
Immunomodulation of IL-33 and IL-37 with Vitamin D in the Neointima of Coronary Artery: A Comparative Study between Balloon Angioplasty and Stent in Hyperlipidemic Microswine


Abstract
:1. Introduction
2. Results
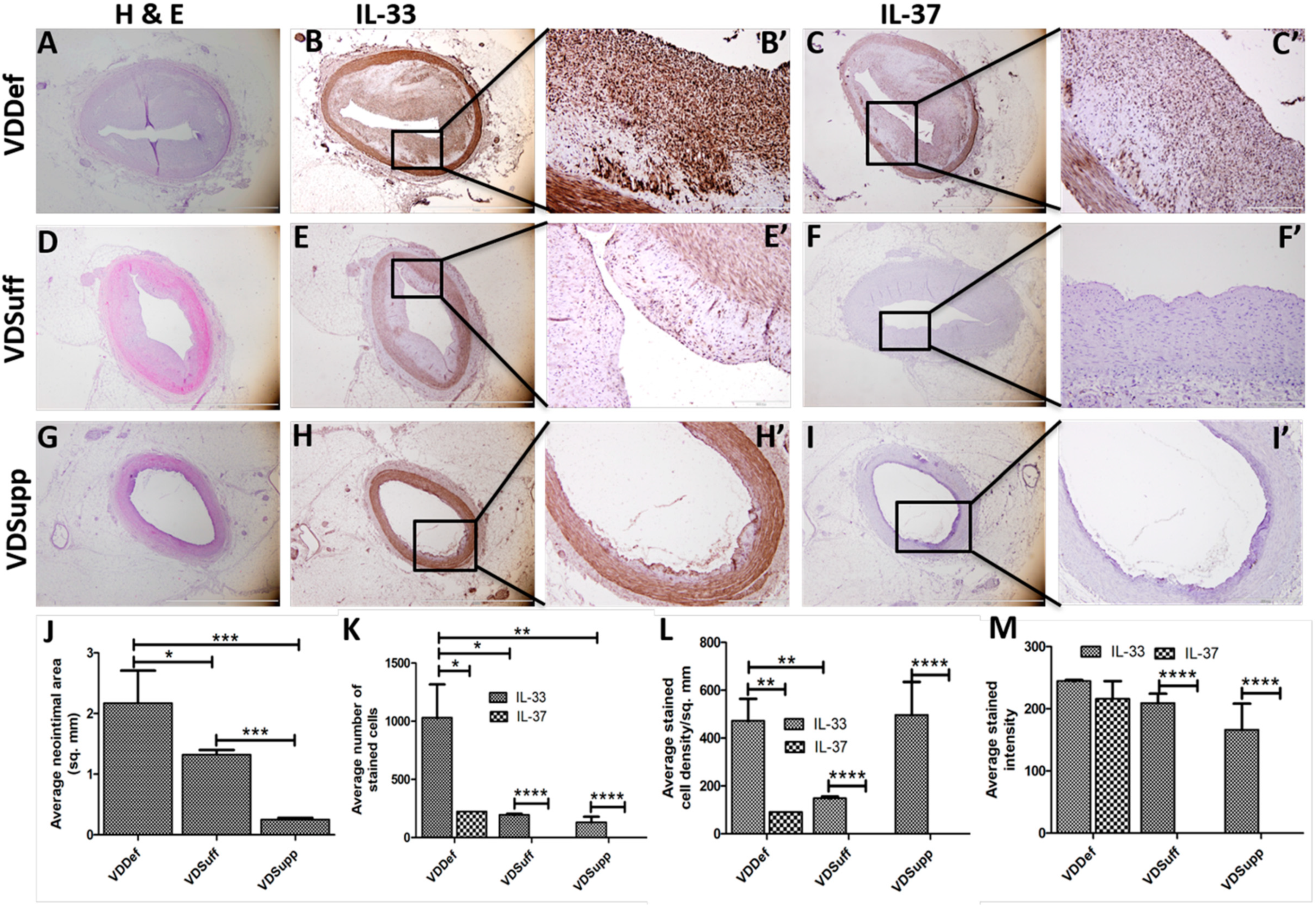
2.1. Vitamin D Supplementation Attenuates Neointima Formation in Coronary Arteries after Balloon Angioplasty and Stent Placement
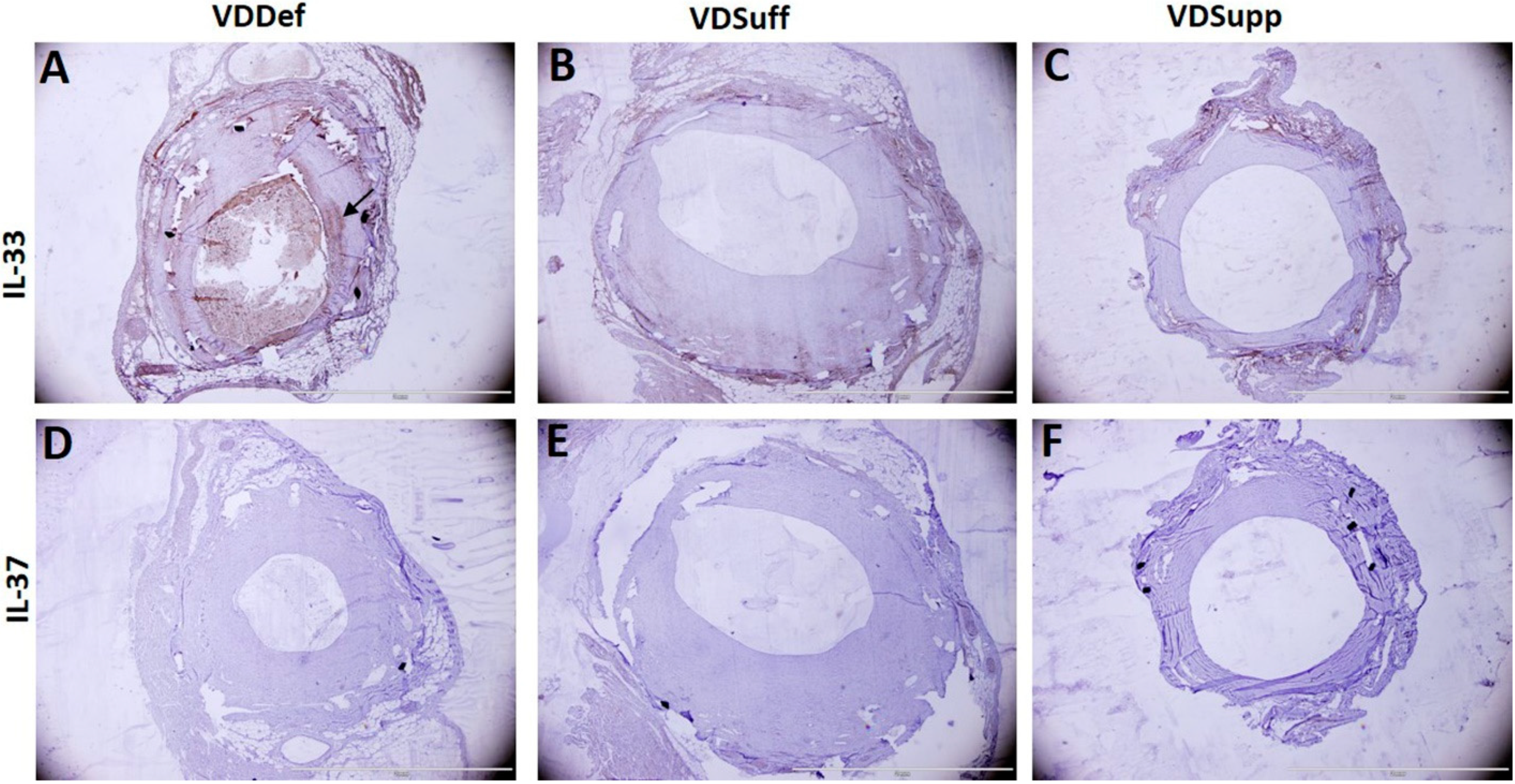
2.2. Vitamin D Supplementation Attenuates IL-33 and IL-37 Expression in Neointima
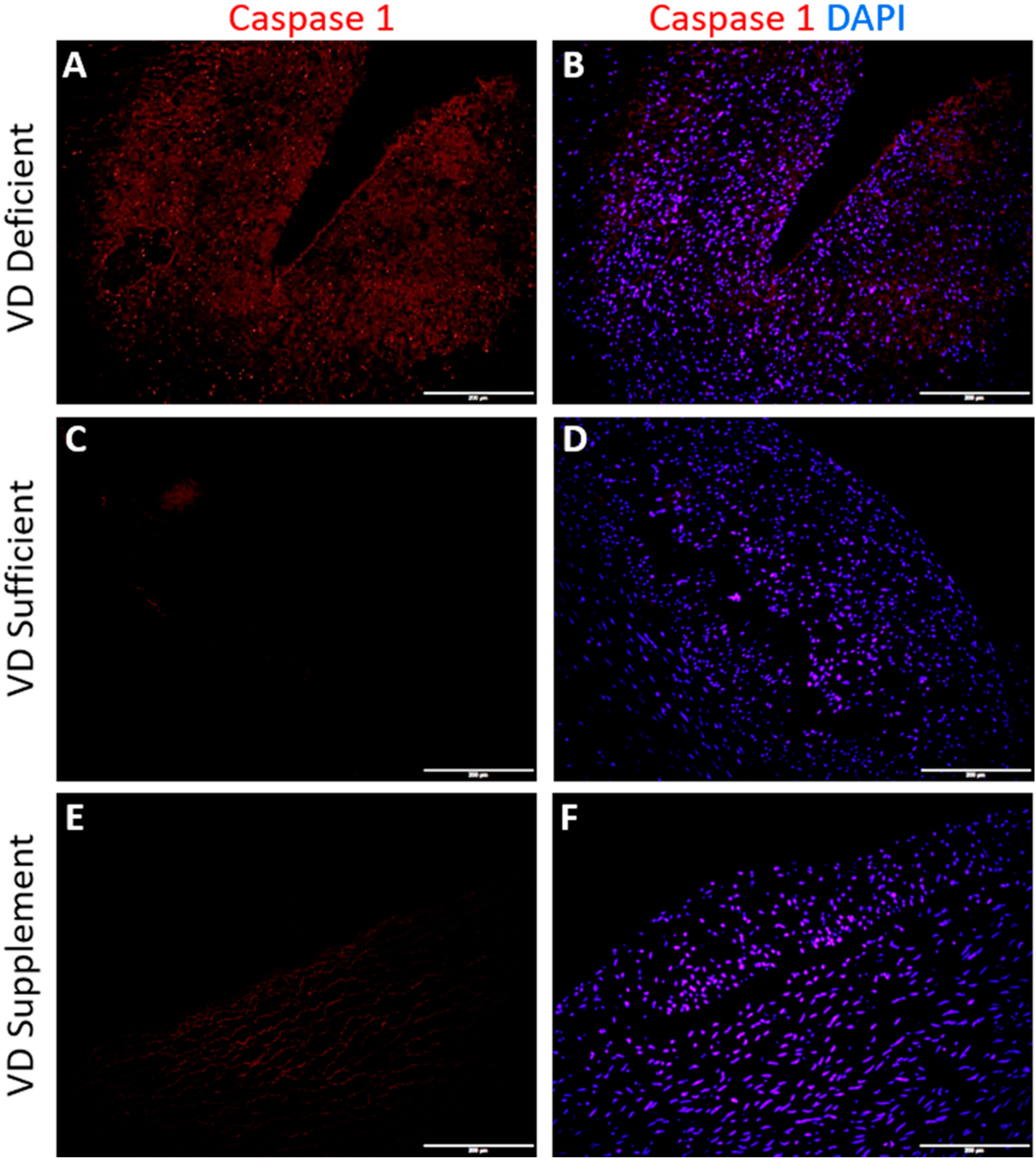
2.3. Vitamin D Deficiency Is Associated with Caspase-1 Expression
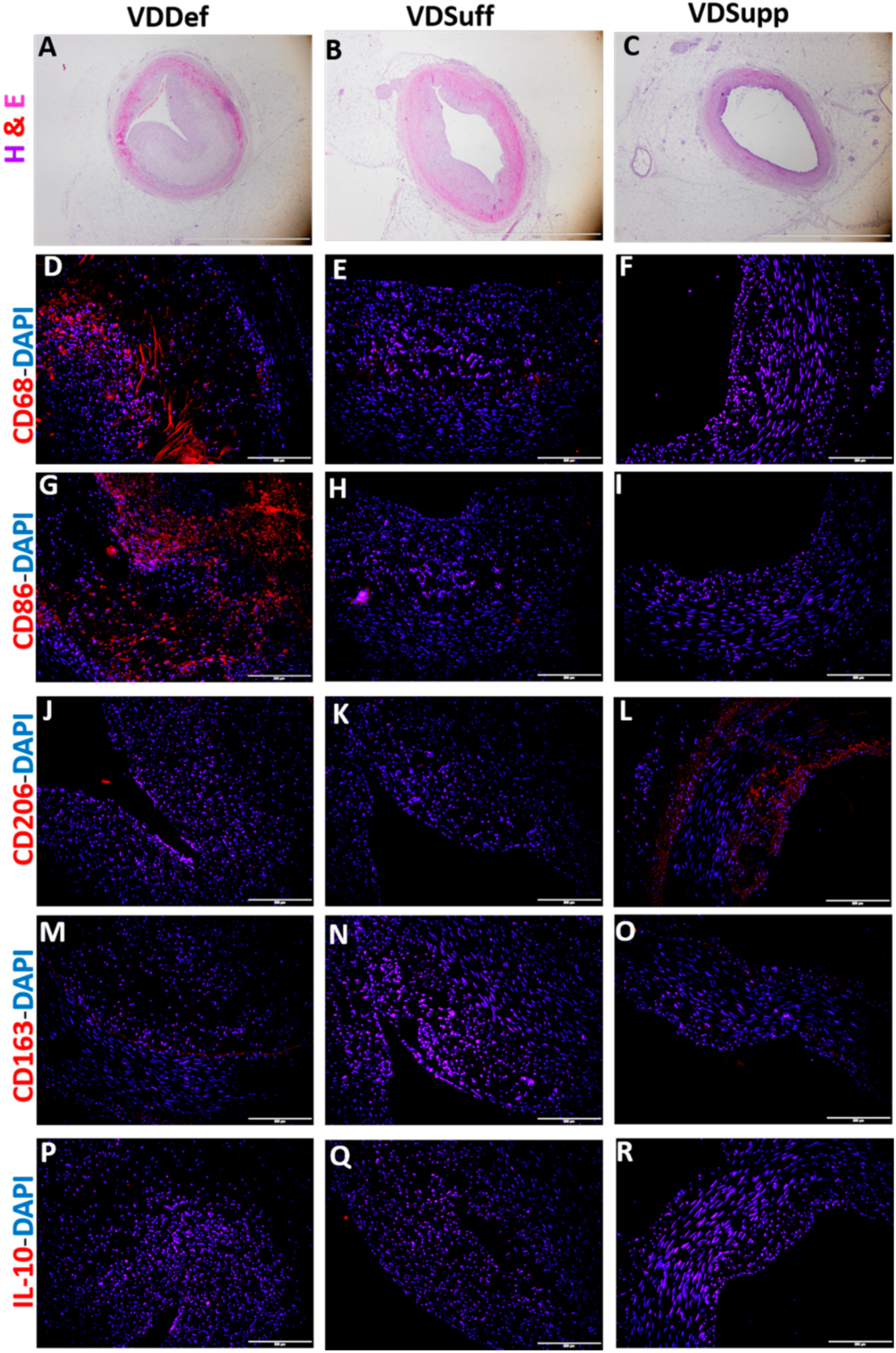
2.4. Vitamin D Supplementation Associates with Decreased Pro-Inflammatory Macrophages
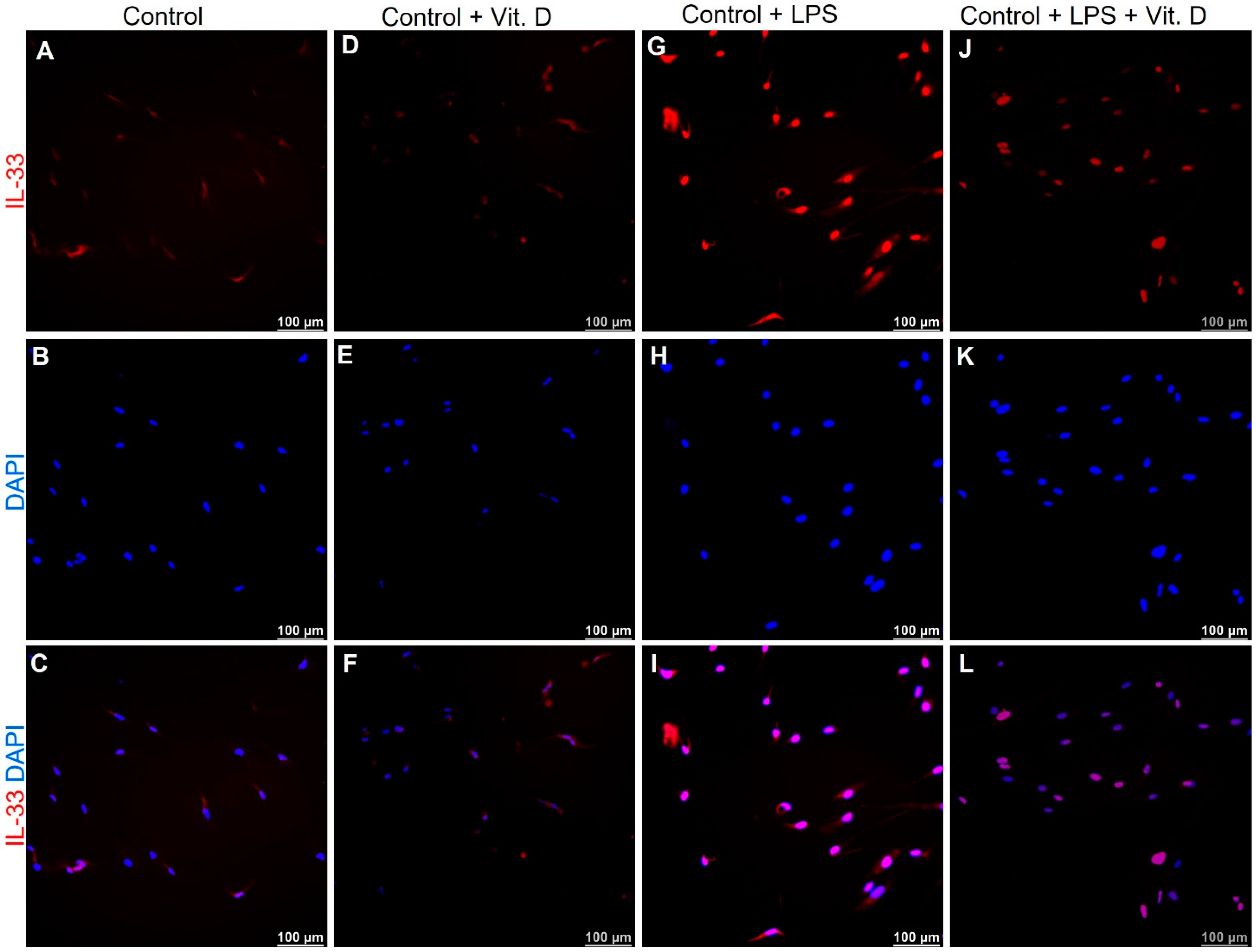
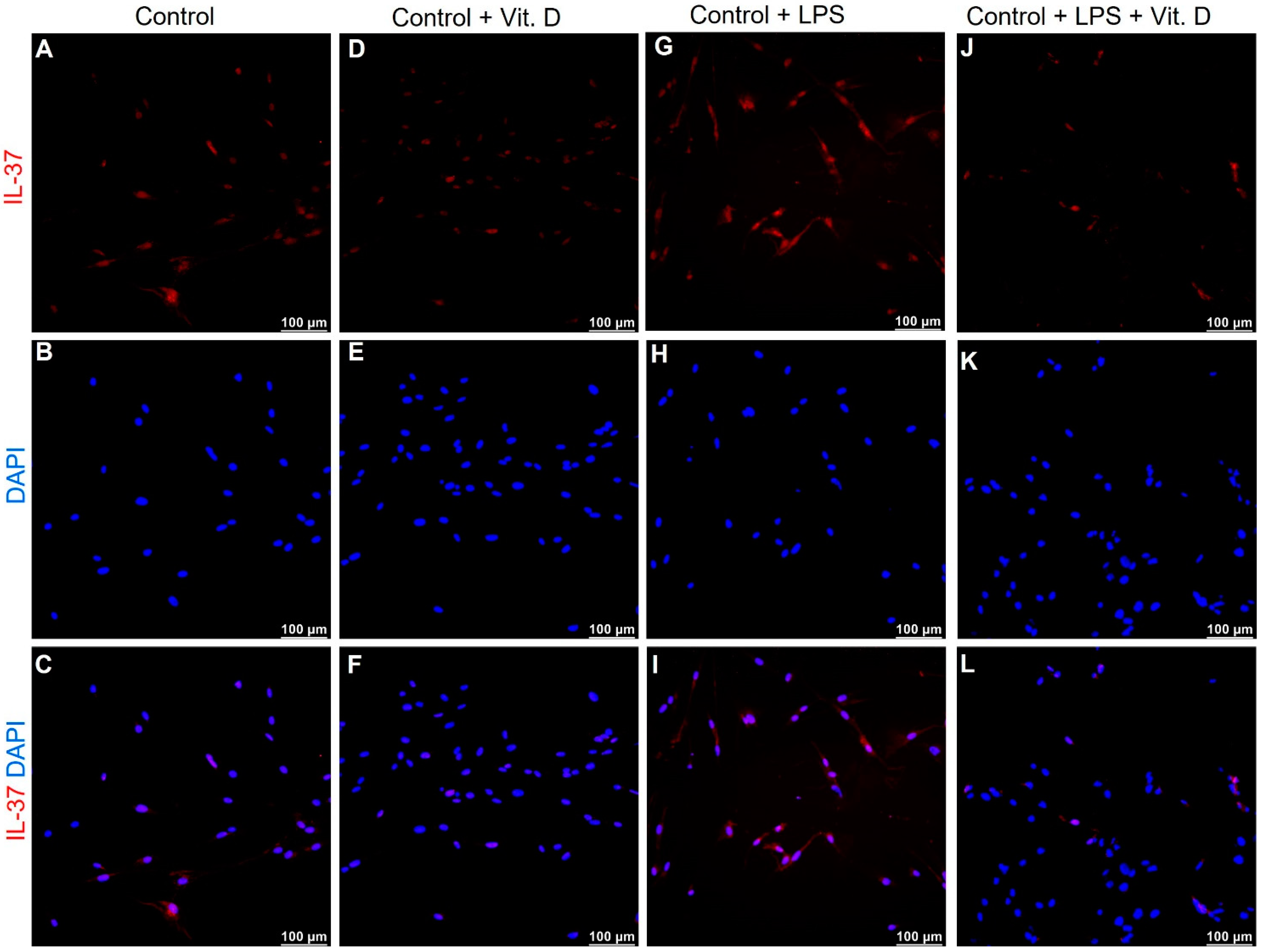
2.5. Vitamin D Attenuates the Expression of IL-33 and IL-37 in Lipopolysaccharides Treated Endothelial Cells In-Vitro
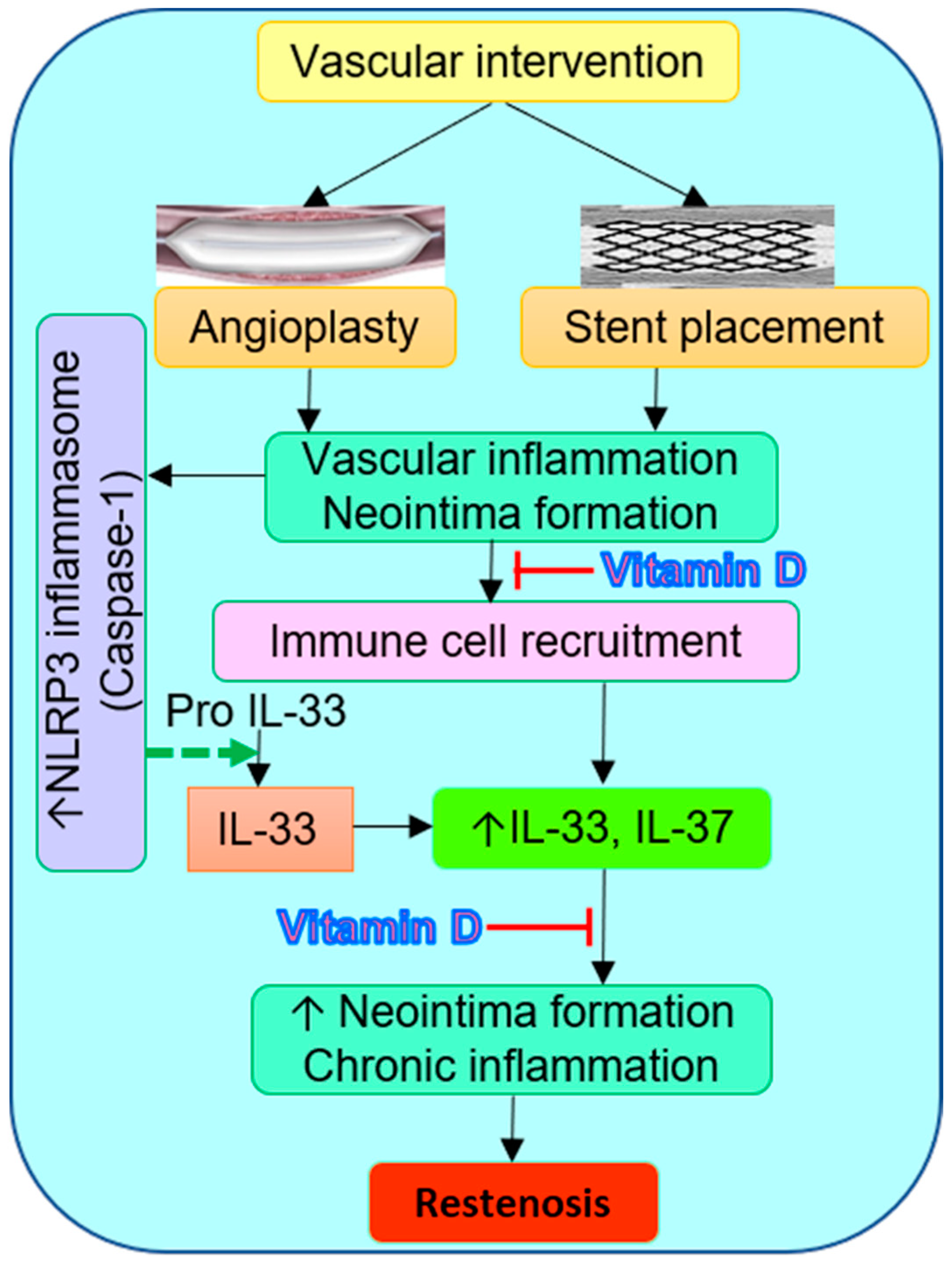
3. Discussion
4. Materials and Methods
4.1. Porcine Model of Neointimal Hyperplasia
4.2. Tissue Harvest and Processing
4.3. Immunohistochemical Studies
4.4. Immunofluorescence Study
4.5. Endothelial Cell Isolation and Primary Culture
4.6. Cell Culture, Stimulation, and Inhibition Studies
4.7. Statistical Analysis
5. Conclusions and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Limitations of the Study
References
- Rao, V.H.; Rai, V.; Stoupa, S.; Subramanian, S.; Agrawal, D.K. Tumor necrosis factor-α regulates triggering receptor expressed on myeloid cells-1-dependent matrix metalloproteinases in the carotid plaques of symptomatic patients with carotid stenosis. Atherosclerosis 2016, 248, 160–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rai, V.; Rao, V.H.; Shao, Z.; Agrawal, D.K. Dendritic cells expressing triggering receptor expressed on myeloid cells-1 correlate with plaque stability in symptomatic and asymptomatic patients with carotid stenosis. PLoS ONE 2016, 11, e0154802. [Google Scholar]
- Yin, K.; Agrawal, D.K. Vitamin D and inflammatory diseases. J. Inflamm. Res. 2014, 7, 69. [Google Scholar] [PubMed] [Green Version]
- Toutouzas, K.; Colombo, A.; Stefanadis, C. Inflammation and restenosis after percutaneous coronary interventions. Eur. Heart J. 2004, 25, 1679–1687. [Google Scholar] [CrossRef]
- Welt, F.G.; Rogers, C. Inflammation and restenosis in the stent era. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 1769–1776. [Google Scholar] [CrossRef] [Green Version]
- Kornowski, R.; Hong, M.K.; Tio, F.O.; Bramwell, O.; Wu, H.; Leon, M.B. In-stent restenosis: Contributions of inflammatory responses and arterial injury to neointimal hyperplasia. J. Am. Coll. Cardiol. 1998, 31, 224–230. [Google Scholar] [CrossRef] [Green Version]
- Nold, M.F.; Nold-Petry, C.A.; Zepp, J.A.; Palmer, B.E.; Bufler, P.; Dinarello, C.A. IL-37 is a fundamental inhibitor of innate immunity. Nat. Immunol. 2010, 11, 1014–1022. [Google Scholar] [CrossRef] [Green Version]
- Gupta, G.K.; Agrawal, T.; Rai, V.; Del Core, M.G.; Hunter, I.I.I.W.J.; Agrawal, D.K. Vitamin D supplementation reduces intimal hyperplasia and restenosis following coronary intervention in atherosclerotic swine. PLoS ONE 2016, 11, e0156857. [Google Scholar] [CrossRef]
- Jozanikohan, Z.; Kazemi, S.D. The effect of vitamin D deficiency treatment on post-PCI coronary restenosis and major adverse cardiac events. Int. Cardiovasc. Res. J. 2016, 10, 12–16. [Google Scholar] [CrossRef] [Green Version]
- Gupta, G.K.; Agrawal, T.; Del Core, M.G.; Hunter, I.I.I.W.J.; Agrawal, D.K. Decreased expression of vitamin D receptors in neointimal lesions following coronary artery angioplasty in atherosclerotic swine. PLoS ONE 2012, 7, e42789. [Google Scholar]
- Rai, V.; Agrawal, D.K. Role of vitamin D in cardiovascular diseases. Endocrinol. Metab. Clin. 2017, 46, 1039–1059. [Google Scholar] [CrossRef] [PubMed]
- Galkina, E.; Ley, K. Immune and inflammatory mechanisms of atherosclerosis. Annu. Rev. Immunol. 2009, 27, 165–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, D.; Ley, K. Immunity and Inflammation in Atherosclerosis. Circ. Res. 2019, 124, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.; Kang, M.; He, F.; Xiao, Z.; Liu, Z.; Yao, H.; Wu, J. Plasma interleukin-37 is increased and inhibits the production of inflammatory cytokines in peripheral blood mononuclear cells in systemic juvenile idiopathic arthritis patients. J. Transl. Med. 2018, 16, 277. [Google Scholar] [CrossRef]
- McCurdy, S.; Liu, C.A.; Yap, J.; Boisvert, W.A. Potential role of IL-37 in atherosclerosis. Cytokine 2019, 122, 154169. [Google Scholar] [CrossRef]
- Liu, J.; Lin, J.; He, S.; Wu, C.; Wang, B.; Liu, J.; Duan, Y.; Liu, T.; Shan, S.; Yang, K.; et al. Transgenic Overexpression of IL-37 Protects against Atherosclerosis and Strengthens Plaque Stability. Cell. Physiol. Biochem. 2018, 45, 1034–1050. [Google Scholar] [CrossRef] [Green Version]
- Schillinger, M.; Exner, M.; Mlekusch, W.; Haumer, M.; Ahmadi, R.; Rumpold, H.; Wagner, O.; Minar, E. Balloon angioplasty and stent implantation induce a vascular inflammatory reaction. J. Endovasc. Ther. 2002, 9, 59–66. [Google Scholar] [CrossRef]
- Shah, P.K. Inflammation, neointimal hyperplasia, and restenosis: As the leukocytes roll, the arteries thicken. Circulation 2003, 107, 2175–2177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, F.; Ahmed, A.S.; Kang, X.; Hu, G.; Liu, F.; Zhang, W.; Zhou, J. MicroRNA-15b/16 attenuates vascular neointima formation by promoting the contractile phenotype of vascular smooth muscle through targeting YAP. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 2145–2152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borovac, J.A.; D’Amario, D.; Vergallo, R.; Porto, I.; Bisignani, A.; Galli, M.; Annibali, G.; Montone, R.A.; Leone, A.M.; Niccoli, G.; et al. Neoatherosclerosis after drug-eluting stent implantation: A novel clinical and therapeutic challenge. Eur. Heart J. Cardiovasc. Pharmacother. 2019, 5, 105–116. [Google Scholar] [CrossRef]
- Niccoli, G.; D’Amario, D.; Borovac, J.A.; Santangelo, E.; Scalone, G.; Fracassi, F.; Vergallo, R.; Vetrugno, V.; Copponi, G.; Severino, A.; et al. Perilipin 2 levels are increased in patients with in-stent neoatherosclerosis: A clue to mechanisms of accelerated plaque formation after drug-eluting stent implantation. Int. J. Cardiol. 2018, 258, 55–58. [Google Scholar] [CrossRef]
- Ghali, R.; Altara, R.; Louch, W.E.; Cataliotti, A.; Mallat, Z.; Kaplan, A.; Zouein, F.A.; Booz, G.W. IL-33 (interleukin 33)/sST2 axis in hypertension and heart failure. Hypertension 2018, 72, 818–828. [Google Scholar] [CrossRef]
- Altara, R.; Ghali, R.; Mallat, Z.; Cataliotti, A.; Booz, G.W.; Zouein, F.A. Conflicting vascular and metabolic impact of the IL-33/sST2 axis. Cardiovasc. Res. 2018, 114, 1578–1594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kipshidze, N.; Dangas, G.; Tsapenko, M.; Moses, J.; Leon, M.B.; Kutryk, M.; Serruys, P. Role of the endothelium in modulating neointimal formation: Vasculoprotective approaches to attenuate restenosis after percutaneous coronary interventions. J. Am. Coll. Cardiol. 2004, 44, 733–739. [Google Scholar] [PubMed] [Green Version]
- Mohammadi-Kordkhayli, M.; Ahangar-Parvin, R.; Azizi, S.V.; Nemati, M.; Shamsizadeh, A.; Khaksari, M.; Moazzeni, S.M.; Jafarzadeh, A. Vitamin D Modulates the Expression of IL-27 and IL-33 in the Central Nervous System in Experimental Autoimmune Encephalomyelitis. Iran. J. Immunol. 2015, 12, 35–49. [Google Scholar] [PubMed]
- Sato, T.; Iwasaki, Y.; Kikkawa, Y.; Fukagawa, M. An efficacy of intensive vitamin D delivery to neointimal hyperplasia in recurrent vascular access stenosis. J. Vasc. Access 2016, 17, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Satish, M.; Gunasekar, P.; Asensio, J.A.; Agrawal, D.K. Vitamin D attenuates HMGB1-mediated neointimal hyperplasia after percutaneous coronary intervention in swine. Mol. Cell. Biochem. 2020, 474, 219–228. [Google Scholar] [CrossRef]
- Kim, D.H.; Sabour, S.; Sagar, U.N.; Adams, S.; Whellan, D.J. Prevalence of hypovitaminosis D in cardiovascular diseases (from the National Health and Nutrition Examination Survey 2001 to 2004). Am. J. Cardiol. 2008, 102, 1540–1544. [Google Scholar] [CrossRef]
- Tete, S.; Tripodi, D.; Rosati, M.; Conti, F.; Maccauro, G.; Saggini, A.; Cianchetti, E.; Caraffa, A.; Antinolfi, P.; Toniato, E. IL-37 (IL-1F7) the newest anti-inflammatory cytokine which suppresses immune responses and inflammation. Int. J. Immunopathol. Pharmacol. 2012, 25, 31–38. [Google Scholar] [CrossRef]
- Tunbridge, M.; França Gois, P.H. Vitamin D and the NLRP3 Inflammasome. Appl. Sci. 2020, 10, 8462. [Google Scholar] [CrossRef]
- Afonina, I.S.; Müller, C.; Martin, S.J.; Beyaert, R. Proteolytic processing of interleukin-1 family cytokines: Variations on a common theme. Immunity 2015, 42, 991–1004. [Google Scholar] [CrossRef] [Green Version]
- McNeill, E.; Channon, K.M.; Greaves, D.R. Inflammatory cell recruitment in cardiovascular disease: Murine models and potential clinical applications. Clin. Sci. 2010, 118, 641–655. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, S.; Moriyama, M.; Miyake, K.; Nakashima, H.; Tanaka, A.; Maehara, T.; Iizuka-Koga, M.; Tsuboi, H.; Hayashida, J.N.; Ishiguro, N.; et al. Interleukin-33 produced by M2 macrophages and other immune cells contributes to Th2 immune reaction of IgG4-related disease. Sci. Rep. 2017, 7, 42413. [Google Scholar]
- Tu, L.; Chen, J.; Xu, D.; Xie, Z.; Yu, B.; Tao, Y.; Shi, G.; Duan, L. IL-33-induced alternatively activated macrophage attenuates the development of TNBS-induced colitis. Oncotarget 2017, 8, 27704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, P.; Li, Q.; Su, S.; Dong, W.; Zong, S.; Ma, Q.; Yang, X.; Zuo, D.; Zheng, S.; Meng, X.; et al. Interleukin 37 suppresses M1 macrophage polarization through inhibition of the Notch1 and nuclear factor kappa B pathways. Front. Cell Dev. Biol. 2020, 8, 56. [Google Scholar] [CrossRef] [PubMed]
- Gunasekar, P.; Swier, V.J.; Fleegel, J.P.; Boosani, C.S.; Radwan, M.M.; Agrawal, D.K. Vitamin D and macrophage polarization in epicardial adipose tissue of atherosclerotic swine. PLoS ONE 2018, 13, e0199411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinberg, E.O.; Ferran, B.; Tsukahara, Y.; Hatch, M.M.; Han, J.; Murdoch, C.E.; Matsui, R. IL-33 induction and signaling are controlled by glutaredoxin-1 in mouse macrophages. PLoS ONE 2019, 14, e0210827. [Google Scholar] [CrossRef] [PubMed]
- Holick, M.F. Vitamin D deficiency. N. Engl. J. Med. 2007, 357, 266–281. [Google Scholar] [CrossRef]
- Hathcock, J.N.; Shao, A.; Vieth, R.; Heaney, R. Risk assessment for vitamin D. Am. J. Clin. Nutr. 2007, 85, 6–18. [Google Scholar] [CrossRef] [Green Version]
- Swier, V.J.; Tang, L.; Krueger, K.D.; Radwan, M.M.; Del Core, M.G.; Agrawal, D.K. Coronary injury score correlates with proliferating cells and alpha-smooth muscle actin expression in stented porcine coronary arteries. PLoS ONE 2015, 10, e0138539. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Swine → | VDDef | VDSuff | VDSupp | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Parameter ↓ | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | |
| Mean neointimal area (mm2) | 1.86 ± 0.08 | 1.43 ± 0.16 | 3.22 ± 0.04 | 1.45 ± 0.24 | 1.17 ± 0.04 | 1.34 ± 0.26 | 0.20 ± 0.02 | 0.28 ± 0.03 | 0.29 ± 0.05 | |
| Mean cell count | IL-33 | 1200 ± 81.30 | 472.67 ± 63.76 | 1419.33 ± 165.17 | 214.33 ± 81.88 | 189.33 ± 68.16 | 183 ± 139.37 | 77.33 ± 6.11 | 90.67 ± 15.53 | 226 ± 69.09 |
| Mean cell density/mm2 | 645.16 ± 27.15 | 330.54 ± 78.67 | 441.24 ± 45.92 | 147.822 ± 76.90 | 161.36 ± 58.55 | 136.23 ± 119.65 | 393.22 ± 63.53 | 327.71 ± 51.46 | 770.45 ± 309.62 | |
| Mean stained intensity | 246.86 ± 3.51 | 241.29 ± 5.92 | 245.72 ± 5.88 | 224.64 ± 12.55 | 224.31 ± 11.39 | 179.35 ± 55.65 | 128.22 ± 16.49 | 120.84 ± 15.25 | 250.23 ± 2.08 | |
| Mean cell count | IL-37 | 84 ± 35.55 | 121 ± 21.28 | 466.67 ± 13.32 | 0 | 0 | 0 | 0 | 0 | 0 |
| Mean cell density/mm2 | 45.16 ± 20.53 | 84.62 ± 5.68 | 145.08 ± 2.61 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Mean stained intensity | 159.88 ± 14.65 | 250.74 ± 2.03 | 237.97 ± 5.70 | 0 | 0 | 0 | 0 | 0 | 0 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rai, V.; Agrawal, D.K. Immunomodulation of IL-33 and IL-37 with Vitamin D in the Neointima of Coronary Artery: A Comparative Study between Balloon Angioplasty and Stent in Hyperlipidemic Microswine. Int. J. Mol. Sci. 2021, 22, 8824. https://doi.org/10.3390/ijms22168824
Rai V, Agrawal DK. Immunomodulation of IL-33 and IL-37 with Vitamin D in the Neointima of Coronary Artery: A Comparative Study between Balloon Angioplasty and Stent in Hyperlipidemic Microswine. International Journal of Molecular Sciences. 2021; 22(16):8824. https://doi.org/10.3390/ijms22168824
Chicago/Turabian StyleRai, Vikrant, and Devendra K. Agrawal. 2021. "Immunomodulation of IL-33 and IL-37 with Vitamin D in the Neointima of Coronary Artery: A Comparative Study between Balloon Angioplasty and Stent in Hyperlipidemic Microswine" International Journal of Molecular Sciences 22, no. 16: 8824. https://doi.org/10.3390/ijms22168824