the causative agent of meningitis and sepsis. Meningococci frequently colonize the respiratory epithelium as a non-invasive commensal, although in rare cases, the bacteria can cross the cell barrier and cause invasive disease (Pace and Pollard, 2012). The initial attachment of bacteria is facilitated by Type IV pili (Tfp), thin filaments that extend from the bacterial surface. After the initial adhesion of meningococci to the epithelium, the bacteria form Tfp-dependent three-dimensional aggregates, called microcolonies. Microcolony formation can develop into bacterial biofilms, i.e., bacterial communities that can persist and avoid removal by mechanical forces (Costerton et al., 1995). Studies have demonstrated the long-term asymptomatic carriage of N. meningitidis in the nasopharynx (Ala’Aldeen et al., 2000; Sim et al., 2000; Wilder-Smith et al., 2003) and the existence of microcolonies in patient biopsies (Harrison et al., 2002).
Several factors have been identified that contribute to meningococcal biofilm formation in vitro. In many bacterial species, the extracellular matrix is composed of polysaccharides, but N. meningitidis does not produce extracellular polysaccharides (Yi et al., 2004; Lappann et al., 2010). In most meningococcal genetic lineages, extracellular DNA (eDNA) is a major component of the extracellular matrix, contributing both to the initial binding to surfaces and the stabilization of the biofilm structure. The release of DNA by meningococci is thought to occur through bacterial lysis, mediated by lytic transglycosylases (MltA and MltB) and N-acetylmuramuyl-L-alanine amidase (AmpD) in early-stage biofilms. In late-stage biofilms, the outer membrane phospholipase A (OMPLA) mediates DNA release, which contributes to the resistance against mechanical forces (Lappann et al., 2010). Positively charged surface attached proteins are involved in biofilm formation by binding to eDNA. When surface exposed, Neisseria heparin-binding antigen (NhbA), as well as the autotransporters IgA protease and AutA can bind eDNA and promote biofilm formation. The autotransporter NalP can proteolytically cleave surface-bound NhbA and IgA protease, leading to reduced biofilm formation (Arenas et al., 2013, 2015). Biofilm formation in the absence of eDNA has also been reported, although this is relatively less protected against shear forces. eDNA-independent biofilms are usually formed by disease isolates and are therefore associated with high transmission rates (Lappann and Vogel, 2010; Lappann et al., 2010).
During biofilm formation, the availability of oxygen is limited (Werner et al., 2004; Barraud et al., 2006). It has been reported that the metabolism of N. gonorrhoeae adapts to growth in anaerobic environments via aniA, ccp, and norB upregulation (Falsetta et al., 2009). Meningococci also possess aniA and norB. Expression of aniA is under control of FNR (fumarate and nitrate reduction regulator), a master regulator of adaptation to anaerobic growth, and the two-component system NarQ/NarP in the presence of nitrite (Householder et al., 1999; Rock et al., 2005; Bartolini et al., 2006; Falsetta et al., 2009). Expression of norB is influenced by the nitric oxide concentrations (Householder et al., 2000; Rock et al., 2007). The meningococcal aniA may not always be active due to frame shift mutation (Barth et al., 2009), whereas norB is more conserved among strains. Further, the gonococcal norB/aniA cassette has been reported to by gene conversion appear in meningococcal strains causing urethritis (Tzeng et al., 2017). Deletion of narP, which encodes the regulator of the NarQ/NarP system, leads to defects in meningococcal biofilm formation (Jamet et al., 2013).
Unencapsulated strains are often isolated from healthy carriers, while for the development of disease, the capsule is almost indispensable (Stephens et al., 2007). Although the polysaccharide capsule is important for meningococcal immune evasion, it inhibits both intimate adhesion of the bacteria to host cells and biofilm formation on abiotic surfaces (de Vries et al., 1996; Yi et al., 2004; Lappann et al., 2006). Downregulation of the capsule is thought to occur upon Tfp-dependent adhesion to host cells and during biofilm formation. The absence of the capsule will expose surface-expressed molecules that support intimate adhesion, invasion, and biofilm modes of growth (Deghmane et al., 2002; Tzeng et al., 2008; O’Dwyer et al., 2009). It has, however, been reported that meningococcal biofilm formation on human airway epithelial cells is not inhibited by the presence of capsule (Neil et al., 2009). Lipooligosaccharide (LOS) has also been shown to have negative effects on meningococcal biofilm formation (Yi et al., 2004; Lappann and Vogel, 2010). A decrease in LOS biosynthesis transcripts has been observed during biofilm growth (O’Dwyer et al., 2009). Tfp is important in the initial attachment of meningococci to host cell surfaces, for bacterial aggregation, and twitching motility (reviewed in Craig et al., 2004). Tfp-mediated bacterial aggregation has been shown to play a role in meningococcal biofilm formation. Additionally, non-pilated and non-aggregative strains might form biofilms, although their flat architectures are different from those of microcolony-based biofilms (Yi et al., 2004; Lappann et al., 2006). It has been proposed that the reduction in twitching motility, rather than aggregation, in these strains affects the establishment of microcolonies within the biofilms (Wolfgang et al., 1998; Lappann et al., 2006). The surface-exposed autotransporter AutA has been reported to promote both aggregation and biofilm formation by binding to itself and eDNA (Arenas et al., 2015).
We have previously found that host-cell derived lactate induces the dispersal of meningococcal aggregates in liquid cultures (Sigurlasdottir et al., 2017). Additionally, lactate dehydrogenases (LDHs) have been implicated in the biofilm formation of multiple bacteria. In iron-supplemented medium, LDH promotes biofilm growth of the gut bacteria Enterococcus faecalis (Keogh et al., 2018). LDH promotes biofilm formation under anaerobic conditions of the oral bacteria Streptococcus sanguinis (Ge et al., 2016). Conversely, in Escherichia coli, deletion of ldhA and other genes that are important in acetate metabolism increased the level of biofilm formation. The loss of ldhA and lactate dehydrogenase regulator (ldhR) in Burkholderia cepacia increased cell viability and production of extracellular matrix, while aggregation and biofilm formation were reduced (Silva et al., 2017). However, the role of the enzymes involved in lactate metabolism in biofilm formation of N. meningitidis is not clearly understood. We, therefore, investigated the roles of the enzymes involved in lactate metabolism in meningococcal biofilm formation.
In the present study, we demonstrate that the absence of D-LDH LdhA increased the amount of biofilm formed by the meningococcal strain FAM20. Additionally, ldhA mutants showed stronger cell surface hydrophobicity and aggregation. The observed increased biofilm formation and aggregation were the results of the increase in the autolytic rate and the amount of eDNA. Taken together, our findings show that the deletion of ldhA promotes biofilm formation and aggregation through autolysis-mediated DNA release.
Materials and Methods
Bacterial Strains and Growth Conditions
Neisseria meningitidis strain FAM20 (FAM18 derivative) and its mutants deficient in capsule operon (Δcap) or lactate metabolic genes (ΔlctP, ΔlldA, ΔldhD, and ΔldhA) have been described previously (Jones et al., 2009; Sigurlasdottir et al., 2017). Bacterial strains were grown on GC agar plates containing a 1% Kellogg’s supplement for 16–18 h at 37°C and 5% CO2. GC liquid containing 1% Kellogg’s supplement was used for experiments performed in liquid culture. For the selection of FAM20 mutant strains, appropriate antibiotics were used: tetracycline (1 μg/ml), chloramphenicol (2 μg/ml), and kanamycin (50 μg/ml). DMEM containing 10% FBS was used as a medium for human cells.
Construction of Mutant Strains
Phusion DNA polymerase (Thermo Fisher Scientific) was used for PCR amplification. All primers used in the study are presented in Table 1. All constructs generated were incorporated into the genome of wild-type N. meningitidis FAM20 or relevant mutants by performing spot transformation, plated and selected on plates containing relevant antibiotics. PCR and sequencing were performed to confirm the correct location and sequence of the constructs in the genome.
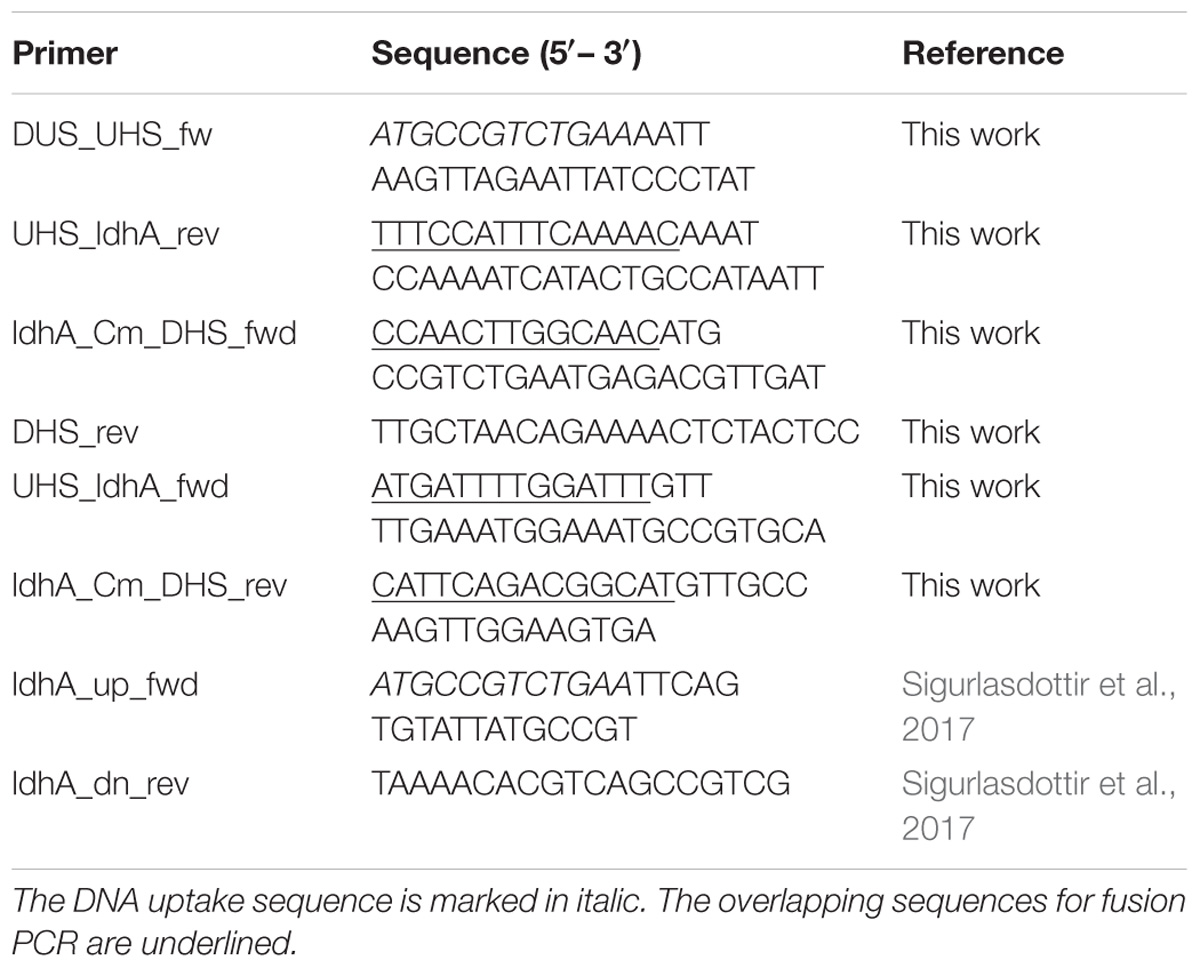
Table 1. Primers used for generation of mutants.
To generate the construct for complementation of the ΔldhA mutation, a fusion PCR was performed. The ldhA gene, including its native promoter, was introduced into a non-coding region between NMC0075 and NMC0080. First, an upstream homologous sequence containing the DUS sequence was amplified from FAM20 genomic DNA using primers DUS_UHS_fw and UHS_ldhA_rev. The downstream homologous sequence containing a chloramphenicol resistance cassette was amplified from a plasmid pDONR P4-P1R, as previously described (Kuwae et al., 2011), with primers ldhA_Cm_DHS_fwd and DHS_rev. The ldhA gene and its native promoter were amplified from FAM20 genomic DNA using primers UHS_ldhA_fwd and ldhA_Cm_DHS_rev. After amplification of all PCR products containing overlapping sequences, two-step fusion PCR reactions were performed. The first reaction was performed in the absence of primers to anneal overlapping sequences of the non-coding regions to ldhA and its native promoter. Later, the primers DUS_UHS_fw and DHS_rev were added to amplify the construct, which was then integrated into the FAM20 ΔldhA genome.
To obtain a Δcap/ΔldhA double mutant, the ΔldhA construct was amplified with the primer pair ldhA_up_fwd and ldhA_dn_rev (Sigurlasdottir et al., 2017). The complete construct was then integrated into the Δcap mutant.
Biofilm Assay Under Static Conditions
The assay has been described previously (Engman et al., 2016). Bacteria were grown on GC plates overnight and suspended to an OD of 0.05. One-hundred microliters of the bacterial solution was added in triplicate to 96-well polystyrene plates and allowed to grow under static conditions for 24 h at 37°C and 5% CO2. After incubation, the samples were washed twice with PBS to remove unbound bacteria. For quantification, the biofilm was stained for 2 min with 0.3% crystal violet, washed twice with PBS to remove unbound dye and solubilized in 30% acetic acid. The absorbance was measured at 630 nm (Yi et al., 2004) using a POLARstar Omega microplate reader. When necessary, DNase I was added at a final concentration of 100 μg/ml. As a control, DNase I was heat inactivated for 10 min at 65°C. Biofilm assays were also performed in the presence of 0.1, 1, and 10 mM sodium acetate. The assay was performed at least three times using triplicate samples. The absorbance values were normalized against the wild-type.
Growth Assays
The wild-type strain and its isogenic mutants were resuspended to an OD600 of 0.1. Cultures were grown under shaking conditions at 37°C and 5% CO2. The absorbance was measured every hour for 7 h. The growth assay was performed three times in duplicate. For OD measurement after 24 h, the wild-type and ΔldhA strains were resuspended to an OD of 0.05, and the cultures were grown under shaking conditions at 37°C and 5% CO2. Samples were measured three times in duplicate. For OD measurement in the presence of DNase I, the wild-type and ΔldhA strains were resuspended to an OD of 0.05 with or without 100 μg/ml DNase I and grown under shaking conditions at 37°C and 5% CO2. Samples were measured twice.
Microscopy
The wild-type, ΔldhA, and ΔldhA/ldhA bacteria were resuspended to an OD600 of 0.05 and one ml was seeded per well containing a glass cover slip. After 24 h of incubation the wells were washed twice with PBS and fixed with 2% formaldehyde/PBS for 30 min. The samples were stained with 330 nM DAPI/PBS for 5 min, washed three times with PBS, and mounted with Vectashield mounting medium (Vector Laboratories). The samples were examined under Zeiss LSM 780 confocal laser scanning microscope. Image stacks at 0.4 μm intervals were acquired using 63×/1.4 NA oil objective and processed using ZEN Black Zeiss software. Nine stacks were combined to create each image.
Quantification of Bacterial Aggregation
The sedimentation assay has been described previously (Helaine et al., 2005). Bacterial suspensions were filtered through 5 μm filters to remove aggregates, and the absorbance was adjusted to OD600 of 0.1. After 3 h of incubation at 37°C and 5% CO2 with agitation, the cultures were moved to static conditions at room temperature. Sedimentation of bacterial aggregates was examined by measuring the OD600 at 20 min intervals. During DNase I treatment, a final concentration of 100 μg/ml was used. The assay was performed three times in duplicate and single samples in the presence of DNase. The absorbance values were normalized against the first time point (0 min). The rate of sedimentation was calculated for the first 20 min using the following formula (OD60020 min - OD6000 min)/(20 min – 0 min). The result gave the decline in absorbance per minute (Abs/min).
Western Blot
Bacteria from plates were resuspended to an OD of 0.05 and incubated for 3 h at 37°C and 5% CO2 under shaking conditions. Samples were taken, centrifuged and resuspended in 1 × sample buffer containing β-mercaptoethanol. The samples were heated at 95°C for 5 min. Proteins were separated on a gradient gel (4–15%, Bio-Rad) and transferred to an Immobilon-P membrane. For detection of PilE, a rabbit polyclonal antibody (1:5000) (Sjolinder and Jonsson, 2007) was used as a primary antibody, and IR-reactive dye conjugated rabbit antibody was used as a secondary antibody. The membrane was stripped to remove antibodies, and EF-Tu monoclonal mouse antibody (Hycult Biotech) was used together with IR-reactive dye conjugated anti-mouse antibody as a loading control. Blots were imaged with Odyssey IR scanner at 700 and 800 nm. For quantification, Image J (version 1.48) was used.
Adherence Assays
The human epithelial cell line FaDu (ATCC HTB-43) was grown to 100% confluence in 24-well plates. Wild-type, ΔldhA, and ΔldhA/ldhA bacteria were resuspended in DMEM supplemented with 1% FBS, filtered through a 5 μm pore filter to break bacterial aggregates, and used to infect cells at an MOI of 100 for 3 h at 37°C in a 5% CO2 environment. Unbound bacteria were washed away three times. The cells were lysed using 1% saponin, and the adhered bacteria were quantified by plating serial dilutions on GC agar plates. Cells were maintained in Dulbecco’s modified Eagle’s medium containing GlutaMAX and pyruvate (DMEM; Thermo Fisher Scientific) and supplemented with 10% heat-inactivated fetal bovine serum (FBS; Sigma-Aldrich).
Microbial Adhesion to Solvents (MATS)
This assay has been described previously (Ly et al., 2006). Briefly, bacteria from plates grown overnight were resuspended in PBS and filtered through 5 μm filters to remove aggregates. The absorbance of the filtered bacterial solution was adjusted to an OD600 of 0.4 and mixed with hexadecane at a ratio of 4:1. To mix the two phases, the solution was vortexed for 30 s and then allowed to stand at room temperature to separate for 15 min. The absorbance of the aqueous phase was measured, and the percentage of hydrophobicity was calculated using the formula [1 – (OD6002/OD6001)] × 100%. The assay was performed three times using triplicate samples.
ELISA
The capsule level was quantified using ELISA. Bacteria were grown overnight on GC plates and then resuspended in PBS to OD600 of 0.1. Bacterial solutions were heat-inactivated at 56°C for 60 min. The ELISA plate was coated with 100 μl of the bacterial solutions overnight at 4°C and then blocked in 2% bovine serum albumin (BSA) for 2 h. Wells were incubated with 95/678 monoclonal anti-capsule antibody (Jones et al., 2009) diluted 1:400 in 2% BSA for 2 h and then horseradish peroxidase-conjugated goat anti-mouse immunoglobulin G (diluted 1:5000 in 2% BSA) for 1 h. 3,3′,5,5′-tetramethylbenzidine (TMB) was used to detect peroxidase bound to the plate, and 1 M HCl was used as a stop solution. The absorbance was measured at 450 nm (reference wavelength, 490 nm) using a Spectramax i3x microplate reader. The assay was performed three times using triplicate samples. The absorbance values were normalized against those of the wild-type.
Autolysis Under Non-growth Conditions
The assay was adapted from a previously described method (Garcia and Dillard, 2006). Wild-type and ΔldhA bacteria from plates were resuspended to an OD600 of 0.05 and grown for 3 h. The bacteria were centrifuged, washed, resuspended in PBS and diluted to OD600 of 0.1 in 30 mM Tris-HCl buffer pH 8.0 or 30 mM Tris-HCl buffer pH 6.0. The decrease in turbidity was measured every 10 min at room temperature for 1 h and after that every 20 min. The initial turbidity (%) was calculated by using the formula (OD600 × min/OD600 0 min) × 100%. The bacteria were resuspended before every measurement to prevent the effects of sedimentation influencing the assay. The assay was performed four times.
DNA Quantification in Culture Supernatants and Biofilms
The assay has been described previously (Hamilton et al., 2005; Tang et al., 2013). Briefly, bacteria were resuspended to an OD600 of 0.05 and grown for 3 h. The bacteria were centrifuged at 13,000 rpm for 3 min, and the supernatants were collected and incubated with Quant-iT PicoGreen dsDNA dye (Thermo Fisher Scientific) at a 1:1 ratio. To determine the eDNA concentration in biofilms, a PicoGreen solution was prepared in TE buffer (1 μl:199 μl) and added directly to biofilms that had been washed two times with PBS. The solution was mixed in the well by pipetting 10 times. Samples mixed with the dye were incubated for 2–5 min before measurements. The dye has been shown to detect both single-stranded and double-stranded DNA (Hamilton et al., 2005; Lappann et al., 2010). DNA was quantified based on a Lambda DNA standard. The fluorescence was measured using the excitation and emission at 485 and 535 nm using a Spectramax i3x microplate reader.
Quantitative Real-Time PCR Analysis
For comparison of gene expression of the wild-type and ΔldhA strains, biofilm assays were performed as indicated above in 24-well glass-bottom plates in 1 ml and incubated for 24 h. Wells were washed carefully two times with PBS, and then the biofilm was resuspended in RNA-protect Bacteria Reagent (Qiagen) diluted at a 1:2 ratio. The samples were mixed extensively and incubated for 5 min. DNase I (final concentration at 0.5 mg/ml) and proteinase K (final concentration at 20 μg/ml) were added to the samples and incubated for 15 min to disrupt the biofilm. For log phase bacterial cultures, the wild-type and ΔldhA bacteria were resuspended to an OD600 of 0.05 and grown under shaking conditions for 3 h; then, samples were collected. Samples were resuspended in RNA-protect Bacteria Reagent (Qiagen) diluted at a 1:2 ratio, vortexed, and incubated for 5 min.
For RNA isolation of both log phase and biofilm samples, the bacteria were pelleted by centrifugation for 1 min at 15,000 × g, and RNA was purified using the RNeasy plus mini kit (Qiagen) according to the manufacturer’s protocol. The RNA yield and quality were analyzed using NanoDrop 8000. SuperScript VILO Master Mix (Thermo Fisher Scientific) with random hexamers was used for cDNA synthesis. LightCycler 480 Real-Time 480 SYBR Green I Master mix (Roche) was used to amplify the resulting cDNA in a LightCycler 480 Real-Time PCR System. The housekeeping gene rpsJ was used as a reference. The PCR program was according to the manufacturer’s instructions, with 40 cycles of amplification and annealing temperatures of 55 or 60°C. Melting curves were analyzed to verify primer pair specificity. Relative expression was analyzed using the LightCycler 480 Real-Time PCR System software. All primers used for the qPCR analysis are listed in Table 2.

Table 2. qPCR primers used in the study.
Acetate Quantification
Culture medium was collected from 24 h old biofilms and centrifuged at 13,000 rpm for 3 min, and the supernatants were collected. The total acetate concentration was measured using Acetate Colorimetric Assay Kit (Sigma-Aldrich) according to manufacturer’s instructions. As recommended by the manufacturer, a sample blank, excluding the Acetate Enzyme mix, was set up for each sample to adjust the effects of ATP and NADH background.
Statistical Analysis
Two-tailed, unpaired Student’s t-tests were used when comparing between two groups. ANOVA with Bonferroni’s post hoc test was used when comparing differences between more than two groups. P-values below 0.05 were accepted as statistically significant.
Results
Deletion of LdhA Influences the Level of Meningococcal Biofilm Formation Under Static Conditions
We have previously shown that lactate induces the dispersal of meningococcal aggregates (Sigurlasdottir et al., 2017). Because aggregation can lead to the establishment of biofilms, we aimed to examine whether deficiencies in lactate metabolism influenced N. meningitidis biofilm formation. Lactate permease (LctP) is responsible for lactate uptake (Exley et al., 2005, 2007), and lactate dehydrogenases (LDHs) catalyze the conversion of lactate into pyruvate. Meningococci are known to contain at least three LDHs (Fischer et al., 1994). LdhA is an NAD+-dependent cytoplasmic D-LDH that catalyzes the reversible conversion of pyruvate into D-lactate. Membrane-bound LdhD (D-LDH) and LldA (L-LDH) are respiratory enzymes that are restricted to the conversion of lactate to pyruvate (Figure 1A; Atack et al., 2014). As the capsule has been shown to have a negative impact on meningococcal biofilm formation on abiotic surface, we used an unencapsulated mutant of N. meningitidis FAM20 as the control (Jones et al., 2009). Analysis of biofilm levels of the four isogenic mutants lacking the genes encoding the three LDHs and LctP showed that ΔldhA formed more biofilm compared to the wild-type. Deletion of ΔlctP,ΔlldA, and ΔldhD did not have any effect on biofilm formation (Figure 1B). We also detected no difference in the growth rate between the mutants and wild-type strains in the medium used (Supplementary Figures 1A,B). Because meningococcal biofilm assays are usually performed in capsule-deficient backgrounds, we constructed a Δcap/ΔldhA double-mutant strain. The double-mutant strain had increased biofilm formation compared to that of Δcap (Figure 1C), further supporting the role of ldhA in biofilm formation. We next constructed a ldhA-complemented strain (ΔldhA/ldhA) with a copy of ldhA containing its native promoter into a non-coding region in the chromosome. The biofilm formation of ΔldhA/ldhA was at a similar level to that of the wild-type strain (Figure 1D). The increase in biofilm formation by the ΔldhA compared to that of the wild-type and ΔldhA/ldhA strains was also detected using confocal microscopy (Figure 2). Taken together, these data suggest that deletion of the lactate dehydrogenase ldhA promotes biofilm formation.
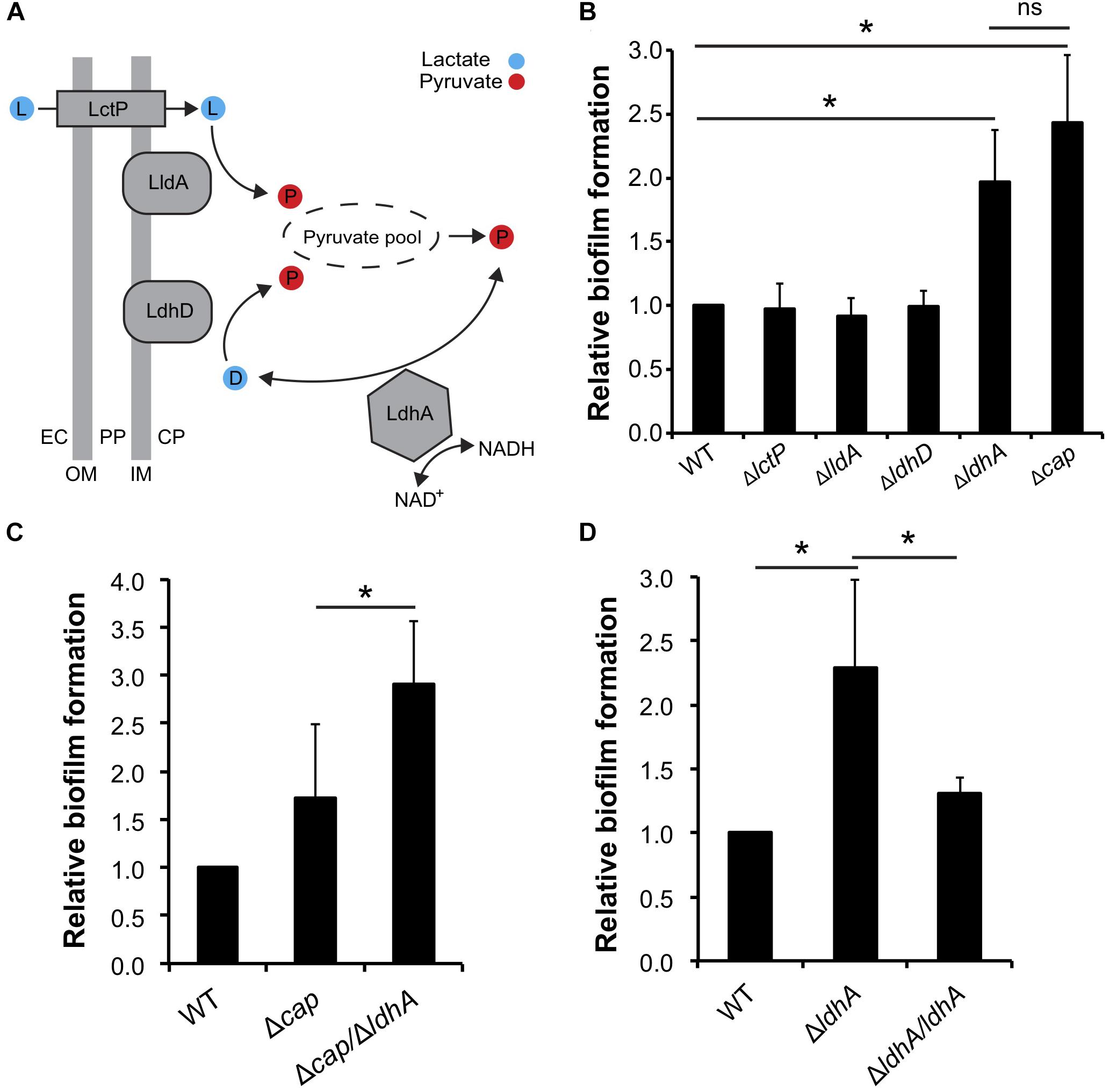
Figure 1. Deletion of ldhA promotes meningococcal biofilm formation in vitro. (A) Predicted role of lactate dehydrogenases (LDHs) in N. meningitidis, adapted from Atack et al. (2014). Abbreviations: LctP, lactate permease; LldA, respiratory L-LDH; LdhD, respiratory D-LDH; LdhA, cytoplasmic NAD+ dependent D-LDH; NAD+, oxidized nicotinamide adenine dinucleotide; NADH, reduced nicotinamide adenine dinucleotide; EC, extracellular space; PP, periplasm; CP, cytoplasm; OM, outer membrane; IM, inner membrane. (B) Biofilm formation by the wild-type, ΔlctP, ΔlldA, ΔldhD, and ΔldhA mutant strains. Δcap was used as a positive control. (C) Biofilm formation by the wild-type, Δcap, and ΔcapA/ΔldhA strains. (D) Biofilm formation by the wild-type, ΔldhA, and ΔldhA/ldhA strains. For all biofilm experiments, bacteria were resuspended in GC liquid supplemented with 1% Kellogg’s supplement to an OD600 of 0.05 and grown under static conditions at 37°C and 5% CO2 for 24 h. The biofilm was washed twice in PBS and then stained with crystal violet. After washing two times with PBS, the biofilm was dissolved with acetic acid and quantified by measuring the absorbance at 630 nm. Experiments were performed at least three times in triplicate. The bars represent the means, with error bars representing the standard deviations. ∗p < 0.05. ns, non-significant.
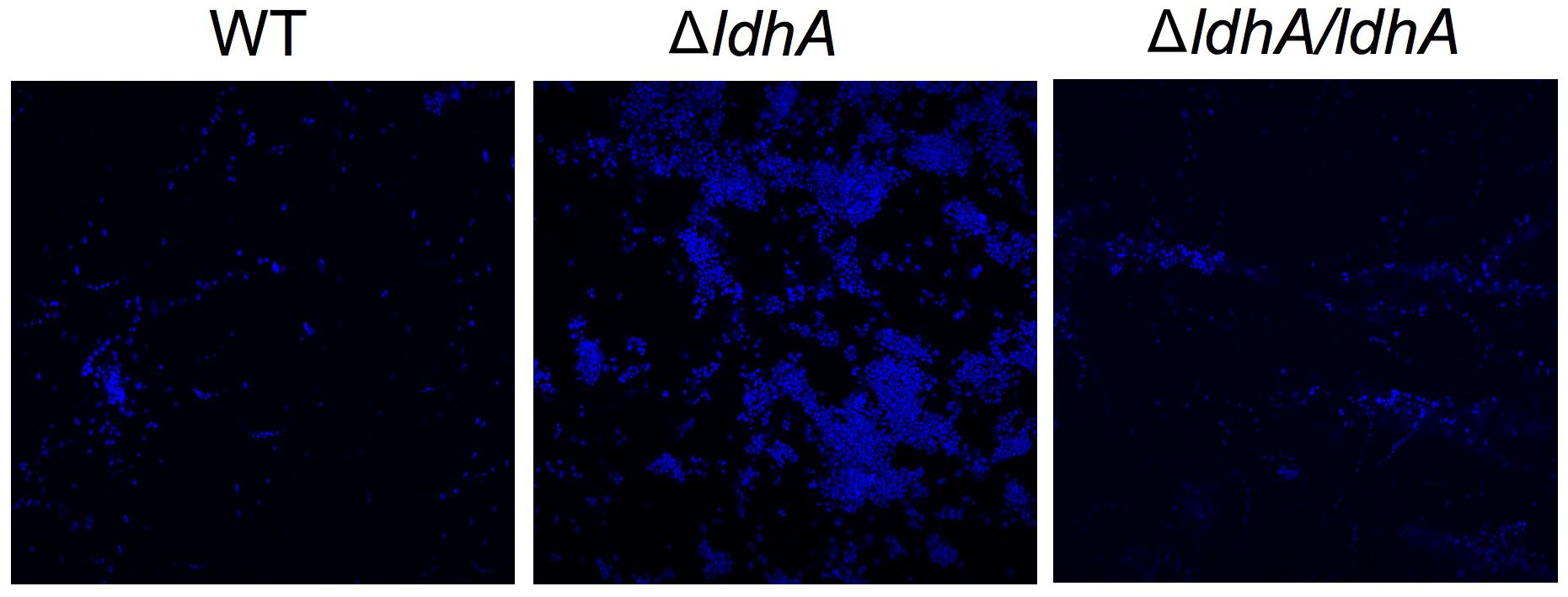
Figure 2. Wild-type, ΔldhA, and ΔldhA/ldhA biofilms visualized by confocal microscopy. Wild-type, ΔldhA, and ΔldhA/ldhA bacteria were resuspended to OD600 of 0.05 and grown under static conditions at 37°C and 5% CO2 for 24 h on glass coverslips. After washing two times with PBS the samples were fixed with 2% formaldehyde and stained with DAPI.
Deletion of ldhA Increases Meningococcal Aggregation and Cell Surface Hydrophobicity
Meningococcal aggregation has been shown to strongly influence biofilm architecture (Yi et al., 2004; Lappann et al., 2006; Arenas et al., 2015). We, therefore, examined whether deletion of ldhA had any effect on bacterial aggregation using a sedimentation assay. Bacteria were grown for 3 h under shaking and were then transferred to static conditions; the absorbance at OD600 of the surface layer was measured every 20 min. The ΔldhA sedimented faster compared to the wild-type and ΔldhA/ldhA strains (Figure 3A). The rate of decline in absorbance was calculated for the first 20 min. The ΔldhA (-0.031 Abs/min) had a significant increase (p < 0.05) in rate compared to that of the wild-type (-0.021 Abs/min) and ΔldhA/ldhA (-0.018 Abs/min) strains. Because pilus expression influences meningococcal aggregation, we performed Western blot analysis to quantify the amount of the major pilus subunit PilE. We did not observe any differences in PilE expression between the wild-type and ΔldhA strains (Figure 3B and Supplementary Figure 3). Also, we did not observe any difference in the level of adhesion to human pharyngeal epithelial FaDu cells for ΔldhA compared to that of the wild-type and ΔldhA/ldhA strains (Figure 3C).
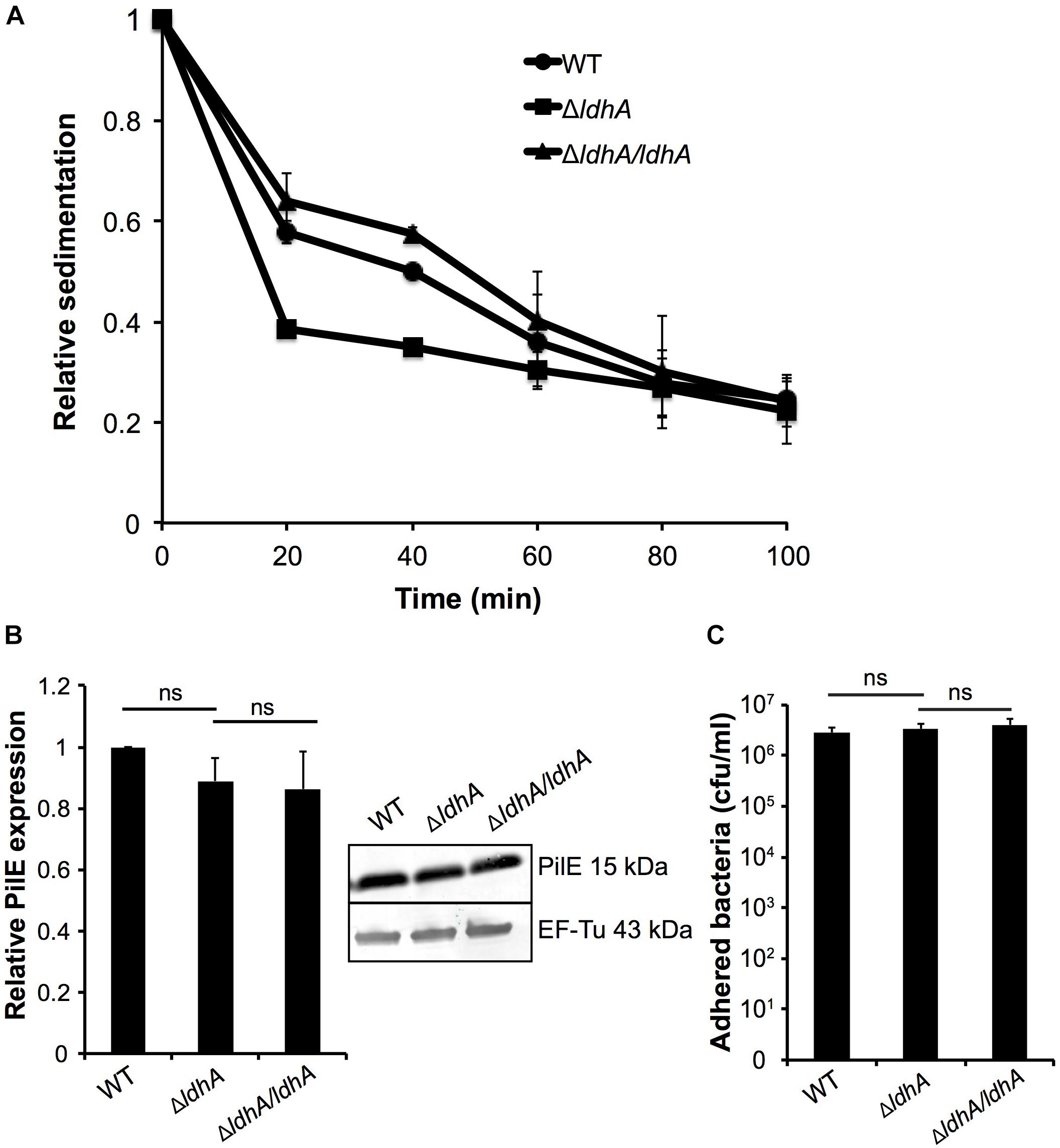
Figure 3. Deletion of ldhA increases meningococcal aggregation. (A) Bacterial aggregation of the wild-type, ΔldhA, and ΔldhA/ldhA strains was examined by the sedimentation assay. Bacteria were grown for 3 h to log phase under shaking conditions at 37°C and 5% CO2 and then moved to static conditions at room temperature. The absorbance (OD600) of the top layer of the culture was measured every 20 min. Data are presented as relative values compared to the 0 min time point (set to 1) for all strains. (B) Western blot analysis and quantification of PilE expression in the wild-type, ΔldhA, and ΔldhA/ldhA strains after 3 h of growth in liquid cultures. After PilE detection, the membrane was stripped, and the level of EF-Tu was examined as a loading control. Data are represented as relative values, and the PilE expression of the wild-type is set to 1. (C) Quantification of the adherence of the wild-type, ΔldhA, and ΔldhA/ldhA strains to the epithelial cell line FaDu. Cells at 100% confluence were infected at an MOI of 100 and incubated for 3 h. All unbound bacteria were removed by washing. Bound bacteria were released by a saponin treatment and quantified by viable counting on GC plates. Experiments were performed at least three times in triplicate. The bars represent the means with error bars representing the standard deviations. ns, non-significant.
As there is a correlation between biofilm formation and hydrophobicity (Yi et al., 2004), we next examined the surface hydrophobicity. We observed elevated cell surface hydrophobicity in ΔldhA compared to wild-type and ΔldhA/ldhA strains (Figure 4A). The capsule-negative mutant (Δcap) was more hydrophobic than the other strains (Figure 4A). To exclude the possibility that the increased ΔldhA surface hydrophobicity was due to reduced capsule expression, we quantified the capsule level. Capsule ELISA revealed no differences in the capsule produced by the ΔldhA mutant compared to that produced by the wild-type strain (Figure 4B). These results indicate that ΔldhA has stronger bacterial aggregation and surface hydrophobicity than the wild-type strain while there was no difference in the level of adhesion to host cells, PilE, and capsule.

Figure 4. Surface hydrophobicity is increased in the ΔldhA, but the capsule level remains constant. (A) A microbial adhesion to solvents (MATS) assay was used to measure the surface hydrophobicity of the wild-type, ΔldhA, and ΔldhA/ldhA strains. The Δcap mutant was used as a control. Bacteria were resuspended in PBS to an OD of 0.4, mixed with hexadecane at a 1:4 ratio and vortexed extensively. After a 15 min incubation, the absorbance (OD600) of the aqueous phase was measured. (B) The level of the polysaccharide capsule was detected with anti-serogroup C capsule antibodies using an indirect ELISA. The plate was coated with a heat-killed bacterial suspension. The absorbance values were normalized against those of the wild-type strain. Experiments were performed at least three times in triplicate. The bars represent the means, with error bars representing the standard deviations. ∗p < 0.05. ns, non-significant.
Increased Aggregation and Biofilm of ΔldhA Is Dependent on Extracellular DNA
Since eDNA is often a major component of the meningococcal extracellular matrix (Lappann et al., 2010) and has been shown to affect bacterial surface hydrophobicity in for example streptococci (Das et al., 2010), we examined whether the increased biofilm formation of ΔldhA was dependent on eDNA. DNase I was added to bacterial solutions before incubation under static conditions for 24 h. The wild-type FAM20 belongs to clonal complex ST-11 strains, which are known to form eDNA-independent biofilms (Lappann et al., 2010). DNase I treatment of the wild-type and Δcap mutant strains did not change the level of biofilm formation (Figure 4A). However, the presence of DNase I significantly (p < 0.05) reduced the biofilm formation of ΔldhA compared to that of those untreated or incubated with heat-inactivated DNase I (Figure 5A). We further show that treatment with DNase did not affect the growth of the wild-type and ΔldhA strains (Supplementary Figure 2). These data indicate that eDNA plays a role in the increased biofilm formation of the ΔldhA mutant.

Figure 5. Increased biofilm formation and aggregation by ΔldhA is associated with an increase in extracellular DNA and autolysis. (A) Biofilm formation by the wild-type, ΔldhA, and Δcap mutant strains in the presence or absence of DNase I. Δcap was used as a positive control. Bacteria were resuspended in GC liquid supplemented with 1% Kellogg’s with or without DNase I to OD600 of 0.05 and grown under static conditions for 24 h. Heat-treated DNase I was used as control (hi–DNase). Washed biofilms were stained with crystal violet and dissolved with acetic acid. The absorbance was measured at 630 nm. The experiment was performed three times in triplicate. (B) Autolysis under non-growth conditions. The wild-type and the ΔldhA strains were resuspended in GC liquid supplemented with 1% Kellogg’s, to an OD600 of 0.05 and grown for 3 h. Bacteria were centrifuged, washed with PBS, and resuspended in 30 mM Tris-HCL buffers at pH 6 or 8 to an OD600 of 1. Absorbance values were acquired every 10 min for the first hour and at 20 min intervals during the second hour. The values were used to calculate the percentage of initial turbidity. One representative experiment, performed in duplicate, out of four is shown. Detection of eDNA in the culture supernatants (C) and in biofilms (D) of the wild-type and ΔldhA strains using a fluorescence-based Quant-IT PicoGreen dsDNA assay kit. Experiments were performed three times in triplicate. (E) Sedimentation of wild-type and ΔldhA aggregates grown in the presence or absence of DNase I. Bacteria were grown for 3 h in the presence or absence of DNase I under shaking conditions at 37°C and 5% CO2 and then moved to static conditions at room temperature. The absorbance (OD600) of the top layer of the culture was measured every 20 min. Data are presented as relative values compared to the 0 min time point (set to 1). Experiments were performed three times. Unless stated, the bars represent the means, with error bars representing the standard deviations. ∗p < 0.05. ns, non-significant.
Because meningococcal eDNA release occurs via cell lysis (Lappann et al., 2010), we next examined autolysis of the ΔldhA and wild-type strains in Tris buffer at pH 6.0 or 8.0. Tris buffer pH at 8.0, but not at pH 6.0, has previously been shown to lead to autolysis in N. gonorrhoeae (Bos et al., 2005; Garcia and Dillard, 2006). Bacteria were resuspended in the buffers, and lysis was quantified by measuring the absorbance every 10 min for 1 h and after that at 20 min intervals. Bacteria were resuspended before every OD measurement to prevent the effects of sedimentation. We detected increased autolysis of ΔldhA compared to that of the wild-type strain at pH 8.0. While Tris buffer at pH 6.0 did not induce autolysis in the wild-type strain, the ΔldhA exhibited autolysis in the pH 6.0 buffer (Figure 5B). To test whether there was a difference in DNA release, we quantified the eDNA from culture supernatants after 3 h of growth. The amount of eDNA measured was higher in ΔldhA culture supernatants and in biofilms compared to that in the wild-type culture supernatants (Figures 5C,D).
These results suggest that an increase in eDNA release mediated by autolysis is the reason for the increased bacterial aggregation and biofilm formation. We, therefore, performed sedimentation assays in the presence or absence of DNase I. We found that the DNase I treatment abolished the enhanced aggregation in ΔldhA, while it did not have an effect on the wild-type strain (Figure 5E). The rate of sedimentation was calculated for the first 20 min. The ΔldhA (-0.030 Abs/min) had a significant (p < 0.05) increase in sedimentation rate compared to that of the wild-type (-0.020 Abs/min) and ΔldhA treated with DNase (-0.022 Abs/min). To summarize, these results indicate that an increase in eDNA release, mediated by autolysis, is the reason for the enhanced aggregation and biofilm formation in ΔldhA mutant.
Differential Expression of Biofilm-Related Genes in Wild-Type and ΔldhA N. meningitidis
To explore whether the deletion of ldhA influences the expression of genes previously linked to biofilm formation, we performed qPCR analyses. Total RNA was isolated from wild-type and ΔldhA mutant harvested from both 3 h log phase shaking cultures and 24 h biofilms. We detected changes in the expression of genes involved in anaerobic respiration. Expression of norB was increased in the ΔldhA mutant both during log phase and biofilm growth. Although the expression of narP was significantly(p < 0.05) induced during log phase, we did not detect within-biofilm changes in the ΔldhA mutant (Figures 6A,B). No significant changes in the expression of aniA were detected (Figures 6A,B).
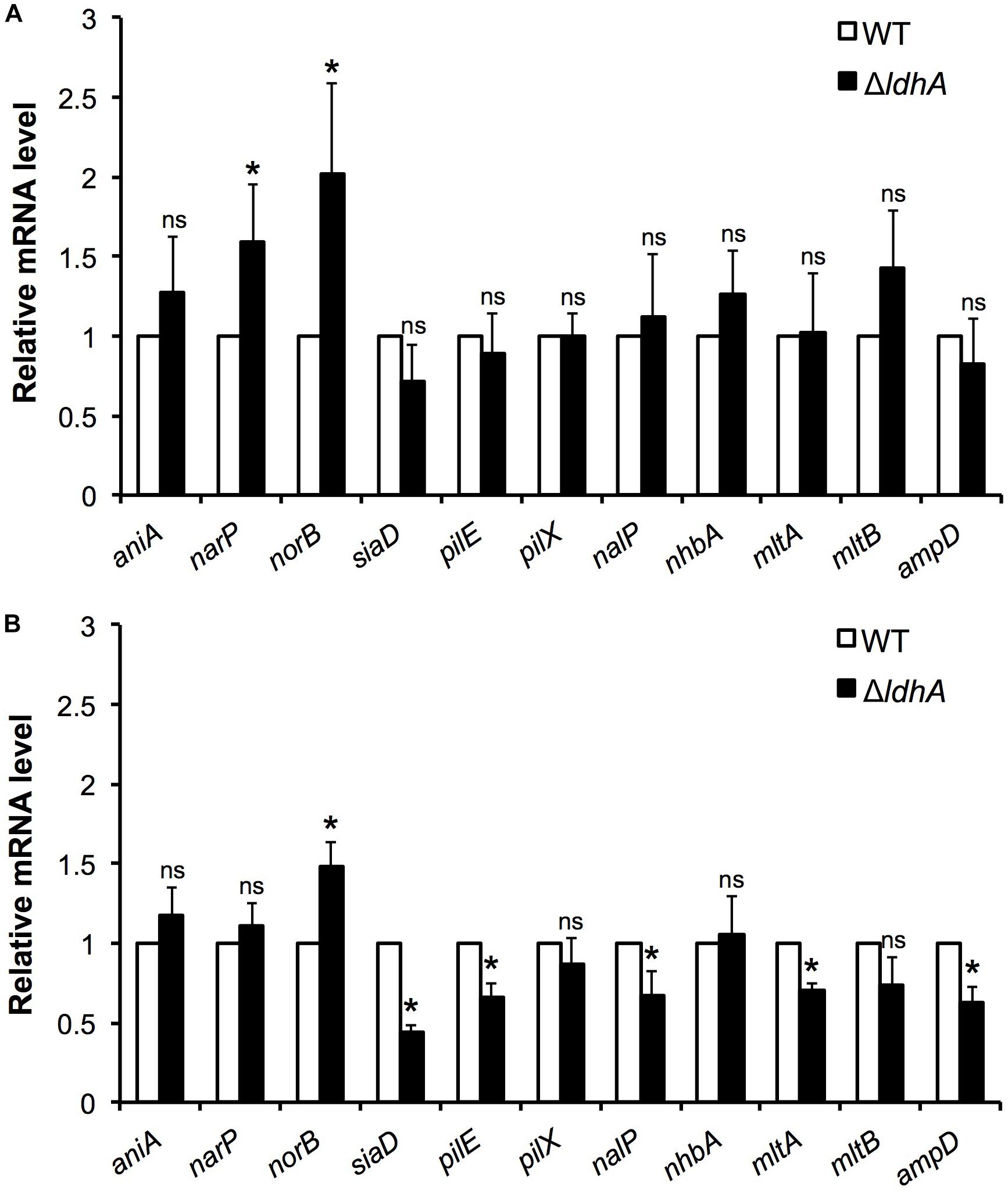
Figure 6. Comparison of gene expression in log phase cultures and biofilms. Gene expression in wild-type and ΔldhA bacterial cultures (A) and biofilms (B). RNA was isolated from bacteria grown to log phase for 3 h and from 24 h old biofilms to generate cDNA to use as a template for qPCR analysis. The gene encoding the ribosomal protein RpsJ was used as a housekeeping gene for data normalization of target mRNA. Experiments were performed four times. The bars represent the means, with error bars representing the standard deviation. ∗p < 0.05. ns, non-significant.
Consistent with the capsule ELISA and PilE detection assay results, we did not find any changes in siaD and pilE expression in log cultures (Figure 6A). However, pilE and siaD were downregulated upon biofilm formation in the ΔldhA mutant (Figure 6B). The expression of nalP and lysis-associated mltA and ampD was downregulated in the ΔldhA mutant during biofilm growth, while no changes were detected in log phase cultures (Figures 6A,B). No changes were identified in the expression of nhbA, mltB or the minor pilin pilX under either condition (Figures 6A,B). To summarize, the expression of pilE, siaD, nalP, mltA, and ampD was downregulated during ΔldhA biofilm formation, while norB, which is involved in anaerobic respiration, was upregulated. Furthermore, the expression of narP and norB was upregulated in the ΔldhA mutant during log phase growth.
Acetate Level Is Reduced in ΔldhA Biofilms but Does Not Influence Biofilm Formation
Accumulation of acetate and acetate intermediates are known to influence biofilm formation of several pathogens (Yang et al., 2005; Sharma-Kuinkel et al., 2009; Chen et al., 2015). We therefore speculated whether acetate played a role in biofilm formation of the ΔldhA mutant. Supernatants were collected from 24 h old biofilms and the amount of acetate was quantified. We detected significant decrease in the amount of acetate in supernatants from ΔldhA biofilms compared to that of the wild-type (Figure 7A). This indicated that reduction in the acetate levels might play a role in the increased biofilm formation by ΔldhA mutant. To explore this possibility, we performed assays to measure biofilm formation by wild-type and ΔldhA mutant in presence of 0.1, 1, and 10 mM sodium acetate. We did not detect significant reduction in biofilm formation by ΔldhA mutant in presence of acetate (Figure 7B). This indicates that although there is a reduction in the level of acetate in ΔldhA biofilm supernatants it does not play a role in the increase in biofilm formation.

Figure 7. Acetate level is reduced in ΔldhA biofilms but does not influence biofilm formation. (A) Quantification of acetate level in biofilm supernatants. Culture medium was collected from wild-type and ΔldhA biofilms and concentration of acetate was quantified using Acetate Colorimetric Assay Kit. Experiment was performed three times in duplicate. (B) Biofilm formation by the wild-type and ΔldhA in the presence of 0.1, 1 and 10 mM sodium acetate. Experiment was performed two times in triplicate. The bars represent the means, with error bars representing the standard deviation. ∗p < 0.05. ns, non-significant.
Discussion
In this study, we examined the role of lactate metabolism in meningococcal biofilm formation in vitro. We provide evidence that a D-lactate dehydrogenase, LdhA, has a negative impact on N. meningitidis biofilm formation. Loss of ldhA increased the level of bacterial aggregation and biofilm formation. This effect was associated with an increased autolytic rate and the amount of eDNA released.
Deletion of ldhA increased the level of biofilm formation. In meningococci, there is a direct relationship between surface hydrophobicity and biofilm formation (Yi et al., 2004). Our results show that the ΔldhA mutant has increased surface hydrophobicity compared to that of the wild-type. Because the capsule affects bacterial hydrophobicity, we quantified the level of capsule. However, there was no difference in the amount of capsule between the wild-type and ΔldhA mutant strains.
Treatment by DNase I abolished the enhanced aggregation and biofilm formation by ΔldhA, suggesting that eDNA plays a role. It is well-known that eDNA plays a major role in biofilm formation in N. meningitidis and other pathogens that colonize the upper respiratory tract, such as Pseudomonas aeruginosa, Streptococcus pneumoniae, and Haemophilus influenza (Whitchurch et al., 2002; Jurcisek and Bakaletz, 2007; Hall-Stoodley et al., 2008; Lappann et al., 2010). The biofilm formation and architecture of meningococci exhibit strain-dependent variations. Biofilm formation by frequently carried meningococcal strains is dependent on eDNA, while hypervirulent strains of clonal complexes (cc) ST-8 and ST-11 do not depend on eDNA (Lappann et al., 2010). The level of biofilm formed by the wild-type FAM20 was not affected by the treatment of DNase I in our studies. Also, the treatment did not have an impact on Δcap biofilm formation. This result is in agreement with the theory that strains such as FAM20, which belong to ST-11 cc, do not depend on eDNA. It is noteworthy that eDNA on streptococcal cell surfaces can increase hydrophobicity (Das et al., 2010). However, eDNA is also known to increase bacterial adhesion to surfaces (Rice et al., 2007; Das et al., 2010; Regina et al., 2014). Additionally, an enhanced aggregation can play a major role in bacterial adhesion to host cells (Mikaty et al., 2009; Kuwae et al., 2011; Engman et al., 2016). However, we did not detect differences in adhesion between the wild-type and ΔldhA strains. The roles of eDNA in meningococcal adhesion and aggregation have not been well-studied, although its role is well established in biofilm formation (Lappann et al., 2010; Pérez-Ortega et al., 2017).
There is a link between eDNA-dependent biofilm formation and autolysis in meningococci (Lappann et al., 2010; Arenas and Tommassen, 2017). The ΔldhA mutant exhibited an increased rate of autolysis, although the underlying mechanisms remain to be elucidated. Meningococcal autolysis can occur through the lytic transglycosylases MltA and MltB and the N-acetylmuramyl-L-alanine amidase AmpD (Lappann et al., 2010). Also, OMPLA is an important inducer of autolysis in meningococci during the stationary phase (Bos et al., 2005). DNA released during OMPLA-mediated autolysis plays a role in meningococcal biofilms and the ability to withstand shear forces and, therefore, contributes to long-term colonization (Lappann et al., 2010). However, ST-8 and ST-11 cc, including FAM20, lack a functional gene encoding OMPLA (Bos et al., 2005). Surprisingly, via RT-qPCR analysis, we detected reduced expression of mltA and ampD, which encode the known meningococcal autolysins in ΔldhA biofilms. Studies have also shown that environmental factors, including pH, growth phase, and NaCl, can be responsible for bacterial autolysis (reviewed in Rice and Bayles, 2008).
We detected downregulation of the capsule biosynthesis gene siaD during ΔldhA biofilm formation. Earlier studies have shown reductions in the transcripts of the capsule biosynthesis genes during biofilm formation (O’Dwyer et al., 2009). However, Neil et al. (2014) have reported biofilm formation by encapsulated meningococci upon adhesion to epithelial cells, with no reduction in capsule observed. Surprisingly, we detected downregulation of pilE in ΔldhA within a biofilm. Deletion of pilE is known to abolish microcolony formation and reduce biofilm thickness, although these results were shown in a strain expressing class I Tfp (Lappann et al., 2006). FAM20 and all strains that belong to ST-8 and ST-11 cc form eDNA-independent biofilms and, express class II Tfp (Cehovin et al., 2010; Wormann et al., 2014). The Tfp is nonetheless important for the establishment of firm adhesion in the presence of mechanical forces (Mikaty et al., 2009). In our assays, we only evaluated the level of biofilm formation in a static environment. However, we did not detect any changes in pilE expression during the log growth phase. The attachment could, therefore, be established at earlier time points, and the pilE downregulation could occur upon biofilm mode of growth.
Reduced expression of nalP was detected during ΔldhA biofilm growth. As mentioned earlier, the autotransporter NalP can influence the level of biofilm formation by cleaving cell-surface eDNA-binding molecules (Arenas et al., 2013). A decrease in the NalP level could potentially contribute to the increased eDNA-binding during biofilm formation in the ΔldhA mutant. In the future, it would be interesting to study the role of NalP in increased biofilm formation in the ΔldhA mutant. A decrease in amount of NalP has been shown to correlate with increased meningococcal biofilm formation, however this might vary depending on the amount of eDNA present (Pérez-Ortega et al., 2017). We did not, however, detect any changes in nalP expression during log phase growth. The autotransporters AutA and AutB have also been linked to meningococcal biofilm formation (Arenas et al., 2015, 2016). However, since the autA gene in FAM20 (FAM18 derivative) is disrupted and the autB is out of frame they were excluded from the study (Peak et al., 1999; Ait-Tahar et al., 2000; van Ulsen et al., 2001).
Adaptations to oxygen limitations are important during the development of biofilms. The expression of norB, which is involved in anaerobic respiration, was induced in the ΔldhA mutant both during log phase and biofilm growth. It has been reported that the expression of norB, which encodes the nitric oxide reductase NorB, increases in the presence of nitric oxide (Householder et al., 2000; Rock et al., 2007). NorB is important for growth in oxygen-deprived environments, and a lack of its activity has a drastic impact on biofilm formation in N. gonorrhoeae (Householder et al., 2000; Falsetta et al., 2009). Additionally, we detected upregulation of narP in ΔldhA log phase cultures. NarP-deficiency reduces biofilm formation in meningococci (Jamet et al., 2013). However, we did not detect any differences in narP expression during biofilm growth in the ΔldhA mutant.
The strain used in this study was of clonal complex ST-11, which is known to form poor biofilms independent of eDNA. It would be of interest to compare the expression of ldhA during growth in both carrier and virulent strains. However, it is highly unlikely that LdhA is directly involved in biofilm formation.
It is tempting to speculate that a metabolic shift due to the lack of ldhA rather than a direct effect is the cause of the observed phenotype. There is accumulating evidence that there is a correlation between energy metabolism and biofilm formation in pathogenic Neisseria. Both transcriptomic and proteomic analyses have revealed changes in metabolic enzymes that are important in pyruvate metabolism, glycolysis/gluconeogenesis, and the citric acid cycle during the biofilm mode of growth (Falsetta et al., 2009; O’Dwyer et al., 2009; van Alen et al., 2010; Phillips et al., 2012). In Neisseria, LdhA can both serve as D-LDH, converting D-lactate to pyruvate, and reversibly convert pyruvate to D-lactate as an NADH-pyruvate reductase. The metabolic shift upon ldhA deletion in Neisseria has not been studied extensively. However, it has been shown that D-lactate production is abolished (Atack et al., 2014). Deletion of ldhA in other pathogens, such as E. coli and S. pneumoniae, also results in the inability to produce D-lactate and an increase in acetate, pyruvate, and ethanol production from glucose. Acetate intermediates have been shown to play a role as metabolic signaling molecules in bacterial biofilms. In E. coli, the deletion of ldhA leads to the accumulation of acetyl-CoA, an intermediate in acetate metabolism, and increases the level of biofilm formation. Acetic acid has been shown to stimulate biofilm formation in Staphylococcus aureus and Bacillus subtilis by activating the expression of genes that encode proteins that are important in bacterial autolysis (Yang et al., 2005; Sharma-Kuinkel et al., 2009; Chen et al., 2015). To examine the possible role of acetate in ΔldhA biofilm formation we measured the concentration of acetate in supernatants collected from biofilms. We detected significant reduction in acetate concentration. However when acetate level was raised we were not able to significantly reduce the amount of biofilm formed by the ΔldhA mutant.
Conclusion
In conclusion, we showed that a lack of ldhA in N. meningitidis increases surface hydrophobicity, bacterial aggregation, and biofilm formation. Based on our results, we suggest that the promoted aggregation and biofilm formation in the ΔldhA mutant is dependent on an increase in autolysis-mediated eDNA release. It is not currently known whether ldhA mutation frequently occurs during colonization, and it would be interesting in the future to investigate ldhA expression during in vivo mimicking conditions, and further elucidate its possible role upon biofilm formation and meningococcal pathogenesis.
Data Availability
All datasets generated for this study are included in the manuscript and/or the Supplementary Files.
Author Contributions
SS, GW, FZ, and A-BJ conceived and designed the experiments, analyzed the data, and wrote the manuscript. SS, GW, FZ, and MA performed the experiments.
Funding
This work was supported by the Swedish Research Council (Dnr 2013-2434, 2016-1279) and the Swedish Cancer Society.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Dr. Sunil Saroj for careful reading and providing useful comments to improve the manuscript, and Dr. Stina Höglund for help with confocal microscopy.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2019.00422/full#supplementary-material
References
Ait-Tahar, K., Wooldridge, K. G., Turner, D. P., Atta, M., Todd, I., and Ala’Aldeen, D. A. (2000). Auto-transporter A protein of Neisseria meningitidis: a potent CD4+ T-cell and B-cell stimulating antigen detected by expression cloning. Mol. Microbiol. 37, 1094–1105. doi: 10.1046/j.1365-2958.2000.02061.x
Ala’Aldeen, D. A., Neal, K. R., Ait-Tahar, K., Nguyen-Van-Tam, J. S., English, A., Falla, T. J., et al. (2000). Dynamics of meningococcal long-term carriage among university students and their implications for mass vaccination. J. Clin. Microbiol. 38, 2311–2316.
Arenas, J., Cano, S., Nijland, R., van Dongen, V., Rutten, L., van der Ende, A., et al. (2015). The meningococcal autotransporter AutA is implicated in autoaggregation and biofilm formation. Environ. Microbiol. 17, 1321–1337. doi: 10.1111/1462-2920.12581
Arenas, J., Nijland, R., Rodriguez, F. J., Bosma, T. N. P., and Tommassen, J. (2013). Involvement of three meningococcal surface exposed proteins, the heparin binding protein NhbA, the peptide of IgA protease and the autotransporter protease NalP, in initiation of biofilm formation. Mol. Microbiol. 87, 254–268. doi: 10.1111/mmi.12097
Arenas, J., Paganelli, F. L., Rodríguez-Castaño, P., Cano-Crespo, S., van der Ende, A., van Putten, J. P., et al. (2016). Expression of the gene for autotransporter AutB of Neisseria meningitidis affects biofilm formation and epithelial transmigration. Front. Cell. Infect. Microbiol. 6:162. doi: 10.3389/fcimb.2016.00162
Arenas, J., and Tommassen, J. (2017). Meningococcal biofilm formation: let’s stick together. Trends Microbiol. 25, 113–124. doi: 10.1016/j.tim.2016.09.005
Atack, J. M., Ibranovic, I., Ong, C.-L. Y. L., Djoko, K. Y., Chen, N. H., Vanden Hoven, R., et al. (2014). A role for lactate dehydrogenases in the survival of Neisseria gonorrhoeae in human polymorphonuclear leukocytes and cervical epithelial cells. J. Infect. Dis. 210, 1311–1318. doi: 10.1093/infdis/jiu230
Barraud, N., Hassett, D. J., Hwang, S. H., Rice, S. A., Kjelleberg, S., and Webb, J. S. (2006). Involvement of nitric oxide in biofilm dispersal of Pseudomonas aeruginosa. J. Bacteriol. 188, 7344–7353. doi: 10.1128/JB.00779-06
Barth, K. R., Isabella, V. M., and Clark, V. L. (2009). Biochemical and genomic analysis of the denitrification pathway within the genus Neisseria. Microbiology 155(Pt 12), 4093–4103. doi: 10.1099/mic.0.032961-0
Bartolini, E., Frigimelica, E., Giovinazzi, S., Galli, G., Shaik, Y., Genco, C., et al. (2006). Role of FNR and FNR-regulated, sugar fermentation genes in Neisseria meningitidis infection. Mol. Microbiol. 60, 963–972. doi: 10.1111/j.1365-2958.2006.05163.x
Bos, M. P., Tefsen, B., Voet, P., Weynants, V., van Putten, J. P., and Tommassen, J. (2005). Function of neisserial outer membrane phospholipase a in autolysis and assessment of its vaccine potential. Infect. Immun. 73, 2222–2231. doi: 10.1128/IAI.73.4.2222-2231.2005
Cehovin, A., Winterbotham, M., Lucidarme, J., Borrow, R., Tang, C. M., Exley, R. M., et al. (2010). Sequence conservation of pilus subunits in Neisseria meningitidis. Vaccine 28, 4817–4826. doi: 10.1016/j.vaccine.2010.04.065
Chen, Y., Gozzi, K., Yan, F., and Chai, Y. (2015). Acetic acid acts as a volatile signal to stimulate bacterial biofilm formation. mBio 6:e00392. doi: 10.1128/mBio.00392-15
Costerton, J. W., Lewandowski, Z., Caldwell, D. E., Korber, D. R., and Lappin-Scott, H. M. (1995). Microbial biofilms. Annu. Rev. Microbiol. 49, 711–745. doi: 10.1146/annurev.mi.49.100195.003431
Craig, L., Pique, M. E., and Tainer, J. A. (2004). Type IV pilus structure and bacterial pathogenicity. Nat. Rev. Microbiol. 2, 363–378. doi: 10.1038/nrmicro885
Das, T., Sharma, P. K., Busscher, H. J., van der Mei, H. C., and Krom, B. P. (2010). Role of extracellular DNA in initial bacterial adhesion and surface aggregation. Appl. Environ. Microbiol. 76, 3405–3408. doi: 10.1128/AEM.03119-09
de Vries, F. P., van Der Ende, A., van Putten, J. P., and Dankert, J. (1996). Invasion of primary nasopharyngeal epithelial cells by Neisseria meningitidis is controlled by phase variation of multiple surface antigens. Infect. Immun. 64, 2998–3006.
Deghmane, A. E., Giorgini, D., Larribe, M., Alonso, J. M., and Taha, M. K. (2002). Down-regulation of pili and capsule of Neisseria meningitidis upon contact with epithelial cells is mediated by CrgA regulatory protein. Mol. Microbiol. 43, 1555–1564. doi: 10.1046/j.1365-2958.2002.02838.x
Engman, J., Negrea, A., Sigurlasdottir, S., Georg, M., Eriksson, J., Eriksson, O. S., et al. (2016). Neisseria meningitidis polynucleotide phosphorylase affects aggregation, adhesion, and virulence. Infect. Immun. 84, 1501–1513. doi: 10.1128/IAI.01463-15
Exley, R. M., Shaw, J., Mowe, E., Sun, Y.-H. H., West, N. P., Williamson, M., et al. (2005). Available carbon source influences the resistance of Neisseria meningitidis against complement. J. Exp. Med. 201, 1637–1645. doi: 10.1084/jem.20041548
Exley, R. M., Wu, H., Shaw, J., Schneider, M. C., Smith, H., Jerse, A. E., et al. (2007). Lactate acquisition promotes successful colonization of the murine genital tract by Neisseria gonorrhoeae. Infect. Immun. 75, 1318–1324. doi: 10.1128/IAI.01530-06
Falsetta, M. L., Bair, T. B., Ku, S. C., Vanden Hoven, R. N., Steichen, C. T., McEwan, A. G., et al. (2009). Transcriptional profiling identifies the metabolic phenotype of gonococcal biofilms. Infect. Immun. 77, 3522–3532. doi: 10.1128/IAI.00036-09
Fischer, R. S., Martin, G. C., Rao, P., and Jensen, R. A. (1994). Neisseria gonorrhoeae possesses two nicotinamide adenine dinucleotide-independent lactate dehydrogenases. FEMS Microbiol. Lett. 115, 39–44. doi: 10.1111/j.1574-6968.1994.tb06611.x
Garcia, D. L., and Dillard, J. P. (2006). AmiC functions as an N-acetylmuramyl-l-alanine amidase necessary for cell separation and can promote autolysis in Neisseria gonorrhoeae. J. Bacteriol. 188, 7211–7221. doi: 10.1128/JB.00724-06
Ge, X., Shi, X., Shi, L., Liu, J., Stone, V., Kong, F., et al. (2016). Involvement of NADH oxidase in biofilm formation in Streptococcus sanguinis. PLoS One 11:e0151142. doi: 10.1371/journal.pone.0151142
Hall-Stoodley, L., Nistico, L., Sambanthamoorthy, K., Dice, B., Nguyen, D., Mershon, W. J., et al. (2008). Characterization of biofilm matrix, degradation by DNase treatment and evidence of capsule downregulation in Streptococcus pneumoniae clinical isolates. BMC Microbiol. 8:173. doi: 10.1186/1471-2180-8-173
Hamilton, H. L., Domínguez, N. M., Schwartz, K. J., Hackett, K. T., and Dillard, J. P. (2005). Neisseria gonorrhoeae secretes chromosomal DNA via a novel type IV secretion system. Mol. Microbiol. 55, 1704–1721. doi: 10.1111/j.1365-2958.2005.04521.x
Harrison, O. B., Robertson, B. D., Faust, S. N., Jepson, M. A., Goldin, R. D., Levin, M., et al. (2002). Analysis of pathogen-host cell interactions in purpura fulminans: expression of capsule, type IV pili, and PorA by Neisseria meningitidis in vivo. Infect. Immun. 70, 5193–5201. doi: 10.1128/IAI.70.9.5193-5201.2002
Helaine, S., Carbonnelle, E., Prouvensier, L., Beretti, J. L., Nassif, X., and Pelicic, V. (2005). PilX, a pilus-associated protein essential for bacterial aggregation, is a key to pilus-facilitated attachment of Neisseria meningitidis to human cells. Mol. Microbiol. 55, 65–77. doi: 10.1111/j.1365-2958.2004.04372.x
Householder, T. C., Belli, W. A., Lissenden, S., Cole, J. A., and Clark, V. L. (1999). cis- and trans-acting elements involved in regulation of aniA, the gene encoding the major anaerobically induced outer membrane protein in Neisseria gonorrhoeae. J. Bacteriol. 181, 541–551.
Householder, T. C., Fozo, E. M., Cardinale, J. A., and Clark, V. L. (2000). Gonococcal nitric oxide reductase is encoded by a single gene, norB, which is required for anaerobic growth and is induced by nitric oxide. Infect. Immun. 68, 5241–5246. doi: 10.1128/IAI.68.9.5241-5246.2000
Jamet, A., Euphrasie, D., Martin, P., and Nassif, X. (2013). Identification of genes involved in Neisseria meningitidis colonization. Infect. Immun. 81, 3375–3381. doi: 10.1128/IAI.00421-13
Jones, A., Georg, M., Maudsdotter, L., and Jonsson, A. B. (2009). Endotoxin, capsule, and bacterial attachment contribute to Neisseria meningitidis resistance to the human antimicrobial peptide LL-37. J. Bacteriol. 191, 3861–3868. doi: 10.1128/JB.01313-08
Jurcisek, J. A., and Bakaletz, L. O. (2007). Biofilms formed by nontypeable Haemophilus influenzae in vivo contain both double-stranded DNA and type IV pilin protein. J. Bacteriol. 189, 3868–3875. doi: 10.1128/JB.01935-06
Keogh, D., Lam, L. N., Doyle, L. E., Matysik, A., Pavagadhi, S., Umashankar, S., et al. (2018). Extracellular electron transfer powers enterococcus faecalis biofilm metabolism. mBio 9, e00626-17. doi: 10.1128/mBio.00626-17
Kuwae, A., Sjolinder, H., Eriksson, J., Eriksson, S., Chen, Y., and Jonsson, A. B. (2011). NafA negatively controls Neisseria meningitidis piliation. PLoS One 6:e21749. doi: 10.1371/journal.pone.0021749
Lappann, M., Claus, H., van Alen, T., Harmsen, M., Elias, J., Molin, S., et al. (2010). A dual role of extracellular DNA during biofilm formation of Neisseria meningitidis. Mol. Microbiol. 75, 1355–1371. doi: 10.1111/j.1365-2958.2010.07054.x
Lappann, M., Haagensen, J. A., Claus, H., Vogel, U., and Molin, S. (2006). Meningococcal biofilm formation: structure, development and phenotypes in a standardized continuous flow system. Mol. Microbiol. 62, 1292–1309. doi: 10.1111/j.1365-2958.2006.05448.x
Lappann, M., and Vogel, U. (2010). Biofilm formation by the human pathogen Neisseria meningitidis. Med. Microbiol. Immunol. 199, 173–183. doi: 10.1007/s00430-010-0149-y
Ly, M. H., Vo, N. H., Le, T. M., Belin, J. M., and Wache, Y. (2006). Diversity of the surface properties of Lactococci and consequences on adhesion to food components. Colloids Surf. B Biointerfaces 52, 149–153. doi: 10.1016/j.colsurfb.2006.04.015
Mikaty, G., Soyer, M., Mairey, E., Henry, N., Dyer, D., Forest, K. T., et al. (2009). Extracellular bacterial pathogen induces host cell surface reorganization to resist shear stress. PLoS Pathog. 5:e1000314. doi: 10.1371/journal.ppat.1000314
Neil, R. B., Shao, J. Q., and Apicella, M. A. (2009). Biofilm formation on human airway epithelia by encapsulated Neisseria meningitidis serogroup B. Microbes Infect. 11, 281–287. doi: 10.1016/j.micinf.2008.12.001
O’Dwyer, C. A. A., Li, M.-S. S., Langford, P. R., and Kroll, J. S. (2009). Meningococcal biofilm growth on an abiotic surface - a model for epithelial colonization? Microbiology 155(Pt 6), 1940–1952. doi: 10.1099/mic.0.026559-0
Pace, D., and Pollard, A. J. (2012). Meningococcal disease: clinical presentation and sequelae. Vaccine 30(Suppl. 2), B3–B9. doi: 10.1016/j.vaccine.2011.12.062
Peak, I. R., Jennings, M. P., Hood, D. W., and Moxon, E. R. (1999). Tetranucleotide repeats identify novel virulence determinant homologues in Neisseria meningitidis. Microb. Pathog. 26, 13–23. doi: 10.1006/mpat.1998.0243
Pérez-Ortega, J., Rodríguez, A., Ribes, E., Tommassen, J., and Arenas, J. (2017). Interstrain cooperation in meningococcal biofilms: role of autotransporters NalP and AutA. Front. Microbiol. 8:434. doi: 10.3389/fmicb.2017.00434
Phillips, N. J., Steichen, C. T., Schilling, B., Post, D. M., Niles, R. K., Bair, T. B., et al. (2012). Proteomic analysis of Neisseria gonorrhoeae biofilms shows shift to anaerobic respiration and changes in nutrient transport and outermembrane proteins. PLoS One 7:e38303. doi: 10.1371/journal.pone.0038303
Regina, V. R., Lokanathan, A. R., Modrzyñski, J. J., Sutherland, D. S., and Meyer, R. L. (2014). Surface physicochemistry and ionic strength affects eDNA’s role in bacterial adhesion to abiotic surfaces. PLoS One 9:e105033. doi: 10.1371/journal.pone.0105033
Rice, K. C., and Bayles, K. W. (2008). Molecular control of bacterial death and lysis. Microbiol. Mol. Biol. Rev. 72:85. doi: 10.1128/MMBR.00030-07
Rice, K. C., Mann, E. E., Endres, J. L., Weiss, E. C., Cassat, J. E., Smeltzer, M. S., et al. (2007). The cidA murein hydrolase regulator contributes to DNA release and biofilm development in Staphylococcus aureus. Proc. Natl. Acad. Sci. U.S.A. 104, 8113–8118. doi: 10.1073/pnas.0610226104
Rock, J. D., Mahnane, M. R., Anjum, M. F., Shaw, J. G., Read, R. C., and Moir, J. W. (2005). The pathogen Neisseria meningitidis requires oxygen, but supplements growth by denitrification. Nitrite, nitric oxide and oxygen control respiratory flux at genetic and metabolic levels. Mol. Microbiol. 58, 800–809. doi: 10.1111/j.1365-2958.2005.04866.x
Rock, J. D., Thomson, M. J., Read, R. C., and Moir, J. W. (2007). Regulation of denitrification genes in Neisseria meningitidis by nitric oxide and the repressor NsrR. J. Bacteriol. 189, 1138–1144. doi: 10.1128/JB.01368-06
Sharma-Kuinkel, B. K., Mann, E. E., Ahn, J. S., Kuechenmeister, L. J., Dunman, P. M., and Bayles, K. W. (2009). The Staphylococcus aureus LytSR two-component regulatory system affects biofilm formation. J. Bacteriol. 191, 4767–4775. doi: 10.1128/JB.00348-09
Sigurlasdottir, S., Engman, J., Eriksson, O. S., Saroj, S. D., Zguna, N., Lloris-Garcera, P., et al. (2017). Host cell-derived lactate functions as an effector molecule in Neisseria meningitidis microcolony dispersal. PLoS Pathog. 13:e1006251. doi: 10.1371/journal.ppat.1006251
Silva, I. N. N., Ramires, M. J., Azevedo, L. A., Guerreiro, A. R., Tavares, A. C., Becker, J. D. D., et al. (2017). Regulator LdhR and d-Lactate dehydrogenase LdhA of burkholderia multivorans play roles in carbon overflow and in planktonic cellular aggregate formation. Appl. Environ. Microbiol. 83:e1343-17. doi: 10.1128/AEM.01343-17
Sim, R. J., Harrison, M. M., Moxon, E. R., and Tang, C. M. (2000). Underestimation of meningococci in tonsillar tissue by nasopharyngeal swabbing. Lancet 356, 1653–1654. doi: 10.1016/S0140-6736(00)03162-7
Sjolinder, H., and Jonsson, A. B. (2007). Imaging of disease dynamics during meningococcal sepsis. PLoS One 2:e241. doi: 10.1371/journal.pone.0000241
Stephens, D. S., Greenwood, B., and Brandtzaeg, P. (2007). Epidemic meningitis, meningococcaemia, and Neisseria meningitidis. Lancet 369, 2196–2210. doi: 10.1016/S0140-6736(07)61016-2
Tang, L., Schramm, A., Neu, T. R., Revsbech, N. P., and Meyer, R. L. (2013). Extracellular DNA in adhesion and biofilm formation of four environmental isolates: a quantitative study. FEMS Microbiol. Ecol. 86, 394–403. doi: 10.1111/1574-6941.12168
Tzeng, Y. L., Bazan, J. A., Turner, A. N., Wang, X., Retchless, A. C., Read, T. D., et al. (2017). Emergence of a new Neisseria meningitidis clonal complex 11 lineage 11.2 clade as an effective urogenital pathogen. Proc. Natl. Acad. Sci. U.S.A. 114, 4237–4242. doi: 10.1073/pnas.1620971114
Tzeng, Y.-L. L., Kahler, C. M., Zhang, X., and Stephens, D. S. (2008). MisR/MisS two-component regulon in Neisseria meningitidis. Infect. Immun. 76, 704–716. doi: 10.1128/IAI.01007-07
van Alen, T., Claus, H., Zahedi, R. P. P., Groh, J., Blazyca, H., Lappann, M., et al. (2010). Comparative proteomic analysis of biofilm and planktonic cells of Neisseria meningitidis. Proteomics 10, 4512–4521. doi: 10.1002/pmic.201000267
van Ulsen, P., van Alphen, L., Hopman, C. T., van der Ende, A., and Tommassen, J. (2001). In vivo expression of Neisseria meningitidis proteins homologous to the Haemophilus influenzae Hap and Hia autotransporters. FEMS Immunol. Med. Microbiol. 32, 53–64. doi: 10.1111/j.1574-695X.2001.tb00534.x
Werner, E., Roe, F., Bugnicourt, A., Franklin, M. J., Heydorn, A., Molin, S., et al. (2004). Stratified growth in Pseudomonas aeruginosa biofilms. Appl. Environ. Microbiol. 70, 6188–6196. doi: 10.1128/AEM.70.10.6188-6196.2004
Whitchurch, C. B., Tolker-Nielsen, T., Ragas, P. C., and Mattick, J. S. (2002). Extracellular DNA required for bacterial biofilm formation. Science 295:1487. doi: 10.1126/science.295.5559.1487
Wilder-Smith, A., Barkham, T. M., Ravindran, S., Earnest, A., and Paton, N. I. (2003). Persistence of W135 Neisseria meningitidis carriage in returning Hajj pilgrims: risk for early and late transmission to household contacts. Emerg. Infect. Dis. 9, 123–126. doi: 10.3201/eid0901.020131
Wolfgang, M., Lauer, P., Park, H. S., Brossay, L., Hébert, J., and Koomey, M. (1998). PilT mutations lead to simultaneous defects in competence for natural transformation and twitching motility in piliated Neisseria gonorrhoeae. Mol. Microbiol. 29, 321–330. doi: 10.1046/j.1365-2958.1998.00935.x
Wormann, M. E., Horien, C. L., Bennett, J. S., Jolley, K. A., Maiden, M. C., Tang, C. M., et al. (2014). Sequence, distribution and chromosomal context of class I and class II pilin genes of Neisseria meningitidis identified in whole genome sequences. BMC Genomics 15:253. doi: 10.1186/1471-2164-15-253
Yang, S.-J. J., Rice, K. C., Brown, R. J., Patton, T. G., Liou, L. E., Park, Y. H., et al. (2005). A LysR-type regulator, CidR, is required for induction of the Staphylococcus aureus cidABC operon. J. Bacteriol. 187, 5893–5900. doi: 10.1128/JB.187.17.5893-5900.2005
Keywords: Neisseria meningitidis, lactate dehydrogenase, eDNA, autolysis, biofilm
Citation: Sigurlásdóttir S, Wassing GM, Zuo F, Arts M and Jonsson A-B (2019) Deletion of D-Lactate Dehydrogenase A in Neisseria meningitidis Promotes Biofilm Formation Through Increased Autolysis and Extracellular DNA Release. Front. Microbiol. 10:422. doi: 10.3389/fmicb.2019.00422
Received: 12 November 2018; Accepted: 18 February 2019;
Published: 05 March 2019.
Edited by:
Satoshi Tsuneda, Waseda University, JapanReviewed by:
Charlene Kahler, University of Western Australia, AustraliaJesús Arenas, Utrecht University, Netherlands
Copyright © 2019 Sigurlásdóttir, Wassing, Zuo, Arts and Jonsson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ann-Beth Jonsson, ann-beth.jonsson@su.se