
An Update on the Role of Leptin in the Immuno-Metabolism of Cartilage

 , ,
, ,  , , , , ,
, , , , ,  ,
,
Abstract
:1. Introduction
2. Physiology and Pathophysiology of Leptin
2.1. Leptin in Pathologies of Articular Cartilage
2.2. Leptin in Chondrocyte Proliferation and Metabolism
2.3. Leptin in Cartilage Inflammation
3. The Role of Leptin in Pathological Joint Diseases
3.1. Leptin in Osteoarthritis
3.2. Leptin in Rheumatoid Arthritis
3.3. Leptin in Systemic Lupus Erythematosus (SLE)
3.4. Leptin in Psoriatic Arthritis
4. Leptin as a Biological Therapy
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| ECM | extracellular matrix |
| WAT | white adipose tissue |
| OA | osteoarthritis |
| RA | rheumatoid arthritis |
| DC | dendritic cells |
| SF | synovial fluid |
| SLE | systemic lupus erythematosus |
References
- Archer, C.W.; Francis-West, P. The chondrocyte. Int. J. Biochem. Cell Biol. 2003, 35, 401–404. [Google Scholar] [CrossRef]
- Sophia Fox, A.J.; Bedi, A.; Rodeo, S.A. The Basic Science of Articular Cartilage: Structure, Composition, and Function. Sports Health 2009, 1, 461–468. [Google Scholar] [CrossRef]
- Bhosale, A.M.; Richardson, J.B. Articular cartilage: Structure, injuries and review of management. Br. Med. Bull. 2008, 87, 77–95. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Liu, W.; Bemis, A.; Wang, E.; Qiu, Y.; Morris, E.A.; Flannery, C.R.; Yang, Z. Comparative proteomic characterization of articular cartilage tissue from normal donors and patients with osteoarthritis. Arthritis Rheum. 2007, 56, 3675–3684. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Sinkeviciute, D.; He, Y.; Karsdal, M.; Henrotin, Y.; Mobasheri, A.; Önnerfjord, P.; Bay-Jensen, A. The minor collagens in articular cartilage. Protein Cell 2017, 8, 560–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becerra, J.; Andrades, J.A.; Guerado, E.; Zamora-Navas, P.; López-Puertas, J.M.; Reddi, A.H. Articular cartilage: Structure and regeneration. Tissue Eng. Part B Rev. 2010, 16, 617–627. [Google Scholar] [CrossRef]
- Krishnan, Y.; Grodzinsky, A.J. Cartilage diseases. Matrix Biol. 2018, 71–72, 51–69. [Google Scholar] [CrossRef]
- Hunter, D.J.; Felson, D.T. Osteoarthritis. BMJ 2006, 332, 639–642. [Google Scholar] [CrossRef]
- Triantaphyllidou, I.-E.; Kalyvioti, E.; Karavia, E.; Lilis, I.; Kypreos, K.E.; Papachristou, D.J. Perturbations in the HDL metabolic pathway predispose to the development of osteoarthritis in mice following long-term exposure to western-type diet. Osteoarthr. Cartil. 2013, 21, 322–330. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Chen, Y.; Heiman, M.; DiMarchi, R. Leptin: Structure, Function and Biology; Academic Press: Cambridge, MA, USA, 2005; Volume 71, pp. 345–372. ISBN 0083-6729. [Google Scholar] [CrossRef]
- Otero, M.; Lago, R.; Lago, F.; Casanueva, F.F.; Dieguez, C.; Gómez-Reino, J.J.; Gualillo, O. Leptin, from fat to inflammation: Old questions and new insights. FEBS Lett. 2005, 579, 295–301. [Google Scholar] [CrossRef] [Green Version]
- Masuzaki, H.; Ogawa, Y.; Hosoda, K.; Miyawaki, T.; Hanaoka, I.; Hiraoka, J.; Yasuno, A.; Nishimura, H.; Yoshimasa, Y.; Nishi, S.; et al. Glucocorticoid Regulation of Leptin Synthesis and Secretion in Humans: Elevated Plasma Leptin Levels in Cushing’s Syndrome1. J. Clin. Endocrinol. Metab. 1997, 82, 2542–2547. [Google Scholar] [CrossRef] [PubMed]
- Boden, G.; Chen, X.; Kolaczynski, J.W.; Polansky, M. Effects of prolonged hyperinsulinemia on serum leptin in normal human subjects. J. Clin. Investig. 1997, 100, 1107–1113. [Google Scholar] [CrossRef]
- Cammisotto, P.G.; Bukowiecki, L.J. Role of calcium in the secretion of leptin from white adipocytes. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 287, R1380–R1386. [Google Scholar] [CrossRef]
- Ahima, R.S.; Prabakaran, D.; Mantzoros, C.; Qu, D.; Lowell, B.; Maratos-Flier, E.; Flier, J.S. Role of leptin in the neuroendocrine response to fasting. Nature 1996, 382, 250–252. [Google Scholar] [CrossRef]
- Zhang, Y.; Proenca, R.; Maffei, M.; Barone, M.; Leopold, L.; Friedman, J.M. Positional cloning of the mouse obese gene and its human homologue. Nature 1994, 372, 425–432. [Google Scholar] [CrossRef]
- Versini, M.; Jeandel, P.-Y.; Rosenthal, E.; Shoenfeld, Y. Obesity in autoimmune diseases: Not a passive bystander. Autoimmun. Rev. 2014, 13, 981–1000. [Google Scholar] [CrossRef]
- Matarese, G.; La Rocca, C.; Moon, H.S.; Huh, J.Y.; Brinkoetter, M.T.; Chou, S.; Perna, F.; Greco, D.; Kilim, H.P.; Gao, C.; et al. Selective capacity of metreleptin administration to reconstitute CD4 + T-cell number in females with acquired hypoleptinemia. Proc. Natl. Acad. Sci. USA 2013, 110, E818–E827. [Google Scholar] [CrossRef] [Green Version]
- Procaccini, C.; Pucino, V.; Mantzoros, C.S.; Matarese, G. Leptin in autoimmune diseases. Metab. Clin. Exp. 2015, 64, 92–104. [Google Scholar] [CrossRef]
- Baumann, H.; Morella, K.K.; White, D.W.; Dembski, M.; Bailon, P.S.; Kim, H.; Lai, C.F.; Tartaglia, L.A. The full-length leptin receptor has signaling capabilities of interleukin 6-type cytokine receptors. Proc. Natl. Acad. Sci. USA 1996, 93, 8374–8378. [Google Scholar] [CrossRef] [Green Version]
- Wada, N.; Hirako, S.; Takenoya, F.; Kageyama, H.; Okabe, M.; Shioda, S. Leptin and its receptors. J. Chem. Neuroanat. 2014, 61–62, 191–199. [Google Scholar] [CrossRef]
- Zhou, Y.; Yu, X.; Chen, H.; Sjöberg, S.; Roux, J.; Zhang, L.; Ivoulsou, A.-H.; Bensaid, F.; Liu, C.-L.; Liu, J.; et al. Leptin Deficiency Shifts Mast Cells toward Anti-Inflammatory Actions and Protects Mice from Obesity and Diabetes by Polarizing M2 Macrophages. Cell Metab. 2015, 22, 1045–1058. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.-C.; Ma, B.; Guo, S.; Yang, M.; Li, L.-J.; Wang, S.-J.; Tan, J. Leptin regulates disc cartilage endplate degeneration and ossification through activation of the MAPK-ERK signalling pathway in vivo and in vitro. J. Cell. Mol. Med. 2018, 22, 2098–2109. [Google Scholar] [CrossRef] [Green Version]
- Simopoulou, T.; Malizos, K.N.; Iliopoulos, D.; Stefanou, N.; Papatheodorou, L.; Ioannou, M.; Tsezou, A. Differential expression of leptin and leptin’s receptor isoform (Ob-Rb) mRNA between advanced and minimally affected osteoarthritic cartilage; effect on cartilage metabolism. Osteoarthr. Cartil. 2007, 15, 872–883. [Google Scholar] [CrossRef] [Green Version]
- Presle, N.; Pottie, P.; Dumond, H.; Guillaume, C.; Lapicque, F.; Pallu, S.; Mainard, D.; Netter, P.; Terlain, B. Differential distribution of adipokines between serum and synovial fluid in patients with osteoarthritis. Contribution of joint tissues to their articular production. Osteoarthr. Cartil. 2006, 14, 690–695. [Google Scholar] [CrossRef] [Green Version]
- Santangelo, K.S.; Radakovich, L.B.; Fouts, J.; Foster, M.T. Pathophysiology of obesity on knee joint homeostasis: Contributions of the infrapatellar fat pad. Horm. Mol. Boil. Clin. Investig. 2016, 26, 97–108. [Google Scholar] [CrossRef]
- Conde, J.; Scotece, M.; López, V.; Abella, V.; Hermida, M.; Pino, J.; Lago, F.; Gómez-Reino, J.J.; Gualillo, O. Differential expression of adipokines in infrapatellar fat pad (IPFP) and synovium of osteoarthritis patients and healthy individuals. Ann. Rheum. Dis. 2014, 73, 631–633. [Google Scholar] [CrossRef]
- Gandhi, R.; Takahashi, M.; Virtanen, C.; Syed, K.; Davey, J.R.; Mahomed, N.N. Microarray analysis of the infrapatellar fat pad in knee osteoarthritis: Relationship with joint inflammation. J. Rheumatol. 2011, 38, 1966–1972. [Google Scholar] [CrossRef]
- Hui, W.; Litherland, G.J.; Elias, M.S.; Kitson, G.I.; Cawston, T.E.; Rowan, A.D.; Young, D.A. Leptin produced by joint white adipose tissue induces cartilage degradation via upregulation and activation of matrix metalloproteinases. Ann. Rheum. Dis. 2012, 71, 455–462. [Google Scholar] [CrossRef]
- Otero, M.; Lago, R.; Gomez, R.; Lago, F.; Dieguez, C.; Gómez-Reino, J.J.; Gualillo, O. Changes in plasma levels of fat-derived hormones adiponectin, leptin, resistin and visfatin in patients with rheumatoid arthritis. Ann. Rheum. Dis. 2006, 65, 1198–1201. [Google Scholar] [CrossRef] [Green Version]
- Dumond, H.; Presle, N.; Terlain, B.; Mainard, D.; Loeuille, D.; Netter, P.; Pottie, P. Evidence for a key role of leptin in osteoarthritis. Arthritis Rheum. 2003, 48, 3118–3129. [Google Scholar] [CrossRef]
- Yusuf, E.; Nelissen, R.G.; Ioan-Facsinay, A.; Stojanovic-Susulic, V.; DeGroot, J.; van Osch, G.; Middeldorp, S.; Huizinga, T.W.J.; Kloppenburg, M. Association between weight or body mass index and hand osteoarthritis: A systematic review. Ann. Rheum. Dis. 2010, 69, 761–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Rong, J.; Wang, Y.; Hu, F.; Bao, C.; Li, X.; Zhao, Y. The relationship between body mass index and hip osteoarthritis: A systematic review and meta-analysis. Jt. Bone Spine 2011, 78, 150–155. [Google Scholar] [CrossRef]
- Scotece, M.; Conde, J.; Gómez, R.; López, V.; Lago, F.; Gómez-Reino, J.J.; Gualillo, O. Beyond fat mass: Exploring the role of adipokines in rheumatic diseases. Sci. World J. 2011, 11, 1932–1947. [Google Scholar] [CrossRef] [Green Version]
- Figenschau, Y.; Knutsen, G.; Shahazeydi, S.; Johansen, O.; Sveinbjörnsson, B. Human articular chondrocytes express functional leptin receptors. Biochem. Biophys. Res. Commun. 2001, 287, 190–197. [Google Scholar] [CrossRef]
- Lee, S.W.; Rho, J.H.; Lee, S.Y.; Kim, J.H.; Cheong, J.-H.; Kim, H.Y.; Jeong, N.Y.; Chung, W.T.; Yoo, Y.H. Leptin protects rat articular chondrocytes from cytotoxicity induced by TNF-α in the presence of cyclohexamide. Osteoarthr. Cartil. 2015, 23, 2269–2278. [Google Scholar] [CrossRef] [Green Version]
- Kishida, Y.; Hirao, M.; Tamai, N.; Nampei, A.; Fujimoto, T.; Nakase, T.; Shimizu, N.; Yoshikawa, H.; Myoui, A. Leptin regulates chondrocyte differentiation and matrix maturation during endochondral ossification. Bone 2005, 37, 607–621. [Google Scholar] [CrossRef]
- Zhao, X.; Dong, Y.; Zhang, J.; Li, D.; Hu, G.; Yao, J.; Li, Y.; Huang, P.; Zhang, M.; Zhang, J.; et al. Leptin changes differentiation fate and induces senescence in chondrogenic progenitor cells. Cell Death Dis. 2016, 7, e2188. [Google Scholar] [CrossRef]
- Zhao, X.; Huang, P.; Li, G.; Lv, Z.; Hu, G.; Xu, Q. Activation of the leptin pathway by high expression of the long form of the leptin receptor (Ob-Rb) accelerates chondrocyte senescence in osteoarthritis. Bone Jt. Res. 2019, 8, 425–436. [Google Scholar] [CrossRef]
- Huang, Z.M.; Du, S.H.; Huang, L.G.; Li, J.H.; Xiao, L.; Tong, P. Leptin promotes apoptosis and inhibits autophagy of chondrocytes through upregulating lysyl oxidase-like 3 during osteoarthritis pathogenesis. Osteoarthr. Cartil. 2016, 24, 1246–1253. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.M.; Shen, C.; Li, H.; Fan, Q.; Ding, J.; Jin, F.C.; Sha, L. Leptin induces the apoptosis of chondrocytes in an in vitro model of osteoarthritis via the JAK2 STAT3 signaling pathway. Mol. Med. Rep. 2016, 13, 3684–3690. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xu, Z.; Wang, J.; Xu, S. DUSP19, a downstream effector of leptin, inhibits chondrocyte apoptosis via dephosphorylating JNK during osteoarthritis pathogenesis. Mol. Biosyst. 2016, 12, 721–728. [Google Scholar] [CrossRef]
- Koskinen-Kolasa, A.; Vuolteenaho, K.; Korhonen, R.; Moilanen, T.; Moilanen, E. Catabolic and proinflammatory effects of leptin in chondrocytes are regulated by suppressor of cytokine signaling-3. Arthritis Res. Ther. 2016, 18, 215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, J.; Xu, L.; Ran, J.; Xiong, Y.; Wu, L. Vaspin prevents leptin induced inflammation and catabolism by inhibiting the activation of nuclear factor κB in rat chondrocytes. Mol. Med. Rep. 2017, 16, 2925–2930. [Google Scholar] [CrossRef]
- Su, Y.-P.; Chen, C.-N.; Huang, K.-C.; Chang, H.-I.; Lee, K.-C.; Lo, C.-M.; Chang, S.-F. Leptin induces MMP1/13 and ADAMTS 4 expressions through bone morphogenetic protein-2 autocrine effect in human chondrocytes. J. Cell. Biochem. 2018, 119, 3716–3724. [Google Scholar] [CrossRef]
- Otero, M.; Lago, R.; Lago, F.; Reino, J.J.G.; Gualillo, O. Signalling pathway involved in nitric oxide synthase type II activation in chondrocytes: Synergistic effect of leptin with interleukin-1. Arthritis Res. Ther. 2005, 7, R581–R591. [Google Scholar] [CrossRef] [Green Version]
- Leonidou, A.; Lepetsos, P.; Mintzas, M.; Kenanidis, E.; Macheras, G.; Tzetis, M.; Potoupnis, M.; Tsiridis, E. Inducible nitric oxide synthase as a target for osteoarthritis treatment. Expert Opin. Ther. Targets 2018, 22, 299–318. [Google Scholar] [CrossRef]
- Pichika, R.; Homandberg, G.A. Fibronectin fragments elevate nitric oxide (NO) and inducible NO synthetase (iNOS) levels in bovine cartilage and iNOS inhibitors block fibronectin fragment mediated damage and promote repair. Inflamm. Res. 2004, 53, 405–412. [Google Scholar] [CrossRef]
- Cao, M.; Westerhausen-Larson, A.; Niyibizi, C.; Kavalkovich, K.; Georgescu, H.I.; Rizzo, C.F.; Hebda, P.A.; Stefanovic-Racic, M.; Evans, C.H. Nitric oxide inhibits the synthesis of type-II collagen without altering Col2A1 mRNA abundance: Prolyl hydroxylase as a possible target. Biochem. J. 1997, 324 Pt 1, 305–310. [Google Scholar] [CrossRef] [Green Version]
- Nishida, K.; Doi, T.; Matsuo, M.; Ishiwari, Y.; Tsujigiwa, H.; Yoshida, A.; Shibahara, M.; Inoue, H. Involvement of nitric oxide in chondrocyte cell death in chondro-osteophyte formation. Osteoarthr. Cartil. 2001, 9, 232–237. [Google Scholar] [CrossRef] [Green Version]
- Vuolteenaho, K.; Koskinen, A.; Kukkonen, M.; Nieminen, R.; Päivärinta, U.; Moilanen, T.; Moilanen, E. Leptin enhances synthesis of proinflammatory mediators in human osteoarthritic cartilage--mediator role of NO in leptin-induced PGE2, IL-6, and IL-8 production. Mediat. Inflamm. 2009, 2009, 345838. [Google Scholar] [CrossRef] [Green Version]
- Abella, V.; Scotece, M.; Conde, J.; Pino, J.; Gonzalez-Gay, M.A.; Gómez-Reino, J.J.; Mera, A.; Lago, F.; Gómez, R.; Gualillo, O. Leptin in the interplay of inflammation, metabolism and immune system disorders. Nat. Rev. Rheumatol. 2017, 13, 100–109. [Google Scholar] [CrossRef]
- Howard, J.K.; Lord, G.M.; Matarese, G.; Vendetti, S.; Ghatei, M.A.; Ritter, M.A.; Lechler, R.I.; Bloom, S.R. Leptin protects mice from starvation-induced lymphoid atrophy and increases thymic cellularity in ob/ob mice. J. Clin. Investig. 1999, 104, 1051–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, B.D.; Solar, G.P.; Yuan, J.Q.; Mathias, J.; Thomas, G.R.; Matthews, W. A role for leptin and its cognate receptor in hematopoiesis. Curr. Biol. 1996, 6, 1170–1180. [Google Scholar] [CrossRef] [Green Version]
- Santos-Alvarez, J.; Goberna, R.; Sánchez-Margalet, V. Human Leptin Stimulates Proliferation and Activation of Human Circulating Monocytes. Cell. Immunol. 1999, 194, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Maya-Monteiro, C.M.; Almeida, P.E.; D’Avila, H.; Martins, A.S.; Rezende, A.P.; Castro-Faria-Neto, H.; Bozza, P.T. Leptin induces macrophage lipid body formation by a phosphatidylinositol 3-kinase- and mammalian target of rapamycin-dependent mechanism. J. Biol. Chem. 2008, 283, 2203–2210. [Google Scholar] [CrossRef] [Green Version]
- Gruen, M.L.; Hao, M.; Piston, D.W.; Hasty, A.H. Leptin requires canonical migratory signaling pathways for induction of monocyte and macrophage chemotaxis. Am. J. Physiol. Cell Physiol. 2007, 293, C1481–C1488. [Google Scholar] [CrossRef]
- Gutierrez, D.A.; Hasty, A.H. Haematopoietic leptin receptor deficiency does not affect macrophage accumulation in adipose tissue or systemic insulin sensitivity. J. Endocrinol. 2012, 212, 343–351. [Google Scholar] [CrossRef]
- Conus, S.; Bruno, A.; Simon, H.-U. Leptin is an eosinophil survival factor. J. Allergy Clin. Immunol. 2005, 116, 1228–1234. [Google Scholar] [CrossRef]
- Amorim, N.R.T.; Souza-Almeida, G.; Luna-Gomes, T.; Bozza, P.T.; Canetti, C.; Diaz, B.L.; Maya-Monteiro, C.M.; Bandeira-Melo, C. Leptin Elicits In Vivo Eosinophil Migration and Activation: Key Role of Mast Cell-Derived PGD(2). Front. Endocrinol. 2020, 11, 572113. [Google Scholar] [CrossRef]
- Bjørbaek, C.; Uotani, S.; da Silva, B.; Flier, J.S. Divergent signaling capacities of the long and short isoforms of the leptin receptor. J. Biol. Chem. 1997, 272, 32686–32695. [Google Scholar] [CrossRef] [Green Version]
- Caldefie-Chezet, F.; Poulin, A.; Tridon, A.; Sion, B.; Vasson, M.-P. Leptin: A potential regulator of polymorphonuclear neutrophil bactericidal action? J. Leukoc. Biol. 2001, 69, 414–418. [Google Scholar] [CrossRef]
- Zarkesh-Esfahani, H.; Pockley, A.G.; Wu, Z.; Hellewell, P.G.; Weetman, A.P.; Ross, R.J.M. Leptin Indirectly Activates Human Neutrophils via Induction of TNF-α. J. Immunol. 2004, 172, 1809–1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamp, V.M.; Langereis, J.D.; van Aalst, C.W.; van der Linden, J.A.; Ulfman, L.H.; Koenderman, L. Physiological concentrations of leptin do not affect human neutrophils. PLoS ONE 2013, 8, e73170. [Google Scholar] [CrossRef] [Green Version]
- Wrann, C.D.; Laue, T.; Hübner, L.; Kuhlmann, S.; Jacobs, R.; Goudeva, L.; Nave, H. Short-term and long-term leptin exposure differentially affect human natural killer cell immune functions. Am. J. Physiol. Metab. 2011, 302, E108–E116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamas, B.; Goncalves-Mendes, N.; Nachat-Kappes, R.; Rossary, A.; Caldefie-Chezet, F.; Vasson, M.-P.; Farges, M.-C. Leptin modulates dose-dependently the metabolic and cytolytic activities of NK-92 cells. J. Cell. Physiol. 2013, 228, 1202–1209. [Google Scholar] [CrossRef] [PubMed]
- Laue, T.; Wrann, C.D.; Hoffmann-Castendiek, B.; Pietsch, D.; Hübner, L.; Kielstein, H. Altered NK cell function in obese healthy humans. BMC Obes. 2015, 2, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Sun, R.; You, L.; Gao, C.; Tian, Z. Expression of leptin receptors and response to leptin stimulation of human natural killer cell lines. Biochem. Biophys. Res. Commun. 2003, 300, 247–252. [Google Scholar] [CrossRef]
- Mattioli, B.; Straface, E.; Quaranta, M.G.; Giordani, L.; Viora, M. Leptin Promotes Differentiation and Survival of Human Dendritic Cells and Licenses Them for Th1 Priming. J. Immunol. 2005, 174, 6820–6828. [Google Scholar] [CrossRef]
- Lord, G.M.; Matarese, G.; Howard, J.K.; Baker, R.J.; Bloom, S.R.; Lechler, R.I. Leptin modulates the T-cell immune response and reverses starvation-induced immunosuppression. Nature 1998, 394, 897–901. [Google Scholar] [CrossRef]
- Saucillo, D.C.; Gerriets, V.A.; Sheng, J.; Rathmell, J.C.; Maciver, N.J. Leptin metabolically licenses T cells for activation to link nutrition and immunity. J. Immunol. 2014, 192, 136–144. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y.; Lim, J.H.; Choi, S.W.; Kim, M.; Kim, S.-T.; Kim, M.-S.; Cho, Y.S.; Chun, E.; Lee, K.-Y. Preferential effects of leptin on CD4 T cells in central and peripheral immune system are critically linked to the expression of leptin receptor. Biochem. Biophys. Res. Commun. 2010, 394, 562–568. [Google Scholar] [CrossRef] [PubMed]
- Martín-Romero, C.; Santos-Alvarez, J.; Goberna, R.; Sánchez-Margalet, V. Human Leptin Enhances Activation and Proliferation of Human Circulating T Lymphocytes. Cell. Immunol. 2000, 199, 15–24. [Google Scholar] [CrossRef]
- De Rosa, V.; Procaccini, C.; Calì, G.; Pirozzi, G.; Fontana, S.; Zappacosta, S.; La Cava, A.; Matarese, G. A key role of leptin in the control of regulatory T cell proliferation. Immunity 2007, 26, 241–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lourenço, E.V.; Liu, A.; Matarese, G.; La Cava, A. Leptin promotes systemic lupus erythematosus by increasing autoantibody production and inhibiting immune regulation. Proc. Natl. Acad. Sci. USA 2016, 113, 10637–10642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frasca, D.; Diaz, A.; Romero, M.; Blomberg, B.B. Leptin induces immunosenescence in human B cells. Cell. Immunol. 2020, 348, 103994. [Google Scholar] [CrossRef] [PubMed]
- Lam, Q.L.K.; Wang, S.; Ko, O.K.H.; Kincade, P.W.; Lu, L. Leptin signaling maintains B-cell homeostasis via induction of Bcl-2 and Cyclin D1. Proc. Natl. Acad. Sci. USA 2010, 107, 13812–13817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agrawal, S.; Gollapudi, S.; Su, H.; Gupta, S. Leptin Activates Human B Cells to Secrete TNF-α, IL-6, and IL-10 via JAK2/STAT3 and p38MAPK/ERK1/2 Signaling Pathway. J. Clin. Immunol. 2011, 31, 472–478. [Google Scholar] [CrossRef] [Green Version]
- Hunter, D.J.; March, L.; Chew, M. Osteoarthritis in 2020 and beyond: A Lancet Commission. Lancet 2020, 396, 1711–1712. [Google Scholar] [CrossRef]
- Hunter, D.J.; Bierma-Zeinstra, S. Osteoarthritis. Lancet 2019, 393, 1745–1759. [Google Scholar] [CrossRef]
- Calvet, J.; Orellana, C.; Gratacós, J.; Berenguer-Llergo, A.; Caixàs, A.; Chillarón, J.J.; Pedro-Botet, J.; García-Manrique, M.; Navarro, N.; Larrosa, M. Synovial fluid adipokines are associated with clinical severity in knee osteoarthritis: A cross-sectional study in female patients with joint effusion. Arthritis Res. Ther. 2016, 18, 207. [Google Scholar] [CrossRef] [Green Version]
- Kroon, F.P.B.; Veenbrink, A.I.; de Mutsert, R.; Visser, A.W.; van Dijk, K.W.; le Cessie, S.; Rosendaal, F.R.; Kloppenburg, M. The role of leptin and adiponectin as mediators in the relationship between adiposity and hand and knee osteoarthritis. Osteoarthr. Cartil. 2019, 27, 1761–1767. [Google Scholar] [CrossRef]
- Stannus, O.P.; Cao, Y.; Antony, B.; Blizzard, L.; Cicuttini, F.; Jones, G.; Ding, C. Cross-sectional and longitudinal associations between circulating leptin and knee cartilage thickness in older adults. Ann. Rheum. Dis. 2015, 74, 82–88. [Google Scholar] [CrossRef] [Green Version]
- Massengale, M.; Reichmann, W.M.; Losina, E.; Solomon, D.H.; Katz, J.N. The relationship between hand osteoarthritis and serum leptin concentration in participants of the Third National Health and Nutrition Examination Survey. Arthritis Res. Ther. 2012, 14, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yusuf, E.; Ioan-Facsinay, A.; Bijsterbosch, J.; Klein-Wieringa, I.; Kwekkeboom, J.; Slagboom, P.E.; Huizinga, T.W.J.; Kloppenburg, M. Association between leptin, adiponectin and resistin and long-term progression of hand osteoarthritis. Ann. Rheum. Dis. 2011, 70, 1282–1284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felson, D.T. Weight and osteoarthritis. Am. J. Clin. Nutr. 1996, 63, 430S–432S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyes, C.; Leyland, K.M.; Peat, G.; Cooper, C.; Arden, N.K.; Prieto-Alhambra, D. Association Between Overweight and Obesity and Risk of Clinically Diagnosed Knee, Hip, and Hand Osteoarthritis: A Population-Based Cohort Study. Arthritis Rheumatol. 2016, 68, 1869–1875. [Google Scholar] [CrossRef] [Green Version]
- Min, S.; Shi, T.; Han, X.; Chen, D.; Xu, Z.; Shi, D.; Teng, H.; Jiang, Q. Serum levels of leptin, osteopontin, and sclerostin in patients with and without knee osteoarthritis. Clin. Rheumatol. 2021, 40, 287–294. [Google Scholar] [CrossRef]
- Xiong, H.; Li, W.; Ke, J.; Fang, W.; Li, B.; Wei, L. Leptin Levels in the Synovial Fluid of Patients with Temporomandibular Disorders. J. Oral Maxillofac. Surg. 2019, 77, 493–498. [Google Scholar] [CrossRef]
- Lübbeke, A.; Finckh, A.; Puskas, G.J.; Suva, D.; Lädermann, A.; Bas, S.; Fritschy, D.; Gabay, C.; Hoffmeyer, P. Do synovial leptin levels correlate with pain in end stage arthritis? Int. Orthop. 2013, 37, 2071–2079. [Google Scholar] [CrossRef] [Green Version]
- Collins, K.H.; Lenz, K.L.; Pollitt, E.N.; Ferguson, D.; Hutson, I.; Springer, L.E.; Oestreich, A.K.; Tang, R.; Choi, Y.R.; Meyer, G.A.; et al. Adipose tissue is a critical regulator of osteoarthritis. Proc. Natl. Acad. Sci. USA 2020, 118. [Google Scholar] [CrossRef]
- Griffin, T.M.; Huebner, J.L.; Kraus, V.B.; Guilak, F. Extreme obesity due to impaired leptin signaling in mice does not cause knee osteoarthritis. Arthritis Rheum. 2009, 60, 2935–2944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conde, J.; Scotece, M.; López, V.; Gómez, R.; Lago, F.; Pino, J.; Gómez-Reino, J.J.; Gualillo, O. Adiponectin and Leptin Induce VCAM-1 Expression in Human and Murine Chondrocytes. PLoS ONE 2012, 7, e52533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conde, J.; Otero, M.; Scotece, M.; Abella, V.; Gómez, R.; López, V.; Pino, J.; Mera, A.; Goldring, M.B.; Gualillo, O. E74-Like Factor (ELF3) and Leptin, a Novel Loop between Obesity and Inflammation Perpetuating a Pro-Catabolic State in Cartilage. Cell. Physiol. Biochem. 2018, 45, 2401–2410. [Google Scholar] [CrossRef] [PubMed]
- Yaykasli, K.O.; Hatipoglu, O.F.; Yaykasli, E.; Yildirim, K.; Kaya, E.; Ozsahin, M.; Uslu, M.; Gunduz, E. Leptin induces ADAMTS-4, ADAMTS-5, and ADAMTS-9 genes expression by mitogen-activated protein kinases and NF-ĸB signaling pathways in human chondrocytes. Cell Biol. Int. 2015, 39, 104–112. [Google Scholar] [CrossRef]
- Otón, T.; Carmona, L. The epidemiology of established rheumatoid arthritis. Best Pract. Res. Clin. Rheumatol. 2019, 33, 101477. [Google Scholar] [CrossRef]
- Smolen, J.S.; Aletaha, D.; McInnes, I.B. Rheumatoid arthritis. Lancet 2016, 388, 2023–2038. [Google Scholar] [CrossRef]
- Rho, Y.H.; Solus, J.; Sokka, T.; Oeser, A.; Chung, C.P.; Gebretsadik, T.; Shintani, A.; Pincus, T.; Stein, C.M. Adipocytokines are associated with radiographic joint damage in rheumatoid arthritis. Arthritis Rheum. 2009, 60, 1906–1914. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Wei, J.; Li, H.; Ouyang, X.; Sun, X.; Tang, Y.; Chen, H.; Wang, B.; Li, X. Leptin Upregulates Peripheral CD4+CXCR5+ICOS+ T Cells via Increased IL-6 in Rheumatoid Arthritis Patients. J Interferon Cytokine Res. 2018, 38, 86–92. [Google Scholar] [CrossRef]
- Olama, S.M.; Senna, M.K.; Elarman, M. Synovial/Serum leptin ratio in rheumatoid arthritis: The association with activity and erosion. Rheumatol. Int. 2012, 32, 683–690. [Google Scholar] [CrossRef]
- de Resende Guimarães, M.F.B.; de Andrade, M.V.M.; Machado, C.J.; Vieira, É.L.M.; da Costa Pinto, M.R.; Júnior, A.L.T.; Kakehasi, A.M. Leptin as an obesity marker in rheumatoid arthritis. Rheumatol. Int. 2018, 38, 1671–1677. [Google Scholar] [CrossRef]
- Busso, N.; So, A.; Chobaz-Péclat, V.; Morard, C.; Martinez-Soria, E.; Talabot-Ayer, D.; Gabay, C. Leptin Signaling Deficiency Impairs Humoral and Cellular Immune Responses and Attenuates Experimental Arthritis. J. Immunol. 2002, 168, 875–882. [Google Scholar] [CrossRef]
- Bottini, N.; Firestein, G.S. Duality of fibroblast-like synoviocytes in RA: Passive responders and imprinted aggressors. Nat. Rev. Rheumatol. 2013, 9, 24–33. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Wei, J.; Tang, Y.; Wang, B.; Zhang, Y.; Shi, L.; Guo, J.; Hu, F.; Li, X. Leptin-induced migration and angiogenesis in rheumatoid arthritis is mediated by reactive oxygen species. FEBS Open Bio 2017, 7, 1899–1908. [Google Scholar] [CrossRef] [Green Version]
- Kontny, E.; Janicka, I.; Skalska, U.; Maśliński, W. The effect of multimeric adiponectin isoforms and leptin on the function of rheumatoid fibroblast-like synoviocytes. Scand. J. Rheumatol. 2015, 44, 363–368. [Google Scholar] [CrossRef]
- Yu, Y.; Fu, S.; Zhang, X.; Wang, L.; Zhao, L.; Wan, W.; Xue, Y.; Lv, L. Leptin facilitates the differentiation of Th17 cells from MRL/Mp-Fas lpr lupus mice by activating NLRP3 inflammasome. Innate Immun. 2020, 26, 294–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazzazi, H.; Aghaei, M.; Memarian, A.; Asgarian-Omran, H.; Behnampour, N.; Yazdani, Y. Th1-Th17 Ratio as a New Insight in Rheumatoid Arthritis Disease. Iran. J. Allergy. Asthma. Immunol. 2018, 17, 68–77. [Google Scholar]
- Pucino, V.; De Rosa, V.; Procaccini, C.; Matarese, G. Regulatory T cells, leptin and angiogenesis. Chem. Immunol. Allergy 2014, 99, 155–169. [Google Scholar] [CrossRef] [PubMed]
- Manson, J.J.; Rahman, A. Systemic lupus erythematosus. Orphanet J. Rare Dis. 2006, 1, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afifi, A.E.-M.A.; Shaat, R.M.; Gharbia, O.M.; Elhanafy, M.; Hasan, A.S.G. Role of serum leptin levels and leptin receptor gene polymorphisms in systemic lupus erythematosus. Clin. Rheumatol. 2020, 39, 3465–3472. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Song, G.G. Association between circulating leptin levels and systemic lupus erythematosus: An updated meta-analysis. Lupus 2018, 27, 428–435. [Google Scholar] [CrossRef]
- Wang, X.; Qiao, Y.; Yang, L.; Song, S.; Han, Y.; Tian, Y.; Ding, M.; Jin, H.; Shao, F.; Liu, A. Leptin levels in patients with systemic lupus erythematosus inversely correlate with regulatory T cell frequency. Lupus 2017, 26, 1401–1406. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Fujii, T.; Mimori, T.; Sato, T.; Nakamura, T.; Iwao, H.; Nakajima, A.; Miki, M.; Sakai, T.; Kawanami, T.; et al. Deficient Leptin Signaling Ameliorates Systemic Lupus Erythematosus Lesions in MRL/Mp- Fas lpr Mice. J. Immunol. 2014, 192, 979–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eder, L.; Haddad, A.; Rosen, C.F.; Lee, K.-A.; Chandran, V.; Cook, R.; Gladman, D.D. The Incidence and Risk Factors for Psoriatic Arthritis in Patients With Psoriasis: A Prospective Cohort Study. Arthritis Rheumatol. 2016, 68, 915–923. [Google Scholar] [CrossRef] [PubMed]
- Caso, F.; Costa, L.; Peluso, R.; Del Puente, A.; Scarpa, R. Psoriatic Arthritis. In Mosaic of Autoimmunity: The Novel Factors of Autoimmune Diseases; Academic Press: Cambridge, MA, USA, 2019; pp. 527–540. ISBN 9780128143087. [Google Scholar] [CrossRef]
- Takahashi, H.; Tsuji, H.; Takahashi, I.; Hashimoto, Y.; Ishida-Yamamoto, A.; Iizuka, H. Plasma adiponectin and leptin levels in Japanese patients with psoriasis. Br. J. Dermatol. 2008, 159, 1207–1208. [Google Scholar] [CrossRef]
- Çerman, A.A.; Bozkurt, S.; Sav, A.; Tulunay, A.; Elbaşı, M.O.; Ergun, T. Serum leptin levels, skin leptin and leptin receptor expression in psoriasis. Br. J. Dermatol. 2008, 159, 820–826. [Google Scholar] [CrossRef]
- Xue, Y.; Jiang, L.; Cheng, Q.; Chen, H.; Yu, Y.; Lin, Y.; Yang, X.; Kong, N.; Zhu, X.; Xu, X.; et al. Adipokines in Psoriatic Arthritis Patients: The Correlations with Osteoclast Precursors and Bone Erosions. PLoS ONE 2012, 7, e46740. [Google Scholar] [CrossRef] [Green Version]
- Paz-Filho, G.; Mastronardi, C.A.; Licinio, J. Leptin treatment: Facts and expectations. Metabolism 2015, 64, 146–156. [Google Scholar] [CrossRef]
- Elinav, E.; Niv-Spector, L.; Katz, M.; Price, T.O.; Ali, M.; Yacobovitz, M.; Solomon, G.; Reicher, S.; Lynch, J.L.; Halpern, Z.; et al. Pegylated Leptin Antagonist Is a Potent Orexigenic Agent: Preparation and Mechanism of Activity. Endocrinology 2009, 150, 3083–3091. [Google Scholar] [CrossRef] [Green Version]
- Fazeli, M.; Zarkesh-Esfahani, H.; Wu, Z.; Maamra, M.; Bidlingmaier, M.; Pockley, A.G.; Watson, P.; Matarese, G.; Strasburger, C.J.; Ross, R.J.M. Identification of a monoclonal antibody against the leptin receptor that acts as an antagonist and blocks human monocyte and T cell activation. J. Immunol. Methods 2006, 312, 190–200. [Google Scholar] [CrossRef]
- Huebner, J.L.; Landerman, L.R.; Somers, T.J.; Keefe, F.J.; Guilak, F.; Blumenthal, J.A.; Caldwell, D.S.; Kraus, V.B. Exploratory secondary analyses of a cognitive-behavioral intervention for knee osteoarthritis demonstrate reduction in biomarkers of ad-ipocyte inflammation. Osteoarthr. Cartil. 2016, 24, 1528–1534. [Google Scholar] [CrossRef] [Green Version]
- Drinda, S.; Franke, S.; Schmidt, S.; Stoy, K.; Lehmann, T.; Wolf, G.; Neumann, T. AGE-RAGE Interaction Does Not Explain the Clinical Improvements after Therapeutic Fasting in Osteoarthritis. Complement. Med. Res. 2018, 25, 167–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, H.; de Toledo, F.W.; Resch, K.L. Fasting followed by vegetarian diet in patients with rheumatoid arthritis: A systematic review. Scand. J. Rheumatol. 2001, 30, 1–10. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
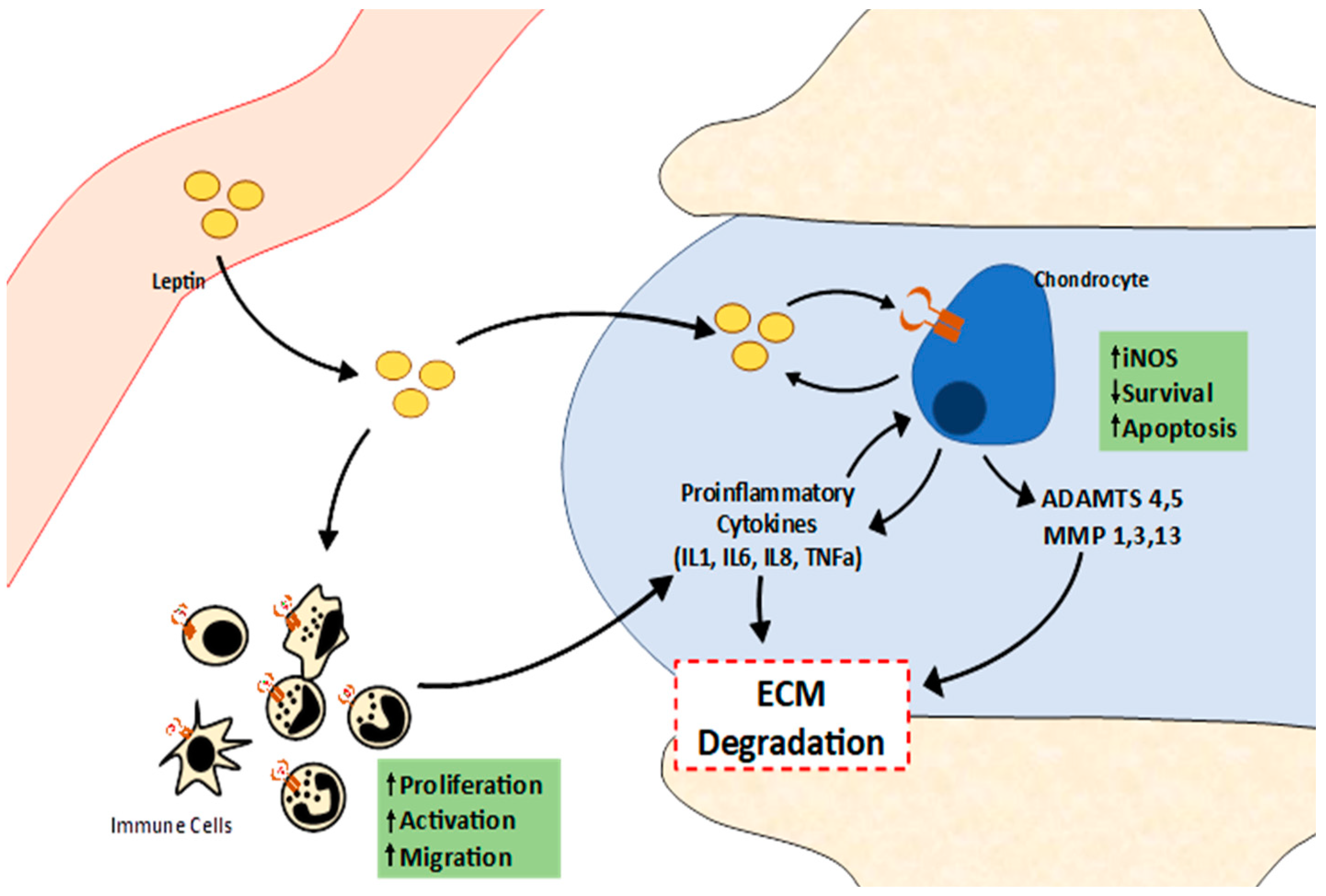{kind=link}
| Author | Year | Disease | Patients | Sample Origin | Leptin Relation with Disease |
|---|---|---|---|---|---|
| Simpoulou, T. et al. [24] | 2007 | OA | 11 | Serum and synovial fluid from Knee and Hips cartilage | Leptin and leptin receptor levels are increased in OA disease and disease progression |
| Calvet, J. et al. [81] | 2016 | OA | 115 | Synovial fluid from Knee | Leptin levels are increased in OA severity |
| Kroon, F.P.B et al [82] | 2019 | OA | 6408 | Serum | Leptin levels are increased in OA disease |
| Massengale, M. et al. [84] | 2019 | OA | 2477 | Serum | No evidence of relation |
| Yusuf, E. et al. [85] | 2011 | OA | 164 | Serum | No evidence of relation |
| Xiong, H. et al. [89] | 2019 | OA | 38 | Synovial fluid from temporomandibular articulation | Leptin levels are increased in OA disease |
| Lübbeke, A. et al. [90] | 2013 | OA | 219 | Synovial fluid from hips and Knees | Leptin levels are increased in OA and are related with join pain |
| Min, S. et al. [88] | 2021 | OA | 148 | Serum | Leptin levels are increased in OA disease |
| Rho, Y.H. et al. [90] | 2009 | RA | 167 | Serum | Leptin levels are increased in RA disease |
| Wang, M. et al. [99] | 2017 | RA | 54 | Serum | Leptin levels are increased in RA |
| Olama, S.M. et al. [100] | 2010 | RA | 40 | Serum and synovial fluid from Knee | Leptin levels are increased in RA |
| Afifi, A.E. et al. [110] | 2020 | SLE | 50 | Serum | Leptin levels are increased in SLE |
| Lee, Y.H. et al. [111] | 2018 | SLE | 1333 | Serum | Leptin levels are increased in SLE |
| Wang, X. et al. [112] | 2017 | SLE | 47 | Serum | Leptin levels are increased in SLE |
| Takahashi, H. et al. [116] | 2008 | PsA | 122 | Serum | Leptin levels are increased in PsA |
| Çerman, A.A. et al. [117] | 2008 | PsA | 43 | Serum | Leptin levels are increased in PsA and related with disease severity |
| Xue, Y. et al. [118] | 2012 | PsA | 41 | Serum | Leptin levels are increased in PsA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cordero-Barreal, A.; González-Rodríguez, M.; Ruiz-Fernández, C.; Eldjoudi, D.A.; AbdElHafez, Y.R.F.; Lago, F.; Conde, J.; Gómez, R.; González-Gay, M.A.; Mobasheri, A.; et al. An Update on the Role of Leptin in the Immuno-Metabolism of Cartilage. Int. J. Mol. Sci. 2021, 22, 2411. https://doi.org/10.3390/ijms22052411
Cordero-Barreal A, González-Rodríguez M, Ruiz-Fernández C, Eldjoudi DA, AbdElHafez YRF, Lago F, Conde J, Gómez R, González-Gay MA, Mobasheri A, et al. An Update on the Role of Leptin in the Immuno-Metabolism of Cartilage. International Journal of Molecular Sciences. 2021; 22(5):2411. https://doi.org/10.3390/ijms22052411
Chicago/Turabian StyleCordero-Barreal, Alfonso, María González-Rodríguez, Clara Ruiz-Fernández, Djedjiga Ait Eldjoudi, Yousof Ramadan Farrag AbdElHafez, Francisca Lago, Javier Conde, Rodolfo Gómez, Miguel Angel González-Gay, Ali Mobasheri, and et al. 2021. "An Update on the Role of Leptin in the Immuno-Metabolism of Cartilage" International Journal of Molecular Sciences 22, no. 5: 2411. https://doi.org/10.3390/ijms22052411