
The Impact of Anthocyanins and Iridoids on Transcription Factors Crucial for Lipid and Cholesterol Homeostasis


Abstract
:1. Introduction
2. Major Transcription Factors Involved in Lipid and Cholesterol Metabolism
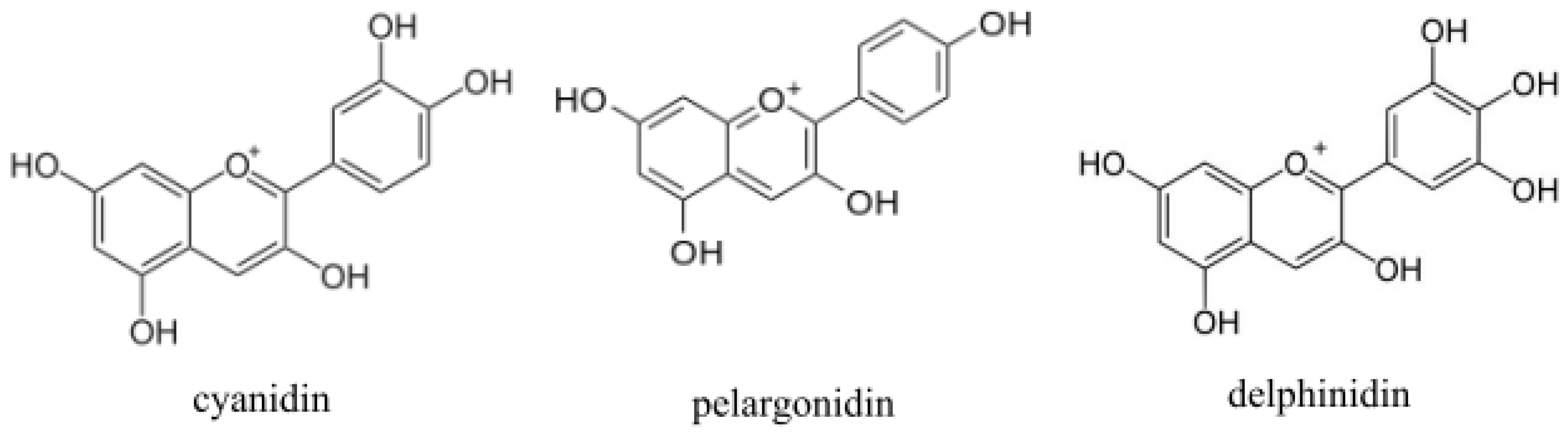
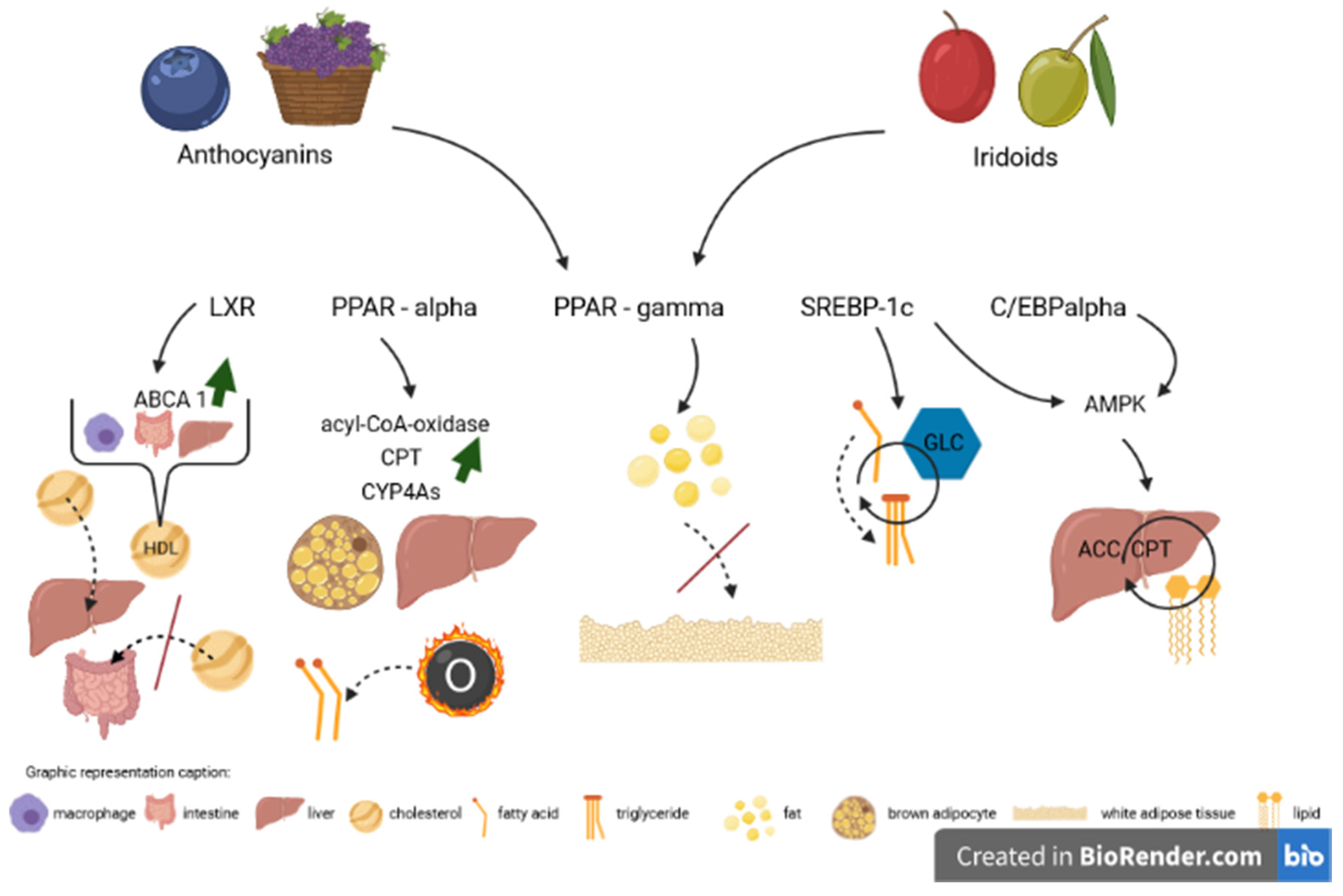
3. Impact of Anthocyanins
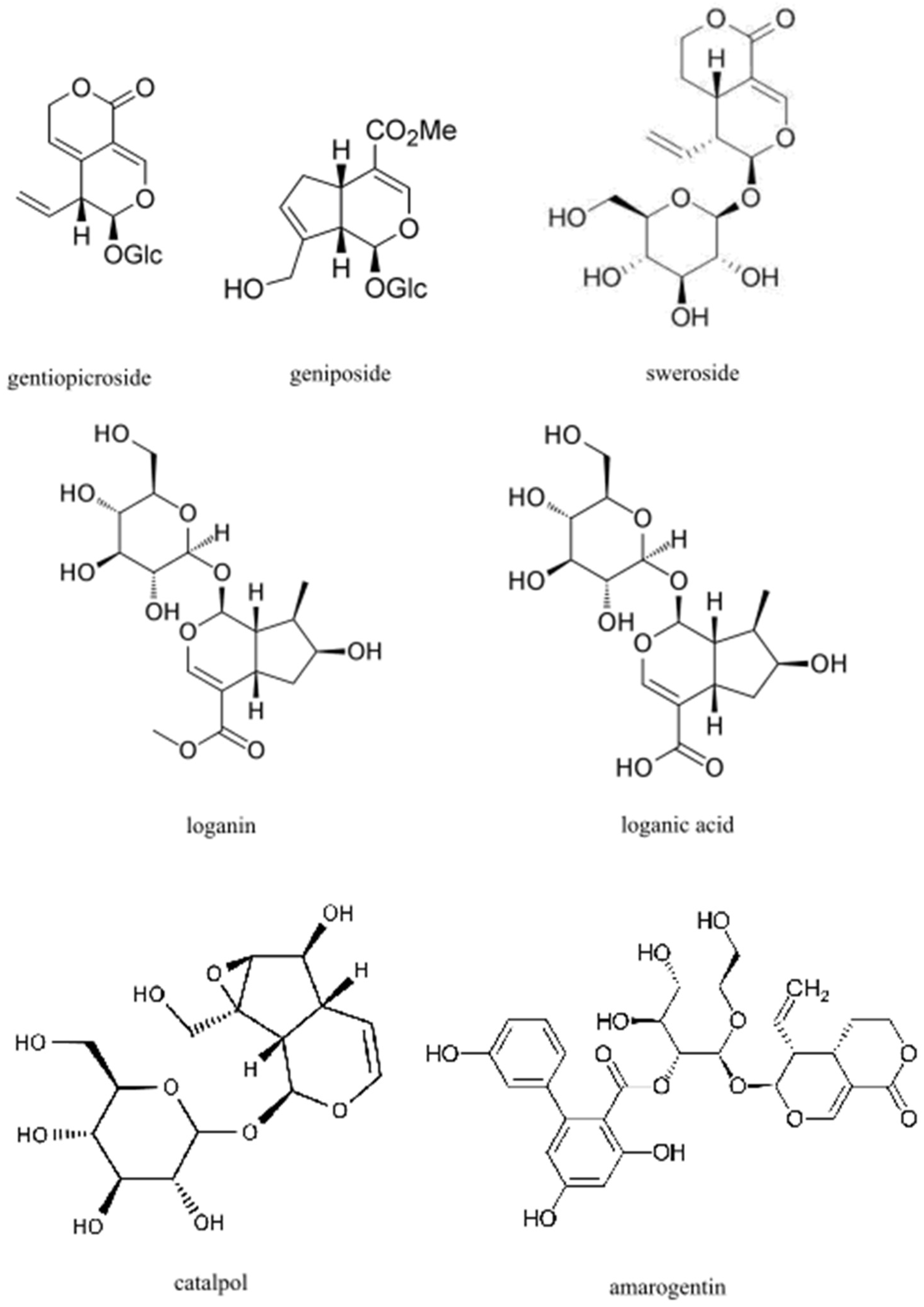
4. Impact of Iridoids
5. Future Perspectives
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ABCA1 | ATP-binding cassette transporter A1 |
| HDL | high-density lipoprotein |
| acyl-CoA-oxidase | acyl coenzyme A oxidase |
| CPT | carnitine palmitoyl transferase |
| CYP4As | cytochrome P450 4A subfamily enzymes |
| GLC | glucose |
| AMPK | AMP-activated protein kinase |
| ACC | acetyl coenzyme A carboxylase |
| O | β-oxidation |
References
- Papavassiliou, A.G. Molecular Medicine. Transcription Factors. N. Engl. J. Med. 1995, 332, 45–47. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.I.; Young, R.A. Transcriptional Regulation and its Misregulation in Disease. Cell 2013, 152, 1237–1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaquerizas, J.M.; Kummerfeld, S.K.; Teichmann, S.A.; Luscombe, N.M. A Census of Human Transcription Factors: Function, Expression and Evolution. Nat. Rev. Genet. 2009, 10, 252–263. [Google Scholar] [CrossRef]
- Papavassiliou, K.A.; Papavassiliou, A.G. Transcription Factor Drug Targets. J. Cell Biochem. 2016, 9999, 1–5. [Google Scholar] [CrossRef]
- Ortega, E.; Rengachari, S.; Ibrahim, Z.; Hoghoughi, N.; Gaucher, J.; Holehouse, A.S.; Khochbin, S.; Panne, D. Transcription Factor Dimerization Activates the p300 Acetyltransferase. Nature 2019, 562, 538–544. [Google Scholar] [CrossRef]
- Lee, T.I.; Young, R.A. Transcription of Eukaryotic Protein-Coding Genes. Annu. Rev. Genet. 2000, 34, 77–137. [Google Scholar] [CrossRef]
- Lemon, B.; Tjian, R. Orchestrated Response: A Symphony of Transcription Factors for Gene Control. Genes Dev. 2000, 14, 2551–2569. [Google Scholar] [CrossRef] [Green Version]
- Orphanides, G.; Lagrange, T.; Reinberg, D. The General Transcription Factors of RNA Polymerase II. Genes Dev. 1996, 10, 2657–2683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartnik, E. Struktura i działanie genów eukariotycznych. In Genetyka Molekularna; Węgleński, P., Ed.; Wydawnictwa Naukowe PWN: Warszawa, Poland, 2002; p. 497. [Google Scholar]
- Cao, G.; Liang, Y.; Broderick, C.L.; Oldham, B.A.; Beyer, T.P.; Schmidt, R.J.; Zhang, Y.; Stayrook, K.R.; Suen, C.; Otto, K.A.; et al. Antidiabetic Action of Liver X Receptor Agonist Mediated by Inhibition of Hepatic Gluconeogenesis. J. Biol. Chem. 2003, 278, 1131–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shuang, R.; Rui, X.; Wenfang, L. Phytosterols and Dementia. Plant Foods Hum. Nutr. 2016, 71, 347–354. [Google Scholar] [CrossRef]
- Sozański, T.; Kucharska, A.Z.; Szumny, A.; Magdalen, J.; Bielska, K.; Merwid-Ląd, A.; Woźniak, A.; Dzimira, S.; Piórecki, N.; Trocha, M. The Protective Effect of the Cornus Mas Fruits (Cornelian Cherry) on Hypertriglyceridemia and Atherosclerosis Through PPARα Activation in Hypercholesterolemic Rabbits. Phytomedicine 2014, 21, 1774–1784. [Google Scholar] [CrossRef]
- Jia, Y.; Hoang, M.H.; Jun, H.J.; Lee, J.H.; Lee, S.J. Cyanidin, a Natural Flavonoid, Is an Agonistic Ligand for Liver X Receptor Alpha and Beta and Reduces Cellular Lipid Accumulation in Macrophages and Hepatocytes. Bioorganic Med. Chem. Lett. 2013, 23, 4185–4190. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Li, L.; Yin, H. Cholesterol Homeostasis and Liver X Receptor (LXR) in Atherosclerosis. Cardiovasc. Hematol. Disord. Drug Targets 2018, 18, 27–33. [Google Scholar] [CrossRef]
- Chen, Z.; Liu, J.; Fu, Z.; Ye, C.; Zhang, R.; Song, Y.; Zhang, Y.; Li, H.; Ying, H.; Liu, H. 24(S)-Saringosterol from Edible Marine Seaweed Sargassum fusiforme is a Novel Selective LXRbeta Agonist. J. Agric. Food Chem. 2014, 62, 6130–6137. [Google Scholar] [CrossRef]
- Hoang, M.H.; Jia, Y.; Jun, H.J.; Lee, J.H.; Lee, B.Y.; Lee, S.J. Fucosterol is a Selective Liver X Receptor Modulator that Regulates the Expression of Key Genes in Cholesterol Homeostasis in Macrophages, Hepatocytes, and Intestinal Cells. J. Agric. Food Chem. 2012, 60, 11567–11575. [Google Scholar] [CrossRef] [PubMed]
- El Kharrassi, Y.; Samadi, M.; Lopez, T.; Nury, T.; El Kebbaj, R.; Andreoletti, P.; El Hajj, H.I.; Vamecq, J.; Moustaid, K.; Latruffe, N.; et al. Biological Activities of Schottenol and Spinasterol, Two Natural Phytosterols Present in Argan Oil and in Cactus Pear Seed Oil, on Murine Microglial BV2 cells. Biochem. Biophys. Res. Commun. 2014, 446, 798–804. [Google Scholar] [CrossRef]
- Bogie, J.; Hoeks, C.; Schepers, M.; Tiane, A.; Cuypers, A.; Leijten, F.; Chintapakorn, Y.; Suttiyut, T.; Pornpakakul, S.; Struik, D.; et al. Dietary Sargassum fusiforme Improves Memory and Reduces Amyloid Plaque Load in an Alzheimer’s Disease Mouse Model. Sci. Rep. 2019, 9, 4908. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Zhong, H.; Leng, L.; Jiang, Z. Effects of Soy Isoflavone on Hepatic Steatosis in High Fat-Induced Rats. J. Clin. Biochem. Nutr. 2017, 61, 85–90. [Google Scholar] [CrossRef]
- Ronis, M.J. Effects of Soy Containing Diet and Isoflavones on Cytochrome P450 Enzyme Expression and Activity. Drug Metab. Rev. 2016, 48, 331–341. [Google Scholar] [CrossRef]
- Illesca, P.; Valenzuela, R.; Espinosa, A.; Echeverría, F.; Soto-Alarcon, S.; Ortiz, M.; Videla, L.A. Hydroxytyrosol Supplementation Ameliorates the Metabolic Disturbances in White Adipose Tissue From Mice Fed a High-Fat Diet Through Recovery of Transcription Factors Nrf2, SREBP-1c, PPAR-γ and NF-κB. Biomed. Pharmacother. 2019, 109, 2472–2481. [Google Scholar] [CrossRef] [PubMed]
- Wakil, S.J.; Abu-Elheiga, L.A. Fatty Acid Metabolism: Target for Metabolic Syndrome. J. Lipid Res. 2009, 50, S138–S143. [Google Scholar] [CrossRef] [Green Version]
- Ramji, D.P.; Foka, P. CCAAT/enhancer-Binding Proteins: Structure, Function and Regulation. Biochem. J. 2002, 365, 561–575. [Google Scholar] [CrossRef] [Green Version]
- Clarke, S.L.; Robinson, C.E.; Gimble, J.M. CCAAT/enhancer Binding Proteins Directly Modulate Transcription From the Peroxisome Proliferator-Activated Receptor Gamma 2 Promoter. Biochem. Biophys. Res. Commun. 1997, 240, 99–103. [Google Scholar] [CrossRef]
- Chan, C.Y.; Wei, L.; Castro-Munozledo, F.; Koo, W.L. (−)-Epigallocatechin-3-gallate Blocks 3T3-L1 Adipose Conversion by Inhibition of Cell Proliferation and Suppression of Adipose Phenotype Expression. Life Sci. 2011, 89, 779–785. [Google Scholar] [CrossRef] [PubMed]
- Angin, Y.; Beauloye, C.; Horman, S.; Bertrand, L. Regulation of Carbohydrate Metabolism, Lipid Metabolism, and Protein Metabolism by AMPK. In AMP-Activated Protein Kinase; Cordero, M., Viollet, B., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 23–43. [Google Scholar]
- Guo, H.; Liu, G.; Zhong, R.; Wang, Y.; Wang, D.; Xia, M. Cyanidin-3-O-β-glucoside Regulates Fatty Acid Metabolism via an AMP-activated Protein Kinase-dependent Signaling Pathway in Human HepG2 cells. Lipids Health Dis. 2012, 11, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bengoechea-Alonso, M.T.; Ericsson, J. SREBP in Signal Transduction: Cholesterol Metabolism and Beyond. Curr. Opin. Cell Biol. 2007, 19, 215–222. [Google Scholar] [CrossRef]
- Chen, G.; Liang, G.; Ou, J.; Goldstein, J.L.; Brown, M.S. Central Role of Liver X Receptor in Insulin-mediated Activation of SREBP-1c Transcription and Stimulation of Fatty Acid Synthesis in Liver. Proc. Natl. Acad. Sci. USA 2004, 101, 11245–11250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferré, P.; Foufelle, F. Hepatic Steatosis: A Role for de novo Lipogenesis and the Transcription Factor SREBP-1c. Diabetes Obes. Metab. 2010, 12, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, T.; Li, D.; Ikaga, R. Effective Food Ingredients for Fatty Liver: Soy Protein β-Conglycinin and Fish Oil. Int. J. Mol. Sci. 2018, 19, 4107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terzo, S.; Caldara, G.F.; Ferrantelli, V.; Puleio, R.; Cassata, G.; Mulè, F.; Amato, A. Pistachio Consumption Prevents and Improves Lipid Dysmetabolism by Reducing the Lipid Metabolizing Gene Expression in Diet-Induced Obese Mice. Nutrients 2018, 10, 1857. [Google Scholar] [CrossRef] [Green Version]
- Danielewski, M.; Matuszewska, A.; Nowak, B.; Kucharska, A.Z.; Sozański, T. The Effects of Natural Iridoids and Anthocyanins on Selected Parameters of Liver and Cardiovascular System Functions. Oxidative Med. Cell Longev. 2020, 2020, 2735790. [Google Scholar] [CrossRef] [PubMed]
- Cassidy, A.; Bertoia, M.; Chiuve, S.; Flint, A.; Forman, J.; Rimm, E.B. Habitual Intake of Anthocyanins and Flavanones and Risk of Cardiovascular Disease in Men. Am. J. Clin. Nutr. 2016, 104, 587–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Beecher, G.R.; Holden, J.M.; Haytowitz, D.B.; Gebhardt, S.E.; Prior, R.L. Concentrations of Anthocyanins in Common Foods in the United States and Estimation of Normal Consumption. J. Agric. Food Chem. 2006, 54, 4069–4075. [Google Scholar] [CrossRef] [PubMed]
- Sozański, T.; Kucharska, A.Z.; Rapak, A.; Szumny, D.; Trocha, M.; Merwid-Ląd, A.; Dzimira, S.; Piasecki, T.; Piórecki, N.; Magdalan, J.; et al. Iridoid-loganic Acid Versus Anthocyanins From the Cornus Mas Fruits (Cornelian Cherry): Common and Different Effects on Diet-Induced Atherosclerosis, PPARs Expression and Inflammation. Atherosclerosis 2016, 254, 151–160. [Google Scholar] [CrossRef]
- Park, M.; Sharma, A.; Lee, H.J. Anti-Adipogenic Effects of Delphinidin-3- O- β-Glucoside in 3T3-L1 Preadipocytes and Primary White Adipocytes. Molecules 2019, 24, 1848. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.; Lee, M.; Lefevre, M.; Kim, H.R. Anthocyanins Inhibit Lipogenesis During Adipocyte Differentiation of 3T3-L1 Preadipocytes. Plant Foods Hum. Nutr. 2014, 69, 137–141. [Google Scholar] [CrossRef]
- Jia, Y.; Kim, J.Y.; Jun, H.J.; Kim, S.J.; Lee, J.H.; Hoang, M.H.; Kim, H.S.; Chang, H.I.; Hwang, K.Y.; Um, S.J.; et al. Cyanidin is an Agonistic Ligand for Peroxisome Proliferator-Activated Receptor-alpha Reducing Hepatic Lipid. Biochim. Biophys. Acta 2013, 1831, 698–708. [Google Scholar] [CrossRef]
- Fu, Y.; Wei, Z.; Zhou, E.; Zhang, N.; Yang, Z. Cyanidin-3-O-β-glucoside Inhibits Lipopolysaccharide-Induced Inflammatory Response in Mouse Mastitis Model. J. Lipid Res. 2014, 55, 1111–1119. [Google Scholar] [CrossRef] [Green Version]
- Du, C.; Shi, Y.; Ren, Y.; Wu, H.; Yao, F.; Wei, J.; Wu, M.; Hou, Y.; Duan, H. Anthocyanins Inhibit High-Glucose-Induced Cholesterol Accumulation and Inflammation by Activating LXRα Pathway in HK-2 Cells. Drug Des. Dev. Ther. 2015, 9, 5099–5113. [Google Scholar]
- Hwang, Y.P.; Choi, J.H.; Han, E.H.; Kim, H.G.; Wee, J.H.; Jung, K.O.; Jung, K.H.; Kwon, K.I.; Jeong, T.C.; Chung, Y.C.; et al. Purple Sweet Potato Anthocyanins Attenuate Hepatic Lipid Accumulation through Activating Adenosine Monophosphate-activated Protein Kinase in Human HepG2 Cells and Obese Mice. Nutr. Res. 2011, 31, 896–906. [Google Scholar] [CrossRef]
- Chang, J.J.; Hsu, M.J.; Huang, H.P.; Chung, D.J.; Chang, Y.C.; Wang, C.J. Mulberry Anthocyanins Inhibit Oleic Acid Induced Lipid Accumulation by Reduction of Lipogenesis and Promotion of Hepatic Lipid Clearance. J. Agric. Food Chem. 2013, 61, 6069–6076. [Google Scholar] [CrossRef]
- Park, M.; Yoo, J.H.; Lee, Y.S.; Lee, H.J. Lonicera caerulea Extract Attenuates Non-Alcoholic Fatty Liver Disease in Free Fatty Acid-Induced HepG2 Hepatocytes and in High Fat Diet-Fed Mice. Nutrients 2019, 11, 494. [Google Scholar] [CrossRef] [Green Version]
- de Sousa, A.R.; de Castro Moreira, M.E.; Lopes Toledo, R.C.; Anjos Benjamin, L.D.; Vieira Queiroz, V.A.; Paranho Veloso, M.; de Souza Reis, K.; Duarte Martino, H.S. Extruded Sorghum (Sorghum Bicolor L.) Reduces Metabolic Risk of Hepatic Steatosis in Obese Rats Consuming a High Fat Diet. Food Res. Int. 2018, 112, 48–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Kang, S.; Jeong, D.Y.; Jeong, S.Y.; Park, J.J.; Yun, H.S. Cyanidin and Malvidin in Aqueous Extracts of Black Carrots Fermented With Aspergillus Oryzae Prevent the Impairment of Energy, Lipid and Glucose Metabolism in Estrogen-Deficient Rats by AMPK Activation. Genes Nutr. 2015, 10, 455. [Google Scholar] [CrossRef]
- Khan, M.I.; Shin, J.H.; Shin, T.S.; Kim, M.Y.; Cho, N.J.; Kim, J.D. Anthocyanins From Cornus Kousa Ethanolic Extract Attenuate Obesity in Association With Anti-Angiogenic Activities in 3T3-L1 Cells by Down-Regulating Adipogeneses and Lipogenesis. PLoS ONE 2018, 13, e0208556. [Google Scholar] [CrossRef] [PubMed]
- Koh, E.S.; Lim, J.H.; Kim, M.Y.; Chung, S.; Shin, S.J.; Choi, B.S.; Kim, H.W.; Hwang, S.Y.; Kim, S.W.; Park, C.W.; et al. Anthocyanin-rich Seoritae Extract Ameliorates Renal Lipotoxicity via Activation of AMP-activated Protein Kinase in Diabetic Mice. J. Transl. Med. 2015, 13, 203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rimando, A.M.; Khan, S.I.; Mizuno, C.S.; Ren, G.; Mathews, S.T.; Kim, H.; Yokoyama, W. Evaluation of PPARα Activation by Known Blueberry Constituents. J. Sci. Food Agric. 2016, 96, 1666–1671. [Google Scholar] [CrossRef]
- Luna-Vital, D.; Weiss, M.; de Mejia, E.G. Anthocyanins From Purple Corn Ameliorated Tumor Necrosis Factor-α-Induced Inflammation and Insulin Resistance in 3T3-L1 Adipocytes via Activation of Insulin Signaling and Enhanced GLUT4 Translocation. Mol. Nutr. Food Res. 2017, 61, 1700362. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Wu, T.; Xu, D.; Chu, Q.; Lin, D.; Zheng, X. Dietary Sweet Cherry Anthocyanins Attenuates Diet-Induced Hepatic Steatosis by Improving Hepatic Lipid Metabolism in Mice. Nutrition 2016, 32, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Kao, E.S.; Tseng, T.H.; Lee, H.J.; Chan, K.C.; Wang, C.J. Anthocyanin Extracted From Hibiscus Attenuate Oxidized LDL-mediated Foam Cell Formation Involving Regulation of CD36 Gene. Chem. Biol. Interact. 2009, 179, 212–218. [Google Scholar] [CrossRef]
- Kim, H.K.; Kim, J.N.; Han, S.N.; Nam, J.H.; Na, H.N.; Ha, T.J. Black Soybean Anthocyanins Inhibit Adipocyte Differentiation in 3T3-L1 Cells. Nutr. Res. 2012, 32, 770–777. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Kim, J.H.; Lee, E.B.; Hur, W.; Kwon, O.J.; Park, H.J.; Yoon, S.K. Aronia melanocarpa Extract Ameliorates Hepatic Lipid Metabolism through PPARγ2 Downregulation. PLoS ONE 2017, 12, e0169685. [Google Scholar] [CrossRef] [Green Version]
- Aboonabi, A.; Aboonabi, A. Anthocyanins Reduce Inflammation and Improve Glucose and Lipid Metabolism Associated With Inhibiting Nuclear factor-kappaB Activation and Increasing PPAR-γ Gene Expression in Metabolic Syndrome Subjects. Free Radic. Biol. Med. 2020, 150, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Rahman, N.; Jeon, M.; Kim, Y.S. Delphinidin, a Major Anthocyanin, Inhibits 3T3-L1 Pre-Adipocyte Differentiation Through Activation of Wnt/β-catenin Signaling. Biofactors 2016, 42, 49–59. [Google Scholar]
- Turner, A.; Chen, S.N.; Nikolic, D.; van Breemen, R.; Farnsworth, N.R.; Pauli, G.F. Coumaroyl Iridoids and a Depside from Cranberry (Vaccinium macrocarpon). J. Nat. Prod. 2007, 70, 253–258. [Google Scholar] [CrossRef] [Green Version]
- Xie, P.J.; Huang, L.X.; Zhang, C.H.; Zhang, Y.L. Phenolic Compositions, and Antioxidant Performance of Olive Leaf and Fruit (Olea europaea L.) Extracts and their Structure–Activity Relationships. J. Funct. Foods 2015, 16, 460–471. [Google Scholar] [CrossRef]
- Kucharska, A.Z.; Szumny, A.; Sokół-Łętowska, A.; Piórecki, N.; Klymenko, S.V. Iridoids and Anthocyanins in Cornelian Cherry (Cornus mas L.) Cultivars. J. Food Compos. Anal. 2015, 40, 95–102. [Google Scholar] [CrossRef]
- Kucharska, A.Z.; Fecka, I. Identification of Iridoids in Edible Honeysuckle Berries (Lonicera caerulea L. var. kamtschatica Sevast.) by UPLC-ESI-qTOF-MS/MS. Molecules 2016, 21, 1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghisalberti, E.L. Biological and Pharmacological Activity of Naturally Occurring Iridoids and Secoiridoids. Phytomedicine 1998, 5, 147–163. [Google Scholar] [CrossRef]
- Desgagne, V.; Bouchard, L.; Guerin, R. MicroRNAs in Lipoprotein and Lipid Metabolism: From Biological Function to Clinical Application. Clin. Chem. Lab. Med. 2017, 55, 667–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, H.; Chen, K.; Feng, M.; Shao, W.; Wu, J.; Chen, K.; Liang, T.; Liu, C. Genipin Alleviates High-Fat Diet-Induced Hyperlipidemia and Hepatic Lipid Accumulation in Mice via miR-142a-5p/SREBP-1c Axis. FEBS J. 2018, 285, 501–517. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Xu, K.; Zhang, X.; Cao, J.; Jia, Z.; Yang, R.; Ma, C.; Chen, C.; Zhang, T.; Yan, Z. Studies on the Regulation of Lipid Metabolism and its Mechanism of the Iridoids Rich Fraction in Valeriana jatamansi Jones. Biomed. Pharmacother. 2016, 84, 1891–1898. [Google Scholar] [CrossRef] [PubMed]
- Malliou, F.; Andreadou, I.; Gonzalez, F.J.; Lazou, A.; Xepapadaki, E.; Vallianou, I.; Lambrinidis, G.; Mikros, E.; Marselos, M.; Skaltsounis, A.L.; et al. The Olive Constituent Oleuropein, as a PPARα Agonist, Markedly Reduces Serum Triglycerides. J. Nutr. Biochem. 2018, 59, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Huang, C.; Zong, G.; Zha, D.; Meng, X.; Li, J.; Tang, W. Hepatoprotective Effects of Geniposide in a Rat Model of Nonalcoholic Steatohepatitis. J. Pharm. Pharmacol. 2011, 63, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Patel, T.P.; Rawal, K.; Soni, S.; Gupta, S. Swertiamarin Ameliorates Oleic Acid Induced Lipid Accumulation and Oxidative Stress by Attenuating Gluconeogenesis and Lipogenesis in Hepatic Steatosis. Biomed. Pharmacother. 2016, 83, 785–791. [Google Scholar] [CrossRef]
- Yang, Q.; Shu, F.; Gong, J.; Ding, P.; Cheng, R.; Li, J.; Tong, R.; Ding, L.; Sun, H.; Huang, W.; et al. Sweroside Ameliorates NAFLD in High-Fat Diet Induced Obese Mice Through the Regulation of Lipid Metabolism and Inflammatory Response. J. Ethnopharmacol. 2020, 255, 112556. [Google Scholar] [CrossRef]
- He, Y.Q.; Ma, G.Y.; Peng, J.N.; Ma, Z.Y.; Hamann, M.T. Liver X Receptor and Peroxisome Proliferator-Activated Receptor Agonist from Cornus alternifolia. Biochim. Biophys. Acta 2012, 1820, 1021–1026. [Google Scholar] [CrossRef] [Green Version]
- Choi, K.M.; Shin, E.; Liu, Q.; Yoo, H.S.; Kim, Y.C.; Sung, S.H.; Hwang, B.Y.; Lee, M.K. Hydroxyframoside B, a Secoiridoid of Fraxinus Rhynchophylla, Inhibits Adipocyte Differentiation in 3T3-L1 Cells. Planta Med. 2011, 77, 1020–1023. [Google Scholar] [CrossRef] [PubMed]
- Bai, N.; He, K.; Ibarra, A.; Bily, A.; Roller, M.; Chen, X.; Rühl, R. Iridoids From Fraxinus Excelsior With Adipocyte Differentiation-Inhibitory and PPARalpha Activation Activity. J. Nat. Prod. 2010, 73, 2–6. [Google Scholar] [CrossRef]
- Drira, R.; Chen, S.; Sakamoto, K. Oleuropein and Hydroxytyrosol Inhibit Adipocyte Differentiation in 3 T3-L1 Cells. Life Sci. 2011, 89, 708–716. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.; Zhu, R.; Tian, Y.; Li, R.; Chen, B.; Zhang, H.; Xia, B.; Zhao, D.; Mo, F.; Zhang, D.; et al. Catalpol in Diabetes and Its Complications: A Review of Pharmacology, Pharmacokinetics, and Safety. Molecules 2019, 24, 3302. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Yao, J.; Gong, C.; Wang, B.; Zhou, P.; Zhou, S.; Yao, X. Gentiopicroside Ameliorates Diabetic Peripheral Neuropathy by Modulating PPAR- Γ/AMPK/ACC Signaling Pathway. Cell Physiol. Biochem. 2018, 50, 585–596. [Google Scholar] [CrossRef]
- Li, X.; Zhang, Y.; Jin, Q.; Xia, K.L.; Jiang, M.; Cui, B.W.; Wu, Y.L.; Song, S.Z.; Lian, L.H.; Nan, J.X. Liver Kinase B1/AMP-activated Protein Kinase-Mediated Regulation by Gentiopicroside Ameliorates P2X7 Receptor-Dependent Alcoholic Hepatosteatosis. Br. J. Pharmacol. 2018, 175, 1451–1470. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Li, J.; Wei, C.; He, Y.; Cao, Y.; Zhang, Y.; Sun, W.; Qiao, B.; He, J. Amelioration of Nonalcoholic Fatty Liver Disease by Swertiamarin in Fructose-Fed Mice. Phytomedicine 2019, 59, 152782. [Google Scholar] [CrossRef]
- Dinda, B.; Dinda, M.; Kulsi, G.; Chakraborty, A.; Dinda, S. Therapeutic Potentials of Plant Iridoids in Alzheimer’s and Parkinson’s Diseases: A Review. Eur. J. Med. Chem. 2019, 169, 185–199. [Google Scholar] [CrossRef]
- Park, E.; Kim, J.; Yeo, S.; Kim, G.; Ko, E.H.; Lee, S.W.; Li, W.Y.; Choi, C.W.; Jeong, S.Y. Antiadipogenic Effects of Loganic Acid in 3T3-L1 Preadipocytes and Ovariectomized Mice. Molecules 2018, 23, 1663. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Authors and Date of Publication | Research Model | Compounds Used in Study | Observed Changes |
|---|---|---|---|
| Aboonabi et al., 2020 [55] | human | berry anthocyanin supplements | PPAR-γ  |
| Chang et al., 2013 [43] | HepG2 cells | mulberry anthocyanin extract | SREBP-1c PPAR-α  |
| de Sousa et al., 2018 [45] | rats | extruded sorghum flour | SREBP-1c PPAR-α |
| Du et al., 2015 [41] | HK-2 cells | cyanidin-3-O-β-glucoside, cyanidin | LXRα PPAR-α |
| Fu et al., 2014 [40] | mice, mice mammary epithelial cells | cyanidin-3-O-β-glucoside | LXRα |
| Hwang et al., 2011 [42] | mice | purple sweet potato anthocyanin fraction | SREBP-1c |
| Jia et al., 2013 [39] | HepG2 cells, CHO-K1 cells | cyanidin | PPAR-α PPAR-δ PPAR-γ |
| Jia et al., 2013 [13] | macrophages, hepatocytes | cyanidin | LXRα, LXRβ SREBP-1c |
| Kao et al., 2009 [52] | mouse macrophage J774A.1 cells | hibiscus anthocyanin extract | PPAR-γ |
| Khan et al., 2018 [47] | 3T3-L1 cells | C. kousa anthocyanin ethanolic leaf extract | PPAR-γ C/EBPα |
| Kim et al., 2012 [53] | 3T3-L1 cells | black soybean anthocyanin extract | PPAR-γ |
| Koh et al., 2015 [48] | mice | Seoritae anthocyanin extract | PPAR-α PPAR-γ SREBP-1c |
| Lee et al., 2014 [38] | 3T3-L1 cells | grape anthocyanin isolate | LXRα PPAR-γ C/EBPα SREBP-1c |
| Luna-Vital et al., 2017 [50] | 3T3-L1 cells | purple corn pericarp anthocyanin extract, pure anthocyanins | PPAR-γ |
| Park et al., 2015 [46] | rats | unfermented and fermented black carrot extract | SREBP-1c PPAR-α |
| Park et al., 2017 [54] | mice, FL83B cells | A. melanocarpa spray-dried ethanol extract | PPAR-γ |
| Park et al., 2019 [44] | HepG2 cells | honeyberry extract | SREBP-1c PPAR-γ C/EBPα PPAR-α |
| Park et al., 2019 [37] | 3T3-L1 cells, primary white adipocytes | delphinidin-3-O-β-glucoside | PPAR-γ C/EBPα SREBP-1c |
| Rahman et al., 2016 [56] | 3T3-L1 cells | delphinidin | PPAR-γ C/EBP |
| Rimando et al., 2016 [49] | hamsters | blueberry peel extract |  PPAR-α PPAR-α |
| Song et al., 2016 [51] | mice | sweet cherry anthocyanins | PPAR-γ |
| Sozański et al., 2014 [12] | rabbits | cornelian cherry fruits lyophilisate | PPAR-α |
| Sozański et al., 2016 [36] | rabbits | mixture of anthocyanins | PPAR-α PPAR-γ |
— up regulation, —down regulation, —unchanged.| Authors and Date of Publication | Research Model | Compounds Used in Study | Observed Changes |
|---|---|---|---|
| Bai et al., 2010 [71] | 3T3-L1 cells | aqueous extract and compounds isolated from the seeds of F. excelsior | PPAR-α |
| Choi et al., 2011 [70] | 3T3-L1 cells | hydroxyframoside B | C/EBPα C/EBPβ PPAR-γ |
| Drira et al., 2011 [72] | 3T3-L1 cells | oleuropein | PPAR-γ C/EBPα SREBP-1c |
| He et al., 2012 [69] | HepG2 cells, CHO cells | leaf extract of C. alternifolia, incl. Kaempferol-3-O-β-glucopyranoside | PPAR-α PPAR-γ LXRα |
| Li et al., 2018 [75] | mice, HepG2 cells, macrophages | gentiopicroside | SREBP-1c PPAR-α |
| Lu et al., 2018 [74] | rats | gentiopicroside | PPAR-γ |
| Ma et al., 2011 [66] | rats | geniposide | PPAR-α |
| Malliou et al., 2018 [65] | mice | oleuropein | PPAR-α |
| Park et al., 2018 [78] | 3T3-L1 cells, mice | loganic acid | PPAR-γ C/EBPα |
| Patel et al., 2016 [67] | HepG2 cells | swertiamarin | SREBP-1c PPAR-α |
| Sozański et al., 2014 [12] | rabbits | cornelian cherry fruits lyophilisate | PPAR-α |
| Sozański et al., 2016 [36] | rabbits | loganic acid | PPAR-α PPAR-γ |
| Yang et al., 2019 [76] | mice | swertiamarin | SREBP-1c |
| Yang et al., 2020 [68] | mice | sweroside | PPAR-α |
| Zhong et al., 2018 [63] | mice, primary hepatocytes | genipin | SREBP-1c |
| Zhu et al., 2016 [64] | rats | iridoids rich fraction in V. jatamansi | LXRα SREBP-1c PPAR-α |
—up regulation, —down regulation.Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Danielewski, M.; Matuszewska, A.; Szeląg, A.; Sozański, T. The Impact of Anthocyanins and Iridoids on Transcription Factors Crucial for Lipid and Cholesterol Homeostasis. Int. J. Mol. Sci. 2021, 22, 6074. https://doi.org/10.3390/ijms22116074
Danielewski M, Matuszewska A, Szeląg A, Sozański T. The Impact of Anthocyanins and Iridoids on Transcription Factors Crucial for Lipid and Cholesterol Homeostasis. International Journal of Molecular Sciences. 2021; 22(11):6074. https://doi.org/10.3390/ijms22116074
Chicago/Turabian StyleDanielewski, Maciej, Agnieszka Matuszewska, Adam Szeląg, and Tomasz Sozański. 2021. "The Impact of Anthocyanins and Iridoids on Transcription Factors Crucial for Lipid and Cholesterol Homeostasis" International Journal of Molecular Sciences 22, no. 11: 6074. https://doi.org/10.3390/ijms22116074